Ökologische Nische
Ökologische Nische bezeichnet die Gesamtheit der biotischen und abiotischen Umweltfaktoren, innerhalb derer eine Art selbst ökologische Funktionen ausüben und überleben kann.[1] Einige Autoren umschreiben die ökologische Nische von Tieren auch als deren „Beruf“ oder „Planstelle“[2] innerhalb der Lebensgemeinschaft der verschiedenen Organismen im jeweiligen Biotop.
Begriffsgeschichte
Die Bezeichnung Nische in ökologischem Zusammenhang wurde in einer Schrift des Naturforschers Joseph Grinnell 1917 erstmals erwähnt.[3][1] (Es gibt verstreute frühere Belege ohne Resonanz). Der Nischenbegriff Grinnells ist im Wesentlichen auf die Umwelt bezogen: Er beschreibt einen Bereich der Erdoberfläche mit seinen spezifischen ökologischen Standorteigenschaften, der für das Überleben der betrachteten Art förderlich ist. Unabhängig davon führte der englische Ökologe Charles Elton im Jahr 1927 den Begriff Nische in ähnlichem Sinn nochmals ein.[4] Elton legt größeren Wert auf die biologischen Interaktionen, sein Nischenbegriff entspricht eher der Rolle oder Funktion einer Art in ihrer Lebensgemeinschaft, vor allem auch ihre Beziehung zu Feinden und Nahrungsressourcen.[1]
Der Begriff in seiner heutigen Verwendung geht auf einen einflussreichen Artikel des Zoologen George Evelyn Hutchinson zurück,[5] der auf den Arbeiten Grinnells und Eltons aufbaut. Hutchinsons wichtigste Neuerung war, dass die Nische nicht als Eigenschaft der Umwelt angesehen wird – in die eine Art „passen“ oder „nicht passen“ kann, die ggf. auch „leer“ bleibt –, sondern als eine Eigenschaft der Art selbst.[6][1]
Das Konzept von Hutchinson
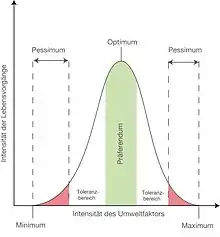
Hutchinsons Konzept geht von der bekannten Tatsache aus, dass eine Art nur leben kann, wenn die Umweltfaktoren sich innerhalb eines bestimmten Bereichs bewegen; hierzu zählen Temperatur, Feuchtigkeit, Bodenbeschaffenheit, Nahrungsangebot, Predationsdruck u. a. Der Wertebereich eines Faktors, z. B. der Temperatur, wird in graphischen Darstellungen üblicherweise in einem einfachen Koordinatensystem auf einer Skala auf der Abszisse („x-Achse“) eingetragen, der Grad der Eignung für den Organismus auf der Ordinate („y-Achse“). Wenn ein zweiter Faktor in derselben Grafik berücksichtigt werden soll, z. B. die Feuchtigkeit, muss ein Flächendiagramm erstellt werden mit der zweiten Dimension für den zweiten Faktor, d. h. für die Kombinationen aus Temperatur und Feuchtigkeit, in denen diese Art leben kann. (Ein Anwendungsbeispiel für die Nische des Wasserflohs Daphnia magna bietet Hooper.)[7] Ein dritter Faktor (z. B. pH-Wert des Bodens) definiert ein Volumen. Mit Hinzufügen weiterer Faktoren ergibt sich ein nicht mehr anschauliches, aber theoretisch mathematisch nachvollziehbares Gebilde: ein n-dimensionaler Hyperraum.
Dieser Hyperraum setzt sich also aus der Gesamtheit aller Umweltfaktoren wie Temperatur, Nahrung, Bodenfeuchte, Lebensraum usw. zusammen, die für die jeweilige ökologische Nische zutreffen, und formt (im mathematischen Sinne) einen sogenannten Nischenraum. Die einzelnen Faktoren lassen sich als Dimensionen graphisch darstellen. Ein Umweltfaktor stellt jeweils eine Dimension dar. Die ökologische Nische wird durch die Grenzen definiert, in denen eine Art leben und sich fortpflanzen kann, was sich als Bereich auf jedem Graph abbilden lässt. Es gibt innerhalb dieses Bereiches für jeden Faktor ein Optimum, bei dem die Art am besten gedeiht. Außerhalb des Optimumbereichs befindet sich zu beiden Seiten ein Toleranzbereich, in dem Individuen der betreffenden Art überleben können. In der Praxis gibt es jedoch aufgrund der Komplexität des Lebens sehr viele Dimensionen. Ein Diagramm einer realistischen mehrdimensionalen Nische kann man sich daher nur schwer vorstellen und nicht zeichnen. Der n-dimensionale Hyperraum der Umweltfaktoren kann daher aus Gründen der Anschaulichkeit in zwei- bis dreidimensionale Nischendiagramme zerlegt werden.

Moderne Anwendungen des Begriffs sind durch Ableitung und Abwandlung des Hutchinsonschen Konzepts entstanden. Besonders in die Synökologie fand der Nischenbegriff Eingang. Hier haben sich die mit dem Begriff zusammenhängenden Termini Nischenbreite, Ökologische Potenz, Nischenüberlappung und Einnischung bewährt.[8]
Nischenüberlappung bedeutet, dass sich zwei Arten die gleiche ökologische Nische teilen. Nischenüberlappung führt in der Regel zu Konkurrenz z. B. durch Verringerung von Ressourcen wie Raum oder Nahrungsangebot oder auch Licht und Wasser bei Pflanzen. Das hat ein Sinken der Überlebensfähigkeit, des Wachstums und der Fortpflanzungsrate der betroffenen Art(en) zur Folge. Ausnahmen sind nur Beziehungen von Arten mit positiven Auswirkungen, vor allem Mutualismus.
Der Begriff ökologische Nische wird gelegentlich missverstanden, da mit dem Begriff „Nische“ umgangssprachlich allgemein eine Räumlichkeit oder ein Ort verbunden wird. Die ökologische Nische ist aber keine räumliche Beschreibung, im Gegensatz zu den Begriffen Habitat (bzw. Standort) und Biotop, die einen physischen Ort bezeichnen. Tatsächlich ist die ökologische Nische ein funktioneller Begriff, der die „ökologische Rolle“ bezeichnet, welche die Art in dem betrachteten Ökosystem spielt. Er beschreibt also, welche biotischen und abiotischen Bedingungen, Umweltfaktoren und evolutionären Faktoren für das Leben bzw. Überleben dieser Art im Ökosystem von Bedeutung sind. Hieraus folgt, dass so definierte Nischen nicht „besetzt“ werden können. Sie werden vielmehr „gebildet“, und zwar durch Interaktion zwischen den Organismen einer Art mit ihrer Umwelt.

So ist beispielsweise die ökologische Nische des Igels unter der Schwankungsbreite der Temperaturen der gemäßigten Zone in seinem „Beruf“ als Vertilger von Schnecken und am Boden laufenden Insekten tätig zu sein, den der Hund nicht erbeuten kann, weil seine Stacheln ihn schützen, während sich zwischen den Stacheln die Flöhe vermehren, die auf den Hund als neuen Wirt überspringen. Der Maulwurf im gleichen Garten, der ebenfalls ein Insektenfresser ist, kann aufgrund anderer körperlicher Merkmale nicht die gleichen ökologischen Funktionen erfüllen.
Nischentheorie und Konkurrenz zwischen Arten
Die artbezogene Definition der ökologischen Nische kann naturgemäß für jede Art unabhängig erfolgen. Andere Arten gehen indirekt (als Umweltfaktoren, z. B. als Prädator oder als Nahrung) in die Nischendefinition ein, spielen aber direkt keine Rolle. Vergleicht man die so gefundenen ökologischen Nischen verschiedener Arten, können diese völlig getrennt voneinander sein, wenn die Arten keinen Bereich eines Lebensraums teilen. Häufiger werden sich die so gebildeten Nischen aber mehr oder weniger stark überlappen. Dies bedeutet, dass Teile des Nischenraums von beiden Arten genutzt werden können. Der Nischenraum einer Art in Gegenwart anderer Arten wird sich also von demjenigen ohne diese unterscheiden. Er kann größer sein (bei symbiontischen oder mutualistischen Beziehungen), häufiger wird er allerdings durch Konkurrenz oder andere Antagonismen kleiner sein.
Fundamentale und realisierte Nische
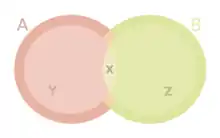
Man unterscheidet daher zwei grundlegende Konzeptionen einer ökologischen Nische:
- Als fundamentale Nische bzw. Fundamentalnische einer Art bezeichnet man den Teil eines Nischenraums, in dem diese Art alleine aufgrund ihrer physiologischen Potenz, also ihrer genetischen Variabilität und Reaktionsnorm und der damit verbundenen Anpassungsfähigkeit leben könnte. Praktisch ist dies nur unter Laborbedingungen feststellbar. Dies ist gewissermaßen die ökologische Gesamtbeschreibung der betreffenden Art.
- Als realisierte Nische bzw. Realnische bezeichnet man den Teil, der unter Berücksichtigung der konkreten aktuellen Standortfaktoren in einem bestimmten Ökosystem tatsächlich von der betreffenden Art belegt wird. Hierbei tritt jedes Individuum in Wechselbeziehung zu Artfremden oder Artgenossen und konkurriert mit ihnen um Ressourcen bzw. wird in sonstiger Weise in seinen Möglichkeiten beschränkt. Die realisierte Nische ist Teil der fundamentalen Nische. Das bedeutet, dass die realisierte Nische neben den biotischen Faktoren weiterhin auch von den abiotischen Faktoren abhängig ist.
Bedeutung des Konkurrenzfaktors
Da Konkurrenz zwischen den Arten nach Meinung sehr vieler Ökologen zu den wesentlichen Faktoren gehört, die natürliche Lebensgemeinschaften bestimmen, liegt ein wesentlicher Anwendungsbereich der Nischentheorie in der Betrachtung dieser Konkurrenz. Ausgangspunkt ist das Konkurrenzausschlussprinzip: Demnach können in einem Lebensraum zwei Arten mit gleichen Lebensansprüchen nicht miteinander koexistieren. Die konkurrenzüberlegene Art wird die unterlegene verdrängen. Gilt das Prinzip in natürlichen Lebensgemeinschaften, ist es erklärungsbedürftig, warum dort so viele Arten offensichtlich miteinander auskommen können, ohne dass die meisten von ihnen durch überlegene Konkurrenten verdrängt würden. Hutchinson selbst sprach z. B. für das Phytoplankton von Seen, das in einem scheinbar homogenen Lebensraum um eine Handvoll essenzieller Ressourcen konkurriert (im Wesentlichen: Licht, Phosphor, Stickstoff) und doch sehr artenreich ist, vom „Paradox des Planktons.“[9]
Übersetzt in die Sprache der Nischentheorie bedeutet Konkurrenzausschluss, dass sich die Nischen zweier Arten für zumindest eine dieser Arten vollständig überlappen (sie brauchen nicht identisch zu sein: Eine Nische kann von einer anderen vollständig eingeschlossen sein). Besitzt eine der Arten einen Anteil des Nischenraums, den der überlegene Konkurrent nicht besiedeln kann, hat sie dort ein Refugium. Damit ist zwar das Überleben der Art gesichert, aber ein gemeinsames Vorkommen wäre weiterhin unmöglich.
Innerhalb der Nischentheorie gibt es verschiedene Lösungsmöglichkeiten für dieses Problem.
- Räumliche Variabilität: Der Lebensraum besteht aus Teil-Lebensräumen, die sich in einem oder mehreren Umweltfaktoren unterscheiden. In jedem Teil-Lebensraum ist eine andere Art konkurrenzüberlegen.
- Zeitliche Variabilität: Die Lebensbedingungen ändern sich innerhalb der Saison, von Jahr zu Jahr oder langfristig und begünstigen dabei jeweils andere Arten. Den überlegenen Arten bleibt nicht genug Zeit, ihre Konkurrenten zu verdrängen, weil sich die Bedingungen bis dahin erneut geändert haben. Die Arten haben also eine „zeitliche Nische“.
- Trade-offs: Es gibt keine „Superarten“, die anderen Arten in allen Belangen überlegen wären. Vorteile in einem Bereich werden durch Nachteile in anderen „erkauft“ (engl. trade-off). So kann eine Art nicht gleichzeitig besonders große und besonders viele Samen haben. Eine Pflanze kann bei hohen Stickstoffgehalten des Bodens konkurrenzüberlegen sein, aber bei geringeren unterliegen.
- Modifikation durch biologische Interaktionen: Die Konkurrenzkraft einer Art kann durch andere Arten, vor allem durch Prädatoren, beeinflusst werden. Bei Gegenwart eines bestimmten Prädators kann eine sonst konkurrenzunterlegene Art z. B. indirekt gefördert werden, auch wenn die Art selbst zum Beutespektrum gehört, wenn der Konkurrent noch stärker unter ihm leidet. Die Beweidung durch den Prädator kann die Populationsdichte der Konkurrenten soweit reduzieren, dass der Faktor, um den normalerweise konkurriert wird, für alle Konkurrenten in ausreichendem Maß zur Verfügung steht.[10]
- Einnischung: Arten können der interspezifischen Konkurrenz ausweichen. Beispielsweise ist es beobachtet worden, dass sich Arten bei gemeinsamen Vorkommen in ihrem Körperbau stärker unterscheiden als bei alleinigem Auftreten („character displacement“). Ein berühmtes Beispiel sind verschiedene Darwinfinkenarten auf den Galapagosinseln.[11] Zur Erklärung für die Einnischung verschiedener Arten wurde eine „diffuse“ Konkurrenz durch verwandte Arten vorgeschlagen.[12]
Neutrale Theorien
Als Alternative zu den genannten Möglichkeiten bestreitet eine Gruppe von Ökologen die überragende Bedeutung der Konkurrenz. In diesem Falle stellen überlappende Nischen kein Problem mehr dar, das es zu lösen gelte. Nischen und Einnischung verlieren in diesen Theorien ihre zentrale Rolle in der Strukturierung von Lebensgemeinschaften. Die lokale Zusammensetzung wird von zufallsabhängigen („stochastischen“) Faktoren geprägt: v. a. Einwanderungs- und Aussterberaten, Kolonisationsgeschwindigkeit, zufällige Reihenfolgeänderungen der eintreffenden Arten (und in evolutionären Zeiträumen: Neuentstehung von Arten). Diese „neutrale Theorie“ ist besonders durch den Ökologen Stephen P. Hubbell ausgearbeitet worden.[13][14] Einen modernen Überblick dazu geben Beeravolu u. a.[15] Auch in der neutralen Theorie ist der Konkurrenzfaktor implizit berücksichtigt, weil der zur Verfügung stehende Raum für jedes Individuum begrenzt ist. Etablierung eines neuen Individuums ist nur in einer Lücke möglich, die beim Tod eines älteren frei wird. Eine solche „Lücken-Lotterie“ um Etablierungsnischen war schon früher zur Erklärung des Artenreichtums von Kalkmagerrasen vorgeschlagen worden. Der Ökologe P. J. Grubb hat dafür den Begriff der Regenerationsnische geprägt.[16]
Moderne Synthesen
Die neue Fassung der neutralen Theorie hat in der Ökologie eine heftige Kontroverse ausgelöst. Obwohl die Theorie in ihrer reinen Form von den meisten Ökologen abgelehnt wird, hat sie zu einer Modifikation der Nischentheorie beigetragen. In den meisten modernen Fassungen werden stochastische Elemente in die (im Kern völlig deterministische) Nischentheorie eingebracht. Mögliche Erklärungsmuster für Artenreichtum sind danach durch eine Kombination beider Faktoren gegeben.[17] Andere Möglichkeiten bieten z. B. multiple Gleichgewichte.[18] Eine moderne Übersicht über die Nischentheorie bietet z. B. Holt.[19]
Anpassung von Nischen
Die ökologische Einnischung von Tierarten ist nach neueren Erkenntnissen flexibler als in den bisherigen Theorien angenommen. Eine Studie zeigte 2011, dass sich die Jagdgewohnheiten tauchender Seevögel sowohl zwischen verschiedenen Arten als auch innerhalb einer Art stark unterscheiden. Die ökologischen Nischen sind dabei nicht starr festgelegt: Unterschiedliche Habitate, das Vermeiden von Konkurrenz mit Nachbarn oder das Ausweichen vor Fressfeinden führen auch innerhalb einer Art zu unterschiedlichem Verhalten. Die Wissenschaftler vom Max-Planck-Institut für Ornithologie in Radolfzell wählten Seevögel als Untersuchungsobjekt, da diese zur Brut ans Land gebunden sind und sich insbesondere zu dieser Zeit viele Tiere den Platz und die Nahrung teilen müssen. Da mehrere in ihren Ansprüchen ähnliche Arten gemeinsam auf einer Insel brüten, war die Frage, wie sie sich in ökologischen Nischen unterscheiden. Die Daten zeigten, dass sich die räumliche und zeitliche Verteilung der Vögel auch innerhalb einer Art sehr stark unterscheiden kann. Die Einnischung innerhalb einer Art durch Spezialisierungen mildert Konkurrenz. Die Tiere tauchen in geographisch weit auseinanderliegenden Meeresgebieten in unterschiedlicher Tiefe und Temperatur nach Nahrung.[20]
Anwendung der Nischentheorie zur Vorhersage von Artarealen
Eine zunehmende Verwendung findet die Nischentheorie in der biogeographischen Forschung. Hier werden bekannte Vorkommen einer Art mit gemessenen Umweltfaktoren (v. a. Klima) korreliert, um so aus der ökologischen Nische der Art ihr potenzielles Areal vorhersagen zu können, indem man die Ergebnisse auf Gebiete mit ähnlichen Umweltfaktoren überträgt, in denen die Art bisher noch nicht nachgewiesen war. Diese Modelle werden als Habitatmodellierung bezeichnet. Besonders verbreitet ist diese Vorgehensweise zur Vorhersage der Auswirkungen des Klimawandels. Hierzu wird, nach den vorliegenden Klimamodellen, ein Modell des Areals unter den veränderten Klimabedingungen konstruiert. Grundlage der Bearbeitung ist die Beobachtung, dass sich die ökologische Einnischung einer Art in kurzen Zeiträumen kaum verändert. Es ist also wahrscheinlicher, dass die Art durch Wanderungsbewegungen ihr Areal verschiebt, als dass sie sich an Ort und Stelle adaptiv an die neuen Bedingungen anpasst. Man spricht anschaulich von der „klimatischen Hülle“ (engl. climatic envelope) der Art.[21] Auf mögliche Gefahren und Fehlschlüsse mit dieser Methode machen einige amerikanische Ökologen aufmerksam, die damit ein Areal für den Yeti konstruieren können.[22]
Abweichende Verwendung des Begriffs Nische außerhalb der Ökologie
Der Begriff der ökologischen Nische hat außerhalb der Ökologie eine weite Verwendung gefunden. Die populären Konzepte, die damit umschrieben werden, unterscheiden sich allerdings stark von der wissenschaftlichen Begriffsverwendung. Eine ökologische Nische im allgemeinen Sprachgebrauch meint ein geschütztes Plätzchen außerhalb des rauen Windes der Konkurrenz, in der man etwa eine zwar beschränkte, aber halbwegs gesicherte Existenz führen kann. Der Politiker Günter Gaus charakterisierte so die Gesellschaft der früheren DDR als „Nischengesellschaft“.
Eine besondere Karriere hat der Begriff in der Ökonomie, insbesondere der Marktpsychologie durchlaufen, in die er von dem Psychologen Bernt Spiegel in einer bewussten Übertragung als „ökonomische Nische“ eingeführt wurde. Die gängigen Begriffe wie „Marktnische“ oder „Nischenprodukt“ gehen so auf die Übertragung eines ökologischen Konzepts in die Ökonomie zurück. Diese Ambivalenz wird von Wirtschaftsjournalisten vor allem im Zusammenhang mit sogenannten „Öko-Produkten“ am Leben gehalten, die Schlagzeilen dazu gern mit Wortspielen in der Art von „Biodiesel – raus aus der Nische“ einleiten.
Siehe auch
- Phytotelma für ein Kleinstgewässer als Mikrobiotop innerhalb einer lebenden Pflanze.
Literatur
- Andrew Cockburn: Evolutionsökologie. Gustav Fischer, Stuttgart 1995, ISBN 3-437-30775-4.
- B. D. Collier, G. W. Cox, A. W. Johnson, P. C. Miller: Dynamic Ecology. London 1974, ISBN 0-13-221309-5.
- Townsend, Harper, Begon: Ökologie. 1. Aufl., Springer-Verlag, Berlin/Heidelberg 2003, ISBN 3-540-00674-5.
- Katharina Munk (Hrsg.): Grundstudium Biologie. Spektrum Akad. Verl., Heidelberg 2000, ISBN 3-8274-0910-1.
Einzelnachweise
- A. Pocheville: The Ecological Niche: History and Recent Controversies. In: Thomas Heams u. a. (Hrsg.): Handbook of Evolutionary Thinking in the Sciences. Springer, Dordrecht 2015, ISBN 978-94-017-9014-7, S. 547–586 (Volltext).
- Werner J. Kloft: Ökologie der Tiere. Ulmer, UTB 729, Stuttgart 1978, S. 161 ff.
- J. Grinnell: The niche-relationships of the California thrasher. In: The Auk. 1017, 34, S. 427–433.
- C. S. Elton: Animal Ecology. Sidgwick and Jackson, London 1927.
- G. E. Hutchinson (1957): Concluding remarks. In: Cold Spring Harbor Symposium on Quantitative Biology. 22, 415–427.
- Im deutschen Sprachraum wird teilweise das Nischenkonzept von Klaus Günther oder das Konzept der ökologischen Lizenz nach Günther Osche verwendet, die sich in Details unterscheiden. Vgl.: Michael Schmitt: Ecological niche sensu Günther and ecological licence sensu Osche – two valuable but poorly appreciated explanatory concepts. In: Zoologische Beiträge N.F. 31, 1987, 49–60.
- Helen L. Hooper u. a: The ecological niche of Daphnia magna characterized using population growth rate. In: Ecology. 89(4), 2008, 1015–1022.
- Munk 2000, S. 14–24.
- G. E. Hutchinson: The paradox of the plankton. In: The American Naturalist. Band 95, Nr. 882, 1961, S. 137–145.
- Andrew Cockburn: Evolutionsökologie. Gustav Fischer, Stuttgart 1995, ISBN 3-437-30775-4, S. 12.
- Peter R. Grant, B. Rosemary Grant: Evolution of character displacement in Darwin’s finches. In: Science. 313, 224, 2006, 224–226.
- Eric R. Pianka: Niche Overlap and Diffuse Competition. Proceedings of the National Academy of Science 71(5), 1974, 2141–2145.
- S. P. Hubbell: A unified theory of biogeography and relative species abundance and its application to tropical rain forests and coral reefs. In: Coral Reefs. 1007, 16, S. 9–21.
- S. P. Hubbell: The Unified Neutral Theory of Biodiversity and Biogeography. Princeton University Press, Princeton, NJ 2001, S. 375 ff.
- Champak R. Beeravolu, Pierre Couteron, Raphaël Pélissier, Francois Munoz: Studying ecological communities from a neutral standpoint: A review of models’ structure and parameter estimation. In: Ecological Modelling. 220, 2009, 2603–2610.
- P. J. Grubb: The maintenance of species-richness in plant communities: the importance of the regeneration niche. In: Biological reviews. 1977, 52, 107–145.
- Z. B. David Tilman: Niche tradeoffs, neutrality, and community structure: A stochastic theory of resource competition, invasion, and community assembly. In: PNAS. 101 (30), 2004, 10854–10861.
- Z. B. Jonathan M. Chase: Stochastic community assembly causes higher biodiversity in more productive environments. Science 328, 2010, 1388–1391.
- Robert D. Holt: Bringing the Hutchinsonian niche into the 21st century: Ecological and evolutionary perspectives. In: PNAS. 106 (17) Suppl. 2, 2009, 19659–19665.
- J. F. Masello, R. Mundry, M. Poisbleau, L. Demongin, C. Voigt, M. Wikelski, P. Quillfeldt: Diving seabirds share foraging space and time within and among species. In: Ecosphere. 1:art19. doi:10.1890/ES10-00103.1.
- R. J. Hijmans, C. H. Graham: The ability of climate envelope models to predict the effect of climate change on species distributions. In: Global Change Biology. 12, 2006, 1–10.
- J. D. Lozier, P. Aniello, M. J. Hickerson: Predicting the distribution of Sasquatch in western North America: anything goes with ecological niche modelling. In: Journal of Biogeography. 2009, 1–5.