Populationsdynamik
Populationsdynamik ist die Veränderung der Größe, aber auch der räumlichen Verbreitung biologischer Populationen in kürzeren oder längeren Zeiträumen. Die Erforschung der Populationsdynamik bei einzelnen oder auch mehreren gekoppelten Populationen ist ein prominenter Gegenstand der Biologie, speziell der Ökologie und der Theoretischen Biologie. Die Populationsdynamik von Arten wird bestimmt durch multifaktorielle Wechselwirkungen sowohl innerhalb der Population als auch mit ihrer belebten und unbelebten Umwelt. Dies gilt für kurze Zeiträume ebenso wie für sehr lange Zeiträume.
Synökologische Aspekte
Natürliche, aber auch die meisten künstlichen Systeme, bestehen aus mehreren Arten, zwischen denen verschiedene Wechselwirkungen bestehen:
Randbedingungen des Populationswachstums
Man unterscheidet bei den Faktoren, welche die Dichte einer Population begrenzen, zwei Gruppen:
Dichteunabhängige Faktoren
Sie sind unabhängig von der Zahl der Individuen, die ein Biotop besiedeln.
- Wetter und Witterung: Das sich in Tages- und Wochenfrist ändernde, von Jahr zu Jahr leicht unterschiedliche klimatische Geschehen mit Faktoren wie Temperatur, Niederschlagsmenge, Wind, Sonneneinstrahlung etc.
- Katastrophen: Unvorhersehbare Ereignisse wie Vulkanausbrüche, verheerende Unwetter, Überschwemmungen, die zum Tod eines Teils der Population oder der ganzen Population (lokales Aussterben) führen können.
- Unspezifische Fressfeinde: Feinde, deren Beutespektrum normalerweise andere Lebewesen umfasst, und deren eigene Populationsgröße damit unabhängig von der zufällig erlegten Beute ist.
- Zwischenartliche (interspezifische Konkurrenz) Konkurrenz: Die Populationsentwicklungen von verschiedenen Tierarten im selben Biotop mit ähnlichen Ansprüchen an Nahrung, Reviere und andere Ressourcen können mehr oder weniger unabhängig voneinander sein, wenn sie unterschiedliche ökologische Nischen besetzen.
- Nicht ansteckende Krankheiten: Im Gegensatz zu Infektionskrankheiten, die sich in einer dichteren Population leichter ausbreiten, sind nicht ansteckende Krankheiten statistisch auftretende Zufallsereignisse, deren relative Häufigkeit mit ansteigender Dichte nicht zunimmt.
- Pestizide: Der Einsatz von Pestiziden (in der Landwirtschaft) führt bei bestimmten Arten, gegen die das Pestizid eingesetzt wird, zum Tod eines Teils der Population oder sogar der gesamten Population, je nach Menge und Intensität der eingesetzten chemischen Substanzen.
- Verhaltensvermittelte Faktoren: Reaktionen von Beutetieren und/oder Beutegreifern auf Umweltreize mit trophischen Kaskaden[1]
Dichteabhängige Faktoren
Die Stärke ihrer Auswirkungen ist von der augenblicklichen Populationsdichte abhängig.
- Intraspezifische Konkurrenz: Die Konkurrenz zwischen Individuen einer Population um Ressourcen wie Nahrung, Lebensraum etc. Diese hängt von den artspezifischen Bedürfnissen ab. So benötigen Individuen einiger Tierarten große Reviere, andere leben in Sozialverbänden (z. B. Herden oder Staaten) auf engerem Raum beisammen.
- Sozialer Stress (Gedrängefaktor): Das Zusammenleben von Tieren verursacht Stress durch Begegnungen und Aggressionen. Mit zunehmender Dichte wird der Stress größer, bis bei manchen Tierarten Verhaltensänderungen, Unfruchtbarkeit oder gar der Tod eintritt.
- Fressfeinde: Tiere, die als Beute für Fressfeinde dienen, nehmen damit auch Einfluss auf deren Populationsdichte. Nimmt die Zahl der Beutetiere zu, so können auch die Fressfeinde entsprechend mehr Junge großziehen, die dann wiederum den Feinddruck auf die Beute erhöhen. (Siehe Räuber-Beute-Beziehung)
- Ansteckende Krankheiten (Infektionskrankheiten): Überall wo Individuen in großer Enge leben, steigt auch die Gefahr, dass sich eine ansteckende Krankheit durch Übertragung der Erreger rasch ausbreitet und so zur Epidemie wird.
- Parasiten: Bei großer Populationsdichte können sie sich, wie auch Infektionskrankheiten, schneller ausbreiten.
Grundformen von Populationsdynamiken
Ein-Spezies-Modelle
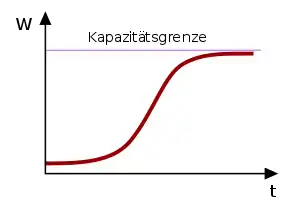
(Sigmoid-Kurve)
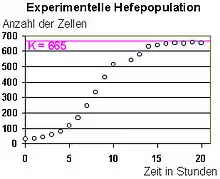
(K = Kapazitätsgrenze)
Eine Form des dichteabhängigen Wachstums wird durch die logistische Gleichung beschrieben.
Logistische Gleichung und Einflussfaktoren
Einflussfaktoren sind
- die Geburtenrate
- die Sterberate
- der begrenzende Kapazitätsfaktor
- die reproduktive Selbstbeschränkung
In der Natur wird die Kapazitätsgrenze durch folgende Faktoren beeinflusst:
- dichteunabhängige Faktoren, z. B.
- Klima
- dichteabhängige Faktoren, z. B.
- Ressourcen, wie Nahrung, Raum
- Versteckmöglichkeiten
- Gedrängefaktor (Sozialer Stress)
- Auswanderung
- innerartliche Konkurrenz (intraspezifische Konkurrenz)
Siehe auch: Gompertz’ verfeinerte Form des logistischen Modells und die mathematische Herleitung der logistischen Funktion.
Typische Phasen der Populationsentwicklung
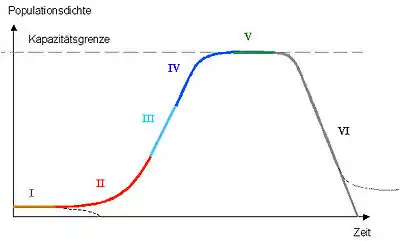
Anmerkung: In einer realen Population müssen nicht alle Phasen auftreten. In der Grafik zeigt die Abzweigung nach unten bei Phase I beispielhaft an, dass die Population schon in Phase I absterben kann. Die Abzweigung nach rechts in Phase VI zeigt an, dass ein Rückgang der Population nicht zum Aussterben führen muss.
Phase I: Lag-Phase (engl. to lag – verlangsamen), Anlaufphase, Nullwachstum auf niedrigem Niveau
Die Populationsgröße ist weit entfernt von der Kapazitätsgrenze. Die Geburtenrate ist ungefähr gleich der Sterberate. Es herrscht kein Mangel an Ressourcen, dichteabhängige Faktoren spielen praktisch keine Rolle, es herrscht keine Konkurrenz. Die Geburtenrate ist niedrig, da bei dieser geringen Populationsdichte innerartliche Begegnungen zufallsgesteuert sind. Die dichteabhängige Mortalität durch Fressfeinde spielt nur eine geringe Rolle, da für die wenigen Individuen genügend Verstecke im Lebensraum vorhanden sind. Auch Infektionskrankheiten wirken sich auf Grund der seltenen innerartliche Begegnungen nur wenig aus.
- Gründerpopulation: Kleine Populationen entstehen durch Auswanderung eines Teils der Population (Beispiel Lemminge) oder durch Katastrophen. Besiedelt eine kleine Populationen einen neuen Lebensraum, kann einige Zeit vergehen, bis die neuen Nahrungsquellen von der Population durch Umstellen der Ernährungsbedingungen optimal genutzt werden.
- Substratanpassung: Bei Mikroorganismen, die verschiedene Substrate verwerten können, dauert es meist einige Zeit (Minuten bis Stunden), bis sie sich auf veränderte Substratangebote umstellen (zum Beispiel Umstellung von anaeroben auf aeroben Stoffwechsel bei Hefezellen). In dieser Umstellungsphase ist kaum Wachstum zu beobachten. Dies gilt allgemein, wenn sich durch verbesserte Lebensbedingungen die Kapazitätsgrenze deutlich erhöht.
Schwankungen der Populationsdichte (Oszillationen) können zum Beispiel genetisch bedingt sein, Fluktuationen zufallsbedingt. Wenn dabei eine bestimmte Populationsdichte unterschritten wird, kann die Population aussterben. Beim Überschreiten eines gewissen Wertes kann die Population in die exponentielle Phase eintreten.
Phase II bis IV: Phase des positiven Wachstums
Die Geburtenrate ist höher als die Mortalitätsrate.
- Phase II: exponentielles Wachstum (Log-Phase). Es findet zunächst exponentielles Wachstum statt, da die Geburtenrate schneller ansteigt als die Mortalitätsrate. Die Ressourcen sind so reichlich vorhanden, dass innerartliche Konkurrenz keine Rolle spielt. Die Population der Fressfeinde, falls vorhanden, ist zunächst so gering bzw. Verstecke sind so zahlreich vorhanden, dass die Mortalitätsrate gering bleibt. Bei sehr günstigen Lebensbedingungen kann es durch das exponentielle Populationswachstum zu einer Massenvermehrung kommen. Mit zunehmender Populationsgröße nimmt aber auch die Sterberate zu.
- Phase III: lineares Wachstum. Steigen Geburten- und Sterberate gleich schnell an, nimmt die Population linear zu.
- Phase IV: verzögertes Wachstum. Mit zunehmender Annäherung an die Kapazitätsgrenze spielen innerartliche und evtl. zwischenartliche Konkurrenz eine immer größere Rolle, so dass der Anstieg der Geburtenrate abgebremst wird und die Sterberate weiter zunimmt. Falls es sich um Beuteorganismen handelt, nimmt mit zunehmender Beutedichte auch die Populationsdichte ihrer Fressfeinde zu, oder die Beutegreifer spezialisieren sich zunehmend auf diese Beute. Das weitere Wachstum wird abgebremst.
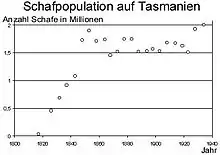
Phase V: Stationäre Phase
Die hohe Geburtenrate wird durch eine hohe dichteabhängige Mortalitätsrate (innerartliche Konkurrenz, Stress, Epidemien etc.) ausgeglichen. Es liegt maximale Besetzung des Lebensraums vor, die Ressourcen werden optimal genutzt, ohne sie zu erschöpfen (siehe Tragfähigkeit von Lebensräumen). Die Populationsdichte schwankt um den Wert K, die Kapazitätsgrenze. Dichteunabhängige Einflüsse (zum Beispiel Jahreszeiten) führen zu einem oszillierenden oder (bei ungünstigen Habitaten mit stark schwankenden Umwelteinflüssen) fluktuierenden Verlauf. Je größer eine Population ist, desto stabiler ist diese stationäre Phase. Ein Überhang erhöht die Sterberate und/oder senkt die Geburtenrate kurzfristig. Er kann durch Auswandern verringert werden (Beispiel: Lemminge).
Phase VI: Absterbephase
Die Geburtenrate ist jetzt geringer als die Sterberate. Bei kleinen Populationen können Zufallsschwankungen zum Aussterben führen. Eine Absenkung der Kapazitätsgrenze, zum Beispiel durch Umweltveränderungen oder Einwanderung neuer Fressfeinde, kann die Einstellung eines neuen Gleichgewichts auf niedrigerem Niveau bewirken.
Multispeziesmodelle
Ein relativ einfacher Fall ergibt sich durch Interaktionen zweier Spezies, die zueinander in einer Räuber-Beute-Beziehung stehen. Dieser Fall wird klassischerweise durch die Lotka-Volterra-Regeln beschrieben.
Komplexere Zusammenhänge ergeben sich durch Interaktion mehrerer Spezies. Es gibt Ansätze, solche Zusammenhänge mit Hilfe mathematischer Modelle zu beschreiben und zu simulieren.
Population als System
Im Folgenden soll auf Grund systemtheoretischer Überlegungen eine mathematische Modellierung entwickelt werden. Es wird zunächst davon ausgegangen, dass die Ressourcen für die Population unbegrenzt vorhanden sind. In Abweichung von manchen Literaturstellen werden Geburtenrate (birthrate) und Sterberate (mortality rate) als systeminterne Steuergrößen angesehen. Eine Populationsänderung zum Beispiel durch Geburt wird also nicht als Folge eines Zuflusses von außen aufgefasst. (Dies träfe eher auf Einwanderung zu). Wechselwirkungen mit der Umwelt in Form von Zu- oder Abwanderungen werden ebenfalls nicht berücksichtigt.
- Die Geburtenrate ist die positive Steuergröße: Je größer die Geburtenrate ist, umso größer wird die Population.
- Die Sterberate ist die negative Steuergröße: Je größer die Sterberate ist, umso kleiner wird die Population.


Allgemein werden derartige Systeme durch Partielle Differentialgleichungen wie z. B. der Fisher-Gleichung beschrieben. Im Folgenden werden Spezialfälle dieser Gleichung behandelt.
Einfluss der Geburtenrate
Es wird nur der Einfluss der Geburtenrate auf die Änderungsgeschwindigkeit der Populationsgröße N betrachtet:
(1)

Daraus ergibt sich (durch Integration) unter der Voraussetzung, dass die Geburtenrate konstant ist () für die Populationsgröße zu jedem beliebigen Zeitpunkt t mit der Ausgangsgröße N(t=0) = N0:

(2)

Dies ergibt ein positives lineares Wachstum, dessen Wachstumsgeschwindigkeit nur von der Geburtenrate abhängt: Je höher die Geburtenrate ist, desto rascher erfolgt das Wachstum.
Dieses System kann als Modell für Insektenstaaten (Bienen, Ameisen, Termiten) oder andere Tierpopulationen (Wolfsrudel) gelten, bei welchen nur ein Weibchen Junge zur Welt bringt, allerdings unter Vernachlässigung der Sterberate. Dabei sollte aber nicht vergessen werden, dass die Modellierung zu einem kontinuierlichen Wachstum führt, das aber in der Natur in diskreten Schritten erfolgt. Auch dürfte die Geburtenrate nur zeitweise unter Idealbedingungen konstant sein.
Einfluss der Sterberate
Für die Sterberate ergibt sich:
(3)

mit = c2 = const. > 0.
(4)

Damit ergibt sich ein negatives lineares Wachstum.
Die Extinktionszeit, die Zeit also, wann die Population ausgestorben ist, lässt sich mit berechnen.

In der Natur gibt es hierfür keine Entsprechung, da hier die Sterberate stets von der Populationsgröße abhängt. Eine regelmäßige Entnahme von Tieren zum Beispiel aus einer Schlachtvieh-Herde würde schon der Veränderung in einem offenen System entsprechen.
Einfluss von Geburten- und Sterberate
Werden Geburten- und Sterberate gleichzeitig berücksichtigt, ergibt sich die zeitliche Veränderung der Population aus:
(5)

Einsetzen der Gleichungen (2) und (4) und anschließende Integration ergeben:
(6)

Zwar liegt jetzt immer noch lineares Wachstum vor, ob aber die Population zu- oder abnimmt oder stagniert, hängt von der Größe der Geburten- und Sterberate ab:
- c1>c2: positives Wachstum
- c1<c2: negatives Wachstum mit
- c1=c2: Nullwachstum (Stagnation)

Population als isoliertes System mit Rückwirkung
In der Natur hängen Geburten- und Sterberate von der Populationsdichte und von der Kapazität des Ökosystems ab.
Unabhängige Geburtenrate und abhängige Sterberate
Im Folgenden soll die Geburtenrate weiterhin von der Populationsgröße unabhängig sein, während die Sterberate von der Populationsdichte abhängt:
(Je größer die Populationsdichte ist, desto größer ist die Sterberate). Aus dieser Proportionalität ergibt sich mit Hilfe des Proportionalitätsfaktors c2 = const. > 0 die Gleichung:

(7)

Für die Änderungsgeschwindigkeit der Populationsgröße auf Grund der Sterberate allein ergibt sich damit:
(8)

Daraus ergibt sich durch Integration:
(9)

Die Einbeziehung der konstanten, von der Populationsdichte unabhängigen Geburtenrate ergibt:
(10)

Durch Integration ergibt sich die Gleichung
(11)

Dieses System kann in einen von N0 verschiedenen Gleichgewichtszustand übergehen. Gleichgewicht herrscht dann, wenn genau so viele geboren werden wie auch wieder sterben, wenn also , oder auch wenn die Änderungsrate der Populationsgröße ist.


Aus Gleichung (10) ergibt sich damit für die Populationsgröße im Gleichgewicht (equilibrium):
(12)

Wenn befindet sich das System von Anfang im Gleichgewicht und damit im stationären Zustand. Trotz Zuwachs durch Geburt und Verlust durch Tod ändert sich die Populationsdichte nicht, es liegt Nullwachstum vor. Andernfalls wird (mathematisch) der Gleichgewichtszustand erst mit erreicht. Der Zeitpunkt , ab dem man praktisch von Gleichgewicht sprechen kann, ließe sich mit der Festlegung berechnen, dass ein im Vergleich zu minimaler Unterschied von N (zum Beispiel 0,01 %) bereits als Gleichgewichtssituation betrachtet wird.




Halbwertszeit der Gleichgewichtskurve:
(13)

In Anlehnung an die Enzymkinetik ließe sich eine Zeit berechnen, die der Michaeliskonstante entspricht. t½Neq wäre der Zeitpunkt, an dem die Population die Hälfte der Gleichgewichtsgröße erreicht hat:
(14)

Geburten- und Sterberate von Populationsgröße abhängig
Tragen in Populationen alle Weibchen zur Geburtenrate bei, ist auch die Geburtenrate von der Populationsdichte abhängig:
(15)

(Je größer die Populationsdichte, desto größer die Geburtenrate.) Aus dieser Proportionalität ergibt sich mit Hilfe des Proportionalitätsfaktors c1 = const. > 0 die folgende Gleichung:
(16)

Für die Änderungsgeschwindigkeit der Populationsgröße auf Grund der Geburtenrate ergibt sich damit:
(17)

Daraus ergibt sich durch Integration:
(18)

Damit ergibt sich für die Änderungsgeschwindigkeit der Populationsgröße unter Einbeziehung von Geburten- und Sterberate:
(19)

Daraus ergibt sich durch Integration:
(20)

Fallunterscheidungen:
- c1 = c2: Nullwachstum (stationär) bei N0
- c1 > c2: positives exponentielles Wachstum (beschleunigte Zunahme)
- c1 < c2: negatives exponentielles Wachstum (verzögerte Abnahme)
Dieses System hat keinen von N0 verschiedenen Gleichgewichtszustand.
Bei einer realen Population hängen allerdings Geburten- und Sterberate nicht nur von der Populationsdichte, sondern auch vom Abstand der Population von der Kapazitätsgrenze, das ist die im System maximal mögliche Populationsgröße, ab: Je näher eine Population an der Kapazitätsgrenze ist (je kleiner also die Differenz K-N), desto niedriger wird die Geburtenrate und desto höher wird die Sterberate.
Vereinfacht werden diese Beziehungen durch das Modell des Logistischen Wachstums (Verhulst) beschrieben:
Mathematische Modellierung
Grundgleichung (nach Pierre-François Verhulst):
(21)

Durch Integration ergibt sich:
(22)

Fallunterscheidung:
- N<K: N(t) ≈ ert, exponentielles (positiv beschleunigtes) Wachstum, wenn die Populationsgröße noch sehr weit weg von der Kapazitätsgrenze entfernt ist.
- N=K: N(t) = K, Nullwachstum (Stagnation), wenn die Populationsgröße der Kapazitätsgrenze entspricht.
- N>K: negatives Wachstum, wenn die Populationsdichte über der Kapazitätsgrenze liegt.
Eine recht gute Übereinstimmung des Modells mit den Beobachtungen ergeben sich für Organismen, die sich durch Zweiteilung vermehren (Bakterien, Hefen und andere eukaryotische Einzeller) sowie höhere Organismen, die eine niedrige Vermehrungsrate pro Generation haben und deren Generationen sich überschneiden.
Nicht beschrieben werden Fluktuationen und Oszillationen der Populationsdichte um die Kapazitätsgrenze sowie Abnahme der Population in der Nähe der Kapazitätsgrenze.
Eine Erweiterung der Formel des logistischen Wachstums um Zeitverzögerungen ergibt im Modell auch periodische Schwankungen:
(23)

- = Reproduktionsverzögerung, zum Beispiel Tragzeit
- = Reaktionszeitverzögerung der Population zum Beispiel beim Überschreiten der Kapazitätsgrenze


Modelle für interagierende Populationen
Räuber-Beute-System
Räuber-Beute-System nach Lotka-Volterra-Gleichung:
(24.1)

(24.2)

N(t): Anzahl der Beutetiere zum Zeitpunkt t
P(t): Anzahl der Räuber zum Zeitpunkt t,
a, b, c, d > 0: Koeffizienten
Wettbewerbsmodell
(25.1)

(25.2)

r(i): lineare Geburtenraten
K(i): Kapazitäten, limitiert durch Ressourcen
b(12), b(21): Wettbewerbseffekte von N2 auf N1, bzw. von N1 auf N2


Fluktuierende Populationsgrößen
Während die theoretischen Modelle zur Interpretation tatsächlich beobachteter Fluktuationen beitragen, so können sie doch diese Fluktuationen nicht immer zweifelsfrei erklären. Mehr oder minder regelmäßige Fluktuationen werden auch als Oszillationen bezeichnet.
Beispiele für Fluktuationen:
- Das bekannteste Beispiel stellen die Lemminge mit einer Fluktuationsrate von 3 bis 5 Jahren, aber eine eindeutige Erklärung steht noch aus.
- Für den Polarfuchs in Skandinavien wird gelegentlich eine Fluktuationrate seiner Populationsgröße von 3, an anderer Stelle von 7 bis 11 und an wieder anderer von 22 bis 25 Jahren angegeben, hier muss das Räuber-Beute-System gleich zweimal angewendet werden: auf ihn als Räuber und auf ihn als Beute.[2]
- Fluktuationen werden auch in europäischen Wildtierbeständen sehr deutlich wahrgenommen, so insbesondere bei Wildschweinen.[3] Im Hochtaunuskreis galten die Jahre 2000 bis 2003 als sehr wildschweinreich, die Jahre 2004 und 2005 als sehr wildschweinarm und 2008 wiederum als außerordentlich wildschweinreich. Die örtliche Presse informiert ausgiebig über Wildschweinschäden in den einen Jahren und über Mangel an Wildschweinfleisch in den anderen. Hier gilt womöglich das Räuber-Beute-System, wobei der Mensch die Rolle des Räubers einnimmt, aber weitere Faktoren spielen ebenfalls bedeutsame Rollen, darunter die Schweinepest und die wegen der Schweinepest verstärkte Bejagung, durch die die Übertragungsrate verringert wurde.
Siehe auch
Literatur
- Josef Hofbauer, Karl Sigmund: Evolutionstheorie und dynamische Systeme. Mathematische Aspekte der Selektion. Parey Verlag, Berlin, Hamburg 1984, ISBN 978-3-489-61834-8.
- James D. Murray: Mathematical Biology. Band I (3. Auflage), Springer-Verlag, 2002.
- Eine kurze Geschichte der mathematischen Populationsdynamik, 2021.
Einzelnachweise
- Paws without claws? Ecological effects of large carnivores in anthropogenic landscapes
- Literaturübersicht, Abschnitt 2.1.5 Populationsdynamik, Abwanderungsverhalten und Populationsstruktur (S. 9)
- Lydia Bauer: Wildschwein und Reh in den Bezirken ... seit 1950: Abschuss- und Bestandsentwicklung, mögliche Einflussfaktoren, Wildschweinschäden in der Landwirtschaft Diplomarbeit, Wien 2006 (PDF; 7,4 MB)