Trilobiten
Die Trilobiten (Trilobita, „Dreilapper“, von altgriechisch τρία tria „drei“ und λοβός lobós „Lappen“) sind eine ausgestorbene Klasse meeresbewohnender Gliederfüßer (Arthropoda).
| Trilobiten | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Isotelus brachycephalus | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Kambrium bis Perm | ||||||||||||
| 521 bis 251 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Trilobita | ||||||||||||
| Walch, 1771 |
Die Trilobiten existierten nahezu während der gesamten Spanne des Paläozoikums (Erdaltertum), von der 2. Serie des Kambriums (Beginn vor 521 Mio. Jahren) bis zum Massenaussterben am Ende des Perms vor etwa 251 Mio. Jahren. Ihre mit Calcit (Calciumcarbonat) zu einem Panzer verstärkten Exoskelette (Außenskelette) sind als Fossilien in großer Zahl erhalten geblieben und ermöglichen so, die Evolution und den Formenreichtum der zahlreichen Arten zu rekonstruieren. Dies, in Verbindung mit ihrer Schichtbeständigkeit und weiten geographischen Ausdehnung, macht Trilobiten zu wichtigen Leitfossilien für das Paläozoikum, insbesondere im Kambrium.
Die Trilobiten gehörten zu den ersten Gliederfüßern (Arthropoda), einem Tierstamm mit Exoskelett, gegliedertem Körperbau und vielen koordiniert arbeitenden Beinen. Ihre Fährten sind ebenfalls vielfach als Ichnofossilien gefunden worden, man nennt diese Spurenfossilien Rusophycus und Cruziana.[1]
Die ausgestorbene Klasse der Trilobita besteht aus neun anerkannten Ordnungen, über 150 Familien, über 5000 Gattungen und mehr als 15.000 beschriebenen Arten. Jedes Jahr werden weitere Arten gefunden und beschrieben. Ihre Vielfalt macht die Trilobiten zur divergentesten Gruppe unter allen ausgestorbenen Lebewesen. Der größte bekannte Trilobit mit mehr als 70 cm Länge ist Isotelus rex aus dem Oberordovizium von Nordamerika.
Namensgebung
Die Bezeichnung „Trilobit“ wurde 1771 von Johann Ernst Immanuel Walch eingeführt,[2][3] doch erst zu Beginn des 19. Jahrhunderts setzte sich der Name in der Wissenschaft durch.
Körperbau
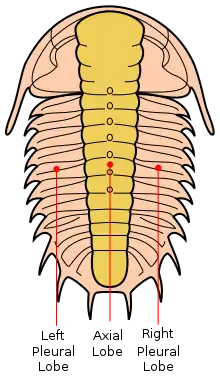
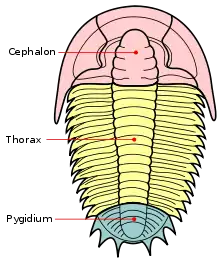
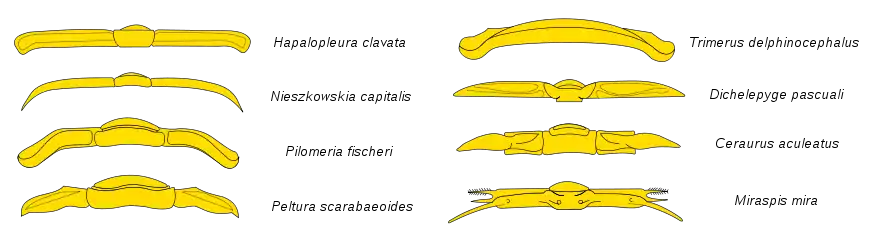
Die Trilobiten („Dreilapper“, früher auch irreführend „Dreilappenkrebse“ genannt) bestehen immer aus drei sagittal verlaufenden Loben („Lappen“), die für diese Tiere namensgebend sind: dem Spindellobus und den beiden pleuralen Loben auf der linken und rechten Seite. Die Trilobiten werden auch transversal in drei Glieder (Tagmata) geteilt: Kopfschild (Cephalon), Thorax und Schwanzschild (Pygidium).
Spindellobus
Der mittlere Lobus wird Spindellobus oder Axislobus genannt. Das Stück des Spindellobus auf dem Kopfschild (Cephalon) heißt Stirnlappen (Glabella) und besteht häufig aus mehreren transversal verlaufenden und oftmals verwachsenen Loben. Zwischen den Loben sind unvollständige oder vollständig teilende Furchen vorhanden. Der vordere Teil der Glabella wird Anteroglabella und der hintere Teil Posteroglabella genannt. Links und rechts neben der Posteroglabella befinden sich bei manchen Arten Erhebungen, die Basalloben genannt werden (beispielsweise bei Agnostida).
Der Begriff Spindel (oder Axis) ist nicht eindeutig. Manchmal werden mit diesem Ausdruck nur die Teile auf dem Thorax bezeichnet, gelegentlich werden die Teile auf Thorax und Schwanzschild (Pygidium) damit beschrieben. Die Spindel ist in verschiedene Spindelringe (auch Axialringe genannt) aufgeteilt. Die Furchen zwischen den Ringen werden Spindelfurchen (Axialfurchen) genannt.
Der transversale Lobus im Übergang von der Glabella zur Rhachis wird Nackenring oder Occipitalring genannt. Er ist bei den meisten Arten ausgeprägt und trägt manchmal einen spitzen Fortsatz (Tuberkel), welcher Occipitaltuberkel oder Nackenknötchen genannt wird.
Rhachis kommt aus dem griechischen und bedeutet eigentlich „Rückenmark“. Die Rhachis ist der Teil der Spindel auf dem Schwanzschild (Pygidium) und ist in verschiedene Rhachisringe aufgeteilt. Die Furchen zwischen den Ringen werden Rhachisfurchen genannt.
Pleuraler Lobus
Als pleuraler Lobus werden die linke und rechte Seite des Trilobiten bezeichnet. Sie gehen von der Spitze des Cephalons seitlich über die Freiwangen, wo sich häufig Facettenaugen befinden, seitlich über den Thorax und seitlich bis zur Spitze des Pygidiums. Die pleuralen Loben werden bei einigen Arten von Furchen auf dem Kopf- und dem Schwanzschild getrennt. Diese Furchen existieren zwischen dem Saum und der Spitze des Spindellobus. Diese Saumfurche wird auf dem Kopfschild mediane Präglabellarfurche und beim Schwanzschild mediane Postaxialfurche genannt. Bei manchen Arten sind diese Furchen verwachsen und nicht mehr zu erkennen.
Als Pleuron oder Pleure wird der rechte und der linke Teil eines Segmentes (auch Schiene) des Thorax bezeichnet. Die Enden eines Pleurons können je nach Art abgerundet sein oder spitzig enden. Furchen auf den Pleuren werden Pleuralfurche genannt.
Kopfschild (Cephalon)
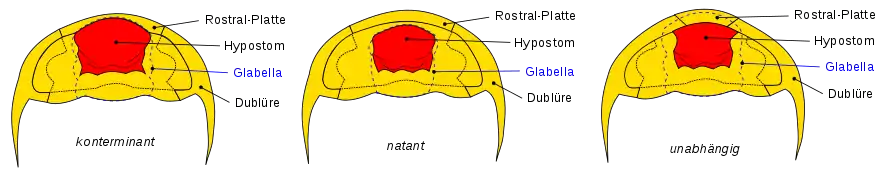
Auf der ventralen Seite des Kopfschildes befindet sich bei einigen Arten eine Rostralplatte, die auch Rostrum genannt wird (lat. für „Schnabel“). Sie kann als Befestigung für das Hypostom dienen. Das Hypostom ist eine Platte auf der Unterseite des Cephalons bei Trilobiten. Es war vermutlich Teil des Mundapparates. Die Form und die Positionierung des Hypostoms sind wesentliche Merkmale bei der systematischen Einteilung der Trilobitenarten.
- Konterminante Positionierung: Bei der sogenannten konterminanten Positionierung ist das Hypostom an der Rostralplatte befestigt. Die Vorderseite der Glabella auf der Oberseite ist bündig zur Vorderseite des Hypostoms.
- Natante Positionierung: Bei der sogenannten natanten („schwimmenden“) Positionierung liegt das Hypostom innerhalb der Unterseite des Cephalons und hat deshalb keine Verbindung mehr zur Rostralplatte. Auch bei dieser Positionierung ist die Glabella-Vorderseite bündig zur Hypostom-Vorderseite.
- Unabhängige Positionierung: Das Hypostom ist an den Dublüren befestigt, ist aber unabhängig von der Glabella positioniert. Die Glabella ist meistens länger und überlappt somit das Hypostom.
Die Trilobiten besaßen nur ein Paar spezialisierte Kopfanhänge, diese waren als lange Gliederantennen ausgebildet und dienten vermutlich als Sinnesorgane. Nur bei der Ordnung Agnostida traten kürzere, stark beborstete Antennen auf, die eher im Dienst der Nahrungsaufnahme gestanden haben könnten (unter anderem deshalb wird die Einbeziehung der Agnostida in die Trilobiten von etlichen Forschern bezweifelt). Die übrigen Gliedmaßen der Kopfsegmente, die gemeinsam vom Kopfschild überdeckt waren, entsprechen völlig den Spaltbeinen der Rumpfsegmente. Man nimmt deshalb an, dass die Beine der Trilobiten unspezialisiert sowohl der Fortbewegung wie auch der Nahrungsaufnahme gleichzeitig dienten. Entsprechende Verhältnisse sind bei vielen heute lebenden Krebstieren gegeben, zum Beispiel bei den meisten Blattfußkrebsen.
Gesichtsnaht
Erläuterung in Anlehnung an Lehmann, 2014[4]
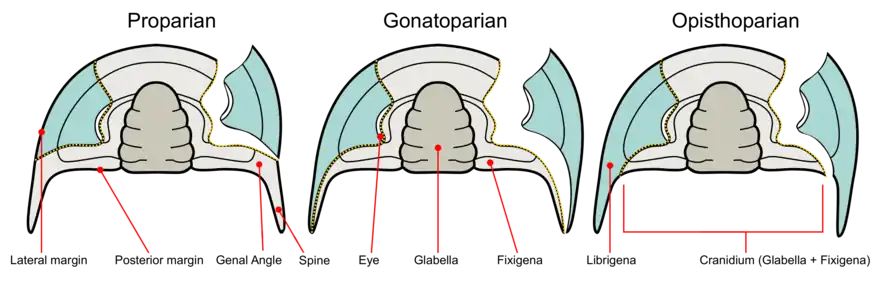
Ein weiteres diagnostisch bedeutsames Merkmal im Bereich des Cephalons ist die Gesichtsnaht (Sutura facialis). Die Gesichtsnaht ist eine Sollbruchstelle im Exoskelett des Cephalons, die es dem Trilobiten ermöglicht, bei der Häutung aus dem alten Panzer zu kriechen. Der Kopfschild zerfällt dabei in das Cranidium, d. h. die Glabella plus die Fixigenae (Sg. Fixigena; feste Wange oder Festwange), und die beiden Librigenae (Sg. Librigena; freie Wange oder Freiwange). Je nach Position und Verlauf der Gesichtsnaht werden mehrere Grundtypen unterschieden:
- Protopare oder hypopare Gesichtsnaht: In diesem Fall verläuft die Häutungsnaht nicht über die Wangen, sondern auf ganzer Länge entlang der Außenkante des Cephalons. Bei der Häutung wird also nur die Dublüre (Saumkante, jener Teil des Panzers der randlich in Richtung zur ungepanzerten Bauchseite umgebogen ist) des Kopfschildes abgetrennt. Freiwangen werden nicht ausgebildet. Dieses Merkmal tritt besonders häufig bei urtümlichen Trilobiten, z. B. innerhalb der Ordnung der Agnostida, (protopar) auf, aber auch sekundär bei einigen hochspezialisierten Formen etwa aus der Ordnung der Harpetida (hypopar).
- Propare Gesichtsnaht: Als propar wird eine Gesichtsnaht bezeichnet, die nur im vordersten Bereich des Cephalons entlang der Außenkante verläuft und dann auf die Oberseite des Kopfschildes übertritt, von dort weiter in Richtung zum Augenhügel (Palpebral-Lobus) und entlang der Innenseite der Facettenaugen zieht, um schließlich noch vor dem Wangeneck bzw. dem Wangenstachel zur Außenkante des Cephalons zurückzukehren.
- Gonatopare Gesichtsnaht: Bei der gonatoparen Gesichtsnaht ist der grundsätzliche Verlauf ähnlich. Die Häutungsnaht endet allerdings nicht an der Außenkante des Kopfschildes, sondern direkt im Wangeneck bzw. an der Spitze des Wangenstachels.
- Opisthopare Gesichtsnaht: Eine opisthopare Gesichtsnaht unterscheidet sich von Proparen und gonatoparen Gesichtsnähten dadurch, dass die Häutungsnaht erst nach dem Wangeneck bzw. dem Wangenstachel, also nicht an der Außenkante, sondern an der Hinterkante des Kopfschildes endet.
- Metapare Gesichtsnaht: Eine metapare Gesichtsnaht unterscheidet sich grundlegend von den übrigen Typen. Hier beginnt die Häutungsnaht an der Hinterkante des Kopfschildes, verläuft in Richtung zum Augenhügel und wieder zurück zu einem ebenfalls an der Hinterkante des Kopfschildes befindlichen Austrittspunkt.
Augen
Nicht alle Trilobitenarten haben Augen ausgebildet. Sind Augen vorhanden, so handelt es sich um Facettenaugen, die wie das Exoskelett aus Calcit bestehen. Somit sind diese Augen nicht direkt mit denjenigen der heutigen Gliederfüßer vergleichbar. Die meisten Forscher nehmen aber an, dass sie zu den Facettenaugen der übrigen Arthropoden homolog sind. Da es sich bei Calcit um anorganisches Material handelt, sind die Facettenaugen bei den fossilierten Exuvien und Individuen sehr gut erhalten geblieben, da sie nicht von Mikroorganismen zersetzt wurden.
Die Augen treten in drei Formen auf: Holochroale, schizochroale oder abathochroale Facettenaugen.[5]
- Bei den holochroalen Facettenaugen sind die Einzelaugen eng aneinandergereiht ohne eine Sclera dazwischen. (Sclera ist zusätzliches Exoskelett-Material, welches eine ähnliche Funktion wie die Sclera des Säugetierauges hat.) Die Hornhaut (Cornea) bedeckt sämtliche Einzelaugen gleichzeitig. Es sind bis zu 15.000 Einzelaugen vorhanden.
- Bei den schizochroalen Facettenaugen sind die Einzelaugen mit einer ausgeprägten dicken Sclera getrennt, welche die Augen fassen. Jedes Einzelauge hat eine eigene Hornhaut, die ebenfalls durch die Sclera gefasst ist und tiefer ins innere Exoskelett hineinreicht. Es sind bei dieser Facettenaugenart nur bis zu 700 Einzelaugen vorhanden.
- Bei den abathochroalen Facettenaugen kommt ebenfalls eine Sclera vor. Diese ist aber einiges dünner als bei den schizochroalen Facettenaugen und höchstens gerade so dick wie die Einzelaugen. Wie bei den schizochroalen Augen hat jedes Einzelauge eine eigene Hornhaut. Diese endet aber bereits bei Beginn der Sclera.
 Holochroale Facettenaugen bei Paralejurus
Holochroale Facettenaugen bei Paralejurus Schizochroale Facettenaugen bei Erbenochile erbeni
Schizochroale Facettenaugen bei Erbenochile erbeni
Thorax
Der Thorax besteht aus Segmenten. Die Anzahl der Segmente und die Form ist systematisch relevant. Kleine Trilobiten der Agnostida haben nur zwei oder drei Segmente. Bei größeren Trilobitenarten sind bis zu 18 Stück vorhanden. Bei Tieren mit besonderer Lebensweise können die Segmente auch unterschiedliche Formen annehmen. So haben sie etwa stachelige Erweiterungen, vermutlich um sich von Fressfeinden zu schützen. Andererseits können sie wie bei den Arten von Paralejurus gebogen sein, um die vermutlich grabende Tätigkeit zu erleichtern.
Spaltbeine
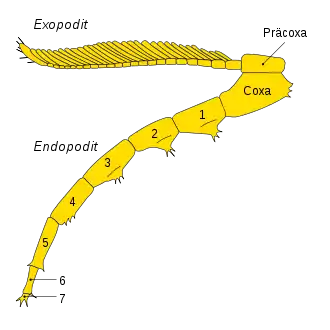
Da nur die Oberseite der Trilobiten hartschalig und verkalkt war, sind Fossilien, die die weichhäutige Unterseite mit den Beinen umfassen, sehr selten und weltweit nur in wenigen Fossillagerstätten gefunden worden. Trilobiten hatten sogenannte zweiästige Spaltbeine. Der erste Ast heißt Schwimm- oder Kiemenbein (Exopodit) und diente der schwimmenden Bewegung im Meer. Der zweite Ast heißt Laufbein (Endopodit) und diente dem Gehen auf dem Meeresgrund.
- Das Laufbein bestand aus der Coxa und sieben weiteren Gliedern. Die Coxa war an der Präcoxa befestigt. Die Glieder konnten zusätzliche Stacheln aufweisen.
- Das Schwimmbein war an der Präcoxa befestigt und bestand je nach Art aus zwei bis mehreren Gliedern. Am letzten Glied oder mehreren dieser Glieder befanden sich fächerartige Erweiterungen, die ein Paddeln im Wasser ermöglichten.
Entwicklung
Trilobiten entwickelten sich über eine Vielzahl von Stadien, bei der Segmente bei jeder Häutung in einer Wachstumszone unmittelbar vor dem Hinterende eingeschoben wurden (Anamorphose). Die meisten Trilobitenarten besaßen als Adulti eine fixierte Anzahl von Segmenten, bei einigen sehr beinreichen Taxa schritt die Einfügung neuer beintragender Segmente aber offenbar auch nach der Geschlechtsreife bis zum Tode fort. Die Entwicklung der Trilobiten ist bei vielen Arten durch zahlreiche Funde verschieden großer Tiere, die sich zu Serien anordnen lassen, und durch Funde von Exuvien (nach einer Häutung zurückbleibenden Hüllen) recht gut bekannt. In der Regel besaßen auch die Larvenstadien eine verkalkte Dorsalhülle (Ausnahme bei den Agnostida und möglicherweise weiteren Gruppen).
Das erste Larvenstadium der Trilobiten wird Protaspis genannt, es bestand aus vier gliedmaßentragenden Kopf-Somiten, einem kurzen Kopfschild und einer knospenartigen Anlage des Thorax. Spätere Stadien lassen zwei Regionen (Tagma), nämlich Kopf und Rumpf, erkennen, sie werden Meraspis genannt. Eine Region am Hinterende konnte ein transitorisches Schwanzschild (Pygidium) ausbilden. Neue Segmente entstanden in der Nähe von dessen Hinterende, wanderten bei weiteren Häutungen durch die Struktur hindurch und wurden am vorderen Ende zu freien Rumpfsegmenten abgeschnürt. Schließlich wurde ein Stadium erreicht, das keine neuen Segmente mehr ausbildete, es wird Holaspis genannt. Tiere im Holaspis-Stadium haben sich noch gehäutet und konnten noch, teilweise erheblich, an Größe gewinnen. Wann genau die Geschlechtsreife der Tiere eintrat, ist am Fossilmaterial nicht zu erkennen. In der Regel sind aber ausgewachsene Exemplare mit klar definierter Segmentzahl, Form und Größe klar erkennbar.
Lebensweise
Trilobiten waren in der Regel Bewohner des Meeresbodens (Benthos). Die meisten Fossilien liegen aus küstennahen (litoralen) Habitaten, z. T. auch aus mittleren Meerestiefen (Schelfgebiete) vor. Tiefseeformen existierten offensichtlich keine. Die Existenz von schwimmenden (pelagialen) Trilobiten erscheint hoch wahrscheinlich, einige ordovizische Formen wiesen stromlinienförmige Körper auf und könnten schnelle, aktive Schwimmer gewesen sein.[6] Einige lebten in sauerstoffarmen, schlammigen Sedimenten. Wenige Formen konnten offenbar Tunnel in das Sediment graben.[7] Die meisten Trilobiten ernährten sich vermutlich als Räuber und/oder Aasfresser. Dies ist vermutlich auch die ursprüngliche Lebensweise. Abgeleitete Formen weisen eine Morphologie auf, die Hinweise auf eine Lebensweise als Detritus-/Sedimentfresser oder als Filtrierer aufweisen.[8] Einige waren offenbar Weidegänger, die Mikrobenmatten und anderen Aufwuchs vom Meeresboden abweideten. Trilobiten wurden selbst offensichtlich auch zur Beute anderer Meeresbewohner. Aus dem mittleren Kambrium liegt z. B. der Fund eines Arthropoden aus einer ausgestorbenen Linie (vermutlich ein basaler Vertreter der Chelicerata) vor, dessen Darminhalt aus zahlreichen Trilobiten bestand.[9]
Trilobiten als Zeugen der Evolution

_4_(15402497771).jpg.webp)

Fossilien von Trilobiten erscheinen im Kambrium mit Beginn der (bisher nur provisorisch benannten und abgegrenzten) 2. Serie in der fossilen Überlieferung. Der Beginn der 2. Serie ist mehr oder weniger durch dieses Ereignis definiert worden. Damit ist ihr Erscheinen um die gesamte Dauer der ersten Serie (inzwischen Terreneuvium genannt) von den einschneidenden Ereignissen, die den Beginn des Kambriums markieren (der „kambrischen Explosion“) getrennt, das sind etwa 13 Millionen Jahre. Die ältesten Trilobiten sind gleichzeitig die ältesten unzweideutigen Körperfunde von Arthropoden überhaupt. In den fossilen Funden der ersten Serie, als Small-Shelly-Fauna bezeichnet, wären Reste von Trilobiten, zumindest von disartikulierten Segmenten oder Platten, zu erwarten, wenn diese in dieser Zeit häufig oder weit verbreitet gewesen wären. Die ältesten Trilobiten[10] treten interessanterweise in verschiedenen Erdregionen (die den Flachmeeren der damaligen Kontinentalplatten des auseinanderbrechenden Superkontinents Rodinia entsprechen) grob gleichzeitig auf, aber mit zwar erkennbar verwandten, aber doch deutlich verschiedenen Formen.[11] Dieses vikariierende Verbreitungsmuster zeigt, dass der tatsächliche Ursprung der Trilobiten älter sein muss, da ihm die Ausbreitung in die anderen Meeresregionen und die anschließende evolutive Auseinanderentwicklung ja vorangegangen sein müssen. Diese nur erschlossene, aber nicht durch Fossilien belegbare Existenzperiode (als „ghost range“ bezeichnet) wurde auf in der Größenordnung etwa 10 Millionen Jahre geschätzt.[12]
Trilobiten zeichnen sich im Fossilbericht durch ihre verkalkten Rückenpanzer aus, die ihre Fossilierung erheblich wahrscheinlicher machen. In den weltweit nur sehr wenigen Lagerstätten, in denen Tiere mit Weichteilerhaltung aus den entsprechenden Epochen gefunden worden sind (sog. Konservatlagerstätten), ist inzwischen ein ganzer Zoo von gleichzeitig lebenden anderen Arthropoden gefunden worden, die zum Teil den Trilobiten ausgesprochen ähnlich waren, nur dass ihr Integument nicht verkalkt war. In der Chengjiang-Faunengemeinschaft aus China, deren Beginn nahe dem erstmaligen Auftreten von Trilobiten lag (tatsächlich wurden einige der ältesten aus der Gattung Parabadiella hier in den wenig älteren liegenden Gesteinsschichten gefunden) sind Arthropoden arten- und individuenreich vertreten. Von den etwa 50 Arten sind nur fünf Trilobiten, die kaum 2 % der Individuen ausmachen.[13][14] Die unverkalkten, trilobiten-ähnlichen Gruppen werden gemeinsam mit den Trilobiten unter den Namen Trilobitoidea, Trilobitomorpha oder Lamellipedia zusammengefasst (jeder dieser Namen umfasst ein etwas unterschiedliches, aber im Kern vergleichbares Konzept).[15] Alle wiesen einen dreigeteilten Körperbau mit hohem Mittelabschnitt und zwei seitlichen Verbreiterungen, einen Kopfschild aus mehreren verschmolzenen Segmenten, einästige (unirame) Antennen und zweiästige (birame) Spaltbeine auf, deren Innenast (Endopodit) ein Laufbein war und dessen Außenast (Exopodit) aus drei Segmenten bestand, deren erstes einen Saum aus langen, lamellenartig verbreiterten Borsten aufwies (bei einigen Vertretern/Gruppen umstritten oder im Fossilmaterial nicht sicher nachweisbar). Fast alle wiesen auch ein Schwanzschild (Pygidium) auf. Die Trilobiten unterscheiden sich von vielen Vertretern im Wesentlichen nur in zwei Merkmalen: der verkalkten Dorsalhülle und den oben (dorsal) liegenden Augen. Direkte Schwestergruppen der Trilobiten sind z. B. die Helmetiida, die Naraoiidae, die Emucarididae[16] (Nectopleura oder Nektaspida) und die Tegopeltida, andere Gruppen wie die Xandarellida sind etwas besser abgrenzbar, aber ebenfalls sehr ähnlich. Die Trilobiten sind letztlich also nur die bekannteste und fossil am besten überlieferte Gruppe einer ursprünglich weitläufigen Verwandtschaft.
Interessanterweise treten einige früher als typisch für Trilobiten geltende Merkmale bei weiteren Fossilien derselben Epoche ebenfalls auf.[17] So weisen viele Vertreter einer Gruppe, die nach einem Merkmal, einem deutlich segmentierten und stark beborsteten ersten Beinpaar, das wohl als Greiforgan für eine räuberische Lebensweise diente, als „great appendage arthropods“ (oder „Megacheira“) zusammengefasst werden, einen sehr ähnlichen dreilappigen Körperbau auf. Diese Gruppe ist in ihrer Verwandtschaft nicht unumstritten, gilt aber meist als Wurzelgruppe der Spinnentiere. Eine weitere Gruppe von fossilen Gliederfüßern mit noch urtümlicheren Merkmalen wie Canadaspis und Fuxianhuia war im Körperbau nicht weit entfernt davon, diese Tiere gelten wahlweise als Vertreter der Stammgruppe der Krebstiere oder sogar der gemeinsamen Stammgruppe aller Arthropoden.[18] Nicht nur der gleichförmige (repetitive) Körperbau mit wenig differenzierten Beinen, sondern zahlreiche weitere Merkmale wie der dreilappige Körperbau, das Spaltbein, Kopf- und Schwanzschild sind damit als gemeinsames Erbe der Arthropoden nachgewiesen. Sie sind bei den Trilobiten zwar zuerst gefunden worden, aber dieser Gruppe nicht eigentümlich, sondern sind von ihren Vorfahren vererbte Plesiomorphien.
Die Verwandtschaft der Trilobiten mit den lebenden (rezenten) Arthropodenordnungen der Spinnentiere und der Krebstiere ist mit diesen neuen Erkenntnissen zu einer offenen Frage geworden. Sie hängt in kritischer Weise von der Interpretation der Homologie verschiedener Körpersegmente, insbesondere des Kopfes[19] und der allerersten Extremitätenpaare, ab. Ob sie gemeinsam mit den Spinnentieren eine Arachnata (oder Arachnomorpha) genannte Gruppe bilden, was der traditionellen Auffassung entspricht, oder doch eher nähere Verwandte der Krebstiere sind, ist vor allem von der Interpretation der Lage des Antennensegments bei zahlreichen fossilen Vertretern abhängig. Die Antennen der Trilobiten sind eindeutig sekundär, d. h., sie sind Extremitäten desjenigen Kopfsegments, das den zweiten Gehirnabschnitt (Deutocerebrum) bildet, sie saßen am Kopflappen (Hypostom) an. Die Antennen der rezenten Stummelfüßer (Onychophora), die als Verwandte der Arthropoden gelten, sitzen hingegen am ersten Kopfabschnitt (der den ersten Gehirnabschnitt bildet). Die Lage der Antennen zahlreicher Fossilien ist umstritten. Je nach Interpretation verschieben sich dann die erschlossenen Verwandtschaftsverhältnisse.
Die spätere Evolution der Trilobiten bringt zwar zahlreiche Neuerungen in Körperbau und Lebensweise, aber eine im Kern fast unveränderte Morphologie. Die allerspätesten fossilen Trilobiten sehen den kambrischen Vertretern ausgesprochen ähnlich. Dem tatsächlichen Aussterben waren bereits einige „Beinahe-Aussterben“ bei früheren Aussterbewellen vorangegangen, die aber von wenigen Arten überlebt worden waren, die sich anschließend wieder differenzieren konnten. Während aber im Kambrium und Ordovizium jeweils zahlreiche neue Arten mit unterschiedlicher Lebensweise entstanden waren, konnten in den späteren Epochen die überlebenden Arten sich nicht mehr zu vergleichbarer Vielfalt entwickeln. Vor ihrem endgültigen Aussterben waren die Trilobiten bereits seit fast 100 Millionen Jahren eine artenarme Reliktgruppe. Einige gut adaptierte Arten konnten offensichtlich in speziellen ökologischen Nischen überleben, die dann bei den mit dem Massenaussterben an der Perm-Trias-Grenze zusammenhängenden Ereignissen verloren gingen (schätzungsweise sind bei diesem Ereignis 95 % aller damals lebenden marinen Wirbellosen ausgestorben).
Ein Überleben einiger Vertreter in unzugänglichen Lebensräumen wie der Tiefsee erscheint so gut wie ausgeschlossen. Es wird vor allem von amerikanischen Kreationisten immer wieder ins Gespräch gebracht, die damit hoffen, Zweifel an der Evolutionstheorie säen zu können.[20]
480 Mio. Jahre alte Fossilien der Art Ampyx priscus aus dem unteren Ordovizium, welche eine Reihenformation der Tiere belegen, wurden als die ersten Zeugnisse von Schwarmintelligenz bei Lebewesen gedeutet.[21]
%252C_linear_cluster_%E2%80%93_480_mya%252C_Morocco.png.webp)
Trilobiten als Leitfossilien
Trilobiten zählen zu den wichtigsten Leitfossilien der Erdgeschichte. Ihre Überreste werden zur relativen Altersbestimmung von Sedimentgesteinen genutzt. Diese Methodik nennt man Biostratigraphie. Bestimmte Trilobitenarten kommen nur in engen zeitlichen Abschnitten vor, sind also für die Ablagerungen dieser Zeit kennzeichnend. So lassen sich die Entstehungszeiträume der Sedimentgesteine bestimmen. Trilobiten sind nur in Gesteinen des Erdaltertums (Paläozoikum) fossil überliefert. Zu den ältesten Trilobiten, die sich gut erhalten haben, zählen die Arten der Gattung Ellipsocephalus, ein Trilobit mit ovalem Kopf. Sie werden auf der ganzen Welt gefunden.
Ordnungen
Gegenwärtig sind neun Ordnungen der Klasse der Trilobiten anerkannt:
- Agnostida: Sehr kleine und kleinste Trilobiten, meist nur wenige Millimeter lang. Zeitalter: Unteres Kambrium bis Oberes Ordovizium.
- Redlichiida: Sehr alte Trilobiten mit vielen, in Pleuralstacheln auslaufenden Thoraxsegmenten. Zeitalter: Unteres Kambrium bis Mittleres Kambrium.
- Corynexochida: Verlängerte Glabella (Kopfpanzer) oft mit konkav laufenden Seiten, in aller Regel gut ausgeprägte Augen. Zeitalter: Unteres Kambrium bis Mittleres Devon.
- Lichida: Zumeist stachelige Trilobiten. Zeitalter: Kambrium bis Devon.
- Phacopida: Vielfältige Gruppe mit zahlreichen Erscheinungsformen. Zeitalter: Unteres Ordovizium bis Oberes Devon.
- Proetida: Normalerweise recht kleine Trilobitenformen. Zeitalter: Ordovizium bis Perm.
- Asaphida: Vielfältige Gruppe mit zahlreichen Erscheinungsformen. Zeitalter: Mittleres/Oberes Kambrium bis zum Oberen Ordovizium/Unterem Silur.
- Harpetida: Haupterkennungsmerkmal: Extrem großer Cephalonsaum. Zeitalter: Oberes Kambrium bis spätes Devon.
- Ptychopariida: Große Gruppe mit unausgereifter Klassifizierung. Zeitalter: Unteres Kambrium bis Oberes Ordovizium.
Fundregionen in Deutschland und Europa

- Perm: in Deutschland nicht berichtet; Europa: (?)
- Karbon: Rheinisches Schiefergebirge, Harz, Frankenwald; Karnische Alpen, sonst in Großbritannien, Frankreich
- Devon: Rheinisches Schiefergebirge, Harz, Eifel, speziell die Trilobitenfelder bei Gees; sonst in den Karnischen Alpen, in Großbritannien, Frankreich, Barrandium, Polen
- Silur: im Geschiebe in Deutschland; sonst in Schweden, Großbritannien, Karnischen Alpen, Barrandium, Estland
- Ordovizium: im Geschiebe in Deutschland, selten im Thüringer Wald und Frankenwald; sonst in Schweden, Norwegen, Bornholm, Estland, Polen, Barrandium, Frankreich, Großbritannien, Spanien und Portugal
- Kambrium: im Geschiebe in Deutschland, selten in der Lausitz und im Frankenwald; sonst hauptsächlich im Barrandium und in Schweden, Norwegen sowie auf Bornholm
Verwendung als Wappentiere
Gehäuse bzw. Schalen von Trilobiten kommen in der Heraldik selten als Wappentiere vor; ihre Verwendung deutet auf lokale Fundstätten hin.




.svg.png.webp)
Siehe auch
Literatur
- Richard Fortey: Trilobiten! Fossilien erzählen die Geschichte der Erde. C.H. Beck, München 2002, ISBN 3-406-49592-3.
- Harry Blackmore Whittington: The Burgess Shale. Yale University Press, New Haven u. a. 1985, ISBN 0-300-03348-6.
Einzelnachweise
- Diese stammen aber nicht immer von Trilobiten. Vgl. John-Paul Zonneveld, S. George Pemberton, Thomas D. A. Saunders, Ronald K. Pickerill: Large, Robust Cruziana from the Middle Triassic of Northeastern British Columbia: Ethologic, Biostratigraphic, and Paleobiologic Significance. In: Palaios. Band 17, Nr. 5, 2002, S. 435–448, doi:10.1669/0883-1351(2002)017<0435:LRCFTM>2.0.CO;2.
- Johann Ernst Immanuel Walch: Die Naturgeschichte der Versteinerungen. Zur Erläuterung der Knorrischen Sammlung von Merkwürdigkeiten der Natur. Theil 3. Felßecker, Nürnberg 1771.
- Robert Kihm, James St.John: Walchs trilobite research. In: Donald G. Mikulic, Ed Landing, Joanne Kluessendorf (Hrsg.): Fabulous Fossils. 300 years of worldwide research on trilobites (= New York State Museum. Bulletin. 507). University of the State of New York – State Education Department, Albany NY 2007, ISBN 1-55557-235-9, S. 115–140.
- U. Lehmann: Paläontologisches Wörterbuch. unveränderter Nachdruck der 4. Auflage, 278 S., Springer, 2014.
- Übersicht in: Euan Clarkson, Riccardo Levi-Setti, Gabor Horvath: The eyes of trilobites: The oldest preserved visual system. In: Arthropod Structure & Development. Band 35, Nr. 4, 2006, S. 247–259, doi:10.1016/j.asd.2006.08.002.
- Richard A. Fortey: Pelagic trilobites as an example of deducing the life habits of extinct arthropods. In: Transactions of the Royal Society of Edinburgh. Earth Sciences. Band 76, Nr. 2/3, 1985, S. 219–230, doi:10.1017/S0263593300010452.
- Lesley Cherns, James R. Wheeley, Lars Karis: Tunneling trilobites: Habitual infaunalism in an Ordovician carbonate seafloor. In: Geology. Band 34, Nr. 8, 2006, S. 657–660, doi:10.1130/G22560.1.
- Richard A. Fortey, Robert M. Owens: Feeding habits in trilobites. In: Palaeontology. Band 42, Nr. 3, 1999, S. 429–465, doi:10.1111/1475-4983.00080.
- Mao-Yan Zhu, Jean Vannier, Heyo Van Iten, Yuan-Long Zhao: Direct Evidence for Predation on Trilobites in the Cambrian. In: Proceedings of the Royal Society of London. Series B: Biological Sciences. Band 271, Supplement 5, 2004, S. S277–S280, doi:10.1098/rsbl.2004.0194.
- J. Stuart Hollingsworth: The first trilobites in Laurentia and elsewhere. In: Isabel Rábano, Rodolfo Gozalo, Diego García-Bellido (Hrsg.): Advances in Trilobite Research (= Cuadernos del Museo Geominero. 9). Instituto Geológico y Minero de España, Madrid 2008, ISBN 978-84-7840-759-0, S. 171–175.
- Jinling Yuan, Xuejian Zhu, Jihpei Lin, Maoyan Zhu: Tentative correlation of Cambrian Series 2 between South China and other continents. In: Bulletin of Geosciences. Band 86, Nr. 3, 2011, S. 397–404, doi:10.3140/bull.geosci.1274.
- Bruce S. Lieberman: Taking the Pulse of the Cambrian Radiation. In: Integrative and Comparative Biology. Band 43, Nr. 1, 2003, S. 229–237, doi:10.1093/icb/43.1.229.
- Xianguang Hou, Jan Bergström: The Chengjiang fauna – the oldest preserved animal community. In: Paleontological Research. Band 7, Nr. 1, 2003, S. 55–70, doi:10.2517/prpsj.7.55.
- James W. Hagadorn: Chengjiang. Early Record of the Cambrian Explosion. In: David H. Bottjer, Walter Etter, James W. Hagadorn, Carol M. Tang (Hrsg.): Exceptional Fossil Preservation. A Unique View on the Evolution of Marine Life. Columbia University Press, New York NY 2002, ISBN 0-231-10255-0, S. 35–60, (Digitalisat (PDF; 6,55 MB)).
- Jan Bergström, Xian-Guang Hou: Arthropod origins. In: Bulletin of Geosciences. Band 78, Nr. 4, 2003, S. 323–334, (Digitalisat (PDF; 270,32 KB)).
- John R. Paterson, Gregory D. Edgecombe, Diego C. Garcia-Bellido, James B. Jago, James G. Gehring: Nektaspid arthropods from the Lower Cambrian Emu Bay Shale Lagerstätte, South Australia, with a reassessment of lamellipedian relationships. In: Palaeontology. Band 53, 2010, S. 377–402, doi:10.1111/j.1475-4983.2010.00932.x.
- Übersicht in: Nigel C. Hughes: The Evolution of Trilobite Body Patterning. In: Annual Review of Earth and Planetary Sciences. Band 35, 2007, S. 401–434, doi:10.1146/annurev.earth.35.031306.140258.
- Graham E. Budd, Maximilian J. Telford: The origin and evolution of arthropods. In: Nature. Band 457, Nr. 7231, 2009, S. 812–817, doi:10.1038/nature07890.
- Gerhard Scholtz. Gregory D. Edgecombe: The evolution of arthropod heads: reconciling morphological, developmental and palaeontological evidence. In: Development Genes and Evolution. Band 216, Nr. 7/8, 2006 = Special Issue: Development and Phylogeny of Arthropods. S. 395–415, doi:10.1007/s00427-006-0085-4.
- Linda A. Ritterbush: A History of Trilobites as „Living Fossils“. In: Perspectives on Science and Christian Faith. Band 56, Nr. 2, 2004, S. 131–134, (Digitalisat (PDF; 330,86 KB)).
- Verhaltensbiologie: Fossilien belegen Schwarmintelligenz schon vor 480 Millionen Jahren. Abgerufen am 18. Oktober 2019 (Artikel über die englische Veröffentlichung: https://doi.org/10.1038/s41598-019-51012-3).