Dendrit (Biologie)
Dendriten (altgr. δένδρον dendron ‚Baum‘ bzw. dendrites ‚zum Baum gehörend‘) heißen in der Biologie Zellfortsätze von Nervenzellen, die aus dem Zellkörper hervorgehen und vorwiegend der Reizaufnahme dienen.[1][2]
| Übergeordnet |
| Neuron / Zelle |
| Untergeordnet |
| Dornenfortsatz Terminus Schaft Verzweigung dendr. Zytoplasma |
| Gene Ontology |
|---|
| QuickGO |
| Aufbau einer Nervenzelle |
|---|
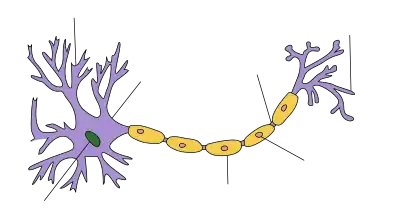
Eine Nervenzelle besteht typischerweise aus drei Anteilen: dem Zellkörper, Soma oder Perikaryon genannt, und Zellfortsätzen, die Dendriten einerseits und der Neurit – in Gliahülle das Axon – andererseits.[3] Es gibt auch spezialisierte Neuronen, die kein Axon haben (z. B. die Amakrinzellen der Netzhaut) oder die keine Dendriten besitzen (z. B. die Stäbchen und Zapfen der Netzhaut) oder solche, bei denen der Zellkörper nicht mehr zwischen Dendritenstamm und Axon liegt und die Fortsätze so ineinander übergehen (pseudounipolare Nervenzellen wie die sensiblen Spinalganglienzellen).
Dendriten als Anteile einer Zelle sind nicht mit dendritischen Zellen des Immunsystems zu verwechseln.
Dendritenwachstum
Trotz der Bedeutung der Dendriten für die Neuronen ist erst wenig darüber bekannt, wie Dendriten wachsen und sich in vivo orientieren und verzweigen. Eine Vermutung über diese Bildungsprozesse stellt die synaptotrophe Hypothese dar, nach der die Bildung von Synapsen eine besondere Rolle für das Wachstum von Dendriten spielt.
Ansonsten wird das Dendritenwachstum ähnlich dem von Neuriten erklärt, und zwar über sogenannte Wachstumskegel (englisch growth cones). Danach besitzen sowohl Neuriten wie Dendriten an ihren Spitzen kegelförmige oder kolbenartige Auftreibungen, die in einer forschenden Wechselwirkung mit der näheren Umgebung stehen und über Fortgang oder Fortsetzung – Ausmaß und Richtung des Auswachsens von Fortsätzen – entscheiden und das weitere Verhalten der Neuronen wesentlich bestimmen. Zellkulturtechniken mit Zeitrafferaufnahmen können eine deutliche Anschauung davon geben, wie Nervenzellfortsätze ihre Umgebung absuchend aussprossen.[4] Im Körper gibt es verschiedene Signale und unterschiedliche Prozesswege, über die Beginn, Richtung und Geschwindigkeit sowie Pausen des Dendritenwachstums geregelt werden können.
Der Großteil des Wachstums von Dendriten im menschlichen Gehirn läuft während der späten embryonalen und frühkindlichen Hirnentwicklung ab. In dieser Phase wachsen aus den 100 Milliarden Nervenzellen unseres Gehirns Dendriten mit einer Gesamtlänge von vielen hundert Kilometern aus.[5] Als ein für das Wachstum des Zellskeletts während der Dendritenentwicklung wichtiges Protein wird das Enzym Nedd4-1 angesehen, das für ein normales Dendritenwachstum unverzichtbar sein soll.[5]
Neuriten bzw. Axone und Dendriten unterscheiden sich in ihrem Wachstum und nach Wachstumsphasen. Zellulär betrachtet bedarf es grob gesprochen für das Fortsatzwachstum zunächst eines stabilisierten Stützskeletts aus Mikrotubuli, um die Wachstumsspitze vorzuschieben. Dann aber braucht es für das hin-und-her-spielende Wachstum in dieser Region – in labilem Gleichgewicht – Auf- und Abbauprozesse, mit denen einzelne Aktin-Moleküle (kugelförmiges, globuläres G-Aktin) sich zu Ketten (fadenförmiges, filamentäres F-Aktin) aneinanderreihen – und wieder zerfallen können. Bei instabilen Mikrotubuli und/oder stabilen Aktin-Filamenten ist kein Wachstum möglich. In frühen Entwicklungsphasen kann derart über In-/Stabilisation auf der einen Seite das Dendritenwachstum vorübergehend eingestellt werden, zugunsten des Längenwachstums des Neuriten auf der anderen. Prinzipiell gelten diese Zusammenhänge auch später noch, beispielsweise bei Regenerationsprozessen nach Läsionen.[6]
Anatomie von Dendriten
Form
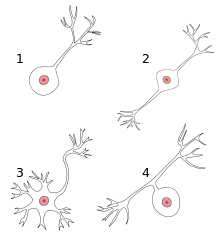
1 unipolare Nervenzelle
2 bipolare Nervenzelle
3 multipolare Nervenzelle
4 pseudounipolare Nervenzelle
Die Formen- und Funktionsvielfalt der Neuronen sind im Wesentlichen durch die unterschiedliche Ausprägung der Dendriten bestimmt. Die Abbildung zeigt die morphologische Unterscheidung von Nervenzellen, die u. a. danach getroffen wird, ob eine Nervenzelle keine, einen oder mehrere Dendriten besitzt. Manche Neuronen verfügen über regelrechte Dendritenbäume, bei anderen ist das Verhältnis Somaoberfläche zu Dendritenoberfläche ausgewogener. Schließlich gibt es auch Neuronen, die keine Dendriten haben.[7] Dieser morphologischen Einteilung folgend kann man sagen, dass Dendriten nur bei bipolaren Nervenzellen und multipolaren Nervenzellen vorkommen. Bei pseudounipolaren Nervenzellen hat das distale Ende des peripheren Fortsatzes typisch dendritischen Charakter.[3]
Zahl und Form der Dendriten tragen ganz wesentlich zur Vergrößerung der rezeptiven Oberfläche der Nervenzellen bei. Es wurde geschätzt, dass an den Dendriten einer einzelnen Purkinje-Zelle bis zu 200 000 Axone enden.[3] In der Regel sind die Dendriten baumartig verzweigte, verästelte Fortsätze des Perikaryons.
Zellbestandteile
Dem Aufbau nach steht der Dendrit dem Zellkörper näher als der Neurit. Dendriten und Perikaryon können in mancher Hinsicht sogar als funktionelle Einheit aufgefasst werden und werden auch als somatodendritisches Kompartiment bezeichnet.[8] Die Zusammensetzung des dendritischen Zytoplasmas entspricht im Wesentlichen der des Perikaryons.[3] Es ist daher unmöglich, eine scharfe Grenze zwischen den Teilen der Nervenzelle zu ziehen.[9]
Die Kenntnis von Zytoplasma, Organellen und Zytoskelett erlaubt einen fundierten Ansatz zur Unterscheidung der Fortsätze (Axon/Dendriten).
Folgende morphologische Besonderheiten finden sich:
- Im Gegensatz zum Axon sind Dendriten unmyelinisiert.
- In den größeren Stammdendriten finden sich ähnliche Organellen wie im Perikaryon. Besonders im breitbasigen Ursprung (perikaryonnah) lassen sich z. T. sogar noch Nissl-Schollen (raues endoplasmatisches Retikulum) finden.[8] Neben dem glatten und rauen endoplasmatischen Retikulum gibt es freie Ribosomen, Mikrofilamente (Aktin) und auch Bündel parallel verlaufender Mikrotubuli. Deren funktionelle Ausrichtung ist jedoch nicht gleichförmig (wie beim Axon), sondern ihre Polarität ist variabel, d. h. ihr Plus-Ende kann entweder zur Peripherie oder zum Perikaryon zeigen.[8]
Während Fibrillen und Nissl-Schollen noch lichtmikroskopisch zu erkennen sind, sind die anderen Bestandteile nur im Elektronenmikroskop sichtbar. - Mit jeder Aufzweigung wird der Durchmesser der Dendriten kleiner. In sehr dünnen Dendriten fehlen Mitochondrien.[10] Die Endabschnitte der Dendriten enthalten wenige Organellen, und auch das Zytoskelett ist nur noch gering ausgebildet.
- Im Vergleich zu den Axonen (die beim Menschen teilweise über 1 m lang sein können) sind Dendriten sehr klein und erreichen nur Längen von einigen hundert Mikrometern (µm). Neuriten, die in die Peripherie auswachsen, können allerdings eine Länge von 1 bis 1,20 m erreichen bei einem Durchmesser von nur 2–16 µm.[11]
Unterscheidungen von Dendriten
Es finden sich in der Literatur verschiedene Unterscheidungsmerkmale von Dendriten.
Betrachtet man Pyramidenzellen (eine recht große Nervenzelle), lassen sich zwei Arten von Dendriten unterscheiden: Apikaldendriten und Basaldendriten.[12] Beide entspringen an der Spitze der Pyramidenzellen, apikale Dendriten sind jedoch länger als basale Dendriten. Die apikalen Dendriten weisen in die dem Axon entgegengesetzte Richtung und erstrecken sich quer vertikal durch die Schichten der Großhirnrinde. Sowohl Apikal- als auch Basaldendriten besitzen Dornen.[13] Während es viele Basaldendriten gibt, steigt zur Kortexoberfläche nur ein langer, starker Apikaldendrit auf.[14]
Manchmal werden die Apikaldendriten noch unterschieden in distale und proximale Dendriten. Die distalen Apikaldendriten sind länger und projizieren in die dem Axon entgegengesetzte Richtung. Wegen ihrer Länge bilden sie nicht-lokale Synapsen, die weit entfernt von der Nervenzelle liegen. Proximale Apikaldendriten sind kürzer und empfangen Impulse von näher gelegenen Neuronen, etwa Interneuronen.[15]
Weiterhin kann man Dendriten danach unterscheiden, ob sie Dendritische Dornen besitzen oder nicht. Man spricht dementsprechend von glatten („smooth dendrites“) oder dornigen („spiny dendrites“) Dendriten.[15] Bei glatten Dendriten wird der Nervenimpuls direkt aufgenommen. Bei dornigen Dendriten nehmen sowohl Dendritenstamm als auch Dornen den Impuls auf.
In der Regel empfangen die dornigen Dendriten exzitatorische Signale, inhibitorische Synapsen finden sich dagegen eher auf glatten Dendriten(abschnitten).
Dendritische Dornen
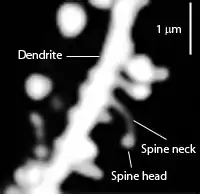
Die kleinen stachelartigen Fortsätze an den Oberflächen verzweigter Dendritenbäume werden dendritische Dornen (engl. spines, lat. spinula dendritica oder gemmula dendritica) oder Dornenfortsätze genannt. Hier sind oft die meisten der synaptischen Kontakte lokalisiert.[16]
In der Regel erhält ein Dornenfortsatz Input von genau einer Synapse eines Axons. Diese feinen Fortsätze (auf einem Dendriten als Nervenzellfortsatz) unterstützen die afferente Übertragung elektrischer Signale hin zum Zellkörper des Neurons. Die Dornen können verschiedene Gestalt annehmen, deutlich ausgebildete haben oft einen knolligen Kopf und einen dünnen Hals, der den Kopf mit dem Dendritenstamm verbindet. Die Dendriten eines einzelnen Neurons können hunderte oder tausende von Dornen tragen. Zusätzlich zu ihrer Funktion als postsynaptische Region (Postsynapse) – einige mit einem Dornenapparat als Calciumspeicher – und der ermöglichten Verstärkbarkeit der synaptischen Übertragung (Langzeit-Potenzierung LTP) können Dendriten außerdem dazu dienen, die mögliche Zahl an Kontakten zwischen Neuronen zu erhöhen.
Dornenfortsätze stellen eine Art Subkompartimentierung der Dendritenmembran dar. Die dadurch mögliche Feinabstimmung des einzelnen Dornenfortsatzes durch sein besonderes Ionenmilieu bzw. sein spezifisches cAMP-Niveau kann für die Selektivität bzw. Speicherung von Informationen wichtig sein.[17]
Funktionen
Den größten Anteil an der Versorgung der Neurone übernehmen die Gliazellen, eine Art Stützgewebe. Aber auch die Dendriten sind an der Ernährung der Nervenzelle beteiligt.[18] Ihre Hauptaufgabe ist jedoch das Empfangen von Reizen oder von Signalen zumeist von anderen Nervenzellen und das Weiterleiten der daraufhin gebildeten Impulse zum Perikaryon (Nervenzellkörper) hin (afferent oder zellulipetal) – im Gegensatz zum Neuriten bzw. dem Axon, über das Signale dieses Neurons am Axonhügel beginnend fort (efferent) und anderen Zellen zugeleitet werden.
Signalaufnahme
| Aufbau einer chemischen Synapse |
|---|
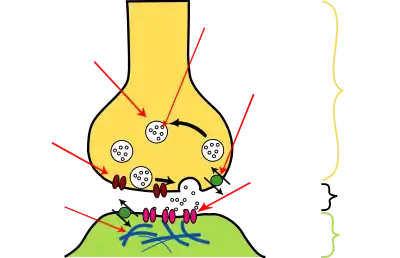
Eine Nervenzelle kann eine Sinneszelle sein – wie etwa Riechzellen (Geruchsrezeptoren) oder Sehzellen (Fotorezeptoren) – oder Signale von vorgeschalteten Zellen erhalten – beispielsweise von anderen Nervenzellen, indem Neurotransmitter an spezifische Rezeptoren in den postsynaptischen Membranregionen dieser Nervenzelle andocken. Zumeist liegen diese Postsynapsen nicht im Bereich von Axon, Axonhügel oder Soma (Körper) der Nervenzelle, sondern auf ihren Dendriten. Kontaktstellen zwischen Neuronen heißen interneuronale Synapsen, wobei mehrere Typen unterschieden werden (siehe auch Klassifikationen von Synapsen). Dendriten sind an den folgenden Typen beteiligt:
- Dendro-dendritische Synapsen: Sie verbinden verschiedene Dendriten miteinander.
Manche Dendriten zeigen präsynaptische Spezialisierungen, über die sie mit anderen (postsynaptischen) Dendriten in Kontakt treten und so dendrodendritische Synapsen bilden können. Als chemische Synapsen können diese mit präsynaptischen Vesikeln und postsynaptischen Membranregionen ausgebildet sein. Manchmal verfügen Synapsen auch als gap junctions weder über Vesikel noch über die sonst üblichen Membranverdichtungen und können Signale nicht nur in eine Richtung (unidirektional) übertragen, sondern bidirektional. Auch chemische dendrodendritische Synapsen mit beiderseits spiegelbildlich synaptischen Bläschen und Membrananlagerungen kommen vor als sogenannte reziproke Synapsen, bei denen in die eine Richtung ein erregender Transmitter (z. B. Glycin) und in die andere ein hemmender (z. B. GABA) ausgeschüttet wird.[19]
Beispiele für dendrodendritischen Synapsenkontakt in der Tierwelt sind bidirektionale Synapsen im stomatogastrischen Ganglion des Hummers, das die Mundhöhle mit dem Magen nerval verbindet, oder die Reichardt-Bewegungsdetektoren im Fliegenauge.[20] Beim Menschen kommen zum Beispiel reziproke Synapsen im Bulbus olfactorius der Riechbahn vor.[21] - Axodendritische Synapsen: Gewöhnlich enden Axone bzw. deren Abzweigungen (Axonkollateralen) innerhalb des Nervensystems als präsynaptische Endigung an einem Dendriten und bilden so axodendritische Synapsen.
- Axospinale dendritische Synapsen: In diesem Spezialfall axodendritischer Synapsen umgreift das Axon den Dornenfortsatz eines Dendriten.
Der an einer der vielen verschiedenen Synapsen einer Nervenzelle eingehende (afferente) Impuls verändert das Membranpotential in dieser Region (postsynaptisches Potential). Diese Potentialänderung breitet sich rasch über die benachbarten Membranbereiche aus, mit zunehmender Entfernung schwächer werdend, und kann entweder depolarisierend (EPSP) oder hyperpolarisierend (IPSP) sein. Durch hyperpolarisierte Regionen kann die Weiterleitung depolarisierender Potentiale aufgehoben werden. Laufen zu einem bestimmten Zeitpunkt hinreichend starke Depolarisationen am Axonhügel zusammen, sodass ein gewisser Schwellenwert überschritten wird – dann wird ein Aktionspotential ausgelöst, das Neuron ist erregt. Nahezu gleichzeitig einlaufende Reize können sich dabei in ihrer Wirkung addieren und am Axonhügel durch Summation ein Erregungspotential aufbauen. Generell gilt, je näher dem Axonhügel eine Synapse liegt, desto stärker ist ihr Einfluss auf die Erregung dieser Nervenzelle, die Bildung von Aktionspotentialen – denn je weiter sich postsynaptische Potentialänderungen (elektrotonisch) ausbreiten, umso mehr werden sie abgeschwächt. Untersuchungen zum Dendritenpotential wurden schon sehr früh angestellt.[22]
Einzelnachweise
- Dendrit – Definition beim Roche Lexikon für Medizin
- Dendriten – Definition im Kompaktlexikon der Biologie.
- L.C.U. Junqueira, José Carneirohofer: Histologie. Springer Berlin Heidelberg (15. September 2004). ISBN 978-3-540-21965-1. S. 109–112.
- 34.3.4.1 Exploratorische Wachstumskegel suchen dem Axon den besten Weg – Buchauszug bei zum.de.
- Wie Nervenzellen wachsen – Pressemitteilung vom Max Planck Institut, 16. Februar 2010.
- Die Wachstumsbremse im Rückenmark lösen – Pressemitteilung beim Deutschen Zentrum für Neurodegenerative Erkrankungen (DZNE), Bonn, 8. Februar 2012.
- Niels Birbaumer, Robert F. Schmidt: Biologische Psychologie. Springer; Auflage: 7., vollst. überarb. u. ergänzte Aufl. (21. Juli 2010). ISBN 978-3-540-95937-3. S. 23.
- Karl Zilles, Bernhard Tillmann: Anatomie. Springer Berlin Heidelberg; Auflage: 1. Aufl. (10. August 2010). ISBN 978-3-540-69481-6. S. 47.
- Werner Linß, Jochen Fanghänel: Histologie. Gruyter; Auflage: 1 (4. November 1998). ISBN 978-3-11-014032-3. S. 81.
- Theodor H. Schiebler, Horst-W. Korf: Anatomie: Histologie, Entwicklungsgeschichte, makroskopische und mikroskopische Anatomie, Topographie. Steinkopff; Auflage: 10., vollst. überarb. Aufl. (21. September 2007). ISBN 978-3-7985-1770-7. S. 72.
- Johannes W. Rohen: Funktionelle Anatomie des Nervensystems. Lehrbuch und Atlas. 5. Aufl., S. 61. Schattauer, Stuttgart 1994, ISBN 3-7945-1573-0.
- Clemens Kirschbaum: Biopsychologie von A bis Z. Springer Berlin Heidelberg; Auflage: 1 (Februar 2008). ISBN 978-3-540-39603-1. S. 20/34
- Nervensystem - Großhirn (Memento vom 24. Oktober 2013 im Internet Archive) – Artikel bei der Uni Freiburg
- Werner Kahle, Michael Frotscher: Taschenatlas Anatomie. Band 3. Thieme, Stuttgart; Auflage: 10., überarbeitete Auflage. (26. August 2009). ISBN 978-3-13-492210-3. S. 242.
- What Are the Different Types of Dendrites? – Artikel bei wisegeek.com.
- Michaela Hartmann, Maria Anna Pabst, Gottfried Dohr: Zytologie, Histologie und Mikroskopische Anatomie: Licht- und elektronenmikroskopischer Bildatlas. Facultas; Auflage: 5., überarbeitete Auflage. (Dezember 2010). ISBN 978-3-7089-0682-9. S. 51.
- Lüllmann-Rauch - Taschenlehrbuch der Histologie Kap. 9 Nervengewebe.
- Pschyrembel, 257. Aufl., 1994, S. 308; Roche Lexikon Medizin, 5. Aufl. 2003, S. 406.
- Roger Eckert, David Randall, Warren Burggren, Kathleen French; et alii: Tierphysiologie, 4. A, Thieme Verlag, 2002, ISBN 3-13-664004-7, S.256.
- Katharina Munk: Taschenlehrbuch Biologie: Zoologie: Thieme, Stuttgart; Auflage: 1 (10. November 2010). ISBN 978-3-13-144841-5. S. 461.
- Wissenschaftliche Zeitschrift der Karl-Marx-Universität: Mathematisch-Naturwissenschaftliche Reihe, Band 33, 1984, S. 479.
- Über die Beziehungen zwischen Dendritenpotential und Gleichspannung an der Hirnrinde – Artikel von HEINZ CASPERS (1959), beim Springer Verlag.
Literatur
- Oliver Arendt: Untersuchungen zur diffusiblen Mobilität kalziumbindender Proteine in Dendriten von Nervenzellen. Leipziger Universitätsvlg; Auflage: 1 (30. November 2009). ISBN 978-3-86583-393-8
- Arne Blichenberg: Dendritische Lokalisation neuronaler mRNAs: Charakterisierung cis-agierender Elemente in Transkripten des mikrotubulusassoziierten Proteins 2 und der Ca2+/Calmodulin-abhängigen Proteinkinase II. Der Andere Verlag; Auflage: 1. Aufl. (2000). ISBN 978-3-934366-98-5
- Jan Eschrich: Zur Signalausbreitung und Konvergenz im Dendritensystem am Beispiel der elektrosensorischen Afferenz des clusterbildenden Welses Schilbe mystis. 2003. ISBN 3-933508-21-5
- Greg Stuart, Nelson Spruston, Michael Hausser: Dendrites. Oxford University Press; Auflage: 2nd Revised edition (REV). (27. September 2007). ISBN 978-0-19-856656-4
- Rafael Yuste: Dendritic Spines. Mit Pr (24. September 2010). ISBN 978-0-262-01350-5
Weblinks
- Eintrag zu Dendrit im Flexikon, einem Wiki der Firma DocCheck
- Dendro-dendritic synapse – Artikel über dendrodendritische Synapsen bei scholarpedia.org