Insektenpheromone
Insektenpheromone sind Botenstoffe, die der chemischen Kommunikation zwischen Individuen einer Insekten-Art dienen. Sie unterscheiden sich damit von Kairomonen, also Botenstoffen, die Information an artfremde Organismen übertragen. Insekten produzieren Pheromone in speziellen Drüsen und geben sie an die Umgebung ab. In den Pheromonrezeptoren der Sinneszellen des Empfängers erzeugen sie bereits in sehr geringer Konzentration einen Nervenreiz, der schließlich zu einer Verhaltensantwort führt. Die innerartliche Kommunikation der Insekten über diese Stoffe erfolgt in vielfältiger Weise und dient unter anderem zum Finden von Geschlechtspartnern, der Aufrechterhaltung der Harmonie in einer Kolonie sozial lebender Insekten, der Markierung von Territorien oder dem Auffinden von Nestplätzen und Nahrungsquellen.

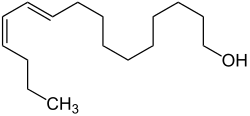
Im Jahr 1959 identifizierte und synthetisierte der deutsche Biochemiker und Nobelpreisträger Adolf Butenandt den ungesättigten Fettalkohol Bombykol, den Sexuallockstoff des Seidenspinners (Bombyx mori), als erstes bekanntes Insektenpheromon. Bei den Sexualpheromonen weiblicher Schmetterlinge handelt es sich meist um mono- oder bis-olefinische Fettsäuren beziehungsweise deren Ester, Fettalkohole, deren Ester oder die entsprechenden Aldehyde. Männliche Falter verwenden ein breites Spektrum von Chemikalien als Sexualpheromone, zum Beispiel Pyrrolizidinalkaloide, Terpene und aromatische Verbindungen wie Benzaldehyd.
Die Erforschung der chemischen Kommunikation von Insekten erweitert das Verständnis darüber, wie diese ihre Nahrungsquellen oder Plätze zur Eiablage auffinden. So nutzen Imker ein künstlich hergestelltes Nasanov-Pheromon, das Terpene wie Geraniol und Citral enthält, um Bienen zu einem ungenutzten Bienenstock zu locken. Die Land- und Forstwirtschaft verwendet Insektenpheromone kommerziell bei der Schädlingsbekämpfung mittels Lockstofffallen zur Verhinderung der Eiablage und bei der Praktizierung der Verwirrmethode. Es besteht die Erwartung, dass Insektenpheromone auf diese Weise auch zur Eindämmung von durch Insekten übertragenen Infektionskrankheiten wie Malaria, Denguefieber oder Afrikanische Trypanosomiasis beitragen können.
Etymologie und Einteilung
Adolf Butenandt und Peter Karlson schlugen im Jahr 1959 den Begriff der Pheromone für Stoffe vor, die der intraspezifischen Kommunikation dienen.[2] Die Definition des Pheromonbegriffs erfolgte im selben Jahr durch Karlson und den Schweizer Zoologen Martin Lüscher. Demnach sind Pheromone
„Substanzen, die von einem Individuum nach außen abgegeben werden und bei einem anderen Individuum der gleichen Art spezifische Reaktionen auslösen.“
Das Wort Pheromon besteht aus den altgriechischen Wortteilen φέρειν phérein, überbringen, melden und ὁρμᾶν hormān, antreiben, erregen.[3][4] Laut Karlson und Lüscher war es das Ziel, für eine Klasse von Substanzen basierend auf einer klaren Definition einen international verständlichen wissenschaftlichen Begriff zu prägen. Es sollte ein kurzes Wort sein, das in vielen Sprachen gesprochen werden kann. Die Endung mon diente als Suffix, wie es in den Wörtern Hormon, Kairomon und Allomon vorkommt und damit deren Verwandtschaft unterstrich.[5] Der Begriff Pheromon löste den Ausdruck Ektohormon beziehungsweise Homoiohormon ab, den Albrecht Bethe bereits im Jahr 1932 mit gleicher Definition vorgeschlagen hatte.[6] Die Bezeichnung von Bethe setzte sich nicht durch, weil die Bezeichnungen Ekto und Hormon sich laut Butenandt gegenseitig ausschlossen. Der Wirkmechanismus eines Pheromons entspricht auch nicht dem eines von einem anderen Individuum in den Blutkreislauf aufgenommenen Hormons und wurde daher als irreführend empfunden.
Die Einordnung der intraspezifisch wirkenden Pheromone in die Gruppe der Semiochemikalien, also der Botenstoffe, die der Kommunikation zwischen Organismen dienen, zeigt die folgende Grafik:[7]
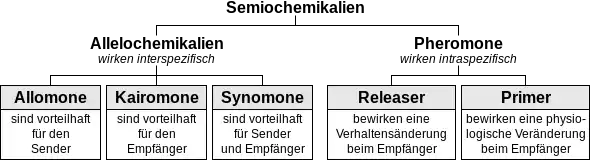
Karlson unterteilte sie nach der Art des Empfangs weiter in olfaktorisch wirkende und oral wirkende Insektenpheromone.[8] Im Jahr 1963 führten Edward O. Wilson, der im Jahr zuvor die Spurenpheromone der Ameisen entdeckt hatte, und William H. Bossert die Begriffe der Releaser- und Primerpheromone ein.[9][10] Releaserpheromone, die meist olfaktorisch wahrgenommen werden, bewirken eine augenblicklich beobachtbare Verhaltensreaktion, wohingegen Primerpheromone, die häufig oral wirken, physiologische Veränderungen beim Empfänger auslösen. Primerpheromone unterdrücken zum Beispiel die Ausbildung der Eierstöcke bei Arbeitsbienen.
Häufig werden Pheromone nach ihrer verhaltensauslösenden Funktion definiert. Neben den bekannten Sexuallockstoffen wirken sie unter anderem als Aggregationspheromone, Dispersionspheromone, Alarmpheromone, Spurpheromone, Markierungspheromone, Bruterkennungspheromone, Eiablagepheromone, Rekrutierungspheromone oder als Kastenerkennungsstoffe.[11]
Vincent Dethier teilte die Insektenpheromone nach ihrer generellen verhaltensauslösenden Wirkung in sechs Kategorien ein.[12] Dazu zählen die normalerweise nur auf kurze Entfernung wahrnehmbaren Arrestants, die ein in Bewegung befindliches Insekt zum Anhalten veranlassen, sowie die Locomotor Stimulants, welche die Geschwindigkeit der Insekten erhöhen oder die Anzahl von Richtungsänderungen verringern. Attractants sind Lockstoffe, die eine orientierte Bewegung zur Riechquelle hin auslösen, wohingegen Repellents eine Fluchtbewegung von dieser weg auslösen. Feeding- beziehungsweise Oviposition Stimulants lösen die Fütterung oder die Eiablage aus. Deterrents dagegen hemmen den Fraß oder die Eiablage.
Funktional definierte Insektenpheromone enthalten oft Mischungen verschiedener Komponenten in genau definierten Mengenverhältnissen. Diese sogenannten Pheromoncocktails enthalten häufig Stoffe verschiedener Kategorien mit Nah- und Fernorientierungsfunktion. So enthält der Aggregationspheromoncocktail der Deutschen Schabe Blattella germanica sowohl Stoffe, die als Attractant wirken als auch Stoffe, die als Arrestant wirken.[13]
Zum Teil werden Insektenpheromone nach dem Ort ihrer biologischen Produktion benannt. Männchen verschiedener Falterarten wie etwa der Bananenfalter besitzen im Hinterleib sogenannte androconiale Organe, die Pheromone abgeben. Diese Insektenpheromone werden entsprechend als Androconialpheromone bezeichnet.[14] Die Königinnen der Westlichen Honigbiene produzieren das Bienenköniginnenpheromon in Mandibeldrüsen. Im englischen Sprachraum werden sie daher oft als Queen Mandibular Gland Pheromones, Königinnenmandibeldrüsenpheromone, bezeichnet.[15]
Geschichte
Erste Entdeckungen

Der englische Imker Charles Butler beobachtete im Jahr 1609, dass durch den Stich einer Biene eine Flüssigkeit freigesetzt wurde. Diese Flüssigkeit zog andere Bienen an und diese begannen daraufhin in Massen zu stechen.[16] Butler stellte damit erstmals die Wirkung eines Alarmpheromons der Bienen dar, das in den 1960er Jahren als Isoamylacetat identifiziert wurde.[17]
Sir John Ray vermutete bereits 1690, dass Birkenspannerweibchen männliche Artgenossen über einen Duft anlockten:
“It emerged out of a stick-shaped geometer caterpillar: it was a female and came out from its chrysalis shut up in my cage: the windows were open in the room or closet where it was kept, and two male moths flying round were caught by my wife who by a lucky chance were into the room in the night: they were attracted, as it seems to me, by the scent of the female and came in from outside.”
„Es entwickelte sich aus einer stabförmigen Birkenspannerraupe: Es war ein Weibchen und kam aus ihrer Puppe, die in meinen Käfig eingeschlossen war: Die Fenster waren offen in dem Raum oder der Kammer, wo sie gehalten wurde, und zwei männliche Falter, die herumflogen, wurden von meiner Frau gefangen, die durch einen glücklichen Zufall in dieser Nacht im Raum war: Sie wurden, wie es mir scheint, durch den Duft des Weibchen angelockt und kamen von außen rein.“

Der französische Entomologe Jean-Henri Fabre berichtete Mitte des 19. Jahrhunderts ebenfalls über Versuche mit Nachtpfauenaugen und Eichenspinnern, bei denen in Drahtkäfigen gefangene Weibchen innerhalb weniger Tage zu bestimmten Uhrzeiten Hunderte von Männchen anlockten.[19] Bei Versuchen mit markierten Seidenspinnermännchen fanden noch 40 % der Männchen aus einer Entfernung von vier Kilometern und 26 % der Männchen aus elf Kilometern zu einem gefangenen Weibchen.[19]
Bei vielen Insektenarten rätselten Forscher lange Zeit über den Mechanismus des Zusammenfindens der Geschlechtspartner: Visuelle oder akustische Reize konnten weder die von Fabre durchgeführten Versuche noch, wie Nachtfalter mit großer Sicherheit paarungsbereite Weibchen fanden, erklären. Theorien über eine Lockwirkung durch Infrarot- oder andere Strahlung bestätigten sich nicht.[19] Ebenso unerklärlich blieb lange Zeit die Organisation von Insektenstaaten. Der Schriftsteller und Bienenforscher Maurice Maeterlinck spekulierte über den Spirit of the hive, den (Team)-Geist des Bienenstocks, ohne dessen Wesen näher bestimmen zu können.[20]
Definitionen von Bethe
Zu Beginn des 20. Jahrhunderts entdeckte Ernest Starling die Hormone als erste biologische Botenstoffe.[21] Im Jahr 1932 veröffentlichte der Neurophysiologe Albrecht Bethe, der zu dieser Zeit das Institut für Tierphysiologie an der Universität Frankfurt am Main leitete, einen Artikel über ein erweitertes Hormonkonzept, bei dem er zwischen Endohormonen und Ektohormonen unterschied.[6] Die Endohormone wirken demnach im produzierenden Organismus selbst und entsprechen der klassischen Hormondefinition. Im Gegensatz dazu gibt der Organismus Ektohormone nach außen ab und überträgt sie auf andere Individuen. Als Beispiel führte Bethe die Wirkung des Laktationshormons an, das von einem Fötus an die Mutter abgegeben wird und bei dieser das Wachstum der Brustdrüse und anschließend die Milchsekretion hervorruft.[6] Dieses Konzept schlug er auch für die chemische Kommunikation unter Insekten vor.
„Bei den Bienen z. B. vermögen die Arbeiterinnen (also nicht die Mütter) aus einem Ei oder einer jungen Larve […] durch besondere Nahrung und Übertragung von Sekreten ihrer Speicheldrüsen eine geschlechtsfähige Königin heranzuziehen. Es kann wohl kaum ein Zweifel darüber bestehen (obwohl es nicht bewiesen ist), daß bei dieser Umdifferenzierung Ektohormone des Speicheldrüsensekrets die Hauptrolle spielen.“
Bethe unterteilte die Ektohormone weiter in Homoiohormone, die – entsprechend der heutigen Definition eines Pheromons – auf Individuen der gleichen Art wirken, und Alloiohormone, die auf Individuen einer anderen Art wirken. Damit prägte er den Vorläuferbegriff der Allelochemikalien.[6]
Arbeiten von Butenandt
Auch Adolf Butenandt vermutete, dass die Kommunikation unter Insekten auf Botenstoffen basierte, und begann in den 1940er Jahren ein Projekt zur Identifizierung des Sexuallockstoffs des Seidenspinners (Bombyx mori). Es handelt sich dabei um einen ursprünglich in China beheimateten Schmetterling aus der Familie der Echten Spinner, der dem Seidenbau dient und dessen Aufzucht und Haltung gut bekannt war. Erst nach fast 20-jähriger Arbeit gelang die endgültige Extraktion und Reinigung eines Stoffes aus mehr als 500.000 Insekten, den Butenandt später Bombykol nannte.
Durch Elementaranalyse bestimmte Butenandt die Summenformel des Stoffes zu C16H30O. Infrarotspektroskopische Untersuchungen wiesen auf die Anwesenheit von konjugierten Doppelbindungen hin. Mit damals gängigen Methoden wie der katalytischen Hydrierung, der Schmelzpunktbestimmung und dem oxidativen Abbau durch Kaliumpermanganat zeigte Butenandt, dass es sich bei dem gesuchten Stoff um einen ungesättigten Fettalkohol, das (10E,12Z)-10,12-Hexadecadien-1-ol, handelte.[22]
Butenandt synthetisierte anschließend Bombykol aus Vernolsäure [(12R,13S)-Epoxy-9-cis-octadecensäure] in mehreren Schritten über die Diolbildung, dessen Spaltung in den Aldehyd, Doppelbindungsisomerisierung und Wittig-Olefinierung. Er synthetisierte die vier möglichen Stereoisomere und testete sie auf ihre biologische Aktivität.[23] Nur ein Isomer zeigte dieselbe Aktivität wie das Extrakt. Damit erbrachte Butenandt den Nachweis, dass die Kommunikation unter Insekten auf stofflicher Basis erfolgt.
„Durch Extraktions- und Kondensationsversuche ist jedoch überzeugend gezeigt worden, daß ein stoffliches Prinzip vorliegen muss, das von den weiblichen Schmetterlingen aus Duftorganen der letzten Hinterleibssegmente sezerniert und von den Männchen mit ihren Antennen wahrgenommen wird.“
Primer- und Releaserpheromone

Gegen Ende der 1950er Jahre definierte Edward O. Wilson Stoffe, die das Alarm- und Grabverhalten von Ameisen auslösen, als Chemical Releaser.[25] Der britische Biochemiker Robert Kenneth Callow identifizierte im Jahr 1961 mit der Verbindung (E)-9-Oxo-dec-2-ensäure, kurz 9-ODA, ein weiteres Pheromon, auch bekannt als Bienenköniginnenpheromon.[26] Die Wirkung dieses Pheromons war offensichtlich anders geartet als die der Alarmpheromone, da es sich langfristig auf die Physiologie der Empfänger auswirkte.
Im Jahr 1963 führten Wilson, der im Jahr zuvor bereits die Spurenpheromone der Ameisen entdeckte, und William H. Bossert dafür den Begriff der Releaser- und Primerpheromone ein, um die verhaltenssteuernde Wirkung von zum Beispiel Sexuallockstoffen von den Pheromonen, die in das Hormonsystem des Empfängers eingreifen, zu unterscheiden.[27][28]
Moderne Forschungsrichtungen
Durch die im Laufe der Jahre enorm verfeinerten Extraktions- und Analyseverfahren identifizierten Chemiker und Biologen zahlreiche weitere Pheromone. Zum Nachweis der zweiten Komponente des Pheromoncocktails von Bombyx mori, des Bombykals [(10Z,12E)-Hexadecadienal], genügte im Jahr 1978 bereits ein Extrakt von 460 Drüsen, aus denen 15 Nanogramm des Aldehyds isoliert wurden.[29]

Neben der Erforschung der Funktion und des Empfangs von Pheromonen und der chemischen Identifizierung untersuchten Wissenschaftler eingehend die Biochemie der Pheromonproduktion. Im Jahr 1984 entdeckten Ashok Raina und Jerome Klun, dass die Produktion des weiblichen Sexuallockstoffs des Eulenfalters Helicoverpa zea durch hormonelle Substanzen, die sogenannten Pheromon-Biosynthese-aktivierenden Neuropeptide (PBAN) im Gehirn weiblicher Falter gesteuert wird.[30] Andere moderne Forschungsschwerpunkte sind die Untersuchung des Empfangs von Insektenpheromonen mittels des Geruchs- und Geschmacksinns, genetische Faktoren und evolutionsbiologische Fragestellungen, wie die Koevolution der weiblichen Sexualpheromonproduktion und der Empfang beim Männchen.[31][32]
Die Bekämpfung von Krankheitsüberträgern wie der Malariamücken bildet einen weiteren Schwerpunkt der Forschung. Nach Schätzungen der Weltgesundheitsorganisation betrug die Zahl der Malariainfektionen im Jahr 2012 etwa 207 Millionen mit 627.000 Todesfällen.[33] Culexmücken übertragen den Erreger der Filariose oder das West-Nil-Virus. Eine Möglichkeit zur Eindämmung dieser Populationen bieten mit Eiablagepheromonen ausgestattete Fallen. Um diese zu optimieren, werden die duftstoffbindenden Proteine in den Antennen der Weibchen, die eine entscheidende Rolle bei der Erkennung der Eiablageplätze spielen, intensiv untersucht.[34]
Herstellung
Als Pheromone bei Insekten dienen oft die Folgeprodukte von Fettsäuren, wie gesättigte und ungesättigte Kohlenwasserstoffe, Fettalkohole, Ester und Aldehyde, aber auch Isoprenoide und andere Verbindungen. Pheromone sind oft nicht reine Stoffe, sondern sogenannte Pheromoncocktails, die aus verschiedenen Komponenten bestehen. Oft löst nur ein spezielles Enantiomer einer Verbindung eine Verhaltensreaktion aus, während das andere Enantiomer keine oder eine andere Reaktionen auslöst.
Manchmal erfolgt die Biosynthese des Pheromons nur, wenn die biochemischen Vorstufen in Form bestimmter Alkaloide aus Nahrungspflanzen aufgenommen wurden. Der Sexuallockstoff signalisiert in diesem Fall gleichzeitig das Vorkommen von Nahrungsquellen.[35]
Durch die potentielle kommerzielle Anwendung im Pflanzenschutz nahm die Intensität der Untersuchung von Pheromonen nach Butenandts Entdeckung stark zu und führte zur Entwicklung hochempfindlicher Analysemethoden[36] und der breiten Anwendung chemo-, regio- und stereoselektiver Synthesen in der organischen Chemie.
Biosynthese

Insektenpheromone werden von einer Vielzahl von exokrinen Drüsen, die vorwiegend aus modifizierten Epidermalzellen an verschiedenen Stellen des Insektenkörpers bestehen, hergestellt. So geben die Hinterleibsdrüsen des Seidenspinnerweibchens neben dem Sexuallockstoff Bombykol Spuren des (E,E)-Isomers des Alkohols als auch den analogen (E,Z)-Aldehyd Bombykal ab.[37][38] Geeignete Oberflächengeometrien in der Umgebung der Drüsen, etwa geriefte Porenplatten, können die effektive Verdampfung eines ausgetretenen Pheromons begünstigen.[39] Honigbienen besitzen 15 Drüsen, mit denen sie eine Reihe verschiedener Substanzen herstellen und abgeben und damit ein komplexes, auf Pheromonen basierendes Kommunikationssystem unterhalten.[40][41] Männchen verschiedener Schmetterlingsarten besitzen im Hinterleib sogenannte androconiale Organe, mit denen sie Pheromone verbreiten können, andere Falter geben diese über Duftschuppen oder Duftborsten an ihren Vorderflügeln oder dem Hinterleibsende ab. Die Duftborsten und Duftschuppen dienen der Oberflächenvergrößerung und erleichtern das Abdampfen der Insektenpheromone.[39]
Anstatt eine völlig einmalige Reihe von Enzymen für die Pheromon-Biosynthese zu entwickeln, modifizieren Insekten oft normale Stoffwechselprodukte zu Pheromonen mit hoher Regio-, Chemo-, (E/Z)-, Diastereo- oder Enantioselektivität und in genau definierten Mengenverhältnissen.[42] Die Biosynthese der Insektenpheromone geschieht entweder de novo nach dem Schema der Fettsäuresynthese durch sukzessiven Anbau von Malonyl-CoA an ein initiales Acetyl oder durch Aufnahme von Precursorn aus der Nahrung. Viele Schmetterlinge nutzen die biosynthetische Möglichkeit, eine bestimmte Mischung von Derivaten einfacher Fettsäuren herzustellen. Die Entwicklung des Enzyms Δ-11-Desaturase in Kombination mit kettenverkürzenden Reaktionen erlaubt es ihnen, eine Vielzahl von ungesättigten Acetaten, Aldehyden und Alkoholen in verschiedenen Kombinationen zu produzieren.[43]
Durch spezielle Enzymsysteme erfolgt gegebenenfalls eine Dehydrierung der Kohlenstoffkette und die Reduktion der Säurefunktion zum Alkohol. Weitere Schritte können die Oxidation zum Aldehyd oder die Acetylierung zum Essigsäureester sein.[44] Bei Bombyx mori wird die Biosynthese tagesperiodisch von Pheromonen durch ein Neurohormon, das sogenannte Pheromon-Biosynthese-aktivierende Neuropeptid (PBAN), aktiviert.[44]
Die hormonellen Mechanismen der Pheromonproduktion unterscheiden sich von Art zu Art erheblich.[45] Juvenilhormone etwa kontrollieren die Pheromonproduktion des Eulenfalters Mythimna unipuncta. Diese werden in hinter dem Gehirn liegenden meist paarig vorkommenden Corpora allata gebildet und in die Hämolymphe abgegeben. Dort binden sie an bestimmte Transportproteine. Werden die Corpora allata entfernt, produzieren die Weibchen keine Pheromone. Juvenilhormone greifen aber eher indirekt in die circadiane Freisetzung von PBAN ein.[46]
Männliche Schmetterlinge aus der Familie der Danainae verwunden zum Teil mit winzigen Klauen an ihren Füßen Raupen, die Alkaloide aus Seidenpflanzen aufgenommen haben, um die austretende Flüssigkeit aufzunehmen, ein als Kleptopharmakophagie beschriebenes Verhalten. Die Falter nutzen die aufgenommenen Alkaloide für die Verteidigung gegen Fressfeinde und um Sexualpheromone herzustellen.[47]
Pheromone aus Pflanzeninhaltsstoffen
Männliche Feuerkäfer der Art Neopyrochroa flabellata und auch verschiedene andere Käferarten nutzen das Terpenoid Cantharidin als Sexualpheromon beziehungsweise Aphrodisiakapheromon. Dieses Isoprenoid wird von Neopyrochroa flabellata mit der Nahrung aufgenommen und beim Paarungsakt auf die Weibchen und anschließend auf die Brut übertragen.[48] Die Weibchen prüfen den Gehalt einer Drüse am Kopf des Männchens vor der Paarung. Das Cantharidin wirkt als Fraßgift und macht die Eier für Räuber ungenießbar; Weibchen bevorzugen daher Männchen mit einem hohen Cantharidingehalt.[48]
Falter wie Utetheisa ornatrix und Tirumala limniace nehmen im Larvenstadium Pyrrolizidinalkaloide aus Nahrungspflanzen wie Crotalaria, Sonnenwenden oder Leberbalsam-Schafgarbe auf, die das erwachsene Männchen durch Oxidation in Pheromone wie Hydroxydanaidal umwandelt. Wie beim Feuerkäfer werden die Alkaloide, die starke Fraßgifte sind und gegen Fressfeinde wie Spinnen, Ameisen oder Netzflügler wirken, auf Weibchen und Eier übertragen.[49] Erwachsene Monarchfalter nehmen sekundäre Pflanzenstoffe auf und erhöhen damit ihre pheromonale Attraktivität.[50] Manchmal erfolgt die Biosynthese des Pheromons nur, wenn die biochemischen Vorstufen in Form bestimmter Alkaloide aus Nahrungspflanzen aufgenommen wurden. Der Sexuallockstoff signalisiert in diesem Fall gleichzeitig das Vorkommen von Nahrungsquellen.[51]
Die Aufnahme von Pheromonvorstufen aus Pflanzen ist auch für bestimmte Arten von Prachtbienen und Bohrfliegen bekannt. Männliche Bienen sammeln eine Mischung von Terpenoiden aus Orchideen und nutzen sie als Aggregationspheromon zur Bildung von Balzplätzen. Manchmal steuern die Pflanzeninhaltsstoffe die Entwicklung der Pheromondrüsen von männlichen Schmetterlingen.[52]
Laborsynthese
Karl Ziegler und Günther Otto Schenck gelang bereits 1941 die Synthese von Cantharidin.[53][54] Die Darstellung der Pheromone erfordert die Anwendung hoch chemo-, regio- und stereoselektiver Synthesen. In den siebziger Jahren gelang es mittels asymmetrischer Synthese unter Verwendung der SAMP-Methode, verschiedene Pheromone enantiomerenrein herzustellen.[55] Des Weiteren setzten Chemiker asymmetrische Epoxidierungen, asymmetrische Dihydroxylierung, Biokatalyse, Olefinmetathese und viele weitere stereoselektiv verlaufende Reaktionen zur Synthese von Pheromonen ein.[56] Die Wittig-Reaktion eignet sich zur Synthese von Pheromonen mit (Z)-olefinischen Doppelbindungen.[57]
Auch gentechnisch veränderte Tabakpflanzen können Sexualpheromone produzieren. Die daraus durch Extraktion gewonnenen Fettalkohole werden anschließend acetyliert, um die jeweiligen Zielsexualpheromone zu gewinnen. Dieser halbsynthetische Weg der Herstellung produziert Insektenpheromone in relativ großer Menge und mit hoher Reinheit.[58]
Eigenschaften

Die chemische Kommunikation zwischen Lebewesen mittels Pheromonen erfolgt nach den gleichen Prinzipien wie die technische Datenübertragung. Ein Sender, zum Beispiel die Drüse eines weiblichen Insekts, gibt das Signal in Form einer chemischen Substanz ab. Sowohl die chemische Struktur der Moleküle als auch ihr Mengenverhältnis bestimmen den Informationsgehalt und dienen als gemeinsamer Zeichenvorrat der Art. Die physikalischen Eigenschaften der Stoffe wie der Dampfdruck determinieren die Funktion ihrer Moleküle als Kurz- oder Fernwegsinformationsüberträger.[59]
Das Insektenpheromon wird durch direkten Kontakt oder über ein Medium wie Wasser oder Luft übertragen. Vom Empfänger, zum Beispiel den Pheromonrezeptoren in der Antenne eines Insektenmännchens, wird der Stoff empfangen und löst eine Verhaltensreaktion aus. Der Begriff der Antenne wurde zunächst für die Fühler der Insekten und danach in der Technik verwendet.[32] Insektenpheromone wirken hochgradig artspezifisch, das heißt, dass sie die gewünschte Verhaltensreaktion nur bei Artgenossen hervorrufen, jedoch nicht bei Individuen anderer Arten. Obwohl zum Beispiel die chemischen Verbindungen, die als Sexualpheromone bei Schmetterlingen wirken, bei verschiedenen Arten gleich sein können, ist die Zusammensetzung des Pheromoncocktails bei allen Arten verschieden.[59] Daneben enthalten die Pheromoncocktails oft Substanzen, die als Verhaltensinhibitoren für andere Arten wirken und etwa die Anflugrate von Männchen fremder Arten auf ein lockendes Weibchen erheblich reduzieren.[60]
Physikalisch-chemische Eigenschaften
Die Pheromone werden meist als Flüssigkeit hergestellt und entweder durch direkten Kontakt übertragen oder als Flüssigkeit oder Dampf in die Umgebung entlassen. Sie können sowohl schwer- als auch leichtflüchtig sein. Die Diffusionsfähigkeit beeinflusst die Funktion des Pheromons maßgeblich.[61] Alarmpheromone sind oft leichtflüchtig, um sich schnell durch Diffusion zu verbreiten. Es handelt sich daher oft um kurzkettige Stoffe mit relativ hohem Dampfdruck und geringer Komplexität.[44] Eine hohe Anforderung an die artspezifische Wirkung der Codierung wie bei Sexualpheromonen besteht nicht. Sexuallockstoffe weisen eine höhere Komplexität als die meisten Alarmpheromone auf, jedoch eine niedrigere molare Masse als Markierungspheromone, die dauerhaft ein Gebiet anzeigen.[62]
Bei fliegenden Insekten – wie Schmetterlingen – darf das Pheromon als Molekül nicht zu groß sein, da sonst Dampfdruck und Flüchtigkeit zu gering sind. So handelt es sich bei über 200 identifizierten Sexuallockstoffen von Schmetterlingsarten um mono- und bis-olefinische Fettaldehyde, Fettalkohole und deren Acetate mit Ketten von 10 bis 18 Kohlenstoffatomen.[44]
Je nach Funktion gibt es verschiedene Emissions- und Empfangsszenarien. Ameisen emittieren etwa Alarmpheromone stoßweise oder kontinuierlich in der meist windstillen Umgebung des Ameisenbaus. Spurenpheromone werden von einer Ameise als bewegliche Quelle ausgesandt. Die Sexualpheromone des Seidenspinners werden in diskreten Duftfäden in einem Luftstrom ausgestoßen.[61]

Männliche Monarchfalter emittieren keine flüchtigen Pheromone, sondern pheromonbeladene Nanoteilchen, Pheromon-Transfer-Partikel genannt, mit deren Hilfe sie Arrestants oder Aphrodisiakapheromone auf die Weibchen übertragen. Die Pheromon-Transfer-Partikel positionieren die Männchen auf ihren Pinselhaaren und verstreuen sie während des Balzflugs. Die Nanoteilchen bleiben auf den mit Pheromonrezeptoren ausgestatteten Fühlern der Weibchen haften, wo sie die Pheromone langsam abgeben und so zu einem lange anhaltenden Reiz für das Weibchen führen.[63]
Weibchen der arktischen Bärenspinnerspezies Pyrrharctia isabella emittieren ein Aerosol, das ausschließlich aus Sexualpheromontröpfchen besteht. Die dabei freigesetzte Pheromonmenge ist wesentlich größer als bei anderen bekannten weiblichen Faltern. Die scheinbare Verschwendung des Sexualpheromons erklärt sich mit der aufgrund des kurzen arktischen Frühlings knappen Zeitspanne, die ein erwachsenes Tier hat, um einen Fortpflanzungspartner zu finden.[64]
Die Empfänger nehmen Pheromone meist in einem Umfeld wahr, das durch die Präsenz vieler anderer Chemikalien geprägt ist. Um eine spezifische Wahrnehmung zu gewährleisten, muss die Pheromonchemikalie entweder so komplex sein, dass sie in der Natur nicht mehrfach vorkommt, oder das richtige Verhältnis mehrerer Einzelkomponenten muss den Reiz auslösen. Es hat sich jedoch gezeigt, dass nur in Ausnahmefällen eine einzige Substanz die Botschaft vermittelt. Oft muss ein Gemisch von Substanzen in sehr präzisen Mengenanteilen vorliegen, die neben der chemischen Struktur der einzelnen Pheromone den Informationsinhalt des Pheromoncocktails bestimmen.[44]

Die chemische Struktur von Pheromonen steht in direktem Zusammenhang mit ihrer Signalfunktion und Signalumgebung. An die Luft abgegebene Pheromone weisen oft eine Kohlenstoffkette von 5 bis 20 Atomen und eine molare Masse von etwa 80 bis 300 g·mol−1 auf. Bei einer Kohlenstoffkette von weniger als fünf Kohlenstoffatomen ist die Anzahl der möglichen Isomere gering und eine gezielte artspezifische Codierung schwierig.[62] Bei längeren Kohlenstoffketten steigt die Zahl der möglichen Isomere schnell an.
Periplanon B, das Sexualpheromon der Amerikanischen Großschabe, ist ein Beispiel einer komplexen Einzelsubstanz, auf die Männchen bereits in extrem geringen Mengen von 10−5 Nanogramm ansprechen.[65][66]
Biologische Eigenschaften

Der von einem Insektenweibchen ausgestoßene Sexualpheromoncocktail breitet sich windabwärts aus. Beim Empfängermännchen treffen die Moleküle auf die Antennen, wo der Empfang der Pheromone mittels olfaktorischer Zellen auf den Riechhaaren oder Sensillien erfolgt. Die Antennen adsorbieren etwa 30 % der in einem Luftstrom enthaltenen Pheromonmoleküle.[60] Die übrigen Moleküle treffen auf die äußere Körperdecke und werden dort enzymatisch abgebaut.
Die Pheromonmoleküle gelangen zunächst auf die Cuticula der Riechhaare und diffundieren über Poren in einen Porenkessel und von dort aus in Tubuli. Von dort diffundieren die Moleküle weiter zur Dendritenmembran.[44] Diese Membran besitzt Rezeptoren, die beim Empfang eines Pheromons über die Öffnung von Ionenkanälen eine Veränderung des elektrischen Widerstands hervorrufen und ein elektrisches Potential erzeugen, das einen Sinnesreiz zur Folge hat.[44] Schon ein einzelnes Pheromonmolekül kann einen Nervenimpuls auslösen.[60] Die Erkennung eines speziellen Pheromoncocktails erfordert jedoch eine gewisse Erregungshöhe verschiedener Zelltypen unterschiedlicher Spezifität.[60] Es wird angenommen, dass die von den verschiedenen Rezeptoren im Zentralnervensystem eingehenden charakteristischen Erregungen dort zu einem Erregungsmuster moduliert werden. Stimmt dieses Erregungsmuster, das vom Mengenverhältnis der empfangenen Pheromonmoleküle abhängt, mit der Codierung eines angeborenen Verhaltensmusters überein, führt dies zur Auslösung einer entsprechenden Verhaltensantwort, etwa dem Gegenwindanflug auf eine Pheromonquelle.[59]
Pheromonarten
Nach ihrer Wirkung lassen sich zwei Klassen von Pheromonen, die Primer- und die Releaserpheromone, unterscheiden. Unter bestimmten Bedingungen wirken gewisse Pheromone sowohl als Releaser- als auch als Primerpheromone.[67]
Releaserpheromone
Releaserpheromone haben eine kurze, unmittelbar verhaltenssteuernde Wirkung. Das erste entdeckte Pheromon, Bombykol, ist ein Beispiel dafür. Zu den Releaserpheromonen gehören typischerweise neben den bekannten Sexuallockstoffen unter anderem Aggregationspheromone, Dispersionspheromone, Alarmpheromone, Spurpheromone und Markierungspheromone.[68]
Aggregationspheromone

Aggregationspheromone werden von beiden Geschlechtern produziert und dienen der geschlechtsunspezifischen Anziehung von Individuen derselben Art. Diese sind zum Beispiel beim Borkenkäfer und anderen Käferarten, Zweiflüglern, Schnabelkerfen und Heuschrecken bekannt. Insekten nutzen Aggregationspheromone zur Verteidigung gegen Fressfeinde, bei der Partnerwahl und zur Überwindung der Resistenz von Wirtspflanzen bei einem Massenangriff. Eine Gruppe von Individuen an einem Standort wird unabhängig vom Geschlecht als Aggregation bezeichnet.[69] Die Aggregationspheromone spielen neben den Sexuallockstoffen eine bedeutende Rolle bei der Entwicklung von Pheromonfallen zur selektiven Schädlingsbekämpfung.[70]
Untersuchungen mittels Elektroantennogrammtechnik zeigten, dass Aggregationspheromone relativ hohe Rezeptorpotentiale geschlechterunspezifisch auslösten, wogegen die Sexualpheromone hohe Rezeptorpotentiale nur bei einem Geschlecht bewirken. Bei den Aggregationspheromonen handelt es sich daher möglicherweise um evolutionäre Vorläufer der Sexualpheromone.[39]
Sexualpheromone

Sexualpheromone signalisieren die Bereitschaft des weiblichen Tieres zur Paarung. Männliche Tiere emittieren ebenfalls Pheromone; sie enthalten Informationen über das Geschlecht und den Genotyp. Viele Insekten setzen Sexualpheromone frei; manche Schmetterlingsarten nehmen dabei das Pheromon noch in einer Entfernung von 10 Kilometern wahr. Die Sinneszellenantwort beim männlichen Seidenspinner beginnt bereits bei einer Konzentration von etwa 1000 Molekülen pro Kubikzentimeter Luft.[60] Das Duftsignal eines Weibchens löst, sobald ein gewisser Konzentrationsgrenzwert überschritten wird, beim Seidenspinnermännchen zunächst einen orientierten Gegenwindflug aus.[60] Bei anderen Arten wie dem Apfelwickler dagegen prüft das Männchen die stereochemische Reinheit des Lockstoffmoleküls. Sobald eine geringe Beimengung eines anderen Stereoisomers im Pheromoncocktail vorhanden ist, bleibt der Anflug zur Quelle aus.[60] Das andere Stereoisomer wirkt in diesem Fall als Repellent.[44] Manche Arten geben neben den Hauptkomponenten noch sogenannte Nahbereichskomponenten in geringer Menge ab, welche die Verhaltensreaktion beeinflussen.[60]
Fouragierende Honigbienen verbreiten den Duft von (Z)-11-Eicosen-1-ol. Bienenwolfweibchen lassen sich von diesem Duft leiten, um Honigbienen zu erbeuten. Die Bienenwolfmännchen nutzen diese Komponente und damit die existierende sensorische Präferenz der Weibchen für Bienenduft als Teil ihres Sexualpheromoncocktails, um diese anzulocken.[71]
Aphrodisiakapheromone
-_%2526_(S)-Oleane_Structural_Formula_V.1.svg.png.webp)
Aphrodisiaka-Pheromone stimulieren die Paarungsbereitschaft. Das Spiroacetal Olean etwa ist das Aphrodisiakapheromon der Olivenfruchtfliege (Bactrocera oleae). Nur das (R)-Enantiomer wirkt auf die Männchen, das (S)-Enantiomer ist bei ihnen unwirksam. Das Weibchen produziert das Racemat, spricht auf (R)- und (S)-Olean an und stimuliert sich damit auch selbst.[72]
Genau umgekehrt wirken sogenannte Anti-Aphrodisiaka. Nymphen der Bettwanze schützen sich mit einem solchen Pheromon, das ein bestimmtes Mischungsverhältnis der Aldehyde (E)-2-Hexenal, (E)-2-Octenal und 4-Oxo-(E)-2-Hexenal aufweist, gegen Begattungsversuche von Bettwanzenmännchen. Dieses bohrt direkt ein Loch in den Hinterleib geschlechtsreifer weiblicher Wanzen und injiziert dort seine Spermien (traumatische Insemination). Für begattete Nymphen kann eine solche Verletzung jedoch tödlich sein.[73]
Alarmpheromone
Einige Insektenarten geben bei einem Angriff Alarmpheromone ab. Diese lösen entweder die Flucht oder gesteigerte Aggression aus. Bei Bienen etwa sind zwei Alarmpheromon-Gemische bekannt. Eines wird durch die Koschewnikow-Drüse in der Nähe des Stachels freigesetzt und enthält mehr als 40 verschiedene Verbindungen, wie das bereits von Butler in der Wirkung beschriebene Isoamylacetat, daneben Butylacetat, 1-Hexanol, 1-Butanol, 1-Octanol, Hexylacetat, Octylacetat und 2-Nonanol. Diese Komponenten haben eine niedrige molare Masse, sind flüchtig und sind die unspezifischsten aller Pheromone. Alarmpheromone werden freigesetzt, wenn eine Biene ein anderes Tier sticht, um andere Bienen anzuziehen und zum Angriff zu verleiten. Rauch unterdrückt die Wirkung von Alarmpheromonen, was von Imkern ausgenutzt wird.[74]
Das andere Alarmpheromon der Honigbiene enthält hauptsächlich 2-Heptanon, eine ebenfalls flüchtige Substanz, die von den Kieferdrüsen freigesetzt wird.[75] Diese Komponente hat einen abstoßenden Effekt auf räuberische Insekten. Der Alarmpheromoncocktail der Bettwanze enthält ungesättigte Hexen- und Octenaldehyde, die in von Wanzen befallenen Zimmern als charakteristischer, süßlicher Geruch wahrgenommen werden.[76][77]
Markierungs- und Dispersionspheromone
Gewisse Insekten wie die Kirschfruchtfliege markieren ihre Eiablageplätze in einer Weise, dass andere Weibchen derselben Art den Ort meiden und ihre Eier an anderen Plätzen ablegen, um unter dem Nachwuchs Konkurrenz um Nahrung zu vermeiden.[78] Auch territoriale soziale Insekten, wie zum Beispiel Kolonien von Ameisen, markieren von ihnen beanspruchte Territorien mit Pheromonen.[79]
Zu den Markierungspheromonen gehören die Dispersionspheromone, mit denen zum Beispiel Borkenkäfer eine Überbesiedlung eines Baumes verhindern.[80] Die Weibchen und Nymphen der Deutschen Schabe übertragen Dispersionspheromone im direkten Kontakt über ihren Speichel. Diese dienen im Nymphenstadium der Abschreckung erwachsener Schaben und damit zum Schutz vor Kannibalismus. Bei erwachsenen Tieren verhindern sie die Übersiedlung eines Lebensraums.[81]
Spurpheromone

Spurpheromone sind vor allem bei in Kolonien lebenden Insekten bekannt, die ihre Pfade mit schwerflüchtigen Substanzen wie höhermolekularen Kohlenwasserstoffen markieren. Vor allem Ameisen markieren oft auf diese Weise den Weg von einer Nahrungsquelle zum Nest.[82] Solange die Nahrungsquelle besteht, wird die Spur erneuert. Beim Versiegen der Nahrungsquelle übersprühen die Ameisen das Spurpheromon mit einem abstoßenden Pheromon.[83] Der US-amerikanische Naturforscher Charles William Beebe berichtete 1921 über das Phänomen der Ameisenmühle, das Spurpheromone bei Wanderameisen auslösen können: Werden die Tiere von der Hauptspur der Kolonie getrennt, folgen die blinden Ameisen den Pheromonspuren vor ihnen laufender Ameisen. Diese laufen in großen Kreisen bis zu vollkommener Erschöpfung oder dem Tod, ohne zur Kolonie zurückzufinden.[84]
Rekrutierungspheromone
Rekrutierungspheromone sind als Element der chemischen Kommunikation weit verbreitet bei sozialen Insekten und wurden für Bienen, Termiten und Ameisen nachgewiesen. Diese Pheromone werden von Insekten verwendet, um andere Mitglieder der Kolonie zur Nahrungssuche bei einer Nahrungsquelle anzuregen.[68] Hummeln führen einen dem Bienentanz ähnlichen Tanz auf, der primär zur Verteilung von Rekrutierungspheromonen dient.[85]
Primerpheromone
In der Ordnung der Hautflügler findet sich die größte Gruppe eusozialer Insekten, darunter viele Bienen, insbesondere der Unterfamilie Apinae, Ameisen sowie einige Arten der Faltenwespen, insbesondere der Unterfamilie der Echten Wespen. Die Merkmale sind oft das Vorhandensein einer reproduktiven Königin sowie Kasten mit spezialisierten Arbeiterinnen und Soldaten. Termiten bilden die zweite große Gruppe eusozial lebender Insekten. Die Kolonien sind in verschiedene Kasten unterteilt, mit einer Königin und einem König als reproduktionsfähige Individuen, Arbeitern und Soldaten, die die Kolonie verteidigen.[86] Primerpheromone haben einen großen Einfluss auf die Organisation der von Hautflüglern gebildeten Hymenopterenstaaten und von Termitenkolonien. Diese Pheromone beeinflussen das Hormonsystem des Empfängers; oft greifen sie über eine Signalkaskade in den Stoffwechsel ein oder aktivieren Proteine, die an die DNA binden können. Im Gegensatz zu den Releaserpheromonen sind die Primerpheromone weniger gut untersucht. So war lange Zeit nur ein Primerpheromon, das 9-ODA, bekannt.
Primerpheromone der Bienen
Ein bekanntes Beispiel für Primerpheromone sind die Bienenköniginnenpheromone.[87] Diese Pheromone steuern das soziale Verhalten, die Instandhaltung der Waben, das Ausschwärmen und die Ausbildung der Eierstöcke der Arbeitsbienen. Bei den Komponenten handelt es sich um Carbonsäuren und aromatische Verbindungen. (E)-9-Oxo-dec-2-ensäure (9-ODA) unterdrückt beispielsweise die weitere Zucht von Königinnen und hemmt die Entwicklung der Eierstöcke von Arbeitsbienen. Es handelt sich auch um ein starkes Sexualpheromon für Drohnen auf dem Hochzeitsflug.[88]
Bruterkennungspheromone werden von Larven und Puppen emittiert und halten Arbeiterbienen davon ab, den Stock zu verlassen, solange noch Nachwuchs zu pflegen ist. Weiterhin unterdrücken sie die Ausbildung der Eierstöcke bei den Arbeitsbienen. Die Pheromone bestehen aus einer Mischung von zehn Fettsäureestern, unter anderem Glyceryl-1,2-dioleat-3-palmitat.[89] Arbeiterpuppen enthalten 2 bis 5, Drohnenpuppen etwa 10 und Königinnenpuppen 30 Mikrogramm des Pheromons.
Ältere, fouragierende Arbeitsbienen setzen Ölsäureethylester frei, was die Entwicklung der Ammenbienen hemmt und diese länger zur Brutpflege veranlasst.[90] Der Ölsäureester wirkt als Primerpheromon und stabilisiert das Verhältnis von brutpflegenden und nahrungsbeschaffenden Bienen. Die Sammlerinnen produzieren es aus mit Spuren von Ethanol versetztem Nektar, den sie an die Ammenbienen verfüttern. Deren Entwicklung wird dadurch solange verzögert, bis die Zahl der älteren Sammlerinnen abnimmt und damit die Exposition der Ammenbienen mit Ölsäureethylester.
Kastendeterminierende Pheromone
Die Gelbfüßige Bodentermite nutzt Terpene wie γ-Cadinen und γ-Cadinenal als kastenstimulierende oder -hemmende Primerpheromone. Diese unterstützen das Juvenilhormon bei der Bestimmung über die Position totipotenter Arbeiter im Kastensystem.[91] Bei Ameisen besitzen die weiblichen Larven einige Zeit Bipotentialität und damit die Möglichkeit, sich entweder als Königinnen oder Arbeiterinnen zu entwickeln. Zu einem gewissen Zeitpunkt der Larvenentwicklung bestimmt die weitere Ernährung das Schicksal der Larve. Wird der Juvenilhormon-Titer über einen bestimmten Schwellenwert gehoben, entwickeln sich Gynomorphe, ansonsten Arbeiterinnen. Die Steuerung der Larvenernährung wird über ein Primerpheromon der Ameisenkönigin gesteuert.[92]
Anwendung
Im 19. Jahrhundert entkamen dem Entomologen Étienne Léopold Trouvelot in Massachusetts Schwammspinner, die sich bis zur Mitte des 20. Jahrhunderts über die gesamten USA ausbreiteten und heute zu den am meisten gefürchteten Schädlingen zählen. Bereits 1898 unternahmen Edward Forbush und Charles Fernald Versuche, die Population des Schwammspinners durch Anlockung der Männchen in Fallen, die mit lockenden Weibchen besetzt waren, einzudämmen.[93] Das Landwirtschaftsministerium der Vereinigten Staaten führte diese Versuche in den 1930er Jahren fort, wobei zur Attraktion männlicher Falter Extrakte weiblicher Abdominalspitzen eingesetzt wurden.[94] Die Anwendung von Insektenpheromonen im Pflanzenschutz ist vor allem seit den ersten Synthesen intensiv untersucht worden, mit dem Ziel, umweltschonende Methoden zur Kontrolle der Populationsdynamik zu entwickeln.[95]

Im Pflanzenschutz ist der Einsatz von Pheromonen in Lockstofffallen zur Bekämpfung von Insekten gängige Praxis. Dabei können die Insekten angelockt werden, um sie mit einem Insektizid oder physikalisch zu töten, um sie einzufangen oder zum Monitoring. Borkenkäfer werden mit Aggregationspheromonen angelockt, um sie in Fallen zu fangen. Der Lockstoff wird normalerweise beim Einbohren in das Fichtenholz freigesetzt und signalisiert, dass der Baum besiedelt werden kann. Die Borkenkäferfalle ist ein wichtiges Instrument zur Bekämpfung der Borkenkäfer.[96] Die Verwendung von Lockstofffallen birgt jedoch das Problem, dass das Pheromon gegebenenfalls als Kairomon wirkt und somit räuberische Insekten anlockt. Durch die Reduktion der Population natürlicher Fressfeinde des Borkenkäfers wirkt die Pheromonfalle in diesem Falle kontraproduktiv.[97] Das Monitoring mittels Lockstofffallen, etwa Fensterfallen, dient der quantitativen Erfassung von Schädlingen, um sie mit Insektiziden in Abhängigkeit von der festgestellten Aktivität gezielter zu bekämpfen. Daneben werden sie bei der Identifizierung neuer Arten eingesetzt.[98]
Fangbäume funktionieren nach demselben Prinzip wie Lockstofffallen. Die Borkenkäfer des Erstbefalls locken durch Aggregationspheromone weitere Artgenossen an. Als Fangbäume eignet sich Sturmholz, das zur Verstärkung der Lockwirkung mit Pheromondispensern ausgestattet werden kann. Die so präparierten Bäume lenken anfliegende Borkenkäfer vom Bestand ab und binden diese an kontrollierbare Stämme. Die Verwendung von Fangbäumen erfordert eine regelmäßige Kontrolle der Bäume. Beim Auftreten von Larvengängen werden die Bäume entrindet, wobei Larven und Puppen vertrocknen. Gegebenenfalls kann der befallene Baum mit Insektiziden behandelt werden oder er wird verbrannt, um den Ausflug der nächsten Generation zu unterbinden.[99]
Eiablageverhindernde Markierungspheromone sind in der Insektenwelt weit verbreitet. In verschiedenen Experimenten konnte die Möglichkeit einer Kontrolle der Populationsdynamik durch diese Pheromone aufgezeigt werden.[100] Die Anwendung des eiablageverhindernden Markierungspheromons der Kirschfruchtfliegen, die sich zum Beispiel mit Gelbtafeln nicht bekämpfen lassen, reduzierte den Befall der Kirschen um 90 %.[101]
Eine weitere Anwendung ist die Verwirrmethode oder Paarungsstörung. Dabei wird eine hohe Stoffkonzentration von künstlich hergestellten Pheromonen ausgebracht. Dadurch ist es den männlichen Tieren nicht mehr möglich, den Pheromonen der Weibchen zu folgen, wodurch die Vermehrung des Schädlings behindert wird. Die Verwirrmethode wirkt artspezifisch.[102] Sie ist bei genügender Ausbringung von Dispensern meist erfolgreich in Bezug auf eine Art, teilweise besetzen jedoch verwandte Arten die freiwerdende ökologische Nische.
Bienen nutzen das Nasanov-Pheromon, um Arbeitsbienen zurück zum Stock zu führen. Das Pheromon enthält Terpene wie Geraniol und Citral. Imker nutzen ein künstlich hergestelltes Produkt, um Bienen zu einem ungenutzten Bienenstock zu locken.[103] Das Verfahren eignet sich zum Fang afrikanisierter Honigbienen in Fangboxen.[104]
Toxikologie
Toxikologische Untersuchungen wurden hauptsächlich im Zusammenhang mit der Zulassung von Pheromonfallen und -dispensern durchgeführt. Eine Gesundheitsgefährdung ist aufgrund der großen chemischen Vielfalt der Pheromone nicht allgemein zu beurteilen, wird jedoch meist ausgeschlossen, weil nur geringe Mengen emittiert werden. In höheren Dosen führen jedoch oral verabreichte Pheromone wie Cantharidin in seltenen Fällen zum Tod.[105]
Nachweis
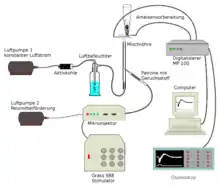
Die kommerzielle Anwendung im Pflanzenschutz intensivierte die Untersuchung von Pheromonen und führte zur Entwicklung hochempfindlicher Analysemethoden.[106] Die Identifizierung eines Pheromons verläuft über mehrere Stufen. Zunächst wird ein Extrakt des Pheromons gewonnen. Dies erfolgt nach der schon von Butenandt angewandten Methode der Extraktion von Drüsen oder ganzen Tieren mit einem leicht verdampfbaren Lösungsmittel, idealerweise zum Zeitpunkt hoher Pheromonproduktion. Alternativ wird das Pheromon an Aktivkohle aus der Gasphase adsorbiert und mit wenig Lösungsmittel ein Extrakt gewonnen.[107] Für sehr geringe Spuren eignet sich die Festphasenmikroextraktion. Zur Identifizierung werden die Extrakte beziehungsweise die Festphasenmikroextraktionsproben mittels Gaschromatographie mit Massenspektrometrie-Kopplung untersucht.[107]
Zur Untersuchung der biologischen Aktivität von Insektenpheromonen eignet sich die Elektroantennogrammtechnik.[108][59] Eine in den Antennenhauptstamm und einen Antennenast eingebrachte Elektrode misst dabei die Änderung der elektrische Spannung als Funktion der Konzentration von auf der Antenne auftreffenden Pheromonmolekülen, die durch einen Luftstrom in definierter Weise zur Antenne transportiert werden.[59] Durch Variation des Pheromonmoleküls lässt sich der Einfluss bestimmter funktioneller Gruppen ermitteln, die mit den chiralen Elementen der Rezeptoren wechselwirken.[59]
Die Kopplung von Gaschromatographie und Elektroantennogramm erlaubt die Überprüfung der biologischen Aktivität der in einem Extrakt vorliegenden Verbindungen.[59] Die Form des Elektroantennogramms ist abhängig von der Duftkomponente im Luftstrom, die Amplitude steigt mit der Konzentration und der Strömungsgeschwindigkeit der Luft an.[109]
Literatur
- Edward O. Wilson, W. H. Bossert (1963): Chemical communication among animals. In: Recent Progress in Hormone Research. Bd. 19, S. 673–716, PMID 14284035.
- Hans Jürgen Bestmann, Otto Vostrowsky (1993): Chemische Informationssysteme der Natur: Insektenpheromone. In: Chemie in unserer Zeit. Bd. 27, Nr. 3, S. 127–133, doi:10.1002/ciuz.19930270304.
- Stefan Schulz: The Chemistry of Pheromones and Other Semiochemicals II. Springer, 2005, ISBN 3-540-21308-2.
- R. T. Carde, A. K. Minks: Insect Pheromone Research: New Directions. Springer, 1997, ISBN 978-0-412-99611-5.
Weblinks
- Insektenpheromon-Datenbank. In: pherobase.com. Abgerufen am 15. November 2013.
Einzelnachweise
- Albert Gossauer: Struktur und Reaktivität der Biomoleküle. Verlag Helvetica Chimica Acta, Zürich 2006, S. 134, ISBN 978-3-906390-29-1.
- Peter Karlson, Adolf Butenandt: Pheromones (Ectohormones) in Insects. In: Annual Review of Entomology. 4, 1959, S. 39–58, doi:10.1146/annurev.en.04.010159.000351.
- Peter Karlson, Martin Lüscher: Pheromones: a New Term for a Class of Biologically Active Substances. In: Nature. 183, 1959, S. 55–56, doi:10.1038/183055a0.
- Wilhelm Gemoll: Griechisch-Deutsches Schul- und Handwörterbuch. Neubearbeitung. Oldenbourg Schulbuchverlag, München 2006, ISBN 3-637-00234-5.
- Peter Karlson, Martin Lüscher: The Proposed Biological Term „Pheromone“. In: Nature. 183, 1959, S. 1835–1835, doi:10.1038/1831835b0.
- Albrecht Bethe: Vernachlässigte Hormone. In: Die Naturwissenschaften. 20, 1932, S. 177–181, doi:10.1007/BF01504737.
- D. A. Nordlund, R. L. Jones, W. J. Lewis: Semiochemicals: Their Role in Pest Control. Wiley, New York 1981, ISBN 0-471-05803-3, S. 13–28.
- Peter Karlson, Martin Lüscher: Pheromone. In: Die Naturwissenschaften. 46, 1959, S. 63–64, doi:10.1007/BF00599084.
- Edward O. Wilson: Chemical communication among workers of the fire ant Solenopsis saevissima (Fr. Smith) 1. The Organization of Mass-Foraging. In: Animal Behaviour. 10, 1962, S. 134–147, doi:10.1016/0003-3472(62)90141-0.
- Edward O. Wilson, William H. Bossert: Chemical communication among animals. In: Recent progress in hormone research. 19 (1963): S. 673.
- Otto Vostrowsky, Wolfram Sander, Lothar Jaenicke, Lothar Jaenicke: Wissenschaft Aktuell. In: Chemie in unserer Zeit. 30, 1996, S. 252–255, doi:10.1002/ciuz.19960300506.
- Vincent G. Dethier, Barton L. Browne: The Designation of Chemicals in Terms of the Responses They Elicit from Insects 1. In: Journal of Economic Entomology. 53.1 (1960), S. 134–136.
- Masayuki Sakuma, Hiroshi Fukami: Aggregation arrestant pheromone of the German cockroach, Blattella germanica (L.) (Dictyoptera: Blattellidae): Isolation and structure elucidation of blattellastanoside-A and -B. In: Journal of Chemical Ecology. 19, 1993, S. 2521–2541, doi:10.1007/BF00980688.
- Bonnie Blaimer: Zur Struktur und Funktion der androconialen Organe und Sekrete bei Brassolini (Lepidoptera). Abgerufen am 23. April 2014.
- T. Pankiw, Z-Y. Huang, M. L. Winston, G. E. Robinson: Queen mandibular gland pheromone influences worker honey bee (Apis mellifera L.) foraging ontogeny and juvenile hormone titers. In: Journal of Insect Physiology. 44, 1998, S. 685–692, doi:10.1016/S0022-1910(98)00040-7.
- Donald A. Nordlund, W. J. Lewis: Terminology of chemical releasing stimuli in intraspecific and interspecific interactions. In: Journal of Chemical Ecology. 2, 1976, S. 211–220, doi:10.1007/BF00987744.
- J. B. Free, J. Simpson: The alerting pheromones of the honeybee. In: Zeitschrift für Vergleichende Physiologie. 61, 1968, S. 361–365, doi:10.1007/BF00428008.
- Charles E. Raven: John Ray, Naturalist: His Life and Works. 506 Seiten, Verlag Cambridge University Press (1950), S. 395
- Adolf Butenandt: Fettalkohole als Sexual-Lockstoffe der Schmetterlinge. In: Fette, Seifen, Anstrichmittel. 64, 1962, S. 187–192, doi:10.1002/lipi.19620640302.
- Maurice Maeterlinck: The Life of the Bee. In: Projekt Gutenberg. 1. Mai 1901, abgerufen am 9. Mai 2014.
- W. M. Bayliss, E. H. Starling: Die chemische Koordination der Funktionen des Körpers. In: Ergebnisse der Physiologie. 5, 1906, S. 664–697, doi:10.1007/BF02321027.
- Adolf Butenandt, Rüdiger Beckmann, D. Stamm: Über den Sexuallockstoff des Seidenspinners. In: Zeitschrift für Naturforschung B. 14, 1959, S. 283–284 (online).
- Adolf Butenandt, Erich Hecker, Manfred Hopp, Wolfgang Koch: Über den Sexuallockstoff des Seidenspinners, IV. Die Synthese des Bombykols und der cis-trans-Isomeren Hexadecadien-(10.12)-ole-(1). In: Justus Liebigs Annalen der Chemie. 658, 1962, S. 39–64, doi:10.1002/jlac.19626580105.
- Adolf Butenandt, Rüdiger Beckmann, Erich Hecker: Über den Sexuallockstoff des Seidenspinners, I. Der biologische Test und die Isolierung des reinen Sexuallockstoffes Bombykol. In: Hoppe-Seyler’s Zeitschrift für physiologische Chemie. 324, 1961, S. 71–83, doi:10.1515/bchm2.1961.324.1.71.
- Edward O. Wilson: A Chemical Releaser of Alarm and Digging Behavior in the Ant Pogonomyrmex Badius (Latreille). In: Psyche: A Journal of Entomology. 65, 1958, S. 41–51, doi:10.1155/1958/57483.
- C. G. Butler, R. K. Callow, N. C. Johnston: The Isolation and Synthesis of Queen Substance, 9-oxodec-trans-2-enoic Acid, a Honeybee Pheromone. In: Proceedings of the Royal Society B: Biological Sciences. 155, 1962, S. 417–432, doi:10.1098/rspb.1962.0009.
- Edward O. Wilson: Chemical communication among workers of the fire ant Solenopsis saevissima (Fr. Smith) 1. The Organization of Mass-Foraging. In: Animal Behaviour. 10, 1962, S. 134–147, doi:10.1016/0003-3472(62)90141-0.
- Edward O. Wilson, William H. Bossert: Chemical communication among animals. In: Recent progress in hormone research. 19, 1963, S. 673.
- Gerhard Kasang, Karl Ernst Kaißling, Otto Vostrowsky, Hans Jürgen Bestmann: Bombykal, eine zweite Pheromonkomponente des Seidenspinners Bombyx mori L. In: Angewandte Chemie. 90, 1978, S. 74–75, doi:10.1002/ange.19780900132.
- Ashok K. Raina, Jerome A. Klun: Brain Factor Control of Sex Pheromone Production in the Female Corn Earworm Moth. In: Science. 225, 1984, S. 531–533, doi:10.1126/science.225.4661.531.
- C. Löfstedt u. a.: No linkage between genes controlling female pheromone production and male pheromone response in the European corn borer, Ostrinia nubilalis Hübner (Lepidoptera; Pyralidae). In: Genetics. 123.3, 1989, S. 553–556.
- Peter Karlson: Evolution der chemischen Kommunikation im Tierreich. In: Information und Kommunikation. Naturwissenschaftliche, medizinische und technische Aspekte. Wissenschaftliche Verlagsgesellschaft mbH, Stuttgart 1985, ISBN 3-8047-0814-5, S. 23–42.
- 10 facts on malaria. In: WHO.int. März 2014, abgerufen am 9. Juli 2014.
- Y. Mao, X. Xu, W. Xu, Y. Ishida, W. S. Leal, J. B. Ames, J. Clardy: Crystal and solution structures of an odorant-binding protein from the southern house mosquito complexed with an oviposition pheromone. In: Proceedings of the National Academy of Sciences. 107, 2010, S. 19102–19107, doi:10.1073/pnas.1012274107.
- Michael Boppre, Dietrich Schneider: Pyrrolizidine alkaloids quantitatively regulate both scent organ morphogenesis and pheromone biosynthesis in male Creatonotos moths (Lepidoptera: Arctiidae). In: Journal of Comparative Physiology A. 157, 1985, S. 569–577, doi:10.1007/BF01351351.
- Athula B. Attygalle, E. David Morgan: Pheromones in Nanogram Quantities: Structure Determination by Combined Microchemical and Gas Chromatographic Methods [New Analytical Methods (35)]. In: Angewandte Chemie. International Edition in English. 27, 1988, S. 460–478, doi:10.1002/anie.198804601.
- K. E. Kaissling, G. Kasang, H. J. Bestmann, W. Stransky, O. Vostrowsky: A new pheromone of the silkworm moth Bombyx mori. In: Naturwissenschaften. 65, 1978, S. 382–384, doi:10.1007/BF00439702.
- Gerhard Kasang, Karl Ernst Kaissling, Otto Vostrowsky, Hans Jürgen Bestmann: Bombykal, a Second Pheromone Component of the Silkworm MothBombyx mori L. In: Angewandte Chemie. International Edition in English. 17, 1978, S. 60–60, doi:10.1002/anie.197800601.
- Hermann Levinson, Anna Levinson: Zu Struktur und Wirkungsweise der Pheromondrüsen vorratsschädlicher Insektenarten. In: Forschungsarbeiten über Insekten und andere Gliedertiere sowie deren Kulturgeschichte – Nachtrag 2008. 10. Oktober 2008, abgerufen am 23. April 2014.
- C. G. Butler, D. H. Calam: Pheromones of the honey bee – The secretion of the Nassanoff gland of the worker. In: Journal of Insect Physiology. 15, 1969, S. 237–244, doi:10.1016/0022-1910(69)90271-6.
- Ken Naumann, Mark L. Winston, Keith N. Slessor, Glenn D. Prestwich, Francis X. Webster: Production and transmission of honey bee queen (Apis mellifera L.) mandibular gland pheromone. In: Behavioral Ecology and Sociobiology. 29, 1991, S. 321–332, doi:10.1007/BF00165956.
- Julie A. Tillman, Steven J. Seybold, Russell A. Jurenka, Gary J. Blomquist: Insect pheromones – an overview of biosynthesis and endocrine regulation. In: Insect Biochemistry and Molecular Biology. 29, 1999, S. 481–514, doi:10.1016/S0965-1748(99)00016-8.
- Wendell L. Roelofs: Chemistry of sex attraction. In: Proceedings of the National Academy of Sciences. 92.1, 1995, S. 44–49.
- Hans Jürgen Bestmann, Otto Vostrowsky: Chemische Informationssysteme der Natur: Insektenpheromone. In: Chemie in unserer Zeit. 27, 1993, S. 123–133, doi:10.1002/ciuz.19930270304.
- R. T. Carde, A. K. Minks: Insect Pheromone Research: New Directions. Springer, 1997, ISBN 978-0-412-99611-5, S. 5.
- Michel Cusson, Stephen S. Tobe, Jeremy N. McNeil: Juvenile hormones: Their role in the regulation of the pheromonal communication system of the armyworm moth, Pseudaletia unipuncta. In: Archives of Insect Biochemistry and Physiology. 25, 1994, S. 329–345, doi:10.1002/arch.940250408.
- Yi‐Kai Tea, Jonathan Soong Wei, Ethan P. Beaver, David J. Lohman: Kleptopharmacophagy? Milkweed butterflies scratch and imbibe from Apocynaceae‐feeding caterpillars. In: Ecology. , doi:10.1002/ecy.3532.
- Jerrold Meinwald: Alkaloids and isoprenoids as defensive and signalling agents among insects. In: Pure Appl. Chem. 62, 1990, S. 1325–1328.
- Gadi V. P. Reddy, Angel Guerrero: Interactions of insect pheromones and plant semiochemicals. In: Trends in Plant Science. 9, 2004, S. 253–261, doi:10.1016/j.tplants.2004.03.009.
- Michael Boppré: Pheromonbiologie am Beispiel der Monarchfalter (Danaidae). In: Biologie in unserer Zeit. 7, 1977, S. 161–169, doi:10.1002/biuz.19770070604.
- Michael Boppré, Dietrich Schneider: Pyrrolizidine alkaloids quantitatively regulate both scent organ morphogenesis and pheromone biosynthesis in male Creatonotos moths (Lepidoptera: Arctiidae). In: Journal of Comparative Physiology A. 157, 1985, S. 569–577, doi:10.1007/BF01351351.
- Michael Boppré: Pharmakophagie: Drogen, Sex und Schmetterlinge. In: Biologie in unserer Zeit. 25, 1995, S. 8–17, doi:10.1002/biuz.19950250103.
- Karl Ziegler, Günther Otto Schenck, E. W. Krockow: Synthese des Cantharidins. In: Die Naturwissenschaften. 29, 1941, S. 390–391, doi:10.1007/BF01479894.
- Fritz Eiden: Cantharidin: Hochzeitsgabe, Schutz- und Lockstoff, Blasenzieher und Enzymhemmer. In: Chemie in unserer Zeit. 40, 2006, S. 12–19, doi:10.1002/ciuz.200600354.
- Dieter Enders, Herbert Eichenauer: Asymmetrische Synthese von Ameisen-Alarmpheromonen – α-Alkylierung von acyclischen Ketonen mit praktisch vollständiger asymmetrischer Induktion. In: Angewandte Chemie. 91, 1979, S. 425–427, doi:10.1002/ange.19790910512.
- Kenji Mori, Takuya Tashiro: Useful Reactions in Modern Pheromone Synthesis. In: Current Organic Synthesis. 1, 2004, S. 11–29, doi:10.2174/1570179043485466.
- Hans Jürgen Bestmann, Otto Vostrowsky: Selected topics of the Wittig reaction in the synthesis of natural products. In: Wittig Chemistry. Springer, Berlin/Heidelberg 1983, ISBN 0-387-11907-8, S. 85–163.
- Bao-Jian Ding, Per Hofvander, Hong-Lei Wang, Timothy P. Durrett, Sten Stymne, Christer Löfstedt: A plant factory for moth pheromone production. In: Nature Communications. 5, 2014, S. 1–7 doi:10.1038/ncomms4353.
- Hans Jürgen Bestmann: Synthese und Wirkungsweise von Pheromonen. In: Information und Kommunikation. Naturwissenschaftliche, medizinische und technische Aspekte. Wissenschaftliche Verlagsgesellschaft mbH, Stuttgart 1985, ISBN 3-8047-0814-5, S. 301–316.
- Ernst Priesner: Pheromone als Sinnesreize. In: Information und Kommunikation. Naturwissenschaftliche, medizinische und technische Aspekte. Wissenschaftliche Verlagsgesellschaft mbH, Stuttgart 1985, ISBN 3-8047-0814-5, S. 207–226.
- William H. Bossert, Edward O. Wilson: The analysis of olfactory communication among animals. In: Journal of Theoretical Biology. 5, 1963, S. 443–469, doi:10.1016/0022-5193(63)90089-4.
- Tristram D. Wyatt: Pheromones and Animal Behaviour. Communication by Smell and Taste. Cambridge University Press, 2003, ISBN 0-521-48526-6, S. 13.
- Michael Boppré: Pheromon-Transfer-Partikel auf einem Duftpinselhaar eines Monarchfalters (Danaus formosa). In: Naturwissenschaftliche Rundschau. 29.9, 1976.
- Stuart B. Krasnoff, Wendell L. Roelofs: Sex pheromone released as an aerosol by the moth Pyrrharctia isabella. In: Nature. 333, 1988, S. 263–265, doi:10.1038/333263a0.
- Ring Cardé: Advances in Insect Chemical Ecology. Cambridge University Press, 2011, ISBN 978-0-521-18893-7, S. 190.
- Kentaro Okada, Masataka Mori, Kazuko Shimazaki, Tatsuji Chuman: Behavioral responses of male Periplaneta americana L. to female sex pheromone components, periplanone-A and periplanone-B. In: Journal of Chemical Ecology. 16, 1990, S. 2605–2614, doi:10.1007/BF00988072.
- Otto Vostrowsky, Wolfram Sander, Lothar Jaenicke, Lothar Jaenicke: Wissenschaft Aktuell. In: Chemie in unserer Zeit. 30, 1996, S. 252–255, doi:10.1002/ciuz.19960300506.
- Fred E. Regnier, John H. Law: Insect pheromones. In: Journal of lipid research. 9.5, 1968, S. 541–551.
- S. P. Foster and, M. O. Harris: Behavioral manipulation Methods for Insect Pest-Management. In: Annual Review of Entomology. 42, 1997, S. 123–146, doi:10.1146/annurev.ento.42.1.123.
- Jean Pierre Vite, Wittko Francke: Waldschutz gegen Borkenkäfer: Vom Fangbaum zur Falle. In: Chemie in unserer Zeit. 19, 1985, S. 11–21, doi:10.1002/ciuz.19850190103.
- Gudrun Herzner, Thomas Schmitt, K. Eduard Linsenmair, Erhard Strohm: Prey recognition by females of the European beewolf and its potential for a sensory trap. In: Animal Behaviour. 70, 2005, S. 1411–1418, doi:10.1016/j.anbehav.2005.03.032.
- Bernd Schäfer: Naturstoffe in der chemischen Industrie. Spektrum Akademischer Verlag, 2007, ISBN 978-3-8274-1614-8, S. 522–524.
- Vincent Harraca, Camilla Ryne, Rickard Ignell: Nymphs of the common bed bug (Cimex lectularius) produce anti-aphrodisiac defence against conspecific males. In: BMC Biology. 8, 2010, S. 121, doi:10.1186/1741-7007-8-121.
- P. Kirk Visscher, Richard S. Vetter, Gene E. Robinson: Alarm pheromone perception in honey bees is decreased by smoke (Hymenoptera: Apidae). In: Journal of Insect Behavior. 8, 1995, S. 11–18, doi:10.1007/BF01990966.
- D. A. Shearer, R. Boch: 2-Heptanone in the Mandibular Gland Secretion of the Honey-bee. In: Nature. 206, 1965, S. 530–530, doi:10.1038/206530a0.
- Hermann Levinson, Anna Levinson: Die Bettwanze, ein Ektoparasit der Fledermaus und des Menschen in eiszeitlichen Höhlen und zeitgemäßen Wohnstätten. In: Forschungsarbeiten über Insekten und andere Gliedertiere sowie deren Kulturgeschichte. 6. Juli 2008, abgerufen am 1. Juli 2014.
- Jutta Klasen, Gabriele Schrader: Bettwanzen, Biologie des Parasiten und Praxis der Bekämpfung. (PDF) In: Umweltbundesamt FG IV 1.4 Gesundheitsschädlinge und ihre Bekämpfung. 23. März 2011, abgerufen am 1. Juli 2014.
- E. F. Boller, M. Aluja: Oviposition deterring pheromone in Rhagoletis cerasi L. In: Journal of Applied Entomology. 113, 1992, S. 113–119, doi:10.1111/j.1439-0418.1992.tb00644.x
- Bert Hölldobler, Edward O. Wilson: Colony-specific territorial pheromone in the African weaver ant Oecophylla longinoda (Latreille). In: Proceedings of the National Academy of Sciences (PNAS). USA, Vol. 74, No. 5, 1977, S. 2072–2075.
- Petia Koleva u. a.: Untersuchungen zur Effizienz von insektizidbehandelten Fanghölzern gegen den Buchdrucker Ips typographus (Coleoptera, Curculionidae). In: Forstschutz Aktuell. 54, 2012, S. 16–21.
- M. Faulde, M. E. A. Fuchs, W. Nagl: Further characterization of a dispersion-inducing contact pheromone in the saliva of the German cockroach, Blattella germanica L. (Blattodea: Blattellidae). In: Journal of Insect Physiology. 36, 1990, S. 353–359, doi:10.1016/0022-1910(90)90017-A.
- M. D. Breed, B. Bennett: Mass recruitment to nectar sources in Paraponera clavata: A field study. In: Insectes Sociaux. 32, 1985, S. 198–208, doi:10.1007/BF02224233.
- E. J. H. Robinson, K. E. Green, E. A. Jenner, M. Holcombe, F. L. W. Ratnieks: Decay rates of attractive and repellent pheromones in an ant foraging trail network. In: Insectes Sociaux. 55, 2008, S. 246–251, doi:10.1007/s00040-008-0994-5.
- N. R. Franks, N. Gomez, S. Goss, J. L. Deneubourg: The blind leading the blind in army ant raid patterns: Testing a model of self-organization (Hymenoptera: Formicidae). In: Journal of Insect Behavior. 4, 1991, S. 583–607, doi:10.1007/BF01048072.
- Angeles Mena Granero, Jose M. Guerra Sanz, Francisco J. Egea Gonzalez, Jose L. Martinez Vidal, Anna Dornhaus, Junaid Ghani, Ana Roldan Serrano, Lars Chittka: Chemical compounds of the foraging recruitment pheromone in bumblebees. In: Naturwissenschaften. 92, 2005, S. 371–374, doi:10.1007/s00114-005-0002-0.
- Ana-Maria Costa-Leonardo, Ives Haifig: Termite Communication During Different Behavioral Activities. In: Biocommunication of Animals. Springer Netherlands, 2014, S. 161–190.
- Shelley E. R. Hoover, Christopher I. Keeling, Mark L. Winston, Keith N. Slessor: The effect of queen pheromones on worker honey bee ovary development. In: Naturwissenschaften. 90, 2003, S. 477–480, doi:10.1007/s00114-003-0462-z.
- K. W. Wanner, A. S. Nichols, K. K. O. Walden, A. Brockmann, C. W. Luetje, H. M. Robertson: A honey bee odorant receptor for the queen substance 9-oxo-2-decenoic acid. In: Proceedings of the National Academy of Sciences. 104, 2007, S. 14383–14388, doi:10.1073/pnas.0705459104.
- N. Koeniger, H. J. Veith: Spezifität eines Brutpheromons und Bruterkennung bei der Honigbiene (Apis Mellifera L.). In: Apidologie. 15, 1984, S. 205–210, doi:10.1051/apido:19840208.
- I. Leoncini, Y. Le Conte, G. Costagliola, E. Plettner, A. L. Toth, M. Wang, Z. Huang, J.-M. Becard, D. Crauser, K. N. Slessor, G. E. Robinson: Regulation of behavioral maturation by a primer pheromone produced by adult worker honey bees. In: Proceedings of the National Academy of Sciences. 101, 2004, S. 17559–17564, doi:10.1073/pnas.0407652101.
- Matthew R. Tarver, Eric A. Schmelz, Michael E. Scharf: Soldier caste influences on candidate primer pheromone levels and juvenile hormone-dependent caste differentiation in workers of the termite Reticulitermes flavipes. In: Journal of Insect Physiology. 57, 2011, S. 771–777, doi:10.1016/j.jinsphys.2011.02.015.
- Edward Vargo: Primer pheromones in ants. In: Pheromone communication in social insects: ants, wasps, bees, and termites. 1998, S. 293–313.
- Edward Howe Forbush, Charles Henry Fernald: Gypsy moth. Porthetria dispar (Linn.).: A Report of the Work of Destroying the Insect in the Commonwealth of Massachusetts together with an Account of its History and Habits both in Massachusetts and in Europe. Wright & Potter, Boston 1896, ISBN 0-405-10393-X, S. 339.
- Charles Walter Collins, Samuel Frederick Potts: Attractants for the flying gipsy moths as an aid in locating new infestations. In: Technical Bulletin No. 336. U.S. Department of Agriculture (1932).
- H. Z. Levinson: Possibilities of using insectistatics and pheromones in pest control. In: Die Naturwissenschaften. 62, 1975, S. 272–282, doi:10.1007/BF00608953.
- D. Klimetzek, L. Schlenstedt: Waldschutz gegen Borkenkäfer: Der Beitrag von Duftstoffmeteorologie und Populationsdynamik. In: Anzeiger für Schädlingskunde Pflanzenschutz Umweltschutz. 64, 1991, S. 121–128, doi:10.1007/BF01906002.
- H. Wigger: Ökologische Bewertung von Räuber-Beifängen in Borkenkäfer-Lockstoffallen. In: Anzeiger für Schädlingskunde Pflanzenschutz Umweltschutz. 66, 1993, S. 68–72, doi:10.1007/BF01903073.
- Franz Pühringer, Nils Ryrholm: Pheromonanflug europäischer Glasflügler (Lepidoptera, Sesiidae). In: Mitteilungen der Entomologischen Arbeitsgemeinschaft Salzkammergut. 31. Dezember 2000, S. 65–72 (zobodat.at [PDF; abgerufen am 12. Mai 2014]).
- Borkenkäfer – Vorbeugung und Bekämpfung. (PDF; 1,5 MB) (Nicht mehr online verfügbar.) In: Waldverband.at. Juni 2009, archiviert vom Original am 14. Juli 2014; abgerufen am 12. April 2018.
- Donald L. Kachigamba, Sunday Ekesi, Mary W. Ndung’u, Linus M. Gitonga, Peter E. A. Teal, Baldwyn Torto: Evidence for Potential of Managing Some African Fruit Fly Species (Diptera: Tephritidae) Using the Mango Fruit Fly Host-Marking Pheromone. In: Journal of Economic Entomology. 105, 2012, S. 2068–2075, doi:10.1603/EC12183.
- B. I. Katsoyannos, E. F. Boller: Second field application of oviposition-deterring pheromone of the European cherry fruit fly, Rhagoletis cerasi L. (Diptera: Tephritidae). In: Zeitschrift für Angewandte Entomologie. 89, 1980, S. 278–281, doi:10.1111/j.1439-0418.1980.tb03467.x.
- Stephen C. Welter, Carolyn Pickel, Jocelyn Millar, Frances Cave, Robert A. Van Steenwyk, John Dunley: Pheromone mating disruption offers selective management options for key pests. In: California Agriculture. 59, 2005, S. 16–22, doi:10.3733/ca.v059n01p16.
- R. G. Danka, J. L. Williams, T. E. Rinderer: A bait station for survey and detection of honey bees. In: Apidologie. 21, 1990, S. 287–292, doi:10.1051/apido:19900403.
- G. A. Rowell, M. E. Makela, J. D. Villa, J. H. Matis, J. M. Labougle, O. R. Taylor: Invasive dynamics of africanized honeybees in North America. In: Naturwissenschaften. 79, 1992, S. 281–283, doi:10.1007/BF01175399.
- A. Polettini, O. Crippa, A. Ravagli, A. Saragoni: A fatal case of poisoning with cantharidin. In: Forensic Science International. 56, 1992, S. 37–43, doi:10.1016/0379-0738(92)90144-L.
- Athula B. Attygalle, E. David Morgan: Pheromones in Nanogram Quantities: Structure Determination by Combined Microchemical and Gas Chromatographic Methods [New Analytical Methods (35)]. In: Angewandte Chemie. International Edition in English. 27, 1988, S. 460–478, doi:10.1002/anie.198804601.
- Stefan Schulz: Auf der Spur der chemischen Sprache der Tiere. In: Nachrichten aus der Chemie. 59, 2011, S. 704–709, doi:10.1002/nadc.201173368.
- Dietrich Schneider: Elektrophysiologische Untersuchungen von Chemo- und Mechanorezeptoren der Antenne des Seidenspinners Bombyx mori L. In: Zeitschrift für Vergleichende Physiologie. 40, 1957, S. 8–41, doi:10.1007/BF00298148.
- Dietrich Schneider: Insect pheromone research: some history and 45 years of personal recollections. (PDF) Abgerufen am 23. April 2014.