Massenaussterben
Ein Massenaussterben, auch Faunenschnitt oder Faunenwechsel genannt, ist ein in geologisch kurzen Zeitabschnitten von einigen tausend bis mehreren hunderttausend Jahren stattfindendes großes Artensterben, das sich vom normalen sogenannten Hintergrundaussterben ebenso deutlich abhebt wie von Massensterben: außergewöhnlichen, drastischen Verlusten von Individuen einer oder vieler Populationen einer Art innerhalb kurzer Zeit.
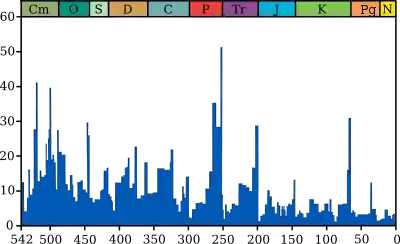
Im Verlauf der jüngeren Erdgeschichte fanden im Abstand von mehreren 10 Millionen Jahren immer wieder mehr oder weniger stark ausgeprägte Faunenwechsel statt. Diese anhand des Fossilberichtes nachvollziehbaren Ereignisse dienten historisch als Grundlage für die Ziehung der Grenzen zwischen den Epochen des Phanerozoikums in der Geologischen Zeitskala (siehe auch Evolutionsgeschichte). Ab den 1970er Jahren wurde anhand geologischer, paläontologischer und paläoklimatologischer Befunde zunehmend nachgewiesen, dass diese Faunenumschwünge auf drastische Erhöhungen der Aussterberaten zurückzuführen sind, die in einem relativ schmalen geologischen Zeitfenster auftraten.[1]
Die in den vergangenen Jahrzehnten erzielten Fortschritte bei den radiometrischen Datierungs- und Nachweisverfahren führten zu einer erheblichen Zunahme der Messgenauigkeit. Dadurch wurde es möglich, verschiedene Massenaussterben zeitlich genauer einzugrenzen, relativ umfassend zu beschreiben und vorher unbekannte biologische und ökologische Krisen im Laufe der Erdgeschichte zu dokumentieren. In der Wissenschaft besteht kein eindeutiger Konsens bei der Definition eines Massenaussterbens. Einige Publikationen verwenden den Begriff nur bei einem Artenverlust[2] von 75 Prozent oder darüber, andererseits wird häufig jeder stärkere Einschnitt in die biologische Vielfalt als Massenaussterben bezeichnet.[3][4]
Verschiedene Expertengremien gehen davon aus, dass das sechste Massenaussterben in der Geschichte des Lebens bereits begonnen hat. Laut dem im Mai 2019 publizierten Globalen Bericht des Weltbiodiversitätsrats der UN-Organisation IPBES könnten eine Million Tier- und Pflanzenarten innerhalb der nächsten Jahrzehnte vom Aussterben bedroht sein. Ein direkter Vergleich dieses Massenaussterbens mit früheren erdgeschichtlichen Krisenzeiten ist insofern problematisch, da gegenwärtig anthropogene Faktoren dominieren. Der IPBES-Bericht nennt als Gründe für das aktuelle Aussterben – in der Reihenfolge ihres Ausmaßes – den Verlust von Lebensraum, Veränderungen in der Landnutzung, Jagd und Wilderei, den Klimawandel, Umweltgifte sowie das Auftreten von Neobiota.
Forschungsgeschichtlicher Hintergrund

Aus geowissenschaftlicher Sicht ist der 6. Juni 1980 ein historisches Datum. Die an diesem Tag veröffentlichte Ausgabe der Fachzeitschrift Science enthielt als Hauptbeitrag einen Artikel mit dem Titel Extraterrestrial Cause for the Cretaceous-Tertiary Extinction (deutsch: Außerirdische Ursache für das Kreide-Tertiär-Aussterben). Als Autoren fungierten der Physiker und Nobelpreisträger Luis Walter Alvarez, sein Sohn, der Geologe Walter Alvarez, sowie die auf dem Gebiet der Kernchemie tätigen Teammitglieder Frank Asaro und Helen W. Michel.[5] Nach den Worten des US-amerikanischen Paläontologen David M. Raup schlug die Alvarez-Studie in der Wissenschaft „wie eine Bombe ein“.[6] Die auf dem Nachweis einer Iridium-Anomalie basierende Hypothese eines Asteroiden-Einschlags an der Kreide-Tertiär-Grenze (heute Kreide-Paläogen-Grenze), der unter anderem die Auslöschung der Dinosaurier bewirkt haben sollte, löste langwierige kontroverse Diskussionen auf Konferenzen und in der Fachliteratur aus. Ein Teil der Ablehnung resultierte aus der Weigerung vieler Forscher, ein Szenario zu akzeptieren, das alle Merkmale eines längst überwunden geglaubten Katastrophismus aufwies.
Dass an den Grenzen beziehungsweise Übergängen der geochronologischen Perioden fast immer ein umfassender Faunenwechsel stattgefunden hatte, war seit dem frühen 19. Jahrhundert bekannt. Erklärt wurden diese biologischen Krisen – in Einklang mit dem Prinzip eines allmählichen Wandels (Aktualismus) – bis weit in das 20. Jahrhundert zumeist mit der Annahme geologischer Prozesse wie Meeresspiegelschwankungen und Kontinentalverschiebungen und damit verknüpften Klima- und Umweltveränderungen über Zeiträume von Jahrmillionen. Zwar äußerten bekannte Wissenschaftler wie der US-amerikanische Nobelpreisträger Harold C. Urey oder der deutsche Paläontologe Otto Heinrich Schindewolf bereits in den 1950er Jahren die Vermutung, dass manche Aussterbe-Ereignisse mit einem Kometeneinschlag beziehungsweise mit den Strahlungsausbrüchen einer erdnahen Supernova in Verbindung stehen könnten, doch ihre Überlegungen fanden in der Fachwelt keine Resonanz.
Hingegen führte die von Alvarez et al. postulierte Katastrophe an der Kreide-Paläogen-Grenze zu einem allmählichen Paradigmenwechsel im Hinblick auf erdgeschichtliche Umbruchzeiten sowie zu einer zunehmend intensiveren Analyse potenzieller Massenaussterben unter Einbeziehung von Forschungsgebieten wie der Geochemie, Mineralogie, Paläontologie, Paläoklimatologie, Sedimentologie, (Event-)Stratigraphie, Vulkanologie, Geophysik sowie des Paläomagnetismus. Als Folge der interdisziplinären Zusammenarbeit wurden multikausale Modelle für Massenaussterben entwickelt, die neben den herkömmlichen Erklärungsmustern auch zusätzliche Faktoren wie Anoxie (das Fehlen von Sauerstoff), Ozeanversauerung, rasche Konzentrationsschwankungen von Treibhausgasen oder den Einfluss von Magmatischen Großprovinzen berücksichtigten. Die Fachliteratur zu diesem Themenbereich hatte sich zwischen 1984 und 2004 annähernd verzehnfacht,[1] mit dem Ergebnis, dass Massenaussterben nicht zwangsläufig an langfristige geologische Abläufe gekoppelt sein müssen, sondern häufig einen katastrophischen und zeitlich eng begrenzten Verlauf genommen haben. Seit Beginn des 21. Jahrhunderts spricht zudem eine wachsende Zahl von Belegen für die Annahme, dass viele Biodiversitätskrisen mit gravierenden klimatischen Veränderungen und deren Folgen verknüpft waren. Auf dieser Basis entstand ein „Ranking“ der verheerendsten Massenaussterben während der letzten 541 Millionen Jahre.[7][4]
Zu den sogenannten „großen Fünf“ (auch Big Five) zählen im Einzelnen:
- das Ordovizische Massenaussterben vor 444 Mio. Jahren
- das Kellwasser-Ereignis im Oberdevon vor 372 Mio. Jahren
- das Ereignis an der Perm-Trias-Grenze vor 252 Mio. Jahren
- die Krisenzeit an der Trias-Jura-Grenze vor 201 Mio. Jahren
- das Massenaussterben an der Kreide-Paläogen-Grenze vor 66 Mio. Jahren
Bei den großen Fünf lag der jeweilige Artenschwund bei wahrscheinlich 70 bis 75 Prozent oder zum Teil darüber. Obwohl gelegentlich eine Revision dieser Gliederung oder eine Neubewertung zugunsten anderer Massenaussterben vorgeschlagen wird[7] (vor allem hinsichtlich des Kellwasser-Ereignisses),[8] sind die Big Five immer noch die inoffizielle Richtschnur bei der Darstellung der schwerwiegendsten biologischen Krisen während des Phanerozoikums. Dies kommt nicht nur in der entsprechenden Fachliteratur zum Ausdruck, sondern wird auch in populärwissenschaftlichen Büchern und Beiträgen häufig thematisiert.[9]
Wahrscheinliche Ursachen von Massenaussterben
Die Ursachen von Massenaussterben sind ein vieldiskutiertes Thema in den verschiedenen Disziplinen der Geowissenschaften. Als eigentliche Auslöser, die ultimaten Ursachen, gelten mittlerweile vor allem der Vulkanismus magmatischer Großprovinzen und Einschläge kosmischer Kleinkörper. Von diesen auslösenden Ereignissen wird versucht, über mitunter komplexe Kausalketten hin zu unmittelbaren, proximaten Ursachen Erklärungen für Aussterbemuster zu finden. Dabei spielen als unmittelbare Faktoren, die massenhaft Arten ausgelöscht haben könnten, rasche Klimaveränderungen, Ozeanversauerung und marine Anoxie – neben anderen – eine wichtige Rolle.[7]
Von großer Bedeutung scheint zudem die Umweltsituation zu sein, in die das auslösende Ereignis tritt. So können die Vegetation über ihren Einfluss auf Verwitterungs- und Erosionsprozesse sowie überhaupt Organismen über ihren Stoffwechsel auf die Zusammensetzung von Meeren und Atmosphäre einwirken. Die Position und Anordnung von Landmassen wird ebenfalls als wichtige Randbedingung für die Wirksamkeit ultimater Aussterbeursachen genannt (ihre Änderung mitunter auch als eigentlicher Auslöser). Einer jüngst entwickelten Hypothese zufolge war die ausgeprägte Kontinentalität des 138 Millionen km² umfassenden Superkontinents Pangaea ein Hauptgrund, warum zu dieser Zeit die Biosphäre besonders verwundbar gegenüber dem Ausstoß vulkanischer Gase und Asche war – es fehlten negative Rückkopplungen, die das Erdsystem stabilisiert hätten. Nach der Auflösung Pangaeas hätten die höhere Verwitterungsrate auf den fragmentierten Kontinenten und eine höhere Produktivität Kohlenstoff bindender Organismen der Atmosphäre vermehrt Kohlenstoffdioxid entzogen und so den Folgen des Vulkanismus und damit letztlich einem Massenaussterben entgegengewirkt.[7][10]
Vulkanismus
Vulkanische Eruptionen der Kategorie VEI-6 oder VEI-7 auf dem Vulkanexplosivitätsindex verursachen durch ihren Ausstoß von vulkanischer Asche einen mehr oder minder ausgeprägten Abkühlungseffekt über mehrere Jahre. Mit Hilfe von Höhenströmungen (Starkwindbänder) verteilen sich die freigesetzten Aerosole in der Stratosphäre – der zweituntersten Schicht der Atmosphäre –, wo sie über Absorption, Streuung und Reflexion die transmittierte Sonneneinstrahlung in erheblichem Maße dämpfen.
Die zur höchsten Kategorie VEI-8 zählenden Supervulkane können aufgrund ihrer Auswurfmenge von über 1.000 km³ an Lava, Asche und Aerosolen (Tephra) Jahrzehnte dauernde Veränderungen der globalen Temperatur hervorrufen und infolge der in der Atmosphäre verteilten Partikel einen vulkanischen Winter auslösen. Alle bekannten Supervulkane hinterließen nach einem Ausbruch, bedingt durch die Größe ihrer Magmakammer, keine Vulkankegel, sondern riesige Calderen. Durch den Ausstoß von pyroklastischem Material wurde in einem Umkreis von mindestens 100 km jedes Leben vernichtet, und die mit vulkanischer Asche bedeckte Fläche umfasste wahrscheinlich Millionen km². Ereignisse dieser Größenordnung hatten Massensterben in den betroffenen Regionen mit einer Abnahme der Biodiversität zur Folge. Für die jüngere Erdgeschichte konnten bisher über 40 derartige Katastrophen eindeutig nachgewiesen werden,[11] darunter der La-Garita-Ausbruch im frühen Oligozän vor etwa 38 bis 36 Millionen Jahren (= 38 bis 36 mya), die bisher letzte VEI-8-Eruption des Yellowstone-Hotspots vor 640.000 Jahren[12] sowie das Toba-Ereignis auf Sumatra vor 74.000 Jahren. Dauerhafte klimatische und ökologische Folgen durch Supervulkane sind jedoch nicht belegt.

Hingegen waren sogenannte Magmatische Großprovinzen (englisch Large Igneous Provinces) mehrfach Ursache für eine gravierende und relativ rasch auftretende globale Erwärmung und ein damit verbundenes Massenaussterben.[13] Dabei handelte es sich um den großvolumigen Austritt magmatischer Gesteine aus dem Erdmantel, überwiegend in Form von Flutbasalten, die sich im Verlauf von einigen Hunderttausend Jahren mitunter über eine Fläche von Millionen km² ergossen. In Abhängigkeit von Ausmaß und Dauer der vulkanischen Aktivitäten wurden erhebliche, klimawirksame Mengen an Kohlenstoffdioxid freigesetzt, in signifikantem Umfang auch Chlorwasserstoff, Fluor und Schwefeldioxid. Da die Kohlenstoffdioxid-Emissionen in diesen Fällen deutlich über dem Aerosol-Ausstoß lagen, waren Magmatische Großprovinzen nicht nur für eine weltweite Temperaturerhöhung verantwortlich, sondern setzten im Extremfall eine zusätzliche Erwärmungsspirale unter Mitwirkung des starken Treibhausgases Methan beziehungsweise Methanhydrat aus ozeanischen Lagerstätten in Gang.[14] Sehr wahrscheinlich stehen viele Massenaussterben der Erdgeschichte mit dem großflächigen Ausfluss von Flutbasalten und der anschließenden Destabilisierung mariner Biotope in direkter Verbindung. Zu den Magmatischen Großprovinzen, die in unterschiedlich starker Weise Umwelt und Biodiversität beeinflussten, gehören unter anderem der Sibirische Trapp (Perm-Trias-Grenze, 252 mya), der Dekkan-Trapp im heutigen Westindien (Kreide-Paläogen-Grenze, 66 mya)[15] sowie der nordamerikanische Columbia-Plateaubasalt (Mittleres Miozän, Hauptaktivität 16,7 bis 15,9 mya).[16]
Klimawandel
Ein Klimawandel auf globaler Ebene beruht auf einem Strahlungsantrieb, der das Erdklimasystem aus einem stabilen thermisch-radiativen Gleichgewicht in ein neues Gleichgewicht (Equilibrium) überführt. Der Strahlungsantrieb resultiert aus Änderungen der atmosphärischen Konzentrationen von Treibhausgasen wie Kohlenstoffdioxid (CO2), Methan (CH4) und Wasserdampf (H2O), aus der variierenden Sonneneinstrahlung aufgrund der sich über längere Zeiträume verändernden Erdbahnparameter (Milanković-Zyklen) sowie aus dem Rückstrahlvermögen (Albedo) der Erdoberfläche einschließlich der Ozeane. Da die Komponenten des Erdsystems eine komplexe Einheit bilden, ist ein Klimawandel auf monokausaler Basis praktisch ausgeschlossen, da sich jede Teilveränderung auf die Gesamtstruktur auswirkt. Dies geschieht überwiegend in Form positiver Feedbacks, das heißt, das System entfernt sich durch eine Vielzahl kumulierender Wechselwirkungen (Rückkopplungs-Effekte) immer weiter vom ursprünglichen Gleichgewicht.[17]
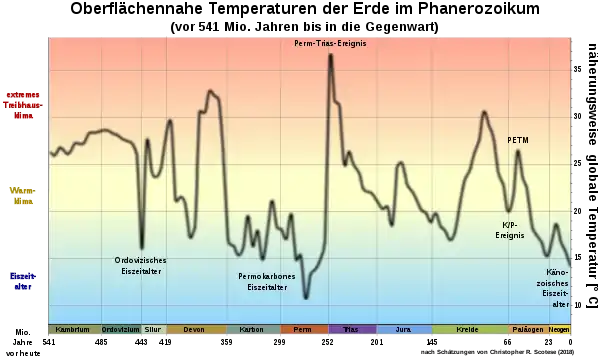
Gewöhnlich wird in der Wissenschaft zwischen den beiden Grundzuständen Warmklima und Kaltklima unterschieden. Um das gesamte klimatische Spektrum vollständig abzudecken, findet mitunter eine Differenzierung in Eishaus, Kühles Treibhaus, Warmes Treibhaus und Hitzehaus statt (Icehouse, Cool Greenhouse, Warm Greenhouse, Hothouse). Demnach besitzt jeder Klimazustand seine eigene Charakteristik, die sich von den übrigen signifikant unterscheidet.[18] Zusätzlich werden am Übergang von Eishaus zu Kühlem Treibhaus sowie von Warmem Treibhaus zu Hitzehaus mehrere Kipppunkte postuliert, die das Erdklimasystem in einen neuen und teilweise irreversiblen Zustand überführen (Kippelemente im Erdsystem).[19]
Im Hinblick auf die Massenaussterben der Erdgeschichte zählten abrupte Klimawechsel fast immer zu den Hauptursachen. Diese konnten sowohl innerhalb weniger Tage eintreten (wie an der Kreide-Paläogen-Grenze) als auch geologisch sehr kurze Zeiträume von einigen tausend oder zehntausend Jahren umfassen (wie bei den meisten Magmatischen Großprovinzen). Insgesamt vollzogen sich die jeweiligen Umweltveränderungen so rasch, dass viele Ökosysteme beim Übergang von einem Klimazustand zum nächsten ihre Anpassungsgrenzen erreichten und kollabierten. Schwerwiegende biologische Krisen korrelierten in den letzten 540 Millionen Jahren mehrmals mit einer Abkühlungsphase (mit einem weltweiten Temperaturrückgang von 4 bis 5 °C), häufiger jedoch mit starken Erwärmungen im Bereich von 5 bis 10 °C.[7] Im letzteren Fall trug ein Bündel von Nebenwirkungen (Vegetationsrückgang, Ausgasungen von Gift- und Schadstoffen, Sauerstoffdefizite, Versauerung der Ozeane etc.) dazu bei, die irdische Biosphäre weiter zu destabilisieren.
Ozeanische anoxische Ereignisse
Ozeanische anoxische Ereignisse (englisch Oceanic Anoxic Events, abgekürzt OAEs) beruhten auf einem Sauerstoffdefizit (unter 2 mg/l) vor allem in tropischen Flachwassermeeren. Davon ausgenommen war lediglich die jeweils oberste durchmischte Wasserschicht. Ein Indikator für das Auftreten anoxischer Ereignisse sind die in einem sauerstofffreien Milieu entstandenen marinen Schwarzschiefer-Horizonte, die sich aus Faulschlamm am Grund des Ozeans bildeten und die sowohl in paläozoischen als auch in mesozoischen Sedimentschichten häufig nachgewiesen wurden.
Ein signifikantes hypoxisches oder anoxisches Ereignis basiert im Normalfall auf folgenden Voraussetzungen:
- eine atmosphärische Kohlenstoffdioxid-Konzentration im Bereich von etwa 1.000 ppm
- ein weltweit subtropisches bis tropisches Klima bei gleichzeitiger Erwärmung der Ozeane mit entsprechend reduzierter Sauerstoffbindung (aus dem Ordovizium ist allerdings ein OAE während einer globalen Abkühlung dokumentiert)
- eine stark verminderte oder zum Stillstand gekommene Tiefenwasserzirkulation zwischen den Polarregionen und dem Äquator
- die Überdüngung der Meere mit festländischen Verwitterungsprodukten aufgrund der in einem Warmklima rasch verlaufenden Erosionsprozesse
Wahrscheinlich wurde ein großer Teil der anoxischen Ereignisse durch die lange anhaltenden CO2- und Schwefeldioxid-Ausgasungen von Magmatischen Großprovinzen mitverursacht oder zumindest deutlich verstärkt.[20][21] OAEs erstreckten sich meistens über einen Zeitraum von 300.000 bis 800.000 Jahren, konnten in Ausnahmefällen jedoch über 2 Millionen Jahre andauern oder wie während der Krisenzeiten im Oberdevon innerhalb weniger 100.000 Jahre mehrmals hintereinander auftreten. Regelmäßige Begleiterscheinungen waren eine das Meerwasser grün färbende Algenblüte sowie die Massenvermehrung sulfatreduzierender Bakterien. Durch die bakterielle Reduktion von Sulfat entstand hochgiftiger Schwefelwasserstoff (H2S), der sich nicht nur in den Meeren, sondern zum Teil auch in der Atmosphäre anreicherte, wobei in diesem Fall wahrscheinlich auch die Ozonschicht nachhaltig geschädigt wurde. Im Jahre 2008 kam man zu Erkenntnissen, dass es in der Erdgeschichte mehrere schwefelwasserstoff-induzierte Massenaussterben gegeben haben müsse,[22] in besonders folgenschwerer Ausprägung während der biologischen Krise an der Perm-Trias-Grenze.[23] Dieses Gas war damals nahezu allgegenwärtig und dessen letaler Wirkung fielen nach diesem Szenario neben Tausenden mariner Arten auch viele Vertreter der Landfauna zum Opfer.
Impakt-Ereignisse
Impakt-Ereignisse wie der Einschlag größerer Asteroiden oder Kometen können nicht nur die Biosphäre in erheblichem Umfang destabilisieren und Massenaussterben wie jenes an der Kreide-Paläogen-Grenze auslösen, sondern auch das Klima kurzfristig verändern (abrupt einsetzender Impaktwinter, in einigen Fällen mit anschließender starker Erwärmung durch freigesetzte Treibhausgase). Auf der Erde gibt es etwa 180 Einschlagkrater mit einer Größe von mehr als 5 bis 10 km, davon nur etwa zwei Dutzend in ozeanischen Sedimenten. Da Meeresböden durch den plattentektonischen Prozess der Subduktion ständig in die Tiefen des Erdmantels „abtauchen“, andererseits an den Spreizungszonen permanent neu gebildet werden, beträgt das Durchschnittsalter der ozeanischen Kruste etwa 80 Millionen Jahre. Somit besteht Grund zu der Annahme, dass viele Impakt-Ereignisse unbekannt bleiben werden und nicht mehr nachweisbar sind.
Die von Vater und Sohn Alvarez im Jahr 1980 veröffentlichte These eines Asteroideneinschlags an der Kreide-Paläogen-Grenze fand nach anfänglichem Widerstand rasche Akzeptanz und konnte sich innerhalb eines Jahrzehnts als wissenschaftlicher Mainstream etablieren. Als 1991 ein von Größe und Alter passender Krater auf der mexikanischen Halbinsel Yucatan entdeckt wurde,[24] war das nicht nur eine Bestätigung der Alvarez-These, sondern ließ auch die Anzahl der Impakt-Befürworter sprunghaft anwachsen. Laut Aussage des Geowissenschaftlers und Biologen Peter Ward hatten viele Forscher um die Jahrtausendwende „an der Vorstellung einen Narren gefressen“,[25] dass fast alle Massenaussterben von Himmelskörpern verursacht wurden, die mit der Erde kollidierten. Im Fokus stand dabei vor allem die Perm-Trias-Krise, die mit Strukturen wie dem hypothetischen Wilkesland-Krater oder der Bedout-Formation vor Australien in Verbindung stehen sollte. Inzwischen wurden diese Annahmen nicht bestätigt und zum Teil widerlegt.[26] Außer dem Einschlag am Kreide-Paläogen-Übergang konnte bisher kein weiterer „Global Killer“ im Verlauf des Phanerozoikums eindeutig identifiziert werden. Dies schließt jedoch nicht aus, dass künftige geologische Entdeckungen das bisherige Bild eventuell revidieren.[27]
Gammablitze

Seit dem ersten sicheren Beleg eines Gammablitzes (gamma-ray burst, abgekürzt GRB) im Jahr 1973 werden Gammablitz-Szenarien als Auslöser früherer Umweltkrisen diskutiert, wobei vor allem das ordovizische Massenaussterben im Fokus steht.[28] Für den Ursprung der Gammablitze existieren mehrere Erklärungsansätze, meist in Verbindung mit der Bildung eines Neutronensterns oder eines Schwarzen Lochs. Die Dauer eines Gammablitzes liegt häufig im Sekundenbereich, wobei in diesem Zeitraum erhebliche und stark gebündelte Energiemengen freigesetzt werden. Daher besteht zumindest theoretisch die Möglichkeit, dass ein die Erde direkt treffender Strahlungsausbruch das Potenzial besitzt, das irdische Leben nachhaltig zu schädigen. Unmittelbare biologische Auswirkungen wären jedoch begrenzt, da die Atmosphäre den Großteil der Strahlung absorbieren würde. Eine nicht zu unterschätzende Gefährdung wäre hingegen die Entstehung von Stickoxiden mit einer daraus resultierenden Zerstörung der Ozonschicht. Die Wahrscheinlichkeit für ein derartiges Ereignis gilt allerdings als äußerst gering.[29]
Supernovae
Gelegentlich wird für verschiedene Massenaussterben eine erdnahe Supernova als Ursache angenommen. Aus der Vergangenheit sind mehrere Supernovae-Signaturen in Form des Eisen-Isotops 60Fe bekannt. Dieses Isotop, das unter irdischen Bedingungen nicht entstehen kann, besitzt eine Halbwertszeit von 2,62 Millionen Jahren und wurde unter anderem in Sedimenten des frühen Quartärs (Gelasium-Stufe) vor 2,2 Millionen Jahren[30] sowie im Unteren Pleistozän (Calabrium) vor 1,5 Millionen Jahren entdeckt.[31] Allerdings stammen diese Belege aus der jüngeren Erdgeschichte. Für Gammablitze und Supernovae gilt gleichermaßen, dass ein Nachweis kosmischer Strahlungseinflüsse in paläozoischen und mesozoischen Gesteinsformationen mit dem gegenwärtig verwendeten Instrumentarium nicht möglich ist.[7]
Nemesis-Hypothese
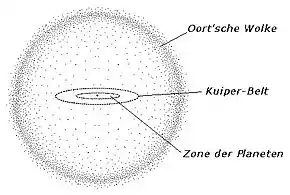
Die Nemesis-Hypothese (in Anlehnung an die griechische Göttin des „gerechten Zorns“) war die hauptsächlich in den 1980er Jahren populäre und vieldiskutierte Annahme, dass die Sonne von einem Braunen Zwerg oder einem leuchtschwachen Zwergstern in 1 bis 3 Lichtjahren Entfernung umkreist werde. In ihrer ursprünglichen Form beruhte die Hypothese auf dem Szenario, dass Nemesis während ihrer etwa 26 bis 27 Millionen Jahre währenden Umlaufperiode am sonnennächsten Punkt (Perihel) bei der Durchquerung einer mit Gestein, Staub und Eis durchsetzten kosmischen Region (Oortsche Wolke) aufgrund ihrer Gravitationswirkung eine Reihe von Kometen in das innere Sonnensystem und damit auch in Richtung Erde gelenkt habe. Dies führe zu einer zeitlich eng begrenzten Häufung von Impakt-Ereignissen und damit zu regelmäßig stattfindenden Massenaussterben. Die (scheinbare) Periodizität biologischer Krisen in der Erdgeschichte wurde zuerst von den Paläontologen David M. Raup und Jack Sepkoski anhand statistischer Analysen postuliert[32] und bildete eine der Grundlagen für die Ausarbeitung der Hypothese.[33]
Begründete Einwände gegen die Nemesis-Hypothese erfolgten sowohl von astronomischer und astrophysikalischer als auch von geowissenschaftlicher Seite. So existieren gegenwärtig keinerlei Hinweise auf einen Begleitstern der Sonne, und zum anderen wird der 26-Millionen-Jahre-Zyklus von Massensterben generell bezweifelt. Laut einer 2017 veröffentlichten Untersuchung geschah die in manchen Epochen nachgewiesene Zunahme von Asteroiden- oder Kometeneinschlägen auf der Erde rein zufällig und ohne ersichtliche Periodizität.[34]
Datierungs- und Analysemethoden
Die in den letzten Jahrzehnten erzielten Fortschritte bei der Untersuchung von Aussterbeereignissen beruhen zu einem großen Teil auf der Anwendung und Weiterentwicklung der Radiometrischen Datierung. Bei dieser Methode der Altersbestimmung werden die Zerfallsraten natürlich vorkommender Isotope und das Isotopenverhältnis gemessen. Bei Einsatz eines modernen Massenspektrometers sind gegenwärtig Probengewichte von einem Milliardstel Gramm oft ausreichend, um präzise Ergebnisse zu erzielen. In den Geowissenschaften existiert eine Vielzahl unterschiedlicher Datierungsmethoden, die drei folgenden Verfahren werden mit am häufigsten verwendet:
- Zirkonkristalle eignen sich durch ihre Hitzeresistenz und ihre stabile Gitterstruktur aufgrund der darin eingeschlossenen radioaktiven Nuklide (wie 235U, 238U oder 232Th) für geochronologische Datierungen über die gesamte Erdgeschichte. Die Fehlertoleranzen bei dieser Form der absoluten Altersbestimmung von Massenaussterben und geologischen Ereignissen liegen bei Anwendung dieses Verfahrens vom Silur bis zur Kreide bei nur wenigen 10.000 Jahren.
- Die 40Ar/39Ar-Datierung ist eine abgewandelte und genauere Methode der herkömmlichen Kalium-Argon-Datierung und wird seit einiger Zeit zur Altersbestimmung von Mineralien und Gesteinen häufig eingesetzt.
- Die Radiokarbonmethode, auch 14C-Datierung genannt, ist ein Verfahren zur Altersbestimmung organischer Substanzen. Aufgrund ihres eingeschränkten Anwendungsbereichs von 300 bis maximal 57.000 Jahren eignet sich die 14C-Datierung jedoch nur für die jüngste Erdgeschichte. Darüber hinaus führen die zunehmenden anthropogenen CO2-Emissionen gegenwärtig zu einer deutlichen Verringerung der 14C-Anteile in der Atmosphäre. Dieser Effekt wird künftige Datierungen auf dieser Basis voraussichtlich beträchtlich erschweren beziehungsweise verfälschen.[35]
Bei der Klimarekonstruktion vergangener Perioden kommen mehrere „Paläothermometer“ zum Einsatz:
- δ18O (Delta-O-18) beschreibt das Verhältnis der stabilen Sauerstoff-Isotope 18O/16O. Dieses vielfältig einsetzbare Messverfahren eignet sich für die Bestimmung von Niederschlagstemperaturen und dient zudem als Indikator von Prozessen der Isotopenfraktionierung wie der Methanogenese. In der Forschung werden 18O/16O-Daten als Temperaturproxy von fossilen Korallen und Foraminiferen sowie von Eisbohrkernen, Tropfsteinen und Süßwassersedimenten verwendet.[36]
- TEX86 (Tetraether-Index von 86 Kohlenstoffatomen) bezeichnet eine biogeochemische Methode zur Ermittlung der Meeresoberflächentemperatur früherer Epochen. Zu diesem Zweck wird die Zellmembran bestimmter mariner Einzeller analysiert.[37] Zeitlicher Anwendungsbereich: Jura, Kreide und gesamtes Känozoikum (etwa die letzten 200 Millionen Jahre).
Eines der wichtigsten Nachweisverfahren in der Geochemie und Paläoklimatologie basiert auf den Eigenschaften des Kohlenstoff-Isotops 13C:
- δ13C (Delta-C-13) ist das Maß für das Verhältnis der stabilen Kohlenstoff-Isotope 13C/12C zwischen einer Probe und einem definierten Standard. Die δ13C-Analyse erlaubt die Bestimmung der atmosphärischen Kohlenstoffdioxid-Konzentration in den verschiedenen erdgeschichtlichen Epochen und lässt Rückschlüsse auf Störungen oder Anomalien des Kohlenstoffzyklus zu. Die Freisetzung großer Mengen Methan wie während des Paläozän/Eozän-Temperaturmaximums hat ebenfalls deutliche Auswirkungen auf die globale δ13C-Signatur.[38]
Chronologie der Massenaussterben
Proterozoikum (vor 2.400 und vor 717/640 Millionen Jahren)
Im frühen Proterozoikum fand im Zuge der Großen Sauerstoffkatastrophe das wahrscheinlich umfassendste Massenaussterben des gesamten Präkambriums statt, bei dem fast alle anaeroben Lebensformen ausgelöscht wurden. Am Beginn dieser Ära wies die irdische Atmosphäre eine relativ hohe Methankonzentration, aber nur sehr geringe Anteile an freiem Sauerstoff auf. Mit großer Wahrscheinlichkeit existierten bereits vor mehr als 3 Milliarden Jahren Cyanobakterien, die die oxygen-phototrophe Photosynthese nutzten und dabei als „Abfallprodukt“ ihres Stoffwechsels in signifikantem Umfang Sauerstoff freisetzten, der jedoch bei der Oxidation von organischen Verbindungen, Schwefelwasserstoff und zweiwertigen Eisen-Ionen Fe2+ in dreiwertige Eisen-Ionen Fe3+ vollständig verbraucht wurde. Dieser Prozess war unmittelbar mit der Entstehung von Bändereisenerz verknüpft (englisch Banded Iron Formation), einem eisenhaltigen, im Präkambrium abgelagerten Sedimentgestein, das sich unter den heutigen geochemischen Bedingungen nicht mehr bilden könnte. Nach Abschluss dieser intensiven Oxidationsphase reicherte sich der Überschuss an freiem Sauerstoff sowohl in der Atmosphäre als auch in den Ozeanen an. Letzteres führte zum Massenaussterben obligat anaerober, also auf ein sauerstoffarmes Milieu angewiesener Organismen, die der Toxizität des Sauerstoffs fast vollzählig zum Opfer fielen. In der Atmosphäre bewirkte der Sauerstoff aufgrund seiner oxidativen Eigenschaften mit Hilfe der UV-Strahlung den Zusammenbruch der Methankonzentration.
Diese Zäsur zog einen gravierenden Klimawandel nach sich. Es gilt als sehr wahrscheinlich, dass die 300 Millionen Jahre dauernde Paläoproterozoische Vereisung (auch Huronische Eiszeit genannt) die unmittelbare Folge aus Methanreduzierung und Sauerstoffzunahme war.[39] Geologische Klimazeugen dieser Glazialphase sind unter anderem in Nordamerika, Skandinavien, Indien sowie im südlichen Afrika zu finden und deuten auf einen globalen Kälteeinbruch hin. Zusätzlich postulieren einige Studien mindestens ein Schneeball-Erde-Ereignis, das zu einer vollständigen Vergletscherung der Erde einschließlich der äquatorialen Zone und der Ozeane führte.[40] Mehrere globale Vereisungszyklen werden auch für die Sturtische Eiszeit (717 bis 660 mya) und für die Marinoische Eiszeit (640 bis 635 mya) im Neoproterozoikum angenommen.[41] Überwiegend wird davon ausgegangen, dass der natürliche Kohlenstoffzyklus dadurch fast zum Erliegen kam und die Biomasseproduktion in den Meeren auf einem sehr niedrigen Niveau verharrte. Wie die meisten Details des Schneeball-Szenarios werden jedoch auch diese Annahmen in der Wissenschaft kontrovers diskutiert.[42]
Ediacarium-Kambrium-Grenze (vor 541 Millionen Jahren)

Die in der Zeit vor etwa 580/570 bis 540 Millionen Jahre existierende Ediacara-Fauna bildete relativ artenarme, aber ausgeprägt heterogene Biotope vielzelliger Tiere (Metazoa), die, abgesehen von einigen Tiefseeformen, in benthischer (bodenbewohnender) Lebensweise die Kontinentalschelfe oder den Grund von Flachmeeren besiedelten. Da die Ediacara-Fauna stammesgeschichtlich ohne direkte Nachfolger blieb, ist ihre taxonomische Einordnung bis heute unklar beziehungsweise umstritten. Als gesichert kann gelten, dass die typischen Vertreter dieser Fauna nur eine geringe oder keine Mobilität besaßen (zum Beispiel als festsitzende, farnähnliche Organismen), sich von gelösten organischen Substanzen oder durch den Abbau mikrobieller Matten ernährten, offenbar über keine Skelettteile verfügten und am Übergang vom Ediacarium zum Kambrium vollständig verschwanden (wobei eine deutliche Reduzierung der Biodiversität bereits vorher stattfand).
Für diesen Faunenwechsel werden in der Wissenschaft zwei mögliche Hauptursachen diskutiert: einmal der gravierende Einfluss abiotischer umweltverändernder Faktoren (was ein „echtes“ Massenaussterben bedeuten würde), zum anderen die Verdrängung der Ediacara-Fauna durch die überlegene Konkurrenz „moderner“ kambrischer Arten.[43] Für die erste Annahme spricht die Tatsache, dass das späte Neoproterozoikum eine Epoche intensiver plattentektonischer Prozesse war. Fast gleichzeitig mit dem Auseinanderbrechen des hypothetisch postulierten Superkontinents Pannotia verschmolzen mehrere Kratone zum Großkontinent Gondwana, verbunden mit umfangreichen Auffaltungen wie der 8.000 km langen Gebirgskette des Transgondwanan Supermountain.[44] Daraus resultierten ein starker Vulkanismus, eine „Überdüngung“ der Ozeane mit festländischen Verwitterungsprodukten aufgrund einer extrem hohen Erosionsrate sowie wahrscheinlich signifikante Klima- und Meeresspiegelschwankungen. Nachgewiesen sind die Existenz von Schwarzschiefersedimenten und die Entstehung von Schwefelwasserstoff in den damaligen Weltmeeren.[45] Beides lässt auf eine Sauerstoffverknappung und auf die Bildung anoxischer Zonen schließen. Zudem konnte anhand der Isotopenanalyse δ13C eine globale Störung des organischen Kohlenstoffzyklus festgestellt werden.[46] Diesem Szenario wird entgegengehalten, dass vergleichbare geochemische und geotektonische Prozesse wie im Ediacarium-Kambrium-Grenzbereich schon vorher auftraten und dass die Auslöschung der Ediacara-Fauna im Wesentlichen durch die rasche Ausbreitung kambrischer Organismen erfolgte, entweder in direkter Form durch freischwimmende Fressfeinde oder indirekt durch wühlende und grabende Tiere (Bioturbation), die auf diese Weise – im Zuge eines eher graduellen Übergangs – die Existenzgrundlage der Ediacara-Arten zerstörten.
Übersicht paläozoischer und mesozoischer Biodiversitäts-Krisen
Mit dem Beginn des Phanerozoikums vor etwa 541 Mio. Jahren kam es zu einer deutlichen Zunahme der Artenvielfalt, auch andere als mikroskopisch kleine Fossilreste lassen sich seitdem nachweisen. In die ersten beiden Ären, das Paläozoikum (Erdaltertum) und das Mesozoikum (Erdmittelalter), fallen die größten Aussterbeereignisse.
| Bezeichnung/Epoche | Datierung | Ungefährer Artenschwund | Wahrscheinlicher Megavulkanismus | Temperaturanomalien1) | Anoxische Ereignisse |
|---|---|---|---|---|---|
| Mittleres Kambrium | 510 mya | 50 %? | Kalkarindji-Vulkanprovinz | Nicht quantifiziert | Ja |
| Oberes Ordovizium | 444 mya | Bis 85 % | – | Abkühlung auf eiszeitliches Klima (ca. −5 °C), Vergletscherungen auf der südlichen Hemisphäre | Ja |
| Kellwasser-Ereignis | 372 mya | 65 bis 75 % | Viluy-Trapp? | Kurzzeitige Erwärmungsspitzen bis +9 °C | Ja |
| Hangenberg-Ereignis | 359 mya | Bis 75 % | – | Rascher Wechsel von Warm- und Glazialphasen, gravierende Absenkung des Meeresspiegels | Ja |
| Capitanium | 260 mya | Um 60 % | Emeishan-Trapp | Wahrscheinlich keine ausgeprägten Klimaschwankungen | Ja (regional?) |
| Perm-Trias-Grenze | 252 mya | 75 % an Land, 95 % im Ozean | Sibirischer Trapp | Erwärmung um +10 °C, in der Unteren Trias (Smithium/Spathium) +6 °C | Ja |
| Trias-Jura-Grenze | 201 mya | Um 70 % | Zentralatlantische Magmatische Provinz | Erwärmung um +4 bis +6 °C | Ja |
| Kreide-Paläogen-Grenze | 66 mya | Bis 75 % | Dekkan-Trapp | Nach Impaktwinter kurzzeitige Erwärmung (+4 °C in den Meeren, +6 bis +8 °C auf dem Festland) | ? |
Kambrium (vor ca. 510, 502 und 485 Millionen Jahren)
Das Kambrium war eine Epoche mit zum Teil stark erhöhtem Vulkanismus, mit Durchschnittstemperaturen um 20 °C oder darüber und einer atmosphärischen CO2-Konzentration um 5000 ppm. Diese Faktoren beeinflussten nachhaltig die chemische Beschaffenheit des Meerwassers, so dass die ozeanischen Lebensgemeinschaften durch Schwefeldioxid-Eintrag, Sauerstoffverknappung sowie Versauerung und das damit verbundene Absacken des pH-Werts häufig an ihre Grenzen stießen.[47]
Die rasche Zunahme der Biodiversität im Zuge der Kambrischen Explosion führte zu einem rapiden Anstieg des Hintergrundaussterbens, das als permanente Begleiterscheinung der biologischen Evolution in der ersten Hälfte des Paläozoikums und hier besonders im Kambrium ein sehr hohes Niveau erreichte. Eine Abgrenzung zwischen dem natürlichen Artenaustausch und einem Massenaussterben ist daher schwierig, zumal einige kambrische Schichten im Hinblick auf die fossile Überlieferung erhebliche Lücken aufweisen. Für die ökologische Krise vor 510 Millionen Jahren konnte vor kurzem ein möglicher Auslöser identifiziert werden. Danach steht das Massenaussterben in Zusammenhang mit der Kalkarindji-Vulkanprovinz im heutigen Westaustralien. Die damals über längere Zeiträume freigesetzten Flutbasalte umfassten eine Fläche von 2 Millionen km² und verursachten (unter gleichzeitiger Destabilisierung des Klimas) durch ihre Ausgasungen eine starke Zunahme anoxischer Zonen in den Ozeanen. Schätzungsweise die Hälfte aller marinen Lebewesen fiel der Katastrophe zum Opfer, darunter viele Trilobiten (Dreilappkrebse), Conodonten und Brachiopoden (Armfüßer).[48]
Oberes Ordovizium (vor ca. 444 Millionen Jahren)

Im ordovizischen Massenaussterben starben etwa 85 Prozent der Arten und 60 Prozent der Gattungen aus, darunter die meisten der Brachiopoden. Die Trilobiten überlebten, doch ihre immense Vielfalt verringerte sich ebenfalls erheblich. Neuere Studien stützen die Annahme, dass die Besiedelung des Festlands durch moosartige Pflanzenteppiche und frühe Pilzformen bereits im Mittleren Kambrium begann und sich im Ordovizium verstärkt fortsetzte.[49] Die zunehmende Vegetationsbedeckung entzog den Böden eine Reihe von Elementen wie Calcium, Magnesium, Phosphor und Eisen. Daraus resultierten eine beschleunigte chemische Verwitterung der Erdoberfläche mit erhöhter Bindung von atmosphärischem Kohlenstoffdioxid und im Zusammenspiel mit anderen Faktoren ein Temperaturrückgang um ca. 5 Grad.[50] Durch die nachlassende „Pufferwirkung“ des Treibhauseffekts aufgrund der allmählichen CO2-Reduktion muss hierbei auch die im Vergleich zur Gegenwart um 4 Prozent verminderte Sonneneinstrahlung berücksichtigt werden (Solarkonstante im Ordovizium 1306 W/m², aktuell 1361 W/m²).[51] Der globale Abkühlungstrend mündete vor ungefähr 460 Millionen Jahren in das Ordovizische Eiszeitalter (auch Hirnantische Eiszeit beziehungsweise Anden-Sahara-Eiszeit). Es erreichte seine prägnanteste Ausprägung zeitgleich mit dem Höhepunkt des Massenaussterbens während der letzten ordovizischen Stufe des Hirnantiums (ab 445,2 mya) und erfasste große Teile der südlichen Hemisphäre. Ein mitbeteiligter Faktor dieses Klimawechsels war die Lage des Großkontinents Gondwana in unmittelbarer Südpolnähe mit umfangreichen Vergletscherungen in den Gebieten des heutigen Nordafrikas und Brasiliens.
In einigen aktuellen Analysen wird die Ansicht vertreten, dass nicht das Glazialklima und die damit verbundene Absenkung des Meeresspiegels (mit einem Schwund mariner Flachwasserbiotope) den primären Aussterbefaktor darstellte, sondern dass geochemische Veränderungen in Verbindung mit einem Ozeanischen anoxischen Ereignis giftige Schwermetalle wie Arsen, Blei oder Mangan am Meeresgrund freisetzten. Anhand von Mikrofossilien lässt sich zum fraglichen Zeitpunkt ein hohes Auftreten fehlgebildeter Organismen belegen, die eine deutliche Kontaminierung durch toxische Elemente aufwiesen.[52] Zusätzlich könnte auch die weitgehende Reduzierung von Spurenelementen eine gravierende Rolle gespielt haben. So erreichte die Konzentration des lebenswichtigen Spurenelements Selen am Übergang vom Ordovizium zum Silur offenbar nur einen Bruchteil des gegenwärtigen Niveaus und lag bei einigen späteren Massenaussterben wie jenen im Oberdevon ebenfalls am Minimum.[53] Diese Konstellation impliziert im Hinblick auf die mögliche Destabilisierung der Ozeane ein komplexes Ursache-Wirkungs-Gefüge, bestehend unter anderem aus den Komponenten Zu- oder Abnahme der globalen Sauerstoff- und Kohlenstoffdioxid-Werte, oxidative Erosion, anoxische Bedingungen, Meeresspiegelschwankungen sowie stark variierende Nährstoffeinträge.
Eine 2004 publizierte Studie postuliert für das Massenaussterben eine extraterrestrische Ursache in Form eines Gammablitzes.[28] Zwar stimmt die rasche Dezimierung der die oberen Meereszonen bewohnenden Organismen mit der Strahlungshypothese überein, es fehlen jedoch darüber hinaus weitere faktische Belege.
Oberdevon (vor ca. 372 und 359 Millionen Jahren)
| System | Serie | Stufe | ≈ Alter (mya) |
|---|---|---|---|
| später | später | später | jünger |
| Devon | Oberdevon | Famennium | 358,9 ⬍ 372,2 |
| Frasnium | 372,2 ⬍ 382,7 | ||
| Mitteldevon | Givetium | 382,7 ⬍ 387,7 | |
| Eifelium | 387,7 ⬍ 393,3 | ||
| Unterdevon | Emsium | 393,3 ⬍ 407,6 | |
| Pragium | 407,6 ⬍ 410,8 | ||
| Lochkovium | 410,8 ⬍ 419,2 | ||
| früher | früher | früher | älter |
Die beiden Paläontologen David M. Raup und Jack Sepkoski, unter anderem bekannt für ihre Hypothese des periodischen Aussterbens,[32] unternahmen im Jahr 1982 erstmals den Versuch, die „Big Five“ statistisch zu quantifizieren. Sie beschrieben dabei im Oberen Devon drei eher schwache, aber „aus dem Hintergrundrauschen hervorstechende“ Aussterbeereignisse (Ende Givetium, Ende Frasnium und Ende Famennium), von denen die Krise an der Frasnium-Famennium-Grenze zwar die prägnanteste war, aber ursprünglich nicht zu den Big Five zählte.[54] In den folgenden Jahrzehnten wurde eine Vielzahl weiterer Krisenzeiten ab dem Emsium identifiziert,[55] wie jenes am Ende des Eifeliums vor 387,7 Millionen Jahren, die manchmal pauschal unter der Bezeichnung mittel- bis oberdevonische Biokrise zusammengefasst wurden.[56] Hingegen konzentriert sich die aktuelle Forschung mit zunehmendem Kenntnisstand auf zwei Schwerpunkte: zum einen auf das Kellwasser-Ereignis vor 372 Millionen Jahren, benannt nach den oberdevonischen Kalksteinschichten des Kellwassertals im Oberharz, sowie auf das mindestens gleichstarke Hangenberg-Ereignis vor 358,9 Millionen Jahren, dessen Bezeichnung auf die sogenannten Hangenberg-Aufschlüsse im nordöstlichen Teil des Rheinischen Schiefergebirges zurückgeht.
- Das Kellwasser-Ereignis (international auch Frasnian-Famennian Mass Extinction) verzeichnete einen Artenschwund bis zu 75 Prozent. Betroffen waren vor allem die Faunengruppen flacher tropischer Meere, wie Fische, Korallen, Trilobiten sowie etliche „Riffbauer“, weshalb die Anzahl der Korallenriffe deutlich abnahm. Darüber hinaus wurde die Biodiversität des Phytoplanktons so drastisch reduziert, dass die ursprüngliche Artenvielfalt erst wieder im Jura erreicht wurde.[57]

- In der geologischen Überlieferung korrespondiert dieses Ereignis mit einer weltweiten Verbreitung von Schwarzschiefer-Horizonten, die auf mehrere Ozeanische anoxische Ereignisse (OAEs) zurückgehen. In der Fachliteratur finden sich die verschiedensten Szenarien für die Ursachen der oberdevonischen Krise, wie zum Beispiel eine durch die sinkende Kohlenstoffdioxid-Konzentration signifikant verstärkte Wirkung der Milanković-Zyklen,[58] verbunden mit einem plötzlichen Umkippen des gesamten Klimasystems,[59] oder der die Ozonschicht zerstörende Strahlungseinfluss einer erdnahen Supernova.[60] Als gesichert gilt, dass gegen Ende des Devons mehrmals starke Schwankungen des Meeresspiegels auftraten, die auf einen extrem kurzfristigen Zyklus verschiedener Kalt- und Warmphasen hindeuten.[61] Die in jüngster Zeit entdeckten Quecksilber-Anomalien in den Kellwasser-Schichten werden als Indikator für die Beteiligung eines Megavulkanismus am damaligen Massenaussterben gewertet[62] und können möglicherweise dem Viluy Trap im östlichen Sibirien zugeordnet werden.[63]
- Mit dem Hangenberg-Ereignis an der Schwelle zum Karbon endete eine mehr als 80 Millionen Jahre herrschende globale Warmklima-Phase. Während das Kellwasser-Ereignis noch mehrere Erwärmungsimpulse verzeichnete, begann nun eine deutliche und relativ abrupte Abkühlung mit umfangreichen Vergletscherungen in den südlichen und westlichen Regionen des Großkontinents Gondwana. Der Meeresspiegel sank um etwa 100 Meter, und die atmosphärische CO2-Konzentration reduzierte sich durch die massive Ablagerung von organischem Kohlenstoff in Schwarzschiefer-Sedimenten um rund 50 Prozent.[64] Aufgrund der sauerstoffarmen Bedingungen in den Ozeanen kam es zum Zusammenbruch mehrerer Ökosysteme und zum Massensterben von Ammoniten, Brachiopoden (Armfüßer), Trilobiten, Conodonten, Stromatoporen, Ostrakoden (Muschelkrebse), Placodermi (Panzerfische) und frühen Landwirbeltieren (Tetrapoden).[65] Die in mehrere kurzzeitige Zyklen untergliederte Krisenzeit (Unteres, Mittleres und Oberes Hangenberg-Ereignis) hatte eine Dauer von ungefähr 100.000 bis 300.000 Jahren.[65][66] Möglicherweise war an den Aussterbewellen und den klimatischen Veränderungen im Oberdevon auch eine Häufung von Impaktkatastrophen wie der australische Woodleigh-Einschlag, der Alamo-Einschlag im heutigen Nevada[67] oder die schwedische Siljan-Impaktstruktur direkt beteiligt.
Perm (vor ca. 260 Millionen Jahren)
Nach dem Abklingen der Permokarbonen Eiszeit, die im Unteren Karbon ihren Anfang genommen hatte und im Verlauf des Perms endete, zeichnete sich der Trend zu einem Warmklima ab, der jedoch mit einer markanten Umweltveränderung vor 260 Millionen Jahren einherging. Die in der Forschung bereits länger bekannte und vom Umfang her unterschiedlich bewertete Abnahme der Biodiversität in tropischen Gewässern war nach neueren Studien ein globales Ereignis, das im sogenannten Capitanium-Massenaussterben gipfelte (benannt nach der obersten chronostratigraphischen Stufe des Mittelperms beziehungsweise der Guadalupium-Serie). Als Ursache wird ein massiver Kohlenstoffdioxid- und Schwefeldioxid-Eintrag in die Ozeane mit Bildung anoxischer Zonen und starker Versauerung des Meerwassers angenommen. Ein Zusammenhang mit den zeitgleich auftretenden Flutbasalten des Emeishan-Trapps im heutigen Südchina (mit Zentrum in der Provinz Sichuan) gilt in der Wissenschaft als sehr wahrscheinlich.[68] Die unterschiedlich stark ausgeprägten Aktivitätszyklen des Trapps dauerten wahrscheinlich knapp zwei Millionen Jahre und bedeckten in dieser Zeit ein Gebiet von mindestens 250.000 km² mit basaltischen Ablagerungen.[69] Untersuchungen fossiler Therapsiden (Diictodon) im südafrikanischen Karoobecken belegen auf dem Höhepunkt des Massenaussterbens eine starke Zunahme arider Bedingungen, während die Temperaturen zumindest in diesem Gebiet konstant blieben.[70]
Obwohl das Aussterbeereignis vor allem die marine Fauna aller Klimazonen in Mitleidenschaft zog, nahm auch bei Landwirbeltieren die Artenzahl deutlich ab, wie bei verschiedenen Gruppen der Dicynodontia, Parareptilien und Pareiasauridae.[71] Erheblich dezimiert wurde darüber hinaus die Landvegetation im Umkreis des Emeishan-Trapps aufgrund von atmosphärischen Schadstoffen und einer Beeinträchtigung der Photosynthese. Aktuellen Analysen zufolge starben während der Capitanium-Krise über 60 Prozent der Arten und 33 bis 35 Prozent der Gattungen aus. Es ist nicht endgültig gesichert, ob das Ereignis im Mittleren Perm das Ausmaß der Big Five erreichte, aber es mehren sich die Hinweise, dass dies durchaus der Fall sein könnte.[7]
Perm-Trias-Grenze (vor ca. 252 Millionen Jahren)
An der Perm-Trias-Grenze ereignete sich das größte Massenaussterben des Phanerozoikums. Bezogen auf die Artenanzahl starben in diesem Zeitraum etwa 95 Prozent aller marinen Lebensformen und etwa 75 Prozent der Landfauna aus, darunter – ein Novum in der Erdgeschichte – auch viele Insektenarten. Die Vegetation wurde durch Brände, Hitze- und Dürreperioden, Luftverschmutzung und Bodenversauerung so stark dezimiert, dass der atmosphärische Sauerstoffgehalt von rund 30 Prozent am Beginn des Perms gegen Ende der Periode auf 10 bis 15 Prozent sank und anschließend Millionen Jahre (über die gesamte Trias und bis in den Jura) auf einem niedrigen Level verharrte. Als Auslöser und Hauptursache für den Zusammenbruch der Ökosysteme gilt der großflächige Flutbasalt-Ausstoß des Sibirischen Trapps, der während seiner Aktivitätszyklen über rund 900.000 Jahre eine Fläche von 7 Millionen Quadratkilometern mit magmatischen Gesteinen bedeckte. Neuere Studien gehen davon aus, dass dabei umfangreiche Mengen an Flugasche aus Kohlebränden und Vegetationsrückständen in die Atmosphäre gelangten und weiträumig verteilt abgelagert wurden.[72][73] Zusätzlich könnten Gase (Kohlenstoffdioxid, Schwefeldioxid, Methan, Chlormethan und Brommethan), die der Kontaktmetamorphose entlang der ausgedehnten unterirdischen Wegsamkeiten des Magmas entstammten – insbesondere entlang schichtparalleler Gänge (Sills) im Tunguska-Sedimentbecken –, die Ausgasungen der an die Oberfläche vordringenden Flutbasalte zusätzlich verstärkt haben.[74][75]
Laut neueren Erkenntnissen lässt sich das damalige Massenaussterben in mehrere Abschnitte untergliedern. Während der Phase 1 stieg die globale Temperatur aufgrund des vom Sibirischen Trapp emittierten Kohlenstoffdioxids innerhalb eines geologisch sehr kurzen Zeitraums um 5 °C. Parallel dazu erwärmten sich auch die oberflächennahen Wasserschichten der Ozeane bis zum Ende des Ereignisses um mindestens 8 °C,[76] was den Übergang zu Phase 2 markierte: Mit der Bildung von sauerstofffreien (anoxischen) Meereszonen, dem rapiden Absacken des pH-Werts[77] sowie der Freisetzung von Methanhydrat begann das Massensterben in den Ozeanen. Durch das in die Atmosphäre entweichende Methan erhöhte sich die weltweite Temperatur um weitere 5 °C, und die Treibhausgas-Konzentration erreichte einen CO2-Äquivalentwert von mindestens 3.000 ppm.[78] Darüber hinaus wird gelegentlich ein kurzfristig auftretender galoppierender Treibhauseffekt postuliert (englisch runaway greenhouse effect) auf der Basis eines Kohlenstoffdioxid-Levels von über 7.000 ppm.[79] Durch die unter diesen Bedingungen verstärkt verlaufenden Erosionsprozesse kam es zu umfangreichen Einschwemmungen festländischer Verwitterungsprodukte in die marinen Biotope, mit anschließenden Überdüngungseffekten (Eutrophierung) wie zum Beispiel Algenblüten.[80]
Eine weitere Ursache für die Destabilisierung der Ökosysteme war vermutlich die Massenvermehrung mariner Einzeller in sauerstofffreien Milieus, die ihre Stoffwechselprodukte in Form von Methan, Halogenkohlenwasserstoffen und großen Mengen Schwefelwasserstoff (H2S) in die Atmosphäre emittierten.[14] Zusätzlich wird vermutet, dass durch die Schadstoffbelastung der Luft auch die Ozonschicht in erheblichem Maße abgebaut wurde, mit der Folge einer verstärkten UV-B-Einstrahlung, die möglicherweise zu Pollenmissbildungen und zu einem großflächigen Vegetationsrückgang beigetragen hat.[81]
Die Dauer der Perm-Trias-Krise wurde bis vor Kurzem auf mehr als 200.000 Jahre veranschlagt, laut einer Analyse von 2014 reduziert sich dieses Zeitfenster auf zwei Kernbereiche, die jeweils 60.000 Jahre (± 48.000 Jahre) umfassten.[82] Hingegen postuliert eine 2018 veröffentlichte Studie ein Einzelereignis mit einem zeitlichen Umfang von maximal rund 30.000 Jahren, möglicherweise beschränkt auf wenige Jahrtausende.[83] Der globale Kollaps der Ökosysteme konnte mithilfe neuentwickelter Präzisionsmessungen auf die unterste Trias vor 251,94 Millionen Jahren datiert werden. Die biologischen, geophysikalischen und klimatischen Spätfolgen des Massenaussterbens reichten zum Teil bis in die Mittlere Trias. Während sich der Formenkreis der Ammoniten, Conodonten und Foraminiferen innerhalb von 1 bis 3 Millionen Jahren erholte, benötigten Korallenriffe 8 bis 10 Millionen Jahre zu ihrer vollständigen Regeneration. Noch länger dauerte die Entstehung neuer Waldhabitate, die sich erst nach etwa 15 Millionen Jahren über größere Areale ausbreiteten. Die schrittweise Erneuerung der durch extreme Erwärmung, Großbrände, sauren Regen und Sauerstoffverknappung geschädigten Biotope („Recovery Phase“) wurde in der Trias mit Schwerpunkt in den chronostratigraphischen Unterstufen Smithium und Spathium durch weitere Aussterbe-Ereignisse mehrmals unterbrochen.[84][85]
Trias-Jura-Grenze (vor ca. 201 Millionen Jahren)
Gegen Ende der Trias kündigte sich mit dem beginnenden Zerfall des seit dem späten Karbon existierenden Superkontinents Pangaea ein geologisches Großereignis an. Entlang der Plattenränder des heutigen Nordamerikas und Europas entstanden ausgedehnte, bis nach Nordafrika reichende Grabenbrüche (Riftsysteme) mit ersten marinen Ingressionen. Aus dieser Entwicklung, hin zur allmählichen Öffnung des späteren Zentralatlantiks, resultierte an der Trias-Jura-Grenze die Entstehung der 11 Millionen km² umfassenden Zentralatlantischen Magmatischen Provinz (englisch Central Atlantic Magmatic Province, abgekürzt CAMP), deren Magmaausflüsse zu den ergiebigsten der bekannten Erdgeschichte zählen. Die Hauptphase des Flutbasalt-Vulkanismus setzte vor 201,5 Millionen Jahren ein, dauerte ungefähr 600.000 Jahre und wies wahrscheinlich vier kurzzeitige Schübe mit stark erhöhter Aktivität auf.[86] Dieser plattentektonische Prozess hatte gravierende Folgen für Atmosphäre, Klima und Biosphäre und gilt allgemein als primäre Ursache für das Massenaussterben mit einem Artenschwund von annähernd 70 bis 75 Prozent.[87][88] Vollständig ausgelöscht wurden in den Meeren die Conodonten, und an Land erfolgte die weitgehende Dezimierung der nicht zu den Dinosauriern zählenden Archosauria. In erheblichem Umfang betroffen waren auch andere Reptilien aus der Gruppe der Diapsida, viele Amphibien sowie einige Vorläufer der Säugetiere.
Mehrere aktuelle Studien kommen zu dem Resultat, dass der Schwerpunkt des Massenaussterbens etwa 100.000 Jahre vor der eruptiven Flutbasaltphase des CAMP-Ereignisses anzusetzen sei. Laut diesen Analysen begann die Aktivität der Zentralatlantischen Magmatischen Provinz mit einem intrusiven Stadium. In dessen Verlauf strömten große Mengen Magma in Evaporit- und Carbonatlagerstätten (mit zusätzlichen Anteilen von Kohlenwasserstoffen) und bewirkten über Zeiträume von einigen tausend oder zehntausend Jahren durch Kontaktmetamorphose die Ausgasung von Kohlenstoffdioxid im fünfstelligen Gigatonnenbereich.[89][90]
Die an dem Massenaussterben beteiligten Faktoren waren somit im Wesentlichen identisch mit jenen an der Perm-Trias-Grenze: ein starker und rascher Anstieg der atmosphärischen Kohlenstoffdioxid-Konzentration und damit verbunden eine rapide Erwärmung, eine durch den CO2- und Schwefeldioxid-Eintrag verursachte Versauerung des Ozeans, eine Biokalzifikationskrise zu Lasten Kalkschalen bildender mariner Organismen sowie möglicherweise die Freisetzung von bakteriell erzeugtem Methan und Schwefelwasserstoff. In der oberen, euphotischen Zone des Tethysmeeres – eines großen Randmeeres des Superkontinentes Pangaea und Biodiversitäts-Hotspots – könnten die hohen Schwefelwasserstoff- und niedrigen Sauerstoffkonzentrationen dazu beigetragen haben, dass sich marine Ökosysteme dort nur sehr langsam erholten.[91]
Kreide-Paläogen-Grenze (vor ca. 66 Millionen Jahren)
Die Kreide-Paläogen-Grenze (früher Kreide-Tertiär-Grenze) markiert gleichzeitig den Wechsel vom Erdmittelalter zur Erdneuzeit. An dieser geologischen Nahtstelle starben rund 70 bis 75 Prozent aller Tierarten aus, darunter mit Ausnahme der Vögel auch die Dinosaurier. Als Ursache werden in der Fachliteratur zwei Ereignisschwerpunkte diskutiert: zum einen der Einschlag (Impakt) eines Asteroiden nahe der Halbinsel Yucatán im heutigen Mexiko, wodurch der 180 km durchmessende Chicxulub-Krater entstand, und zum anderen das alternative Flutbasalt-Szenario des Dekkan-Trapps im westlichen Indien. Einige Studien postulierten zu Beginn der 2000er Jahre, dass der Chicxulub-Krater 300.000 Jahre vor der eigentlichen Kreide-Paläogen-Grenze entstanden sei.[92] Diese Annahme wird in Anbetracht des aktuellen Datenmaterials inzwischen als unwahrscheinlich eingestuft.[93] Die Anwendung von Datierungsmethoden mit sehr geringen Toleranzbereichen führte zu dem Resultat, dass Impaktereignis und Kreide-Paläogen-Grenzschicht zeitlich präzise übereinstimmen.[94] Auch der dem Einschlag folgende Impaktwinter gilt inzwischen als faktisch gesichert.[95] Somit ist der Chicxulub-Impakt nach derzeitigem Kenntnisstand die primäre Ursache des damaligen Massenaussterbens.[96][97]

Das in der Wissenschaft gebräuchlichste Szenario geht davon aus, dass vor 66,040 Millionen Jahren (± 0,032 Mio. Jahre) ein etwa 14 km großer Asteroid mit einer Geschwindigkeit um 20 km/s in einem tropischen Flachmeer detonierte und innerhalb einer Sekunde verdampfte. Durch die Wucht der Explosion wurden mehrere tausend Kubikkilometer Carbonat- und Evaporitgestein als glühende Ejekta bis in die Stratosphäre und zum Teil darüber hinaus geschleudert.[98] Neben den unmittelbaren Folgen des Einschlags wie Megatsunamis, einer überschallschnellen Druckwelle sowie Erdbeben im Bereich der Stärke 11 oder 12 traten weltweit Wald- und Flächenbrände auf, mit einer kurzzeitigen, aber starken Erhitzung der Atmosphäre.[99] Innerhalb weniger Tage verteilte sich in der gesamten Atmosphäre eine große Menge an Ruß- und Staubpartikeln, die das Sonnenlicht über Monate hinweg absorbierten, einen globalen Temperatursturz herbeiführten und die Photosynthese der meisten Pflanzen zum Erliegen brachten. Ein zusätzlicher Abkühlungsfaktor war möglicherweise eine atmosphärische Schicht von Schwefelsäure-Aerosolen, die laut einer aktuellen Untersuchung einen Temperatursturz von 26 °C bewirkt haben könnten und maßgeblich dazu beitrugen, dass die globalen Durchschnittstemperaturen für einige Jahre unter den Gefrierpunkt sanken, mit dramatischen Folgen für die gesamte Biosphäre.[100] Von dieser Krise waren die ozeanischen und festländischen Bereiche gleichermaßen betroffen. 75 Prozent der Arten fielen dem Massenaussterben innerhalb eines wahrscheinlich sehr kurzen, aber nicht genau zu bestimmenden Zeitraums zum Opfer, darunter nicht nur die Saurier, sondern auch die Ammoniten, die großen Meeresreptilien wie Plesio- oder Mosasaurier, fast alle kalkschalenbildenden Foraminiferen sowie verschiedene Planktongruppen. Von den kreidezeitlichen Vogelarten überstand nur ein sehr kleiner Teil die Zäsur des Massenaussterbens[101] (→ #Aussterbe- und Überlebensmuster am Beispiel des Chicxulub-Einschlags).
Eine im April 2015 von mehreren bekannten Geowissenschaftlern vorgelegte Hypothese geht davon aus, dass aufgrund der Impaktenergie von 3×1023 Joule (nach einer anderen Berechnung 1×1024 Joule) und der dadurch ausgelösten tektonischen Schockwellen der lange „schwelende“ Dekkan-Trapp eine erhebliche Zunahme seiner Aktivität verzeichnete. Laut dieser Hypothese ist der kurzfristige Ausstoß von 70 Prozent aller Dekkan-Trapp-Flutbasalte auf den Chicxulub-Einschlag zurückzuführen.[15] Die bisher wenig beachtete Möglichkeit einer kausalen Verbindung zwischen Asteroideneinschlag und Flutbasalt-Vulkanismus wird derzeit in den Geowissenschaften auf breiter Basis diskutiert.[102] In der Wissenschaft herrscht jedoch Uneinigkeit darüber, ob die Hauptaktivität dieser Magmatischen Großprovinz unmittelbar an beziehungsweise kurz nach der Kreide-Paläogen-Grenze oder bereits vorher stattfand.[103]
Eozän-Oligozän-Grenze (vor 33,9 Millionen Jahren)

Im Rahmen des Grande Coupure („Großer Einschnitt“) ereignete sich am Eozän-Oligozän-Übergang vor 33,9 bis 33,7 Millionen Jahren ein großes Artensterben, das mit einem markanten Temperaturabfall an Land und in den Weltmeeren verknüpft war (englisch Eocene-Oligocene extinction event). Betroffen von dem rasch einsetzenden Klimawandel und dessen Folgeerscheinungen waren im europäischen Raum etwa 60 Prozent der eozänen Säugetiergattungen, darunter ein Großteil der damaligen Palaeotherien (frühe Pferdeverwandte), Primaten, Creodonta und andere Tiergruppen. Existierte in Europa während des Eozäns noch eine subtropische Vegetation, entstanden nun aufgrund des kühleren und trockeneren Klimas weitläufige Steppengebiete. Auf globaler Ebene sank die Temperatur der Ozeane bis in tiefere Regionen um 4 bis 5 °C, und der Meeresspiegel fiel um etwa 30 Meter.
Die klimatischen Veränderungen an der Eozän-Oligozän-Grenze lassen sich zumindest teilweise mit plattentektonischen Prozessen erklären. Bis in das spätere Eozän waren Antarktika und Südamerika durch eine Landbrücke miteinander verbunden, ehe sich die Drakestraße allmählich öffnete. Dadurch entstand im Südpolarmeer die stärkste Meeresströmung der Erde, der Antarktische Zirkumpolarstrom, der Antarktika von nun an im Uhrzeigersinn umkreiste, den Kontinent von der Zufuhr wärmeren Meerwassers abschnitt und die Grundlage für die Bildung des Antarktischen Eisschildes schuf. Somit war Antarktika nicht nur geographisch, sondern auch thermisch isoliert, und die auf dem Kontinent einsetzende Inlandsvereisung vor 33,7 Millionen Jahren war gleichbedeutend mit dem Beginn des Känozoischen Eiszeitalters. Auch in der Nordpolarregion konnte parallel dazu eine deutliche Abkühlung festgestellt werden. Funde von Dropstones belegen die zeitweilige Existenz von grönländischem Kontinentaleis vor 38 bis 30 Millionen Jahren.[104] Zusätzlich wird angenommen, dass der lange Zeit isolierte Arktische Ozean nach einem Übergangsstadium als Brackwassermeer im frühen Oligozän (≈ 32 mya) mit dem Einströmen von salzhaltigem Nordatlantikwasser Anschluss an die globale Meereszirkulation fand.[105]
Auffällig in dem Zusammenhang ist der steile Abfall der atmosphärischen CO2-Konzentration. Bewegte sich diese gegen Ende des Eozäns noch in einem Bereich von 700 bis 1.000 ppm, sank dieses Level vor 33,7 Millionen Jahren abrupt um etwa 40 Prozent (und lag eventuell für einen erdgeschichtlich sehr kurzen Zeitraum noch tiefer).[106] Verschiedentlich wird deshalb ein zusätzlicher Abkühlungsfaktor, zum Beispiel in Form eines großen Meteoriteneinschlags, diskutiert. Als Kandidat am ehesten geeignet wäre der Popigai-Impakt im nördlichen Sibirien, mit einem „offiziellen“ Alter von 35,7 Millionen Jahren. Laut einer kürzlich erfolgten Neudatierung liegt das Impakt-Ereignis jedoch unmittelbar an der Eozän-Oligozän-Grenze und könnte, falls sich die Angaben bestätigen, ursächlich mit dem damaligen Massensterben verknüpft gewesen sein.[107]
Pliozän-Pleistozän-Übergang (vor 3,8 bis 2,4 Millionen Jahren)
Die sich am Ende des Pliozäns verstärkende globale Abkühlung ging mit dem Beginn des Pleistozäns vor rund 2,6 Millionen Jahren in die Quartäre Eiszeit über (als jüngster Zeitabschnitt des Känozoischen Eiszeitalters). Damit verbunden war eine relativ rasch einsetzende Vergletscherung großer Teile der nördlichen Hemisphäre sowie eine allmähliche Senkung des Meeresspiegels um mehr als 100 Meter. Dies führte zu einer Reduzierung beziehungsweise Austrocknung flacher Schelfmeere um ungefähr 30 Prozent und damit zu einem Verlust ozeanischer Lebensräume.[108] Davon besonders betroffen war die in Küstennähe beheimatete marine Megafauna, an ihrer Spitze der etwa 15 Meter große Riesenhai Megalodon,[109] daneben verzeichneten Meeressäuger und Meeresschildkröten einen Artenschwund um etwa 50 Prozent. Weltweit nahm die genetische Vielfalt der Megafauna während der Übergangszeit vom Pliozän zum Pleistozän um 15 Prozent ab.
Jungpleistozän (vor ca. 50.000 bis 12.000 Jahren)
_-_Mauricio_Ant%C3%B3n.jpg.webp)
Gegen Ende des Pleistozäns und partiell im frühen Holozän wurde im Verlauf der Quartären Aussterbewelle ein großer Teil der Megafauna Amerikas, Eurasiens und Australiens ausgelöscht. Obwohl nur relativ wenige Tierarten davon betroffen waren, hat das Ereignis in der öffentlichen Rezeption eine gewisse Bedeutung, da sich darunter sehr große und „populäre“ Tierarten befanden, wie das Mammut, das Wollnashorn und die Säbelzahnkatze. Die Gründe für das auf verschiedenen Kontinenten zeitversetzt stattgefundene Aussterben sind weitgehend ungeklärt und Gegenstand einer wissenschaftlichen Kontroverse. Häufig wird die Annahme vertreten, dass der Mensch im Zuge seiner Einwanderung in neue Lebensräume diverse Großsäuger-Arten durch intensive Bejagung (Overkill-Hypothese) ausgerottet hat. Andererseits könnte auch der drastische Klimawandel am Ende der letzten Vereisungsphase ein entscheidender Aussterbeimpuls gewesen sein. Neuere Publikationen kombinieren klimatische und anthropogene Faktoren zu einem Erklärungsmodell, wobei dem menschlichen Einfluss ein deutliches Übergewicht zugeschrieben wird.[110]
Das pleistozäne und das heutige Massensterben gehen in manchen Regionen ohne klare zeitliche Abgrenzung ineinander über. So verschwanden einige Arten, wie etwa das Wollhaarmammut und der Riesenhirsch, je nach Region zu unterschiedlichen Zeiten im Verlauf des Spätpleistozäns und Holozäns. Während die Wollhaarmammuts des eurasischen und nordamerikanischen Festlands das Ende des Pleistozäns nicht mehr erlebten, überdauerten einige Populationen auf abgelegenen Inseln bis in das Holozän. Als letzter Vertreter der eiszeitlichen Megafauna starb eine Zwergform des Mammuts auf der 7608 km² großen Wrangelinsel im Arktischen Ozean erst vor rund 3.700 Jahren aus, nachdem Menschen erstmals die Insel erreicht hatten.[111]
Im Jahr 2007 wurde in einer aufsehenerregenden Studie postuliert, dass ein Asteroid oder ein Kometenfragment vor etwa 12.800 Jahren mit der Erde kollidierte.[112] Die Clovis-Kometen-Theorie geht davon aus, dass der Einschlag (beziehungsweise die Detonation des kosmischen Körpers in der Lufthülle) vor allem im Nordatlantikraum schwerwiegende Folgen zeitigte. Laut dieser Hypothese war der Impakt nicht nur für das Verschwinden der pleistozänen Megafauna verantwortlich, sondern vernichtete auch die frühzeitliche Clovis-Kultur auf dem amerikanischen Kontinent. Darüber hinaus sollen sich aufgrund des Einschlags erhebliche Mengen an Asche und Staub in der Atmosphäre verteilt haben, die die Sonneneinstrahlung erheblich dämpften und maßgeblich zum jähen Kälteeinbruch der Jüngeren Dryaszeit beitrugen. Obwohl die Hypothese in der Fachliteratur ein oft besprochenes und vielfach rezipiertes Thema war, wurde sie mangels überzeugender Belege von der Wissenschaft bis vor kurzem mehrheitlich abgelehnt.[113] Mehrere Studien neueren Datums führen jedoch eine Reihe von Indizien auf, die für einen Impakt sprechen.[114][115]
Weitere ökologische und biologische Krisen
Die folgenden Abschnitte beschreiben jene Ereignisse im Phanerozoikum, die sich zwar deutlich vom natürlichen Hintergrundaussterben abheben, jedoch nicht unter den Begriff Massenaussterben fallen: entweder weil nur relativ wenige Arten oder Gattungen davon betroffen waren oder weil das Ausmaß der Umweltveränderungen durch die Weiterentwicklung analytischer Methoden erst in den letzten Jahren genauer bestimmt werden konnte und gegenwärtig immer noch diskutiert wird.
Mittleres Silur und Silur-Devon-Grenze

Bis Anfang der 1990er Jahre galt das Silur als ereignisarme Epoche mit stabilem Warmklima. Inzwischen wurde auch in diesem Zeitabschnitt eine Reihe biologischer Krisen und Massensterben identifiziert, mit Schwerpunkt in der Wenlock-Serie (433,4 bis 427,4 mya) sowie nahe der Silur-Devon-Grenze (419,2 mya). Während des Wenlocks erfolgte in der letzten Phase der Kaledonischen Gebirgsbildung der endgültige Zusammenschluss der Kontinentalplatten von Laurentia, Baltica und Avalonia zum Großkontinent Laurussia. Daraus resultierte ein intensiver Vulkanismus, der chemische und klimatische Anomalien in der Atmosphäre und den Ozeanen hervorrief und den kurz- und langfristigen Kohlenstoffkreislauf nachhaltig beeinflusste. Davon betroffen waren vor allem die marinen Lebensformen der Conodonten und verschiedene Planktongruppen wie die Graptolithen. Bei letzteren stieg die Aussterberate bis auf 95 Prozent, ehe die Artenvielfalt über längere Zeiträume wieder zunahm.[116] Gegen Ende des Silurs machte sich eine deutliche Abkühlung der Ozeane bemerkbar, gekoppelt mit einer Umgestaltung der Tiefenwasserströmungen und einer Veränderung des organischen Kohlenstoffzyklus. Die daraus entstehende Störung des ökologischen Gleichgewichts einschließlich mehrerer Aussterbewellen ist unter anderem in der fossilen Überlieferung der Cephalopoden (Kopffüßer) im Detail dokumentiert.[117]
Karbon-Perm-Grenze (vor 298,9 Millionen Jahren)
Die über Jahrmillionen kaum veränderte Position des Großkontinents Gondwana im Umkreis der Antarktis trug durch die Wirkung der Eis-Albedo-Rückkopplung wesentlich zur Entstehung des Permokarbonen Eiszeitalters bei, das mit einer Dauer von annähernd 80 Millionen Jahren vom Unterkarbon bis in das Mittlere Perm reichte. Ein zusätzlicher Abkühlungsfaktor war die während des Karbons zunehmende Ausbreitung tief wurzelnder und das Erdreich aufspaltender Gewächse.[118] Die Kombination von verstärkter Bodenerosion mit umfangreichen Inkohlungsprozessen entzog der Atmosphäre große Mengen an Kohlenstoff. Im Laufe dieser Entwicklung sank die globale Temperatur allmählich auf ein eiszeitliches Niveau,[119] und die atmosphärische CO2-Konzentration fiel gegen Ende der Epoche auf die bis dahin niedrigsten Werte im Phanerozoikum, mit einer an die verschiedenen Kalt- und Warmphasen gekoppelten Schwankungsbreite von 150 bis 700 ppm.[120] Laut einer Studie von 2017 verringerte sich der Kohlenstoffdioxid-Gehalt im frühen Perm auf das Minimum von wahrscheinlich 100 ppm. Sollte sich diese Annahme bestätigen, wäre das Erdklimasystem vor rund 298 Millionen Jahren in unmittelbare Nähe jenes Kipppunkts gerückt, der den Planeten in den Klimazustand einer globalen Vereisung überführt hätte, vergleichbar den Schneeball-Erde-Ereignissen im Neoproterozoikum.[121]
Im späten Karbon kam es zum Zusammenbruch der in Äquatornähe angesiedelten Regenwälder (in der Fachliteratur Carboniferous Rainforest Collapse) und damit zum ersten pflanzlichen Massenaussterben.[122] Die tropischen Wälder wurden innerhalb einer geologisch sehr kurzen Zeitspanne bis auf einige Vegetationsinseln dezimiert, und ebenso verschwand die Mehrzahl der Feucht- und Sumpfgebiete.[123] Vom Verlust dieser Biotope besonders betroffen waren verschiedene Gliederfüßer, ein Großteil der Amphibien und frühe Reptilien mit semiaquatischer Lebensweise.[124] Während der Übergangsphase vom Karbon zum Perm entstanden neue Waldbiotope, die an ein kühleres und trockenes Klima mit jahreszeitlich bedingten Temperaturschwankungen angepasst waren. Ein Beispiel für diesen Wandel ist die kälteresistente und laubabwerfende Glossopteris-Flora im südlichen Teil von Gondwana, die sich dort zum vorherrschenden Pflanzentypus entwickelte.
Jura-Kreide-Grenze (vor 145,0 Millionen Jahren)
Ebenso wie das Silur wurde auch der Jura in geowissenschaftlichen Abhandlungen lange als vergleichsweise ruhige Epoche beschrieben. Diese Einschätzung änderte sich in den letzten Jahrzehnten grundlegend. Demnach war der mittlere Abschnitt des Mesozoikums eine Zeit umfangreicher tektonischer Prozesse, starker Klima- und Meeresspiegelschwankungen sowie einer atmosphärischen Sauerstoffkonzentration, die deutlich unter dem gegenwärtigen Niveau lag.[125] Zudem gilt der gegen Ende des Juras einsetzende und bis in die frühe Unterkreide reichende Abkühlungstrend als Indikator für ein mögliches Massenaussterben. In dieser Frage ist die Fachwelt jedoch gespalten. Während einesteils ein signifikanter Artenschwund besonders bei den marinen Teleosauriden und Ichthyosauriern postuliert wird, widersprechen andere Studien dieser Annahme und verweisen auf erhebliche Lücken im Fossilbericht zum fraglichen Zeitpunkt.[126] Insgesamt scheint die Jura-Kreide-Grenze von allen Epochenübergängen des Phanerozoikums in paläobiologischer Hinsicht der unauffälligste zu sein.
Cenomanium-Turonium-Grenze (vor 93,9 Millionen Jahren)

Die einen Zeitraum von 79 Millionen Jahren umfassende kreidezeitliche Periode wies fast durchgehend tropische bis subtropische Bedingungen bis in höhere Breiten auf.[127] Nach relativ kühlem Beginn kam es im Klimaoptimum der Mittleren und Oberen Kreide zu einer der stärksten Erwärmungsphasen im Phanerozoikum, ehe die Temperaturen gegen Ende der Kreide allmählich wieder absanken, im späten Maastrichtium aufgrund des Dekkan-Trapp-Vulkanismus mit abrupten Klimawechseln und vermutlich saisonaler Meereisbedeckung sowie der Bildung von Kontinentaleis in höhergelegenen antarktischen Regionen.[128] Dieser langfristige Temperaturtrend wurde mehrmals von ausgeprägten Klimaschwankungen unterbrochen, die mit variierenden CO2-Konzentrationen, umfangreichen tektonischen Aktivitäten und starkem Vulkanismus einhergingen. Eine Besonderheit der Kreide ist die Häufung von Ozeanischen anoxischen Ereignissen, wobei jenes an der Cenomanium-Turonium-Grenze (kurz OAE-2 genannt, auch Bonarelli-Ereignis) globale Ausmaße erreichte und wahrscheinlich die markanteste Störung des Kohlenstoffkreislaufs der letzten 100 Millionen Jahre war, mit prägnanten klimatischen und biologischen Auswirkungen.[129] In der älteren Literatur wird dem Ereignis eine Dauer von 300.000 Jahren zugeschrieben, neuere Studien veranschlagen rund 700.000 Jahre.[130] Während der anoxischen Umweltkrise, verknüpft mit einer Abkühlung von etwa 4 °C, wurden verschiedene Planktonarten und Riffbauer sowie die Klasse der Cephalopoden (darunter Ammoniten und Belemniten) stark dezimiert, und mit der Gattung Platypterygius starben die letzten Vertreter der Ichthyosaurier aus.[131] Das zeitlich gestaffelte, aber letztlich abrupte Verschwinden der Fischsaurier ist Ausdruck einer Reihe von biotischen Ereignissen während des OAE-2, die anschließend zur Ausbreitung neuer Arten in veränderten marinen Ökosystemen führte.
Paläozän/Eozän-Temperaturmaximum (vor 55,8 Millionen Jahren)
Das Paläozän/Eozän-Temperaturmaximum (PETM) war eine sehr starke Erwärmungsphase mit einer Dauer von 170.000 bis 200.000 Jahren. Der globale Temperaturanstieg erfolgte auf der Basis eines bereits vorhandenen Warmklimas und verlief parallel zu einer deutlichen Konzentrationszunahme von Treibhausgasen in Atmosphäre und Weltmeeren.[132] Während des PETM stieg die globale Temperatur innerhalb weniger Jahrtausende von etwa 18 auf 24 °C. Dies bewirkte durch den umfangreichen Eintrag von Kohlenstoffdioxid eine Versauerung der Meere bis in tiefere Schichten und die Entstehung anoxischer Milieus.[133] Während auf dem Festland die Ausdehnung der tropischen Klimazone zu weiträumigen Migrationsbewegungen von Flora und Fauna[134] und bei einigen Gattungen der Säugetiere zu Kleinwüchsigkeit aufgrund von Nahrungsmangel in Verbindung mit phänotypischen Reaktionen führte,[135][136] geschah in den Ozeanen ein Massensterben der benthischen Foraminiferen mit einem Artenschwund zwischen 30 und 50 Prozent. An der Destabilisierung der marinen Biotope hatte die deutliche Abnahme des pH-Werts entscheidenden Anteil. Davon in Mitleidenschaft gezogen, jedoch nur partiell vom Aussterben bedroht waren in der Tiefsee angesiedelte Organismen (Seeigel, Muscheln, Schnecken) sowie nahezu alle Planktongruppen. Eine Tendenz zur „Verzwergung“ erfasste auch viele marine Arten, darunter die Ostrakoden (Muschelkrebse). Diese Entwicklung resultierte sehr wahrscheinlich aus der Erwärmung und Versauerung der Tiefseeregionen und einer damit verknüpften Störung der Remineralisierungsprozesse von organischem Kohlenstoff.[137]
Das gegenwärtige Massenaussterben
Die vorwiegend auf menschlichen Einflüssen beruhende Aussterbewelle begann am Übergang vom Pleistozän zum Holozän und setzt sich, unter zunehmender Verstärkung, bis in die Gegenwart fort. Die Weltnaturschutzunion (IUCN) geht davon aus, dass der aktuell registrierte Artenschwund die Rate des normalen Hintergrundaussterbens um das 1.000- bis 10.000-fache übertrifft.[139] Eine weitere Untersuchung deutet ebenfalls darauf hin, dass das sechste Massenaussterben bereits begonnen hat.[140] Seit der Sesshaftwerdung des Menschen in der Jungsteinzeit wurde die Tierwelt, insbesondere die Megafauna, stark zurückgedrängt. Als Menschen ab etwa 800 n. Chr. weitere Inseln besiedelten, rotteten sie die dortige Megafauna, die eine niedrige Reproduktionsrate besaß und über keine Fluchtreflexe verfügte, durch exzessive Bejagung aus (Madagaskar, die Moas in Neuseeland) oder brachten die ansässige Vogelwelt zum Verschwinden (Hawaii, Polynesien). Eine Beschleunigung des Aussterbens trat mit dem Zeitalter der Entdeckungen ab ca. 1500 ein, als Europäer andere Kontinente besiedelten, vermeintliche Schädlinge ausrotteten und Neozoen einschleppten (Ratten, Füchse, Schweine), denen die einheimische Fauna oftmals wehrlos ausgeliefert war.
Der Jahresbericht 2014[141] der Umweltstiftung World Wide Fund For Nature (WWF) spricht von einer teilweise dramatisch zunehmenden Verschlechterung der Lage vieler Arten wie etwa Nashörner (von einer Unterart, dem Nördlichen Breitmaulnashorn, gibt es laut WWF nur noch wenige Exemplare), Elefanten (Wilderer in Afrika erlegen mehr Elefanten, als Nachwuchs geboren wird), Löwen (in Westafrika stehen sie vor dem Aussterben, in Indien gibt es nur noch Restbestände) oder Walrosse (sie werden Opfer des Klimawandels, ihre Ruheplätze auf Eisschollen verschwinden mit dem Rückgang des arktischen Packeises). Auch viele andere Tiere verlieren laut WWF ihren Lebensraum: Menschenaffen wie die Bonobos verlieren ihre letzten Schutzgebiete, beispielsweise infolge einer in einem Nationalpark im Kongo geplanten Erdölförderung. Bei den Primaten finden sich inzwischen 94 Prozent auf der Roten Liste in einer der drei höchsten Gefährdungskategorien (Stand 2014). Laut WWF hat die Artenvielfalt seit den 1970er-Jahren stark gelitten; die Zahl der Säugetiere, Vögel,[142] Reptilien und Fische habe sich seither im Schnitt halbiert, die Welt verliere täglich 380 Tier- und Pflanzenarten.[143]
„Der Mensch verursacht gerade das größte globale Artensterben seit dem Verschwinden der Dinosaurier.“
Aktuelle Daten
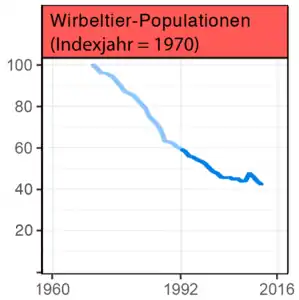
Der im September 2020 veröffentlichte, zusammen mit der Zoologischen Gesellschaft London (ZSL) und dem Global Footprint Network (GFN) erstellte Living Planet Report („Lebender-Planet-Bericht“) des WWF meldete bei weltweit über 14.000 untersuchten Tierpopulationen einen Rückgang der Bestände um ca. 70 Prozent innerhalb von 50 Jahren.[145][146]
Laut dem Bericht aus 2016 haben die weltweiten Tierbestände in Flüssen und Seen im Schnitt um 81 Prozent abgenommen.[147] Der Report von 2020 identifizierte die zunehmende Entwaldung und landwirtschaftliche Expansion als Hauptursachen, wobei der stärkste Rückgang (94 Prozent) in den subtropischen Gebieten Amerikas registriert wurde.[146]
„Wenn wir die ökologische Landwirtschaft nicht deutlich ausdehnen, können wir nicht sicher sagen, ob in zehn oder 20 Jahren von der jetzigen Vogelpopulation noch etwas übrig ist.“
Eine 2020 publizierte Untersuchung zu vom Aussterben bedrohter Wirbeltiere kam zu dem Ergebnis, dass sich das „sechste Massenaussterben“ beschleunigt, wobei ein wesentlicher Aspekt darin besteht, dass das Verschwinden mancher Tierarten wiederum die Existenz anderer Arten gefährdet.[149][150]
Nach einem Bericht des Weltbiodiversitätsrats (IPBES-Artenschutzkonferenz in Paris) vom Mai 2019 sind bis zu eine Million Arten vom Aussterben bedroht,[151] 500.000 davon werden als „dead species walking“ (sinngemäß lebende Tote) bezeichnet. Bei den Pflanzen sind die Palmfarne besonders bedroht, einige Arten sind bereits ausgestorben.[152]
Ursachen
Die unten aufgeführten (Risiko-)Faktoren tragen in ihrer Gesamtheit und auf unterschiedlich langen Zeitskalen zur Möglichkeit eines anthropogen bedingten Massenaussterbens bei:
- Intensive Landnutzung, vor allem Abholzung beziehungsweise Rodung von Wäldern sowie die Umgestaltung natürlicher Ökosysteme zu landwirtschaftlich nutzbaren Flächen (Monokulturen), insbesondere durch die Ausweitung der Massentierhaltung und dem dazu benötigten Futtermittelanbau, unter Ausbringung von Pestiziden mit der Gefährdung z. B. zahlreicher Insektenarten[153]
- Großräumige Flächenversiegelung
- Gewerbsmäßige Jagd auf geschützte Tierarten
- Klimaerwärmung auf regionaler und globaler Ebene durch anthropogene Emissionen, mit Folgeerscheinungen wie Verschiebung der Klimazonen, zunehmendes Extremwetter, Erwärmung und Versauerung der Ozeane[154]
- Gefährdung der Biodiversität in den Meeren u. a. durch Fischerei, Offshorebauwerke, Schifffahrt, Marikultur und Meeresbodenbergbau[155]
- Schadstoff-Einträge in marine Systeme: Abwässer aus Industrie und Landwirtschaft, Verklappung von Schweröl, diverse Ölpest-Ereignisse, halogenierte und chlorierte Kohlenwasserstoffe, Schwermetalle wie Blei, Cadmium oder Quecksilber, Atommüll oder radioaktiv kontaminierte Abwässer, feste Abfallstoffe (hauptsächlich in Form von Kunststoffen, darunter auch signifikante Mengen an Nanopartikeln)
- Einwanderung von Neophyten, mit möglicher Verdrängung autochthoner (einheimischer) Arten bei Flora und Fauna
- Virusinfektionen[156]
- Der Straßenverkehr, die Wirkung von Lichtverschmutzung auf lebende Organismen und die Verunruhigung der Landschaft etwa durch deren Nutzung als Naherholungsgebiete stellen weitere wichtige Faktoren dar.[148]
Massenaussterben und Evolution
Die Entstehung neuer Arten
Einem Massenaussterben folgte für gewöhnlich eine beschleunigte Phase der Adaptiven Radiation durch die überlebenden Arten, die im Zuge ihrer Vermehrung jene Habitate besiedelten, die zuvor von konkurrierenden Lebensformen besetzt waren. Die Geschwindigkeit dieser Ausbreitung hing in hohem Maße von den geophysikalischen Rahmenbedingungen und damit von der Nutzbarkeit der verfügbaren Biotope ab. Während zum Beispiel die Säugetiere nach dem Umbruch an der Kreide-Paläogen-Grenze von der relativ zügig verlaufenden Regeneration der natürlichen Systeme profitierten und im Paläozän im Zeitraum von 0,4 bis 1,0 Millionen Jahre nach dem Aussterbe-Ereignis eine erste Zunahme der Biodiversität und damit die Bildung neuer Arten verzeichneten,[157] vollzog sich die Normalisierung im Anschluss an das Massenaussterben am Perm-Trias-Übergang nur schleppend. Bedingt durch die anhaltende Umweltkrise mit zahlreichen Folgeerscheinungen dauerte es zum Teil 10 bis 15 Millionen Jahre, bis sich ein stabiles ökologisches Gleichgewicht eingestellt hatte.[84][158] In dieser Übergangszeit okkupierten mitunter einige robustere Spezies unter massiver Vermehrung viele verwaiste Lebensräume („Katastrophentaxa“), ehe sie durch den Konkurrenzdruck einwandernder Populationen zurückgedrängt wurden.[159] Eine ähnlich verzögerte Regeneration könnte auch in der (allerdings umstrittenen) Fossilienlücke des frühen Karbons aufgetreten sein („Romers’s gap“), die möglicherweise eine Spätfolge des vorhergehenden Hangenberg-Ereignisses war.[160]
Aussterbe- und Überlebensmuster am Beispiel des Chicxulub-Einschlags

Das späte Maastrichtium, die letzte Stufe der Kreide, wies neben einer deutlichen Abkühlungstendenz mehrere kurzzeitige und stark ausgeprägte Klimaschwankungen auf, die wahrscheinlich auf den lang anhaltenden Flutbasalt-Vulkanismus des Dekkan-Trapp im heutigen Westindien zurückgehen. Demzufolge wurde in der Wissenschaft über Jahrzehnte die Ansicht vertreten – scheinbar gestützt durch den Fossilbericht –, dass sich viele Gattungen der mesozoischen Fauna lange vor ihrem endgültigen Verschwinden „auf dem absteigenden Ast“ befanden.[161] Neuere Studien entwerfen hingegen ein differenziertes Bild der damaligen Ereignisse. Insgesamt liegen keine eindeutigen Beweise vor, dass Dinosaurier und andere Tiergruppen bereits vor dem drastischen Einschnitt an der Kreide-Paläogen-Grenze von einem langfristigen Rückgang betroffen waren.[162][163] Dieser Befund trifft auch für die Pterosauria (Flugsaurier) zu, deren Artenvielfalt während der obersten Kreide offenbar stabil blieb und die bis zu ihrem abrupten Aussterben möglicherweise eine Diversitätszunahme verzeichnen konnten.[164] Somit deutet Vieles darauf hin, dass die biologische Krise am Ende des Mesozoikums ausschließlich durch den Chicxulub-Einschlag verursacht wurde.[97][103]
Das Impakt-Ereignis vor 66 Millionen Jahren bewirkte eine Reihe gravierender Umweltveränderungen, die in gestaffelter zeitlicher Abfolge auftraten und eine weltweite Katastrophe auslösten, der alle Nichtvogel-Dinosaurier sowie ein großer Teil der übrigen Fauna zum Opfer fielen.[26] Bereits in der ersten Stunde nach dem Einschlag kam es auch in weiterem Umkreis des Epizentrums durch extrem starke Erdbeben sowie durch eine überschallschnelle Druck- und Hitzewelle in Verbindung mit einem schauerartigen Tektitregen zu einem Massensterben der Land- und Wasserfauna.[165] Sich unmittelbar daran anschließende Ereignisse waren ein Megatsunami[166] und die Entstehung globaler Flächenbrände durch den weiträumig verteilten Auswurf glühender Gesteinstrümmer (Ejecta). Im nächsten Stadium bildete sich innerhalb weniger Tage eine die Erde umhüllende Staub- und Rußwolke, die das Sonnenlicht absorbierte, die pflanzliche Photosynthese weitgehend unterband und den Beginn eines rasch einsetzenden Impaktwinters markierte. Daraus resultierte eine starke Dezimierung der Landvegetation mit entsprechender Rückwirkung auf große Pflanzenfresser.
Die vollständige Auslöschung der relativ artenarmen Dinosaurier-Population beruhte vermutlich auf einer Kombination verschiedener Faktoren. Bedingt durch die Größe der einzelnen Individuen gab es keine Möglichkeit, den Primärfolgen des Asteroideneinschlags zu entkommen und – anders als etwa Säugetiere – in natürliche Schutzräume wie Erdhöhlen zu fliehen.[98] Als überwiegend endotherme (warmblütige) Tiere benötigten Dinosaurier zur Aufrechterhaltung ihrer Körpertemperatur und ihrer Vitalfunktionen ein ausreichendes Nahrungsangebot, das jedoch im Verlauf der Impaktkatastrophe deutlich schrumpfte. Im Wesentlichen gliederte sich die Gruppe der Dinosaurier in Herbivoren und Karnivoren, ob sich darunter auch Omnivoren (Allesfresser) befanden, wie eventuell bei den Ornithomimosauria, ist gegenwärtig nicht belegt. Von den Säugern scheinen keine rein pflanzen- oder fleischfressenden Formen das Ereignis überlebt zu haben. Hingegen konnten sich Omnivoren und Insektenfresser einigermaßen behaupten, da sie Kerbtiere, Würmer und Schnecken als Nahrungsquelle nutzten.[157] Das untere Ende der Nahrungskette wurde vom sogenannten Detritus gebildet, eine noch nicht humifizierte organische Substanz im oder auf dem Erdboden, bestehend aus abgestorbenem Pflanzengewebe, Pollen, Aas, Mikroorganismen und anderem organischen Material. Es wird angenommen, dass die Detritus-Depots an der Kreide-Paläogen-Grenze keine signifikanten Einbußen erlitten und somit die Existenz vieler kleinerer Lebewesen sicherten.
Mit am schwersten geschädigt wurden Flora und Fauna im heutigen Nordamerika. Laut einer statistischen Untersuchung starben in dieser Region fast 60 Prozent aller Pflanzenarten aus.[167] Weniger auffällig gestaltete sich die Florenreduzierung in der südlichen Hemisphäre. Dennoch kam es hier wie überall zu einer Ausbreitung von Pilzen, Moosen und Flechten und nach einiger Zeit zu einer Wachstumsspitze bei Farngewächsen. Dass die Vegetationskrise nur von vorübergehender Dauer war, ist wahrscheinlich der bei vielen höheren Pflanzen auftretenden Polyploidie geschuldet. Polyploidie bezeichnet das genetische Merkmal, wenn ein Organismus mehrere Chromosomensätze in einer Zelle besitzt. Diese Eigenschaft führt bei Pflanzen oftmals zu erhöhter Vitalität und zur besseren Anpassung an sich verschlechternde Klimabedingungen und könnte zum Überleben im drastisch veränderten Umfeld der Post-Impakt-Ära beigetragen haben.[168]

Neben den kreidezeitlichen Vögeln, von denen sich nur wenige Arten in das Paläogen retteten,[101] verzeichneten auch Eidechsen und Schlangen erhebliche Verluste.[169] Da kein Landtier mit mehr als 15 bis 20 kg Körpergewicht die Krisenzeit überstand, war Kleinwüchsigkeit generell von Vorteil. Krokodile stellten in dieser Hinsicht eine gewisse Ausnahme dar. Zwar starben ihre größten Vertreter in Form von Meeresreptilien ebenfalls aus, doch konnten sie als ektotherme (wechselwarme) Lebewesen sehr lange auf Nahrung verzichten, aufgrund ihrer semiaquatischen Lebensweise in Flüssen und Seen die gröbsten Beeinträchtigungen vermeiden und so den Artenschwund auf rund 50 Prozent begrenzen.[170] Das lässt die Vermutung zu, dass Süßwasser-Biotope unter den ökologischen Nischen zu den vergleichsweise sicheren Lebensbereichen zählten.
Im Gegensatz dazu begann das Massenaussterben in den Ozeanen mit leichter Zeitverzögerung, dann aber mit gravierenden Folgen. Ursächlich daran beteiligt waren rasch auftretende geochemische Veränderungen, hervorgerufen durch den massiven Eintrag von Schadstoffen wie Stickoxiden und Schwefeldioxid beziehungsweise der daraus entstandenen Schwefelsäure.[171] Diese Prozesse gingen vor allem zu Lasten Kalkschalen bildender Organismen, deren Aussterberaten bei Ammoniten 100 Prozent betrug und bei Foraminiferen und verschiedenen Planktonarten 90 Prozent überschritt. Dies deutet auf eine starke Versauerung der ozeanischen Bereiche über mehrere 10.000 Jahre hin, und vermutlich dauerte es über eine Million Jahre, bis der nachhaltig gestörte ozeanische Gleichgewichtszustand in vollem Umfang wiederhergestellt war.[97][172] Fest steht, dass mit der weitgehenden Reduzierung des Nannoplanktons viele davon abhängige Lebensformen ihre Nahrungsgrundlage einbüßten, was sich als Kettenreaktion auf das gesamte marine Ökosystem und in letzter Konsequenz auch auf die darin heimische Megafauna auswirkte. Somit erreichte das Massenaussterben in den Meeren ähnliche Dimensionen wie das auf dem Festland.
Der über mehrere Jahrzehnte herrschende Impaktwinter gilt als eine der Hauptursachen des damaligen Massenaussterbens. Über das Ausmaß des Kälteeinbruchs gibt es in der Wissenschaft unterschiedliche Annahmen. Während einerseits ein Temperatursturz um 26 °C mit globalem Dauerfrost durch eine atmosphärische Schicht von Schwefelsäure-Aerosolen postuliert wird,[100] fand laut einer anderen Studie eine vergleichsweise moderate und zeitlich begrenzte Abkühlung statt.[173] Hingegen besteht Einigkeit darüber, dass offenbar die Erholung der südhemisphärischen Biotope bedeutend rascher vonstatten ging als die der entsprechenden Regionen auf der nördlichen Halbkugel.[174] Nach dem Ende der Kältephase trat die Erde in das Stadium einer globalen Erwärmung, zum Teil bedingt durch Milliarden Tonnen Kohlenstoffdioxid, die der Chicxulub-Einschlag infolge der Verdampfung ozeanischer Böden freigesetzt hatte, zu einem größeren Teil verursacht von den erhöhten CO2-Ausgasungen des Dekkan-Trapps, möglicherweise initiiert durch die tektonische Erschütterung des Asteroideneinschlags.[175] Welchen Anteil das rapide einsetzende und mindestens 50.000 Jahre währende Treibhausklima am Massenaussterben hatte, ist im Detail noch nicht hinreichend geklärt.
Die Autoren einer Ende 2017 veröffentlichten Studie kommen zu dem Ergebnis, dass der Asteroid in Gesteinsschichten einschlug, die hohe Konzentrationen an Kohlenwasserstoffen und Sulfaten enthielten. Durch die großflächige Erhitzung und Verdampfung der ozeanischen Sedimente in dieser Region verteilten sich umfangreiche Mengen an Ruß- und Schwefelaerosolen in der Stratosphäre, wodurch sich die klimatischen Auswirkungen des folgenden Impaktwinters im Hinblick auf dessen Dauer und Ausmaß erheblich verstärkten. Hätte der Asteroid ein Gebiet mit geringeren Kohlenwasserstoff-Anteilen getroffen (etwa 87 Prozent der Erdoberfläche), wäre die biologische Krise wahrscheinlich weitaus glimpflicher verlaufen, mit einer deutlich höheren Überlebensrate der mesozoischen Fauna.[176] Eine 2020 publizierte Studie postulierte anhand von Computersimulationen, dass der bei 45 bis 60 Grad liegende wahrscheinliche Aufprallwinkel des Asteroiden von allen Einschlagszenarien die verheerendsten Auswirkungen zeitigte.[177]
Siehe auch
Literatur
Englischsprachige Werke
- Ashraf Elewa (Hrsg.): Mass Extinction. Springer, Berlin u. a. 2008, ISBN 978-3-540-75915-7.
- George R. McGhee Jr: When the Invasion of Land Failed. The Legacy of the Devonian Extinctions. Columbia University Press, New York 2013, ISBN 978-0-231-16057-5 (Darstellung verschiedener biologischer Krisen in Verbindung mit den devonischen Massenaussterben).
- George R. McGhee Jr.: Carboniferous Giants and Mass Extinction. The Late Paleozoic Ice Age World. Columbia University Press, New York 2018, ISBN 978-0-231-18097-9 (Beschreibung von Flora und Fauna während des Permokarbonen Eiszeitalters mit Schwerpunkt auf dem Karbon).
Deutschsprachige Werke
- Matthias Glaubrecht: Das Ende der Evolution: der Mensch und die Vernichtung der Arten. C. Bertelsmann, München 2019, ISBN 978-3-570-10241-1.
- Peter Ward, Joe Kirschvink: Eine neue Geschichte des Lebens. Wie Katastrophen den Lauf der Evolution bestimmt haben. Deutsche Verlags Anstalt, München 2016, ISBN 978-3-421-04661-1.
- Norman MacLeod: Arten Sterben – Wendepunkte der Evolution. Theiss Verlag – Wissenschaftliche Buchgesellschaft, Darmstadt 2016, ISBN 978-3-8062-3284-4 (populärwissenschaftliche Darstellung eines Paläontologen zum gesamten Themenkomplex).
- Jens Boenigk, Sabina Wodniok: Biodiversität und Erdgeschichte. Springer Verlag, Berlin – Heidelberg 2014 (Springer Spektrum), DOIː 10.1007/978-3-642-55389-9, ISBN 978-3-642-55388-2 (Lehrbuch u. a. über die Entstehung von Diversität im erdgeschichtlichen Kontext).
- Elizabeth Kolbert: Das sechste Sterben. Wie der Mensch Naturgeschichte schreibt. Aus dem Englischen übersetzt von Ulrike Bischoff. Suhrkamp, Berlin 2015, ISBN 3-515-08518-1.
- Ted Nield: Superkontinent. Das geheime Leben unseres Planeten: Eine abenteuerliche Reise durch die Erdgeschichte. Verlag Antje Kunstmann, München 2008, ISBN 978-3-88897-526-4.
- József Pálfy: Katastrophen der Erdgeschichte. Globales Artensterben? Schweizerbart, Stuttgart 2005, ISBN 3-510-65211-8.
- David M. Raup: The Nemesis Affair. A Story of the Death of Dinosaurs and the Ways of Science, W. W. Norton 1986, deutsche Ausgabe: Der schwarze Stern. Wie die Saurier starben – der Streit um die Nemesis-Hypothese. Rowohlt, Reinbek 1990, ISBN 3-498-05720-0 (zur Nemesis-Hypothese einer periodischen extraterrestrischen Ursache, die in den 1980er-Jahren populär war).
Weblinks
- astris.de: Die Geschichte der Erde – Massensterben
- scinexx.de: Massenaussterben
- oekosystem-erde.de: Massenaussterben
- Peter D. Ward: Tod aus der Tiefe. In: Spektrum der Wissenschaft, 3/2007, S. 26–33.
Einzelnachweise
- Richard J. Twitchett: The palaeoclimatology, palaeoecology and palaeoenvironmental analysis of mass extinction events. (PDF) In: Palaeogeography, Palaeoclimatology, Palaeoecology. 232, Nr. 2–4, März 2006, S. 190–213. doi:10.1016/j.palaeo.2005.05.019.
- Der Artenverlust kann nicht nur durch eine erhöhten Aussterberate, sondern auch eine geringe Artbildungsrate mitverursacht werden, siehe Bond und Grasby (2017), S. 8, und Anthony D. Barnosky, Nicholas Matzke, Susumu Tomiya, Guinevere O. U. Wogan, Brian Swartz, Tiago B. Quental, Charles Marshall, Jenny L. McGuire, Emily L. Lindsey, Kaitlin C. Maguire, Ben Mersey, Elizabeth A. Ferrer: Has the Earth’s sixth mass extinction already arrived?. In: Nature. 471, Nr. 7336, März 2011, S. 51–52.
- Anthony D. Barnosky, Nicholas Matzke, Susumu Tomiya, Guinevere O. U. Wogan, Brian Swartz, Tiago B. Quental, Charles Marshall, Jenny L. McGuire, Emily L. Lindsey, Kaitlin C. Maguire, Ben Mersey, Elizabeth A. Ferrer: Has the Earth’s sixth mass extinction already arrived?. In: Nature. 471, Nr. 7336, März 2011, S. 51–57. doi:10.1038/nature09678.
- Richard K. Bambach: Phanerozoic biodiversity mass extinctions. In: Annual Review of Earth and Planetary Sciences. 34, Mai 2006, S. 127–155. doi:10.1146/annurev.earth.33.092203.122654.
- L. W. Alvarez, W. Alvarez, F. Asaro, H. W. Michel: Extraterrestrial Cause for the Cretaceous-Tertiary Extinction. (PDF) In: Science. 208, Nr. 4448, Juni 1980, S. 1095–1108.
- David M. Raup: Der schwarze Stern. Wie die Saurier starben – der Streit um die Nemesis-Hypothese. Rowohlt, Reinbek 1990, ISBN 3-498-05720-0, S. 73.
- David P. G. Bond, Stephen E. Grasby: On the causes of mass extinctions. (PDF) In: Palaeogeography, Palaeoclimatology, Palaeoecology. 478, Nr. 15, Juli 2017, S. 3–29. doi:10.1016/j.palaeo.2016.11.005.
- Alycia L. Stigall: Speciation collapse and invasive species dynamics during the Late Devonian “Mass Extinction”. In: GSA Today (Geological Society of America). 22, Nr. 1, Januar 2012, S. 4–9. doi:10.1130/G128A.1.
- Ulf von Rauchhaupt: Fünfmal ging die Welt schon unter. In: FAZ.NET
- Paul B. Wignall: The Worst of Times. Princeton University Press, 2016, ISBN 978-0-691-17602-4.
- Ben G. Mason, David M. Pyle, Clive Oppenheimer: The size and frequency of the largest explosive eruptions on Earth. (PDF) In: Bulletin of Volcanology. 66, Nr. 8, Dezember 2004, S. 735–748. doi:10.1007/s00445-004-0355-9.
- Robert B. Smith, Lawrence W. Braile: Crustal Structure and Evolution of an Explosive Silicic Volcanic System at Yellowstone National Park In Geology of Yellowstone Park Area; 33rd Annual Field Conference Guidebook, 1982, S. 233–250.
- David P. G. Bond, Paul B. Wignall: Large igneous provinces and mass extinctions: An update. (PDF) In: The Geological Society of America (GSA) Special Paper. 505, September 2014, S. 29–55. doi:10.1130/2014.2505(02).
- Daniel H. Rothman, Gregory P. Fournier, Katherine L. French, Eric J. Alm, Edward A. Boyle, Changqun Cao, Roger E. Summons: Methanogenic burst in the end-Permian carbon cycle. In: PNAS. 111, Nr. 15, April 2014, S. 5462–5467. doi:10.1073/pnas.1318106111.
- Mark A. Richards, Walter Alvarez, Stephen Self, Leif Karlstrom, Paul R. Renne, Michael Manga, Courtney J. Sprain, Jan Smit, Loÿc Vanderkluysen, Sally A. Gibson: Triggering of the largest Deccan eruptions by the Chicxulub impact. (PDF) In: Geological Society of America Bulletin. April 2015. doi:10.1130/B31167.1.
- Jennifer Kasbohm, Blair Schoene: Rapid eruption of the Columbia River flood basalt and correlation with the mid-Miocene climate optimum. (PDF) In: Science Advances. 4, Nr. 9, September 2018. doi:10.1126/sciadv.aat8223.
- Marten Scheffer, Victor Brovkin, Peter M. Cox: Positive feedback between global warming and atmospheric CO2 concentration inferred from past climate change. In: Geophysical Research Letters. 33, Nr. 10, Mai 2006, S. L10702 1–4. doi:10.1029/2005GL025044.
- David L. Kidder, Thomas R. Worsley: A human-induced hothouse climate?. (PDF) In: GSA Today (The Geological Society of America). 22, Nr. 2, Februar 2012, S. 4–11. doi:10.1130/G131A.1.
- Timothy M. Lenton, Hermann Held, Elmar Kriegler, Jim W. Hall, Wolfgang Lucht, Stefan Rahmstorf, Hans Joachim Schellnhuber: Tipping elements in the Earth’s climate system. In: PNAS. 105, Nr. 6, 2008, S. 1786–1793. doi:10.1073/pnas.0705414105.
- Vincent E. Courtillot, Paul R. Renne: Time-correlation of mass-extinctions and flood-basalt-Events. (PDF) In: Comptes Rendus Geoscience. 335, Nr. 1, Januar 2003, S. 113–140. doi:10.1016/S1631-0713(03)00006-3.
- Vincent E. Courtillot, Paul R.Renne: On the ages of flood basalt events Comptes Rendus Geoscience 335, 2003
- Katja M. Meyer, Lee R. Kump: Oceanic Euxinia in Earth History: Causes and Consequences. (PDF) In: Annual Review of Earth and Planetary Sciences. 36, Mai 2008, S. 251–288. doi:10.1146/annurev.earth.36.031207.124256.
- Gregory A. Brennecka, Achim D. Herrmann, Thomas J. Algeo, Ariel D. Anbar: Rapid expansion of oceanic anoxia immediately before the end-Permian mass extinction. In: PNAS. 108, Nr. 43, Oktober 2011, S. 17631–17634. doi:10.1073/pnas.1106039108.
- Alan R. Hildebrand, Glen T. Penfield, David A. Kring, Mark Pilkington, Antonio Camargo, Stein B. Jacobsen, William V. Boynton: Chicxulub Crater: A possible Cretaceous/Tertiary boundary impact crater on the Yucatán Peninsula, Mexico. (PDF) In: Geology. 19, Nr. 9, September 1991, S. 867–871.
- Peter Ward, Joe Kirschvink: Eine neue Geschichte des Lebens. Wie Katastrophen den Lauf der Evolution bestimmt haben. Deutsche Verlags-Anstalt, München 2016, ISBN 978-3-421-04661-1, S. 36.
- Christian Koeberl: Massensterben und Impaktereignisse in der Erdgeschichte: Ein kurzer Überblick. Ursprünglich in: Jahrbuch der Geologischen Bundesanstalt (Österreich), Band 147/Heft 1+2, Festschrift zum 65. Geburtstag von HR Univ.-Prof. Dr. Hans Peter Schönlaub, Direktor der Geologischen Bundesanstalt (zobodat.at [PDF]).
- A. Y. Glikson, A. J. Meixner, B. Radke, I. T. Uysal, E. Saygin, J. Vickers, T. P. Mernagh: Geophysical anomalies and quartz deformation of the Warburton West structure, central Australia. (PDF) In: Tectonophysics. 643, März 2015, S. 55–72. doi:10.1016/j.tecto.2014.12.010.
- A. L. Melott, B. S. Lieberman, C. M. Laird, L. D. Martin, M. V. Medvedev, B. C. Thomas, J. K. Cannizzo, N. Gehrels, C. H. Jackman: Did a gamma-ray burst initiate the late Ordovician mass extinction?. (PDF) In: International Journal of Astrobiology. 3, Nr. 1, April 2004, S. 55–61. doi:10.1017/S1473550404001910.
- K. Z. Stanek, O. Y. Gnedin, J. F. Beacom, A. P. Gould, J. A. Johnson, J. A. Kollmeier, M. Modjaz, M. H. Pinsonneault, R. Pogge, D. H. Weinberg: Protecting Life in the Milky Way: Metals Keep the GRBs Away. (PDF) In: Acta Astronomica. 56, Dezember 2006, S. 333–345. arxiv:astro-ph/0604113.
- K. Knie, G. Korschinek, T. Faestermann, E. A. Dorfi, G. Rugel, A. Wallner: 60Fe Anomaly in a Deep-Sea Manganese Crust and Implications for a Nearby Supernova Source. (PDF) In: Physical Review Letters. 93, Nr. 17, Oktober 2004, S. 171103-1–171103-4. doi:10.1103/PhysRevLett.93.171103.
- A. Wallner, J. Feige, N. Kinoshita, M. Paul, L. K. Fifield, R. Golser, M. Honda, U. Linnemann, H. Matsuzaki, S. Merchel, G. Rugel, S. G. Tims, P. Steier, T. Yamagata, S. R. Winkler: Recent near-Earth supernovae probed by global deposition of interstellar radioactive 60Fe. In: Nature. 532, Nr. 7597, April 2016, S. 69–72. doi:10.1038/nature17196.
- David M. Raup, J. John Sepkoski: Periodicity of extinctions in the geologic past. (PDF) In: Proceedings of the National Academy of Sciences (PNAS). 81, Nr. 4539, 1984, S. 801–805.
- Marc Davis, Piet Hut, Richard A. Muller: Extinction of species by periodic comet showers. (PDF) In: Nature. 308, April 1984, S. 715–717. doi:10.1038/308715a0.
- Matthias M. M. Meier, Sanna Holm-Alwmark: A tale of clusters: no resolvable periodicity in the terrestrial impact cratering record. In: Monthly Notices of the Royal Astronomical Society. 467, Nr. 3, Juni 2017, S. 2545–2551. doi:10.1093/mnras/stx211.
- Heather D. Graven: Impact of fossil fuel emissions on atmospheric radiocarbon and various applications of radiocarbon over this century. In: pnas. 112, Nr. 31, Juli 2015, S. 9542–9545. doi:10.1073/pnas.1504467112.
- Melanie J. Leng, Jim D. Marshall: Palaeoclimate interpretation of stable isotope data from lake sediment archives. In: Quaternary Science Reviews. Band 23, Nr. 7–8, April 2004, S. 811–831, doi:10.1016/j.quascirev.2003.06.012.
- Jung-Hyun Kim, Stefan Schouten, Ellen C. Hopmans, Barbara Donner, Jaap S. Sinninghe Damsté: Global sediment core-top calibration of the TEX86 paleothermometer in the ocean. (PDF) In: Geochimica et Cosmochimica Acta. 72, Nr. 4, Februar 2008, S. 1154–1173. doi:10.1016/j.gca.2007.12.010.
- K. Panchuk, A. Ridgwell, L. R. Kump: Sedimentary response to Paleocene-Eocene Thermal Maximum carbon release: A model-data comparison. In: Geology. 36, Nr. 4, April 2008, S. 315–318. doi:10.1130/G24474A.1.
- Heinrich D. Holland: The oxygenation of the atmosphere and oceans. (PDF) In: Philosophical Transactions of Royal Society B. 361, Nr. 1470, Juni 2006, S. 903–915. doi:10.1098/rstb.2006.1838.
- Robert E. Kopp, Joseph L. Kirschvink, Isaac A. Hilburn, Cody Z. Nash: The Paleoproterozoic snowball Earth: A climate disaster triggered by the evolution of oxygenic photosynthesis. In: PNAS. 102, Nr. 32, 2005, S. 11131–11136. doi:10.1073/pnas.0504878102.
- Alan D. Rooney, Justin V. Strauss, Alan D. Brandon, Francis A. Macdonald: A Cryogenian chronology: Two long-lasting synchronous Neoproterozoic glaciations. (PDF) In: Geology. 43, Nr. 5, Mai 2015, S. 459–462. doi:10.1130/G36511.1.
- Frank A. Corsetti, Stanley M. Awramik, David Pierce: A complex microbiota from snowball Earth times: Microfossils from the Neoproterozoic Kingston Peak Formation, Death Valley, USA. In: PNAS. 100, Nr. 8, April 2003, S. 4399–4404. doi:10.1073/pnas.0730560100.
- Marc Laflamme, Simon A. F. Darroch, Sarah M. Tweedt, Kevin J. Peterson, Douglas H. Erwin: The end of the Ediacara biota: Extinction, biotic replacement, or Cheshire Cat?. (PDF) In: Gondwana Research. 23, Nr. 2, März 2013, S. 558–573. doi:10.1016/j.gr.2012.11.004.
- Richard J. Squire, Ian H. Campbell, Charlotte M. Allen, Christopher J. L. Wilson: Did the Transgondwanan Supermountain trigger the explosive radiation of animals on Earth?. (PDF) In: Earth and Planetary Science Letters. 250, Nr. 1–2, Oktober 2006, S. 116–133. doi:10.1016/j.epsl.2006.07.
- Lawrence M. Och, Graham A. Shields-Zhou, Simon W. Poulton, Christina Manning, Matthew F. Thirlwall, Da Li, Xi Chen, Hongfei Ling, Tony Osborn, Lorenzo Cremonese: Redox changes in Early Cambrian black shales at Xiaotan section, Yunnan Province, South China. (PDF) In: Precambrian Research. 225, Februar 2013, S. 166–189.
- Tomoko Ishikawa, Yuichiro Ueno, Degan Shu, Yong Li, Jian Han, Junfeng Guo, Naohiro Yoshida, Tsuyoshi Komiya: Irreversible change of the oceanic carbon cycle in the earliest Cambrian: High-resolution organic and inorganic carbon chemostratigraphy in the Three Gorges area, South China. (PDF) In: Precambrian Research. 225, Februar 2013, S. 190–208. doi:10.1016/j.precamres.2011.10.004.
- Benjamin C. Gill, Timothy W. Lyons, Seth A. Young, Lee R. Kump, Andrew H. Knoll, Matthew R. Saltzman: Geochemical evidence for widespread euxinia in the Later Cambrian ocean. (PDF) In: Nature. 469, Nr. 7328, Januar 2011, S. 80–83. doi:10.1038/nature09700.
- F. Jourdan, K. Hodges, B. Sell, U. Schaltegger, M. T. D. Wingate, L. Z. Evins, U. Söderlund, P. W. Haines, D. Phillips, T. Blenkinsop: High-precision dating of the Kalkarindji large igneous province, Australia, and synchrony with the Early-Middle Cambrian (Stage 4–5) extinction. (PDF) In: Geology. 42, Nr. 6, Juni 2014, S. 543–546. doi:10.1130/G35434.1.
- Jennifer L. Morris, Mark N. Puttick, James W. Clark, Dianne Edwards, Paul Kenrick, Silvia Pressel, Charles H. Wellman, Ziheng Yang, Harald Schneider, Philip C. J. Donoghue: The timescale of early land plant evolution. In: PNAS. Februar 2018. doi:10.1073/pnas.1719588115.
- Timothy M. Lenton, Michael Crouch, Martin Johnson, Nuno Pires, Liam Dolan: First plants cooled the Ordovician. (PDF) In: Nature Geoscience. 5, Februar 2012, S. 86–89. doi:10.1038/ngeo1390.
- Pascale F. Poussart, Andrew J. Weaver, Christopher R. Barne: Late Ordovician glaciation under high atmospheric CO2: A coupled model analysis. (PDF) In: Paleoceanography. 14, Nr. 4, August 1999, S. 542–558.
- Thijs R. A. Vandenbroucke, Poul Emsbo, Axel Munnecke, Nicolas Nuns, Ludovic Duponchel, Kevin Lepot, Melesio Quijada, Florentin Paris, Thomas Servais, Wolfgang Kiessling: Metal-induced malformations in early Palaeozoic plankton are harbingers of mass extinctions. In: Nature Communications. 6, August 2015. doi:10.1038/ncomms8966.
- John A. Long, Ross R. Large, Michael S. Y. Lee, Michael J. Benton, Leonid V. Danyushevsky, Luis M. Chiappe, Jacqueline A. Halpin, David Cantrill, Bernd Lottermoser: Severe selenium depletion in the Phanerozoic oceans as a factor in three global mass extinction events. (PDF) In: Gondwana Research. 36, August 2016, S. 209–218. doi:10.1016/j.gr.2015.10.001.
- David M. Raup, J. John Sepkoski: Mass Extinctions in the Marine Fossil Record. (PDF) In: Science. 215, Nr. 4539, März 1982, S. 1501–1503.
- R. T. Becker, P. Königshof, C. E. Brett: Devonian climate, sea level and evolutionary events: an introduction. (PDF) In: Geological Society, London, Special Publications. 423, August 2016, S. 1–10. doi:10.1144/SP423.15.
- „Middle to Upper Devonian biotic crisis“, siehe Thomas J. Algeo, Stephen E. Scheckler: Terrestrial-marine teleconnections in the Devonian: links between the evolution of land plants, weathering processes, and marine anoxic events. In: Philosophical Transactions of the Royal Society of London B (Biological Sciences). 353, Nr. 1365, 1998, S. 113–130, doi:10.1098/rstb.1998.0195, PMC 1692181 (freier Volltext)
- Marina Kloppischː Organisch-geochemischer Vergleich ausgewählter Gesteine der Frasnium/Famennium Grenze (Oberdevon) im Bergischen Land und der Eifel (PDF; 5,0 MB). Berichte des Forschungszentrums Jülich, Institut für Chemie und Dynamik der Geosphäre, 2002.
- David De Vleeschouwer, Micha Rakociński, Grzegorz Racki, David P. G. Bond, Katarzyna Sobień, Philippe Claeys: The astronomical rhythm of Late-Devonian climate change (Kowala section, Holy Cross Mountains, Poland). (PDF) In: Earth and Planetary Science Letters. 365, März 2013, S. 25–37. doi:10.1016/j.epsl.2013.01.016.
- Sarah K. Carmichael, Johnny A. Waters, Cameron J. Batchelor, Drew M. Coleman, Thomas J. Suttner, Erika Kido, L. M. Moore, Leona Chadimová: Climate instability and tipping points in the Late Devonian: Detection of the Hangenberg Event in an open oceanic island arc in the Central Asian Orogenic Belt. (PDF) In: Gondwana Research. 32, April 2016, S. 213–231. doi:10.1016/j.gr.2015.02.009.
- Brian D. Fields, Adrian L. Melott, John Ellis, Adrienne F. Ertel, Brian J. Fry, Bruce S. Lieberman, Zhenghai Liu, Jesse A. Miller, Brian C. Thomas: Supernova triggers for end-Devonian extinctions. In: PNAS. August 2020. doi:10.1073/pnas.2013774117.
- Sandra Isabella Kaiser, Ralf Thomas Becker, Thomas Steuber, Sarah Zhor Aboussalam: Climate-controlled mass extinctions, facies, and sea-level changes around the Devonian–Carboniferous boundary in the eastern Anti-Atlas (SE Morocco). (PDF) In: Palaeogeography, Palaeoclimatology, Palaeoecology. 310, Nr. 3–4, Oktober 2011, S. 340–364. doi:10.1016/j.palaeo.2011.07.026.
- Grzegorz Racki, Michał Rakociński, Leszek Marynowski, Paul B. Wignall: Mercury enrichments and the Frasnian-Famennian biotic crisis: A volcanic trigger proved?. (PDF) In: Geology. 46, Nr. 6, Juni 2018, S. 543–546. doi:10.1130/G40233.1.
- J. Ricci, X. Quidelleur, V. Pavlov, S. Orlov, A. Shatsillo, V. Courtillot: New 40Ar/39Ar and K–Ar ages of the Viluy traps (Eastern Siberia): Further evidence for a relationship with the Frasnian–Famennian mass extinction. (PDF) In: Palaeogeography, Palaeoclimatology, Palaeoecology. 386, September 2013, S. 531–540. doi:10.1016/j.palaeo.2013.06.020.
- Leszek Marynowski, Michał Zatoń, Michał Rakociński, Paweł Filipiak, Slawomir Kurkiewicz, Tim J. Pearce: Deciphering the upper Famennian Hangenberg Black Shale depositional environments based on multi-proxy record. (PDF) In: Palaeogeography, Palaeoclimatology, Palaeoecology. 346–347, August 2012, S. 66–86. doi:10.1016/j.palaeo.2012.05.020.
- Sandra Isabella Kaiser, Markus Aretz, Ralph Thomas Becker: The global Hangenberg Crisis (Devonian–Carboniferous transition): review of a first-order mass extinction. (PDF) In: Geological Society, London, Special Publications. 423, August 2016, S. 387–437.
- Paul M. Myrow, Jahandar Ramezani, Anne E. Hanson, Samuel A. Bowring, Grzegorz Racki, Michał Rakociński: High-precision U–Pb age and duration of the latest Devonian (Famennian) Hangenberg event, and its implications. (PDF) In: Terra Nova. 26, Nr. 3, Juni 2014, S. 222–229. doi:10.1111/ter.12090.
- Andrew J. Retzler, Leif Tapanila, Julia R. Steenberg, Carrie J. Johnson, Reed A. Myers: Post-impact depositional environments as a proxy for crater morphology, Late Devonian Alamo impact, Nevada. (PDF) In: Geosphere (Geological Society of America). 11, Nr. 1, Januar 2015, S. 123–143. doi:10.1130/GES00964.1.
- David P. G. Bond, Paul B. Wignall, Michael M. Joachimski, Yadong Sun, Ivan Savov, Stephen E. Grasby, Benoit Beauchamp, Dierk P. G. Blomeier: An abrupt extinction in the Middle Permian (Capitanian) of the Boreal Realm (Spitsbergen) and its link to anoxia and acidification. (PDF) In: GSA Bulletin (Geological Society of America). 127, Nr. 9–10, September 2015, S. 1411–1421. doi:10.1130/B31216.1.
- He Bin, Yi-Gang Xu, Xiao-Long Huang, Zhen-Yu Luo, Yu-Ruo Shi, Qi-Jun Yang, Song-Yue Yu: Age and duration of the Emeishan flood volcanism, SW China: Geochemistry and SHRIMP zircon U-Pb dating of silicic ignimbrites, post-volcanic Xuanwei Formation and clay tuff at the Chaotian section. (PDF) In: Earth and Planetary Science Letters. 255, März 2007, S. 306–323. doi:10.1016/j.epsl.2006.12.021.
- Kévin Reya, Michael O. Day, Romain Amiot, Jean Goedert, Christophe Lécuyer, Judith Sealy, Bruce S. Rubidge: Stable isotope record implicates aridification without warming during the late Capitanian mass extinction. In: Gondwana Research. 59, Juli 2018, S. 1–8. doi:10.1016/j.gr.2018.02.017.
- J. Gregory Shellnutt: The Emeishan large igneous province: A synthesis. In: Geoscience Frontiers (Elsevier). 5, Nr. 3, August 2016, S. 369–394. doi:10.1016/j.gsf.2013.07.003.
- Stephen E. Grasby, Hamed Sanei, Benoit Beauchamp: Catastrophic dispersion of coal fly ash into oceans during the latest Permian extinction. (PDF) In: Nature Geoscience. 4, Februar 2011, S. 104–107. doi:10.1038/ngeo1069.
- L. T. Elkins-Tanton, S. E. Grasby, B. A. Black, R. V. Veselovskiy, O. H. Ardakani, F. Goodarzi: Field evidence for coal combustion links the 252 Ma Siberian Traps with global carbon disruption. (PDF) In: Geology. 48, Juni 2020. doi:10.1130/G47365.1.
- Henrik H. Svensen, Sergei Frolov, Grigorii G. Akhmanov, Alexander G. Polozov, Dougal A. Jerram, Olga V. Shiganova, Nikolay V. Melnikov, Karthik Iyer, Sverre Planke: Sills and gas generation in the Siberian Traps. In: Philosophical Transactions of the Royal Society A. 376, Nr. 2130, Oktober 2018. doi:10.1098/rsta.2017.0080.
- S. D. Burgess, J. D. Muirhead, S. A. Bowring: Initial pulse of Siberian Traps sills as the trigger of the end-Permian mass extinction. In: Nature Communications. 8, Juli 2017. doi:10.1038/s41467-017-00083-9.
- Michael M. Joachimski, Xulong Lai, Shuzhong Shen, Haishui Jiang, Genming Luo, Bo Chen, Jun Chen, Yadong Sun: Climate warming in the latest Permian and the Permian–Triassic mass extinction. (PDF) In: Geology. 40, Nr. 3, März 2012, S. 195–198. doi:10.1130/G32707.
- M. O. Clarkson, S. A. Kasemann, R. A. Wood, T. M. Lenton, S. J. Daines, S. Richoz, F. Ohnemueller, A. Meixner, S. W. Poulton, E. T. Tipper: Ocean acidification and the Permo-Triassic mass extinction. (PDF) In: Science. 348, Nr. 6231, April 2015, S. 229–232. doi:10.1126/science.aaa0193.
- Yadong Sun, Michael M. Joachimski, Paul B. Wignall, Chunbo Yan, Yanlong Chen, Haishui Jiang, Lina Wang, Xulong Lai: Lethally Hot Temperatures During the Early Triassic Greenhouse. (PDF) In: Science. 338, Nr. 6105, Oktober 2012, S. 366–370. doi:10.1126/science.1224126.
- Michael J. Benton, Richard J. Twitchett: How to kill (almost) all life: the end-Permian extinction event. (PDF) In: Trends in Ecology and Evolution. 18, Nr. 7, Juli 2003, S. 358–365. doi:10.1016/S0169-5347(03)00093-4.
- Hana Jurikova, Marcus Gutjahr, Klaus Wallmann, Sascha Flögel, Volker Liebetrau, Renato Posenato, Lucia Angiolini, Claudio Garbelli, Uwe Brand, Michael Wiedenbeck, Anton Eisenhauer: Permian–Triassic mass extinction pulses driven by major marine carbon cycle perturbations. In: Nature Geoscience. Oktober 2020. doi:10.1038/s41561-020-00646-4.
- Jeffrey P. Benca, Ivo A. P. Duijnstee, Cindy V. Looy: UV-B–induced forest sterility: Implications of ozone shield failure in Earth’s largest extinction. In: Science Advances. 4, Nr. 2, Februar 2018. doi:10.1126/sciadv.1700618.
- Seth D. Burgess, Samuel Bowring, Shu-Zhong Shen: High-precision timeline for Earth’s most severe extinction. In: PNAS. 111, Nr. 9, März 2014, S. 3316–3321. doi:10.1073/pnas.1317692111.
- Shu-Zhong Shen, Jahandar Ramezani, Jun Chen, Chang-Qun Cao, Douglas H. Erwin, Hua Zhang, Lei Xiang, Shane D. Schoepfer, Charles M. Henderson, Quan-Feng Zheng, Samuel A. Bowring, Yue Wang, Xian-Hua Li, Xiang-Dong Wang, Dong-Xun Yuan, Yi-Chun Zhang, Lin Mu, Jun Wang, Ya-Sheng Wu: A sudden end-Permian mass extinction in South China. (PDF) In: GSA Bulletin (The Geological Society of America). 131, Nr. 1–2, Januar 2019, S. 205–223. doi:10.1130/B31909.1.
- Michael J. Benton, Andrew J. Newell: Impacts of global warming on Permo-Triassic terrestrial ecosystems. (PDF) In: Gondwana Research. 25, Nr. 4, Mai 2014, S. 1308–1337. doi:10.1016/j.gr.2012.12.010.
- Zhong-Qiang Chen, Michael J. Benton: The timing and pattern of biotic recovery following the end-Permian mass extinction. (PDF) In: Nature Geoscience. 5, Nr. 6, Juni 2012, S. 375–383. doi:10.1038/ngeo1475.
- Terrence J. Blackburn, Paul E. Olsen, Samuel A. Bowring, Noah M. McLean, Dennis V. Kent, John Puffer, Greg McHone, E. Troy Rasbury, Mohammed Et-Touhami: Zircon U-Pb Geochronology Links the End-Triassic Extinction with the Central Atlantic Magmatic Province. (PDF) In: Science. 340, Nr. 6135, Mai 2013, S. 941–945. doi:10.1126/science.1234204.
- Tran T. Huynh, Christopher J. Poulsen: Rising atmospheric CO2 as a possible trigger for the end-Triassic mass extinction. (PDF) In: Palaeogeography, Palaeoclimatology, Palaeoecology. 217, Nr. 3–4, Februar 2005, S. 223–242. doi:10.1016/j.palaeo.2004.12.004.
- Jessica H. Whiteside, Paul E. Olsen, Timothy Eglinton, Michael E. Brookfield, Raymond N. Sambrotto: Compound-specific carbon isotopes from Earth’s largest flood basalt eruptions directly linked to the end-Triassic mass extinction. In: PNAS. 107, Nr. 15, April 2010, S. 6721–6725. doi:10.1073/pnas.1001706107.
- J. H. F. L. Davies, H. Bertrand, N. Youbi, M. Ernesto, U. Schaltegger: End-Triassic mass extinction started by intrusive CAMP activity. (htlm) In: Nature Communications. 8, Mai 2017. doi:10.1038/ncomms15596.
- Thea H. Heimdal, Henrik. H. Svensen, Jahandar Ramezani, Karthik Iyer, Egberto Pereira, René Rodrigues, Morgan T. Jones, Sara Callegaro: Large-scale sill emplacement in Brazil as a trigger for the end-Triassic crisis. In: Nature Scientific Reports. 8, Januar 2018. doi:10.1038/s41598-017-18629-8.
- Sylvain Richoz, Bas van de Schootbrugge, Jörg Pross, Wilhelm Püttmann, Tracy M. Quan, Sofie Lindström, Carmen Heunisch, Jens Fiebig, Robert Maquil, Stefan Schouten, Christoph A. Hauzenberger, Paul B. Wignall: Hydrogen sulphide poisoning of shallow seas following the end-Triassic extinction. (PDF) In: Nature Geoscience. 5, August 2012, S. 662–667. doi:10.1038/NGEO1539.
- G. Keller, T. Adatte, W. Stinnesbeck, M. Rebolledo-Vieyra, J. U. Fucugauchi, U. Kramar, Doris Stüben: Chicxulub impact predates the K-T boundary mass extinction. In: PNAS. 101, Nr. 11, März 2004, S. 3753–3758. doi:10.1073/pnas.0400396101.
- Peter Schulte, Robert P. Speijer, Henk Brinkhuis, Agnes Kontny, Philippe Claeys, Simone Galeotti, Jan Smit: Comment on the paper „Chicxulub impact predates KT boundary: New evidence from Brazos, Texas“ by Keller et al. (2007). (PDF) In: Earth and Planetary Science Letters. 269, Nr. 3–4, Mai 2008, S. 614–620. doi:10.1016/j.epsl.2007.11.06.
- Paul A. Renne, Alan L. Deino, Frederik J. Hilgen, Klaudia F. Kuiper, Darren F. Mark, William S. Mitchell III, Leah E. Morgan, Roland Mundil, Jan Smit: Time Scales of Critical Events Around the Cretaceous-Paleogene Boundary. (PDF) In: Science. 339, Nr. 6120, Februar 2013, S. 684–687. doi:10.1126/science.1230492.
- Johan Vellekoop, Appy Sluijs, Jan Smit, Stefan Schouten, Johan W. H. Weijers, Jaap S. Sinninghe Damsté, Henk Brinkhuis: Rapid short-term cooling following the Chicxulub impact at the Cretaceous-Paleogene boundary. In: PNAS. 111, Nr. 21, Mai 2014, S. 7537–7541. doi:10.1073/pnas.1319253111.
- Stephen L. Brusatte, Richard J. Butler, Paul M. Barrett, Matthew T. Carrano, David C. Evans, Graeme T. Lloyd, Philip D. Mannion, Mark A. Norell, Daniel J. Peppe, Paul Upchurch, Thomas E. Williamson: The extinction of the dinosaurs. In: Biological Reviews, Cambridge Philosophical Society (Wiley Online Library). 90, Nr. 2, Mai 2015, S. 628–642. doi:10.1111/brv.12128.
- Michael J. Henehan, Andy Ridgwell, Ellen Thomas, Shuang Zhang, Laia Alegret, Daniela N. Schmidt, James W. B. Rae, James D. Witts, Neil H. Landman, Sarah E. Greene, Brian T. Huber, James R. Super, Noah J. Planavsky, Pincelli M. Hull: Rapid ocean acidification and protracted Earth system recovery followed the end-Cretaceous Chicxulub impact. In: PNAS. 116, Nr. 43, Oktober 2019. doi:10.1073/pnas.1905989116.
- Douglas S. Robertson, Malcolm C. McKenna, Owen B. Toon, Sylvia Hope, Jason A. Lillegraven: Survival in the first hours of the Cenozoic. (PDF) In: Geological Society of America Bulletin. 116, Nr. 5/6, S. 760–768. doi:10.1130/B25402.
- Douglas S. Robertson, William M. Lewis, Peter M. Sheehan, Owen B. Toon: Reevaluation of the heat-fire hypothesis. In: Journal of Geophysical Research: Biogeoscience. 110, Nr. 1, März 2013, S. 329–336. doi:10.1002/jgrg.20018.
- Julia Brugger, Georg Feulner, Stefan Petri: Baby, it's cold outside: Climate model simulations of the effects of the asteroid impact at the end of the Cretaceous. In: Geophysical Research Letters. 44, Nr. 1, Januar 2017, S. 419–427. doi:10.1002/2016GL072241.
- Nicholas R. Longrich, Tim Tokaryk, Daniel J. Field: Mass extinction of birds at the Cretaceous-Paleogene (K-Pg) boundary. In: PNAS. 108, Nr. 37, September 2011, S. 15253–15257. doi:10.1073/pnas.1110395108.
- Paul R. Renne, Courtney J. Sprain, Mark A. Richards, Stephen Self, Loÿc Vanderkluysen, Kanchan Pande: State shift in Deccan volcanism at the Cretaceous-Paleogene boundary, possibly induced by impact. (PDF) In: Science. 350, Nr. 6256, Oktober 2015, S. 76–78. doi:10.1126/science.aac7549.
- Pincelli M. Hull, André Bornemann, Donald E. Penman, Michael J. Henehan, Richard D. Norris, Paul A. Wilson, Peter Blum, Laia Alegret, Sietske J. Batenburg, Paul R. Bown, Timothy J. Bralower, Cecile Cournede, Alexander Deutsch, Barbara Donner, Oliver Friedrich, Sofie Jehle, Hojung Kim, Dick Kroon, Peter C. Lippert, Dominik Loroch, Iris Moebius, Kazuyoshi Moriya, Daniel J. Peppe, Gregory E. Ravizza, Ursula Röhl, Jonathan D. Schueth, Julio Sepúlveda, Philip F. Sexton, Elizabeth C. Sibert, Kasia K. Śliwińska, Roger E. Summons, Ellen Thomas, Thomas Westerhold, Jessica H. Whiteside, Tatsuhiko Yamaguchi, James C. Zachos: On impact and volcanism across the Cretaceous-Paleogene boundary. (PDF) In: Science. 367, Nr. 6475, Januar 2020, S. 266–272. doi:10.1126/science.aay5055.
- James S. Eldrett, Ian C. Harding, Paul A. Wilson, Emily Butler, Andrew P. Roberts: Continental ice in Greenland during the Eocene and Oligocene. (PDF) In: Nature. 446, März 2007, S. 176–179. doi:10.1038/nature05591.
- Michael Stärz, Wilfried Jokat, Gregor Knorr, Gerrit Lohmann: Threshold in North Atlantic-Arctic Ocean circulation controlled by the subsidence of the Greenland-Scotland Ridge. In: Nature Communications (online). 8, Juni 2017. doi:10.1038/ncomms15681.
- Mark Pagani, Matthew Huber, Zhonghui Liu, Steven M. Bohaty, Jorijntje Henderiks, Willem Sijp, Srinath Krishnan, Robert M. DeConto: The Role of Carbon Dioxide During the Onset of Antarctic Glaciation. (PDF) In: Science. 334, Nr. 6060, Dezember 2011, S. 1261–1264. doi:10.1126/science.1203909.
- Becky Oskin: Russia's Popigai Meteor Crash Linked to Mass Extinction. livescience, 13. Juni 2014.
- Catalina Pimiento, John N. Griffin, Christopher F. Clements, Daniele Silvestro, Sara Varela; Mark D. Uhen, Carlos Jaramillo: The Pliocene marine megafauna extinction and its impact on functional diversity. (PDF) In: Nature ecology & evolution. 2017. doi:10.1038/s41559-017-0223-6.
- Kenshu Shimada: The size of the megatooth shark, Otodus megalodon (Lamniformes: Otodontidae), revisited. In: Historical Biology. September 2019. doi:10.1080/08912963.2019.1666840.
- Sander van der Kaars, Gifford H. Miller, Chris S. M. Turney, Ellyn J. Cook, Dirk Nürnberg, Joachim Schönfeld, A. Peter Kershaw, Scott J. Lehman: Humans rather than climate the primary cause of Pleistocene megafaunal extinction in Australia. In: Nature Communications. 8, Januar 2017. doi:10.1038/ncomms14142.
- David L. Fox, Daniel C. Fisher, Sergey Vartanyan, Alexei N. Tikhonov, Dick Mol, Bernard Buigues: Paleoclimatic implications of oxygen isotopic variation in late Pleistocene and Holocene tusks of Mammuthus primigenius from northern Eurasia. (PDF) In: Quaternary International. 169–170, Juli 2007, S. 154–165. doi:10.1016/j.quaint.2006.09.001.
- R. B. Firestone, A. West, J. P. Kennett, L. Becker, T. E. Bunch, Z. S. Revay, P. H. Schultz, T. Belgya, D. J. Kennett, J. M. Erlandson, O. J. Dickenson, A. C. Goodyear, R. S. Harris, G. A. Howard, J. B. Kloosterman, P. Lechler, P. A. Mayewski, J. Montgomery, R. Poreda, T. Darrah, S. S. Que Hee, A. R. Smith, A. Stich, W. Topping, J. H. Wittke, W. S. Wolbach: Evidence for an extraterrestrial impact 12,900 years ago that contributed to the megafaunal extinctions and Younger Dryas cooling. In: Proceedings of the National Academy of Sciences (PNAS). 103, Nr. 41, Oktober 2007, S. 16016–16021. doi:10.1073/pnas.0706977104.
- Nicholas Pinter, Andrew C. Scott, Tyrone L. Daulton, Andrew Podoll, Christian Koeberl, R. Scott Anderson, Scott E. Ishman: The Younger Dryas impact hypothesis: A requiem. (PDF) In: Earth-Science Reviews (Elsevier). 106, Nr. 3–4, Juni 2011, S. 247–264. doi:10.1016/j.earscirev.2011.02.005.
- Wendy S. Wolbach, Joanne P. Ballard, Paul A. Mayewski, Andrew C. Parnell, Niamh Cahill, Victor Adedeji, Ted E. Bunch, Gabriela Domínguez-Vázquez, Jon M. Erlandson, Richard B. Firestone, Timothy A. French, George Howard, Isabel Israde-Alcántara, John R. Johnson, David Kimbel, Charles R. Kinzie, Andrei Kurbatov, Gunther Kletetschka, Malcolm A. LeCompte, William C. Mahaney, Adrian L. Melott, Siddhartha Mitra, Abigail Maiorana-Boutilier, Christopher R. Moore, William M. Napier, Jennifer Parlier, Kenneth B. Tankersley, Brian C. Thomas, James H. Wittke, Allen West, James P. Kennett: Extraordinary Biomass-Burning Episode and Impact Winter Triggered by the Younger Dryas Cosmic Impact ∼12,800 Years Ago. 2. Lake, Marine, and Terrestrial Sediments. (PDF) In: The Journal of Geology. Februar 2018. doi:10.1086/695704.
- Andrew M. T. Moore, James P. Kennett, William M. Napier, Ted E. Bunch, James C. Weaver, Malcolm LeCompte, A. Victor Adedeji, Paul Hackley, Gunther Kletetschka, Robert E. Hermes, James H. Wittke, Joshua J. Razink, Michael W. Gaultois, Allen West: Evidence of Cosmic Impact at Abu Hureyra, Syria at the Younger Dryas Onset (~12.8 ka): High-temperature melting at >2200 °C. In: Nature Scientific Reports. 10, März 2020. doi:10.1038/s41598-020-60867-w.
- Bradley D. Cramer, Daniel J. Condon, Ulf Söderlund, Carly Marshall, Graham J. Worton, Alan T. Thomas, Mikael Calner, David C. Ray, Vincent Perrier, Ian Boomer, P. Jonathan Patchett, Lennart Jeppsson: U-Pb (zircon) age constraints on the timing and duration of Wenlock (Silurian) paleocommunity collapse and recovery during the “Big Crisis”. (PDF) In: Geological Society of America (Bulletin). 124, Nr. 11–12, Oktober 2012, S. 1841–1857. doi:10.1130/B30642.1.
- Štěpán Manda, Jiří Frýda: Silurian-Devonian boundary events and their influence on cephalopod evolution: evolutionary significance of cephalopod egg size during mass extinctions. (PDF) In: Bulletin of Geosciences. 85, Nr. 3, 2010, S. 513–540. doi:10.3140/bull.geosci.1174.
- Alexander J. Hetherington, Joseph G. Dubrovsky, Liam Dolan: Unique Cellular Organization in the Oldest Root Meristem. In: Current Biology. 26, Nr. 12, Juni 2016, S. 1629–1633. doi:10.1016/j.cub.2016.04.072.
- Peter Franks: New constraints on atmospheric CO2 concentration for the Phanerozoic. (PDF) In: Geophysical Research Letters. 31, Nr. 13, Juli 2014. doi:10.1002/2014GL060457.
- Isabel P. Montañez, Jennifer C. McElwain, Christopher J. Poulsen, Joseph D. White, William A. DiMichele, Jonathan P. Wilson, Galen Griggs, Michael T. Hren: Climate, pCO2 and terrestrial carbon cycle linkages during late Palaeozoic glacial–interglacial cycles. (PDF) In: Nature Geoscience. 9, Nr. 11, November 2016, S. 824–828. doi:10.1038/ngeo2822.
- Georg Feulner: Formation of most of our coal brought Earth close to global glaciation. In: PNAS. 114, Nr. 43, Oktober 2017, S. 11333–11337. doi:10.1073/pnas.1712062114.
- Borja Cascales-Miñana, Christopher J. Cleal: The plant fossil record reflects just two great extinction events. In: Terra Nova. 26, Nr. 3, 2013, S. 195–200. doi:10.1111/ter.12086.
- William A. DiMichele, Neil J. Tabor, Dan S. Chaney, W. John Nelson: From wetlands to wet spots: Environmental tracking and the fate of Carboniferous elements in Early Permian tropical floras. (PDF) In: GSA (Geological Society of America). Special Paper 399, 2006, S. 223–248. doi:10.1130/2006.2399(11).
- Sarda Sahney, Michael J. Benton, Howard J. Falcon-Lang: Rainforest collapse triggered Pennsylvanian tetrapod diversification in Euramerica. (PDF) In: Geology. 38, Nr. 12, November 2010, S. 1079–1082. doi:10.1130/G31182.1.
- Guillaume Dera, Benjamin Brigaud, Fabrice Monna, Rémi Laffont, Emmanuelle Pucéat, Jean-François Deconinck, Pierre Pellenard, Michael M. Joachimski, Christophe Durlet: Climatic ups and downs in a disturbed Jurassic world. (PDF) In: Geology. 53, Nr. 3, März 2011, S. 215–218. doi:10.1130/G31579.1.
- Federico Fanti, Tetsuto Miyashita, Luigi Cantelli, Fawsi Mnasri, Jihed Dridi, Michela Contessi, Andrea Cau: The largest thalattosuchian (Crocodylomorpha) supports teleosaurid survival across the Jurassic-Cretaceous boundary. (PDF) In: Cretaceous Research. 61, Juni 2016, S. 263–274. doi:10.1016/j.cretres.2015.11.011.
- Yongdong Wang, Chengmin Huang, Bainian Sun, Cheng Quan, Jingyu Wu, Zhicheng Lin: Paleo-CO2 variation trends and the Cretaceous greenhouse climate. (PDF) In: Earth-Science Reviews. 129, Februar 2014, S. 136–147. doi:10.1016/j.earscirev.2013.11.001.
- Vanessa C. Bowman, Jane E. Francis, James B. Riding: Late Cretaceous winter sea ice in Antarctica?. (PDF) In: Geology. 41, Nr. 12, Dezember 2013, S. 1227–1230. doi:10.1130/G34891.1.
- James S. Eldrett, Ian Jarvis, John S. Lignum, Darren R. Gröcke, Hugh C. Jenkyns, Martin A. Pearce: Black shale deposition, atmospheric CO2 drawdown, and cooling during the Cenomanian-Turonian Oceanic Anoxic Event. In: Paleoceanography and Paleoclimatology. 26, Nr. 3, September 2011. doi:10.1029/2010PA002081.
- James S. Eldrett, Chao Ma, Steven C. Bergman, Brendan Lutz, F. John Gregory, Paul Dodsworth, Mark Phipps, Petros Hardas, Daniel Minisini, Aysen Ozkan, Jahander Ramezani, Samuel A. Bowring, Sandra L. Kamo, Kurt Ferguson, Calum Macaulay, Amy E. Kelly: An astronomically calibrated stratigraphy of the Cenomanian, Turonian and earliest Coniacian from the Cretaceous Western Interior Seaway, USA: Implications for global chronostratigraphy. (PDF) In: Cretaceous Research. 56, S. 316–344. doi:10.1016/j.cretres.2015.04.010.
- Valentin Fischer, Nathalie Bardet, Roger B. J. Benson, Maxim S. Arkhangelsky, Matt Friedman: Extinction of fish-shaped marine reptiles associated with reduced evolutionary rates and global environmental volatility. In: Nature Communinications. 7, März 2016. doi:10.1038/ncomms10825.
- Richard E. Zeebe, Andy Ridgwell, James C. Zachos: Anthropogenic carbon release rate unprecedented during the past 66 million years. (PDF) In: Nature Geoscience. 9, Nr. 4, April 2016, S. 325–329. doi:10.1038/ngeo2681.
- Donald E. Penman, Bärbel Hönisch, Richard E. Zeebe, Ellen Thomas, James C. Zachos: Rapid and sustained surface ocean acidification during the Paleocene-Eocene Thermal Maximum. (PDF) In: Oceanography. 29, Nr. 5, Mai 2014, S. 357–369. doi:10.1002/2014PA002621.
- Francesca A. McInerney, Scott L. Wing: The Paleocene-Eocene Thermal Maximum: A Perturbation of Carbon Cycle, Climate, and Biosphere with Implications for the Future. (PDF) In: Annual Review of Earth and Planetary Sciences. 39, Mai 2011, S. 489–516. doi:10.1146/annurev-earth-040610-133431.
- Abigail R. D. Ambrosia, William C. Clyde, Henry C. Fricke, Philip D. Gingerich, Hemmo A. Abels: Repetitive mammalian dwarfing during ancient greenhouse warming events. (PDF) In: Science Advances. 3, Nr. 3, März 2017. doi:10.1126/sciadv.1601430.
- Stephen G. B. Chester, Jonathan I. Bloch, Ross Secord, Doug M. Boyer: A New Small-Bodied Species of Palaeonictis (Creodonta, Oxyaenidae) from the Paleocene-Eocene Thermal Maximum. (PDF) In: Journal of Mammalian Evolution. 17, Nr. 4, Dezember 2010, S. 227–243. doi:10.1007/s10914-010-9141-y.
- Tatsuhiko Yamaguchi, Richard D. Norris, André Bornemann: Dwarfing of ostracodes during the Paleocene–Eocene Thermal Maximum at DSDP Site 401 (Bay of Biscay, North Atlantic) and its implication for changes in organic carbon cycle in deep-sea benthic ecosystem. (PDF) In: Palaeogeography, Palaeoclimatology, Palaeoecology. 346–347, Nr. 6384, August 2012, S. 130–144. doi:10.1016/j.palaeo.2012.06.004.
- William J. Ripple, Christopher Wolf, Thomas M. Newsome, Mauro Galetti, Mohammed Alamgir, Eileen Crist, Mahmoud I. Mahmoud, William F. Laurance und 15.364 Biowissenschaftler aus 184 Ländern: World Scientists’ Warning to Humanity: A Second Notice. In: BioScience. Band 67, Nr. 12, 2017, S. 1026–1028, doi:10.1093/biosci/bix125.
- Klaus Jacob: Die sechste Katastrophe. In: Süddeutsche Zeitung, 31. August 2014.
- Gerardo Ceballos, Paul R. Ehrlich, Anthony D. Barnosky, Andrés García, Robert M. Pringle, Todd M. Palmer: Accelerated modern human–induced species losses: Entering the sixth mass extinction. In: Science Advances. 1, 2015, S. e1400253, doi:10.1126/sciadv.1400253.
- wwf.de: Gewinner und Verlierer 2014
- European wild bird indicators and Trends of common birds in Europe, 2018 update. In: ebcc.info. 21. Januar 2019, abgerufen am 31. Januar 2019.
- Christian Mihatsch: Die Welt verliert 380 Tier- und Pflanzenarten pro Tag. Badische Zeitung, 6. Oktober 2014
- WWF Deutschland Pressestelle: Gewinner und Verlierer 2016 WWF: 2016 ist ein durchwachsenes Jahr für den Artenschutz
- World wildlife plummets more than two-thirds in 50 years: index (en). In: phys.org.
- Living Planet Report 2020.
- badische-zeitung.de, Panorama, 28. Oktober 2016: Tierbestand ist in Deutschland, aber auch weltweit zurückgegangen (30. Oktober 2016)
- Südkurier: Ornithologe Peter Berthold: "Unseren Vögeln geht's beschissen"
- Rachel Nuwer: Mass Extinctions Are Accelerating, Scientists Report. In: The New York Times, 1. Juni 2020. Abgerufen am 2. Juli 2020.
- Gerardo Ceballos, Paul R. Ehrlich, Peter H. Raven: Vertebrates on the brink as indicators of biological annihilation and the sixth mass extinction. In: Proceedings of the National Academy of Sciences. 117, Nr. 24, 16. Juni 2020, ISSN 0027-8424, S. 13596–13602. bibcode:2020PNAS..11713596C. doi:10.1073/pnas.1922686117. PMID 32482862. Abgerufen am 2. Juli 2020.
- Alarmierender Uno-Bericht: Bis zu eine Million Arten sind vom Aussterben bedroht. In: nzz.ch. 23. April 2019, abgerufen am 24. April 2019.
- Der Mensch verdrängt eine Million Tier- und Pflanzenarten, Süddeutsche Zeitung, 6. Mai 2019.
- Francisco Sánchez-Bayo, Kris A. G. Wyckhuys: Worldwide decline of the entomofauna: A review of its drivers. In: Biological Conservation. 232, April 2019, S. 8–27. doi:10.1016/j.biocon.2019.01.020.
- Mark C. Urban: Accelerating extinction risk from climate change. (PDF) In: Science. 348, Nr. 6234, Mai 2015, S. 571–573. doi:10.1126/science.aaa4984.
- Living Planet Report 2020 (deutsche Kurzfassung). (PDF; 4,77 MB) In: wwf.de. Abgerufen am 10. Oktober 2020.
- C. D. Harvell, D. Montecino-Latorre u. a.: Disease epidemic and a marine heat wave are associated with the continental-scale collapse of a pivotal predator (Pycnopodia helianthoides). In: Science Advances. 5, 2019, S. eaau7042, doi:10.1126/sciadv.aau7042.
- Gregory P. Wilson: Mammals across the K/Pg boundary in northeastern Montana, U.S.A.: dental morphology and body-size patterns reveal extinction selectivity and immigrant-fueled ecospace filling. (PDF) In: Paleobiology. 39, Nr. 3, Mai 2013, S. 429–469. doi:10.1666/12041.
- Evolution imposes 'speed limit' on recovery after mass extinctions. In: ScienceDaily. Abgerufen am 7. September 2019 (englisch).
- Jens Boenigk, Sabina Wodniok: Biodiversität und Erdgeschichte. Berlin 2014, ISBN 978-3-642-55388-2, S. 69.
- Jason S. Anderson, Tim Smithson, Chris F. Mansky, Taran Meyer, Jennifer Clack: A Diverse Tetrapod Fauna at the Base of 'Romer’s Gap'. In: PLOS ONE. 10, Nr. 4, April 2015. doi:10.1371/journal.pone.0125446.
- Manabu Sakamoto, Michael Benton, Chris Venditti: Dinosaurs in decline tens of millions of years before their final extinction. In: PNAS. 113, Nr. 18, Mai 2016, S. 5036–5040. doi:10.1073/pnas.1521478113.
- Zoltan Siki-Sava, Eric Buffetaut, Attila Ősi, Xabier Pereda-Suberbiola, Stephen L. Brusatte: Island life in the Cretaceous – faunal composition, biogeography, evolution, and extinction of land-living vertebrates on the Late Cretaceous European archipelago. In: ZooKeys. 469, Januar 2015, S. 1–161. doi:10.3897/zookeys.469.8439.
- Stephen L. Brusatte, Carlos R. A. Candeiro, Felipe M. Simbras: The last dinosaurs of Brazil: The Bauru Group and its implications for the end-Cretaceous mass extinction. In: Anais da Academia Brasileira de Ciências (Rio de Janeiro). 89, Nr. 3. doi:10.1590/0001-3765201720160918.
- Nicholas R. Longrich, David M. Martill, Brian Andres: Late Maastrichtian pterosaurs from North Africa and mass extinction of Pterosauria at the Cretaceous-Paleogene boundary. In: PLOS Biology. März 2018. doi:10.1371/journal.pbio.2001663.
- Robert A. DePalma, Jan Smit, David A. Burnham, Klaudia Kuiper, Phillip L. Manning, Anton Oleinik, Peter Larson, Florentin J. Maurrasse, Johan Vellekoop, Mark A. Richards, Loren Gurche, Walter Alvarez: A seismically induced onshore surge deposit at the KPg boundary, North Dakota. In: PNAS. 116, Nr. 17, April 2019, S. 8190–8199. doi:10.1073/pnas.1817407116.
- Katherine Kornei: Huge global tsunami followed dinosaur-killing asteroid impact. In: Eos. 20. Dezember 2018, doi:10.1029/2018EO112419.
- Peter Wilf, Kirk R. Johnson: Land plant extinction at the end of the Cretaceous: a quantitative analysis of the North Dakota megafloral record. (PDF) In: Paleobiology. 30, September 2004, S. 347–368. doi:10.1666/0094-8373(2004)030<0347:LPEATE>2.0.CO;2.
- Jeffrey A. Fawcett, Steven Maere, Yves Van de Peer: Plants with double genomes might have had a better chance to survive the Cretaceous–Tertiary extinction event. In: PNAS. 106, Nr. 14, April 2009, S. 5737–5742. doi:10.1073/pnas.0900906106.
- Nicholas R. Longrich, Bhart-Anjan S. Bhullar, Jacques A. Gauthier: Mass extinction of lizards and snakes at the Cretaceous–Paleogene boundary. In: PNAS. 109, Nr. 52, Dezember 2012, S. 21396–21401. doi:10.1073/pnas.1211526110.
- Stéphane Jouve, Nathalie Bardet, Nour-Eddine Jalil, Xabier Pereda Suberbiola, Baâdi Bouya, Mbarek Amaghzaz: The Oldest African Crocodylian: Phylogeny, Paleobiogeography, and Differential Survivorship Of Marine Reptiles Through the Cretaceous-Tertiary Boundary. (PDF) In: Journal of Vertebrate Paleontology. 28, Februar 2008, S. 409–421. doi:10.1671/0272-4634(2008)28[409:TOACPP]2.0.CO;2.
- Michael J. Henehan, Pincelli M. Hull, Donald E. Penman, James W. B. Rae, Daniela N. Schmidt: Biogeochemical significance of pelagic ecosystem function: an end-Cretaceous case study. In: Philosophical Transactions of the Royal Society B (Biological Sciences). 171, Nr. 1694, Mai 2016. doi:10.1098/rstb.2015.0510.
- Toby Tyrrell, Agostino Merico, David Ian Armstrong McKay: Severity of ocean acidification following the end-Cretaceous asteroid impact. In: PNAS. 112, Nr. 21, Mai 2015, S. 6556–6561. doi:10.1073/pnas.1418604112.
- Kunio Kaiho, Naga Oshima, Kouji Adachi, Yukimasa Adachi, Takuya Mizukami, Megumu Fujibayashi, Ryosuke Saito: Global climate change driven by soot at the K-Pg boundary as the cause of the mass extinction. In: Nature Scientific Reports. 6, Juli 2016. doi:10.1038/srep28427.
- Michael P. Donovan, Ari Iglesias, Peter Wilf, Conrad C. Labandeira, N. Rubén Cúneo: Rapid recovery of Patagonian plant–insect associations after the end-Cretaceous extinction. In: Nature Ecology & Evolution. 1, Dezember 2016. doi:10.1038/s41559-016-0012.
- Sierra V. Petersen, Andrea Dutton, Kyger C. Lohmann: End-Cretaceous extinction in Antarctica linked to both Deccan volcanism and meteorite impact via climate change. In: Nature Communications. 7, Juli 2016. doi:10.1038/ncomms12079.
- Kunio Kaiho, Naga Oshima: Site of asteroid impact changed the history of life on Earth: the low probability of mass extinction. In: Scientific Reports. 7, November 2017. doi:10.1038/s41598-017-14199-x.
- G. S. Collin, N. Patel, T. M. Davison, A. S. P. Rae, J. V. Morgan, S. P. S. Gulick, IODP-ICDP Expedition 364 Science Party: G. L. Christeson, E. Chenot, P. Claeys, C. S. Cockell, M. J. L. Coolen, L. Ferrière, C. Gebhardt, K. Goto, H. Jones, D. A. Kring, J. Lofi, C. M. Lowery, R. Ocampo-Torres, L. Perez-Cruz, A. E. Pickersgill, M. H. Poelchau, C. Rasmussen, M. Rebolledo-Vieyra, U. Riller, H. Sato, J. Smit, S. M. Tikoo, N. Tomioka, J. Urrutia-Fucugauchi, M. T. Whalen, A. Wittmann, L. Xiao, K. E. Yamaguchi, Third-Party Scientists: N. Artemieva, T. J. Bralower: A steeply-inclined trajectory for the Chicxulub impact. In: Nature Communications. 11, Mai 2020. doi:10.1038/s41467-020-15269-x.