Vögel
Die Vögel sind – nach traditioneller Taxonomie – eine Klasse der Wirbeltiere, deren Vertreter als gemeinsame Merkmale unter anderem Flügel, eine aus Federn bestehende Körperbedeckung und einen Schnabel aufweisen. Vögel leben auf allen Kontinenten. Bislang sind über 10.758 existente rezente Vogelarten bekannt, zusätzlich sind 158 Arten in historischer Zeit ausgestorben.[1] Bei Anwendung des moderneren, phylogenetischen Artkonzepts kommt man auf etwa 18.000 (15.845 bis 20.470) Vogelarten, und auch genetische Daten deuten auf eine in etwa doppelt so hohe Anzahl von Vogelarten hin.[2] Die Wissenschaft von den Vögeln ist die Vogelkunde (Ornithologie).
| Vögel | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
.jpg.webp)
Eilseeschwalbe (Thalasseus bergii) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Aves | ||||||||||||
| Linnaeus, 1758 | ||||||||||||
| Unterklassen | ||||||||||||
|
Allgemeine Kennzeichen
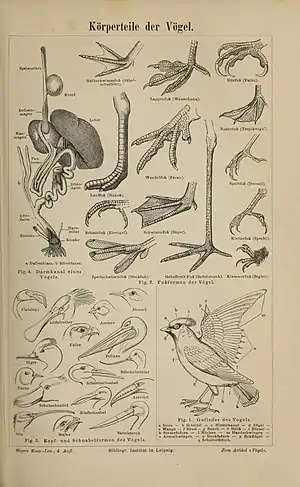
- Vögel haben wie alle Landwirbeltiere (Tetrapoda) zwei Paar Extremitäten, wovon die vorderen bei Vögeln zu Flügeln umgebildet sind.
- Das Gefieder bestimmt das äußere Gesamtbild der Vögel wesentlich: Der Körper ist von Federn bedeckt. Diese Strukturen aus Keratin dienen beim Fliegen als Tragfläche und Steuerfläche, einer aerodynamisch günstigen Verkleidung des Körpers und als Isolation, die sogar, meist je nach Temperatur und Wind, veränderbar ist. Ferner hat das Gefieder Farben und dient oft der sexuellen Werbung. Bei Wasservögeln ist es wasserabweisend und sorgt für Auftrieb. Das Gefieder wird zu bestimmten Zeiten (Mauser) gewechselt. Des Weiteren sind bei allen Vögeln die Beine durch Hautschuppen bedeckt.
- Alle rezenten Vögel besitzen einen Schnabel ohne echte Zähne. Einige ausgestorbene Arten waren dagegen bezahnt. Der Schnabel besteht bei Vögeln aus Knochensubstanz mit einer Hornscheide; nur bei Tauben- (Columbiformes) und Gänsevögeln (Anseriformes) ist die Knochenmasse des Schnabels von einer weichen Haut überzogen.
- Alle bekannten Vogelarten haben eine relativ hohe, konstante Körpertemperatur (Endothermie), die höher ist als bei allen anderen heute lebenden Tieren und ca. 42 °C beträgt. Einige Vogelarten, z. B. Kolibris und Kohlmeisen, senken bei der Nachtruhe ihre Körpertemperatur um ca. 10 °C.
- Die meisten Vögel sind flugfähig. Alle flugunfähigen Arten haben sich aus Arten entwickelt, die ursprünglich fliegen konnten. Dies betrifft zum Beispiel Laufvögel, Pinguine und viele Inselformen, wie die Kiwis (Neuseeland) und den Stummelkormoran (Galapagosinseln).
- Das Vogelskelett ist leicht gebaut. Es besitzt zur Gewichtsreduzierung hohle (pneumatisierte) Knochen. Der Anteil der Knochenmasse macht nur 8 bis 9 Prozent der Gesamtkörpermasse aus, während er bei einigen Säugern bis zu 30 Prozent betragen kann. Das sehr große Brustbein hat einen vorspringenden Kiel (Carina), der als Ansatz für die sehr großen Flugmuskeln dient.
- Die Herzschlagfrequenz ist hoch: Die maximale Herzschlagfrequenz eines Straußes beträgt 178 Schläge pro Minute, diejenige eines Haussperlings 900 und schließlich diejenige eines Blaukehl-Sternkolibris 1.260 Schläge pro Minute.
- Das Zentralnervensystem (ZNS) ist hoch entwickelt, unter den Sinnesorganen ist besonders die Leistungsfähigkeit des Auges bemerkenswert. Einige Vögel sind wahrscheinlich Pentachromaten, haben also fünf Typen von Farbsinneszellen (Zapfen). Das für die Verarbeitung optischer Reize zuständige Hirnareal ist stark vergrößert.
- Die Stimmbildung erfolgt bei Vögeln nicht im Kehlkopf (Larynx), da Stimmbänder fehlen. Dafür liegt an der Gabelung der Trachea ein gesondertes Organ, der Stimmkopf (Syrinx), auch als unterer Kehlkopf bezeichnet.
- Die Vogellunge mit ihren Luftsäcken ist komplizierter gebaut als die aller anderen Wirbeltiere.
- Vögel haben eine Kloake, das heißt, Eier, Urin und Kot gelangen durch eine einheitliche Ausführungsöffnung ins Freie.
- Die Elektrolytausscheidung (Meersalze) erfolgt bei vielen Vögeln wie bei Reptilien auch über die Nasendrüse.
- Vögel scheiden ebenso wie Fledertiere und Reptilien über ihren Urin Stickstoffverbindungen als Guanin sowie Harnsäure aus. Guanin ist zwar energiereicher als Harnstoff, benötigt aber kaum Wasser zur Ausscheidung, sodass die Tiere nicht so viel Trinkwasser wie Säugetiere benötigen und das Wasser im Körper nicht mitgeführt werden muss. Diese Ersparnis an zu bewegender Masse unterstützt die Flugfähigkeit. Ablagerungen aus Vogelkot können abbauwürdige Mächtigkeiten erreichen. Sogenannter Inselguano, der aus Seevogelausscheidungen besteht, wird ebenso wie Höhlenguano (meist aus Fledermauskot bestehend) abgebaut und als phosphatreiches natürliches Düngemittel eingesetzt.
- Wie bei Reptilien fehlt eine Harnblase.
- Die meisten Vogelarten besitzen zur Gefiederpflege eine besondere, Fett absondernde Drüse, die Bürzeldrüse. Bei einigen Arten wird deren Funktion durch sogenannte Puderdunen unterstützt bzw. komplett ersetzt (Kakadus, Taubenvögel, Reiher). Einigen Arten fehlen sowohl Bürzeldrüse als auch Puderdunen (z. B. Schlangenhalsvögel).
- Von den oben genannten Merkmalen kommt keines exklusiv bei den Vögeln vor. So existieren (oder existierten) fliegende Formen bei den Säugetieren (Fledertiere) und Reptilien (Flugsaurier), über Federn verfügten auch die gefiederten Nichtvogeldinosaurier, amniotische Eier werden ebenso von Reptilien und Kloakentieren gelegt und auch ein Schnabel ist nicht auf die Vögel beschränkt.
- Bei den Vögeln sind Oberkiefer (Maxilla) und Unterkiefer (Mandibula) beweglich, während sich bei den anderen Wirbeltieren nur der Unterkiefer bewegt.
Systematik
Äußere Systematik
Die Vögel werden so wie die Amphibien, Reptilien und Säugetiere traditionell als eigene Klasse von Landwirbeltieren (Tetrapoda) betrachtet. Dies ist allerdings aus moderner, kladistischer Sichtweise betrachtet falsch, da die Vögel als einzige überlebende Gruppe der Klade Dinosaurier eine Teilgruppe der Reptilien sind.[3] Um eine einheitliche Terminologie zu bewahren, wird das monophyletische Taxon, welches die traditionellen Klassen Reptilien und Vögel vereint, als Sauropsida bezeichnet. Dementsprechend heißen die drei Klassen der Landwirbeltiere nach heutigem wissenschaftlichem Standard:
- Amphibien (Amphibia)
- Sauropsiden (Sauropsida)
- Säugetiere (Mammalia)
Ordnungen und Familien der Vögel
Die Klasse der Vögel ist die artenreichste der Landwirbeltiere. Sie umfasst etwa 10.700 rezente und etwa 150 in historischer Zeit ausgestorbene Vogelarten.[1] Von ihren zwei Unterklassen (Urkiefer- und Neukiefervögel) ist erstere klein und bis auf fünf Familien ausgestorben. Hierhin gehören insbesondere die Laufvögel, die wegen des fehlenden Brustbeinkamms flugunfähig sind. Von den weiteren etwa 30 Ordnungen der Neukiefervögel umfasst jene der Sperlingsvögel fast 60 Prozent aller Arten. In dieser Gruppe ist wiederum die Unterordnung der Singvögel (Passeri) die umfangreichste.
Unter Pygostylia findet sich eine abweichende Systematik der Vögel, die auch ausgestorbene Gruppen berücksichtigt.
Evolution
Abstammung von Dinosauriern
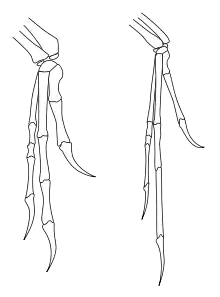
links der Maniraptor Deinonychus, rechts Archaeopteryx (verschiedene Maßstäbe, Deinonychus ist deutlich größer als Archaeopteryx)
Allgemein unstrittig ist, dass die Vögel der diapsiden Entwicklungslinie der Amnioten entstammen (siehe auch Reptilien). Als ihre nächsten lebenden Verwandten gelten die Krokodile. Gemeinsam bilden Krokodile und Vögel die Diapsiden-Untergruppe der Archosauria.
Die Skelette der ersten Vögel stammen aus Ablagerungen des Oberjura (ca. 150 Mio. Jahre vor heute) und zeigen nach Ansicht der Mehrzahl der Forscher eine enge verwandtschaftliche Beziehung zu kleinen Raubdinosauriern (Theropoden) aus der Gruppe der Maniraptora. Nach den Regeln der Kladistik sind Vögel damit ebenfalls Dinosaurier. Die Maniraptora galten allerdings lange als reine Bodenläufer, da ihr Fossilbericht, speziell die Ausbildung der Gliedmaßen aller bis dahin bekannten Vertreter, eine solche Lebensweise nahelegte. Die Anhänger einer Nicht-Dinosaurier-Abstammung der Vögel weisen deshalb unter anderem darauf hin, dass fliegende oder gleitfliegende Tiere immer von baumlebenden Vorfahren abstammten, die sich zunächst zu Baumspringern entwickelt hätten. Die Vögel müssten deshalb zwingend von baumlebenden Diapsiden abstammen und könnten deshalb keine Dinosaurier sein.[4] Erst im Jahr 2000 wurde Microraptor entdeckt, eine sehr wahrscheinlich baumlebende und gleitfliegende Gattung gefiederter Theropoden. Wenngleich Microraptor in der Frühen Kreide (ca. 130 Mio. Jahre vor heute) und somit deutlich später lebte als die ersten Vögel, zeigt seine Entdeckung, dass die Maniraptora auch baumlebende Formen hervorbringen konnten, und stützt die These der Zugehörigkeit der Vögel zu dieser Gruppe (und damit zu den Dinosauriern). Aber auch die Debatte, ob sich die Vögel aus Bodenläufern oder aus Baumspringern entwickelt haben, ist noch nicht entschieden.
Vorläufer der modernen Vögel
Das bekannteste evolutionäre Bindeglied zwischen Reptilien und den Vögeln ist die Gattung Archaeopteryx (wörtlich: „Urflügel“). Die Flügel von Archaeopteryx haben große Ähnlichkeit mit den Flügeln moderner Vögel. Fossilien dieser Gattung wurden in den Solnhofener Plattenkalken aus der Zeit des Oberjura gefunden. Von Archaeopteryx nahm man lange eine vermittelnde Position zwischen den beiden Klassen an, denn er schien mosaikartig sowohl Merkmale von Reptilien als auch solche von Vögeln zu zeigen. Aus diesem Grund sind unvollständig und schlecht erhaltene Exemplare – wie etwa beim „Haarlemer Exemplar“ im Teylers Museum – lange Zeit nicht als Fossilien dieses Tieres erkannt worden. Mit der Entdeckung von immer mehr gefiederten Dinosauriern seit den 1990er Jahren hat sich aber gezeigt, dass sehr viele, früher für typische Vogelmerkmale gehaltene Eigenschaften des Archaeopteryx bei vielen anderen Theropodenspezies ebenfalls vorkamen. Vögel erscheinen daher mehr und mehr als ans Fliegen angepasste, ansonsten aber typische Dinosaurier. Ungeklärt ist nicht zuletzt deshalb, ob Archaeopteryx ein echter „Urvogel“ war, also ein direkter Vorfahre der modernen Vögel. Viele Forscher meinen, dass er einer blind endenden Entwicklungslinie angehörte.
Der oberjurassische Archaeopteryx besaß noch Kiefer mit Zähnen, eine lange Schwanzwirbelsäule und bewegliche bekrallte Mittelhandknochen. Wahrscheinlich besaß er, wie es auch für seine Nicht-Vogel-Dinosaurierverwandten vermutet wird, eine konstante, aktiv geregelte Körpertemperatur (Homoiothermie). Auch die in Sedimentgesteinen der oberen Kreide gefundenen fossilen Wasservögel (Ichthyornis und Verwandte) waren bezahnt. Die heutigen Vogelgruppen mit ihren unbezahnten Kiefern haben sich erst im Känozoikum herausgebildet. Der Verlust des harten Zahnschmelzes muss genetischen Analysen zufolge aber bereits vor mehr als 100 Millionen Jahren in der unteren Kreide in der Entwicklungslinie, die zu den modernen Vögeln (Neornithes, Vogel-Kronengruppe) führt, stattgefunden haben und zwar nach der Abspaltung der Linie, die zu Ichthyornis führt.[5] Beim jüngsten gemeinsamen Vorfahren der beiden Hauptlinien der modernen Vögel (Urkiefervögel und Neukiefervögel) sollen schließlich alle für die Zahnentwicklung zuständigen Gene abgeschaltet gewesen sein.[5] Während der älteste unzweifelhafte Fossilnachweis der Kronengruppen-Vögel aus der späten Oberkreide (Maastrichtium) stammt,[6][7] sind bereits aus der späten Unterkreide Vögel bekannt, deren Anatomie an die der modernen Vögel stark heranreicht. Ein Beispiel für solch einen Vertreter ist Gansus yumenensis aus der Xiagou-Formation (115–105 mya) der chinesischen Gansu-Provinz.[8] Generell besaßen die Vögel bereits in der Kreidezeit eine große Artenvielfalt. Eine mit Gansus relativ eng verwandte Gruppe kreidezeitlicher, anatomisch jedoch weniger fortschrittlicher Vögel sind die Enantiornithes, deren fossile Überreste unter anderem im Nordosten Chinas gefunden wurden.
Entwicklung im Känozoikum
Beim Massenaussterben vor 65,5 Millionen Jahren (Kreide-Tertiär-Grenze) starben die Nicht-Vogel-Dinosaurier aus. Auch die Vögel und viele andere Gruppen von Lebewesen erlitten einen erheblichen Verlust an Arten und höheren Taxa.
Zu Beginn des Känozoikums entwickelte sich in sehr kurzer Zeit aus den vermutlich wenigen überlebenden Arten eine Vielzahl neuer Vogelgruppen, welche die Grundlage der heutigen Avifauna (Vogelwelt) bildete. Einige dieser Gruppen starben wieder aus. Vogelfossilien aus dem Eozän (z. B. aus der Grube Messel) belegen das Vorhandensein einer vielfältigen Avifauna, wobei nicht alle Arten heute noch lebenden Gruppen zuzuordnen sind. Ein Beispiel für Vertreter ausgestorbener Gruppen sind große, fleischfressende, flugunfähige Vögel wie Gastornis, die im Eozän die ökologische Rolle der noch nicht entwickelten Raubtiere eingenommen haben könnten.
Die Evolution der Feder
1995 in der Volksrepublik China entdeckte fossile Vögel aus der Unterkreide ähnelten Archaeopteryx hinsichtlich Krallenhand, Bauchrippen (Gastralia) und Beckenbau. Doch zeigten manche Versteinerungen Federn und ein kräftiges Brustbein wie heutige Vögel, ferner einen Schnabel ohne Zähne und nur noch eine kurze Schwanzwirbelsäule. Die 1998 veröffentlichte Erstbeschreibung des kleinen gefiederten Dinosauriers Caudipteryx trug wesentlich zum Verständnis der Evolution der Vögel, der Gefiederentwicklung und zum Teil des Vogelflugs bei. Demnach entwickelten die Vogelvorfahren zunächst sowohl an den Vorder- als auch an den Hinterextremitäten Federn und konnten damit im Gleitflug von Baum zu Baum gelangen. Die Flügelbildungen an den Hinterextremitäten wurden im Laufe der Evolution reduziert, sodass lediglich die Arm- und Handschwingen zum Fliegen übrigblieben.
Nach einer anderen These bildeten sich die Federn zuerst zum Schutz vor Wärmeverlust bei bodenlebenden, zweibeinig gehenden Sauriern. Auch heutige Vögel haben Tausende von relativ einfach gebauten Flaumfedern, aber nur etwa 50 Schwungfedern. Eine Entwicklung von isolierendem Flaum zu komplexeren Flugfedern macht auch die Zwischenstufen der Entwicklung plausibler, die für das Fliegen noch ungeeignet waren.
Fortpflanzung

Allen Vögeln gemeinsam ist die Fortpflanzung durch die Ablage von Eiern (Oviparie). Es gibt zwar einige lebendgebärende Fische, Lurche und Reptilien (sowie andererseits eierlegende Säugetiere), aber keine einzige lebendgebärende Vogelart. Wahrscheinlich hätte eine längere Tragzeit und die damit verbundene Gewichtsveränderung für die Vögel als Flieger zu viele Nachteile. Zwar sind Fledertiere als Flieger ebenfalls lebendgebärend, aber in dieser Untergruppe der Höheren Säugetiere ist das Gebären lebender Jungen (Viviparie) ein ursprüngliches Merkmal. Das heißt, die Flugfähigkeit der Fledertiere entwickelte sich bei einer bereits lebendgebärenden Stammart. Für Vögel ist das Eierlegen hingegen ein ursprüngliches Merkmal, sodass bei ihnen die Flugfähigkeit die Entstehung von Viviparie verhindert haben könnte.
Die Männchen einiger Vogelgruppen wie beispielsweise Laufvögel und Gänsevögel haben gut entwickelte Kopulationsorgane, während andere gar keinen oder einen einfach gebauten Vogel-Penis besitzen. Bei weiblichen Vögeln ist im Allgemeinen nur ein – und zwar der linke – Eierstock entwickelt, während bei den Männchen zwei Gonaden vorhanden sind. Für das Flugvermögen ist ein möglichst geringes Körpergewicht von großer Bedeutung. So werden die Sexualorgane bei den Vögeln außerhalb der Paarungszeit stark zurückgebildet, sie schrumpfen aber meist nicht sofort nach der Eiablage, um bei Bedarf noch ein zweites Gelege erzeugen zu können. Bei der Begattung, dem sogenannten Tretakt, steigt das Männchen auf den Rücken des Weibchens, und beide drücken ihre Kloaken aufeinander. Die Samenflüssigkeit des Männchens fließt in die Kloake des Weibchens. Die Kopulation dauert meist nur wenige Sekunden, wird jedoch oft wiederholt.
Der Follikelsprung – das Freiwerden der Eizelle – kann durch verschiedenartige Reize (wie zum Beispiel Anblick eines Geschlechtspartners) ausgelöst werden. In der Öffnung des Eileiters verbleibt das Ei einige Minuten und wird von im Eileiter entlang gewanderten Spermien befruchtet. Danach befördern Muskelbewegungen des Eileiters das noch unfertige Ei in Richtung der Geschlechtsöffnung. Im Eileiter werden dem Dotter (bestehend aus einem Drittel Proteinen und zwei Dritteln Fetten und fettähnlichen Stoffen sowie Vitaminen und Mineralsalzen) und dem sich bereits furchenden Embryo erst das Eiklar (Proteine, Salze und Wasser), die Eihaut und schließlich die Eischale angelagert. Im Gegensatz zu den weichschaligen Eiern der Reptilien sind die Schalen von Vogeleiern stark verkalkt (bis zu 94 % Calciumcarbonat); sie lassen jedoch einen Gasaustausch zur Atmung zu. Durch die Muskelbewegungen des Eileiters (dessen letzten Teil man als Uterus bezeichnet) erhalten Vogeleier ihre arttypische Form. Bei weiblichen Vögeln wächst, induziert durch Östrogene, vor der Eiablage eine sehr calciumreiche Knochensubstanz in den Beinknochen zwischen dem harten Außenknochen und dem Knochenmark, in der das Calcium für die Bildung der Eierschalen gespeichert wird. Diese Knochensubstanz, die sich nach der Eiablage wieder zurückbildet, wird als medullärer Knochen bezeichnet.[9] Medullärer Knochen ist auch bei Nicht-Vogel-Dinosauriern und fossilen Vögeln bekannt und dient in der Paläontologie zur Bestimmung weiblicher Tiere.[10][11]
Vogeleier enthalten alle Nährstoffe, Vitamine und Spurenelemente, die der Embryo zu seiner Entwicklung braucht. Der zum Stoffwechsel unerlässliche Sauerstoff wird durch die feste Schale hindurch aufgenommen. Die Eier von Nestflüchtern enthalten mehr Dotter als Eier von gleich großen, als Nesthocker schlüpfenden Vogelarten. In dem einen Fall sind die Küken schon kurz nach dem Schlüpfen weitgehend selbständig, während Nesthocker völlig hilflos, unbefiedert und meist blind aus dem Ei kommen und von den Eltern lange gefüttert werden.
Viele Vögel erbrüten nur ein Ei, während die größten Gelege mit 20 bis 22 Eiern bei Hühnervögeln vorkommen. Bei einigen Arten legen auch zwei oder mehrere Weibchen ihre Eier in ein gemeinsames Nest (s. die großen Laufvögel). Viele Vogelarten ziehen in einer Fortpflanzungsperiode mehrere Jahresbruten hintereinander hoch.
Brut und Schlüpfen der Jungen
Einige Vogelarten (Großfußhühner) nutzen Fremdwärme zum Ausbrüten ihrer Eier. Die meisten Vögel jedoch wärmen ihre Eier im Brust- und Bauchgefieder. Bei einigen Arten haben die an der Brut beteiligten Geschlechter Brutflecke (federlose Hautpartien an Brust und Bauch), an denen die Körperwärme besser zu den Eiern gelangen kann als durch das isolierende Gefieder. Bei vielen Arten brüten beide Partner, bei anderen nur das Weibchen oder seltener ausschließlich das Männchen, beispielsweise die großen Laufvögel sowie der Kaiserpinguin. Die Bruttemperatur liegt bei etwa 34 °C. Die Eier werden während der Brut häufig gewendet, um so eine gleichmäßige Erwärmung zu gewährleisten. Viele andere Faktoren, wie genügend Feuchtigkeit, keine übermäßige Erwärmung durch Sonneneinstrahlung, keine oder nur wenige schwache Erschütterungen u. a. sind wichtig für eine erfolgreiche Brut. Die kürzeste Brutdauer beträgt elf Tage (z. B. Feldlerche 11–12), die längste etwa zwölf Wochen (Streifenkiwi bis 92 Tage).
Kurz vor dem Schlüpfen ist die Kalkschale durch Kalkabbau vom Embryo dünner geworden. Die Jungvögel geben oft schon vor Verlassen des Eies Rufe von sich, die oftmals der Synchronisation des Schlüpfvorganges dienen oder für die Beziehung zwischen Altvogel und Nestling von entscheidender Bedeutung sind. Der Jungvogel reibt und pickt von innen her die Eischale auf, bis ein kleines Loch entsteht. Dazu ist auf der Spitze des Oberschnabels – und bei einigen Vogelarten auf dem unteren Schnabelteil – ein kleiner, harter Höcker, der Eizahn ausgebildet, der wenige Tage nach dem Schlüpfen abfällt oder zurückgebildet wird. Der gesamte Schlüpfvorgang dauert – je nach Vogelart – einige Minuten oder bis zu vier Tagen (Röhrennasen).
Vögel, die alt genug sind, um flugfähig zu sein, sind flügge.
Nestlinge: Nesthocker, Nestflüchter und Platzhocker
Nesthocker
Zum Zeitpunkt des Schlupfes sind die Jungvögel meist nackt und blind. Die Augen und Gehörgänge sind noch verschlossen. Sie sind auf die Wärmezufuhr (hudern) von außen angewiesen und auf eine regelmäßige Fütterung durch die Altvögel. Je nach Vogelart hudern beide Elternteile oder nur das Weibchen. Viele Nesthocker verlassen das Nest erst kurz vor dem Erreichen der Flugfähigkeit. An diesem Punkt heißt der Jungvogel nicht mehr Nestling, sondern Ästling. Die Ästlinge werden weiter bis zum Erreichen der Flugfähigkeit und dann weiter bis zur Selbständigkeit von den Altvögeln gefüttert. Vertreter der Gruppe der Nesthocker sind beispielsweise Störche, Tauben, Greifvögel, Segler, Spechte und alle Singvogelarten.
Nestflüchter

Diese Jungvögel sind gleich beim Schlupf am ganzen Körper mit Daunen befiedert. Bereits voll entwickelt sind Augen und Ohren. Die meisten Nestflüchterarten können ab Schlupf selbständig Nahrung aufnehmen. Ebenso können sie je nach Art sofort laufen, schwimmen und tauchen. Bereits am 1. oder 2. Lebenstag verlassen sie unter der Führung der Altvögel das Nest. Dieses wird nie wieder aufgesucht. In der Anfangszeit werden sie in der Regel durch die Mutter gehudert. Durch leise Rufe teilen die Altvögel ihren Jungen mit, wo sie sich befinden. Ebenso kommunizieren die Jungvögel mit den Eltern und ihren Geschwistern. Bis zur Selbständigkeit der Nestflüchter verbringt die Familie die Zeit zusammen. Zu den Nestflüchtern gehören beispielsweise Entenvögel und Kraniche.
Platzhocker
Im Unterschied zu Nesthockern sind Platzhocker beim Schlupf bereits voll befiedert, ihre Augen und Ohren sind voll entwickelt. Sie halten sich im Nest und in dessen unmittelbarer Nähe auf bis zum Erreichen der Flugfähigkeit. Lange Zeit sind sie nicht in der Lage, ihre Körpertemperatur selber zu regeln, weswegen sie lange gehudert werden. Ebenso erreichen sie die Flugfähigkeit erst sehr spät. Wie beim Nesthocker füttern die Altvögel – manchmal nur das Weibchen, seltener nur das Männchen – die Jungvögel, bis jene selbständig sind. Zu den Platzhockern gehören beispielsweise fast alle Möwen und Pinguine.
Flugunfähige Vögel
Die Flugfähigkeit ist bei einigen Vogelarten bzw. -gruppen sekundär verlorengegangen, das heißt, sie sind flugunfähig. Das kann mehrere Gründe haben:
- Anpassung an das Leben im Wasser, wie dies bei den auf der Südhalbkugel lebenden Pinguinen der Fall ist. Auch die auf der Nordhalbkugel lebenden Alkenvögel zeigen eine Tendenz zur Flugunfähigkeit, der ausgestorbene Riesenalk war flugunfähig. Beide Gruppen „fliegen“ unter Wasser mit ihren Flügeln.
- Die Verdauung von energiearmer Nahrung erfordert ein großes und damit schweres Verdauungssystem. Grasfressende Vögel wie Gänse sind daher besonders schwer. Aufgrund des Flugvermögens können Vögel nicht beliebig an Größe zunehmen. Daher gibt es unter entsprechenden Nahrungsspezialisten ebenfalls sekundär flugunfähige Arten wie beispielsweise den Kakapo.
- Auch die Umstellung auf schnelles Laufen wie beim afrikanischen Strauß und den anderen großen Laufvögeln in Südamerika und Australien kann zu Flugunfähigkeit führen.
- Als letztes kann auch das Nichtvorhandensein von bodenbewohnenden Raubsäugern eine Flugfähigkeit überflüssig machen. Deshalb haben viele Bewohner isolierter Inseln ihre Flugfähigkeit verloren. Vielen Arten wurde dies aber zum Verhängnis, nachdem durch Seefahrer doch Raubsäuger (z. B. Katzen, Ratten, Schweine usw.) eingeführt wurden. Beispiele dafür sind die inzwischen ausgestorbenen Dronten (Raphus cucullatus) auf Mauritius, der neukaledonische Kagu (Rhynochetos jubatus), die neuseeländische Takahe (Porphyrio mantelli) und die ebenfalls flugunfähige Galapagosscharbe (Nannopterum harrisi) sowie die stark gefährdete Südinseltakahe und alle Arten der Kiwis.
Sinnesleistungen der Vögel
Die Sinnesleistungen der Vögel unterscheiden sich nicht grundlegend von denen der Säugetiere. Allerdings gibt es aufgrund der anderen Lebensweise Unterschiede in der Konstruktion und der Gewichtung der einzelnen Sinne, die es oft schwer machen, sich vorzustellen, wie Vögel ihre Umwelt wahrnehmen.
_I_IMG_7881.jpg.webp)
Sehen
Die Augen der Vögel sind relativ zur Körpergröße größer als die der Säugetiere. Die meisten Vogelarten können mehr Bilder pro Sekunde unterscheiden als Menschen (siehe Flimmerfusionsfrequenz). In der Vogelhaltung werden daher keine Leuchtstofflampen mit 50 Hertz betrieben, da dieses Licht für Vögel flimmert.
Je nach Ökologie besitzen Vogelaugen zahlreiche Spezialanpassungen. Zum Beispiel können Vögel, die ihre Nahrung unter Wasser suchen, ihre besonders weichen Augenlinsen durch Muskeln verformen und so das Brechungsvermögen anpassen.
Lichtempfindlichkeit und Sehschärfe
Eulen können bei Nacht zwar mehr sehen als Tagvögel oder der Mensch, ihr Sehen ist allerdings weniger auf Sehschärfe als auf Lichtausbeute ausgerichtet. Dies funktioniert aufgrund gleicher Physik wie bei lichtstarken Objektiven, die ebenfalls mit wenig Licht auskommen, sich dabei aber Schärfeprobleme, besonders mit der Schärfentiefe einhandeln. Wanderfalken sind dagegen auf Tagjagd optimiert; sie können kleine Objekte wie Beutevögel über Entfernungen von über einem Kilometer ausmachen und verfolgen.
Zumindest kleine Vögel sind in der Lage, UV-Licht zu sehen, bei großen Vögeln wird das UV-Licht vom Glaskörper des Auges zu stark ausgefiltert. Viele Arten besitzen nicht nur drei Farbrezeptoren wie der Mensch, sondern vier Farbrezeptoren (beim Menschen haben sich nach neueren Erkenntnissen der Rezeptor für rotes und der für grünes Licht aus einem gemeinsamen entwickelt, so dass Vögel eigentlich zwei Rezeptoren mehr als Säugetiere haben), davon einer für violettes Licht, der sich bei einigen Vogelgruppen zu einem UV-Rezeptor (um 375 Nanometer) entwickelt hat. Evolutionär ist hierzu nur die Mutation von drei Aminosäuren des Opsins notwendig.
Der Nutzen des UV-Lichtes ist sehr unterschiedlich:
- Mäuse-Urin leuchtet im UV-Bereich, mäusejagende Greife können so von oben eine Landschaft auf ihren Mäusereichtum beurteilen.
- Bei Früchten kann der Reifegrad mit Hilfe von UV-Licht besser beurteilt werden; manche Schimmelpilze besitzen im UV-Bereich andere Farben und fallen so besser auf.
- Es gibt einige Vogelarten, bei denen sich die Geschlechter im für uns sichtbaren Licht nicht unterscheiden, wohl aber im UV-Licht. Stare oder einzelne Meisenarten (Blaumeisen) sind dafür ein Beispiel.
Räumliches Sehen
Je nach ökologischer Anpassung ist die Fähigkeit zum räumlichen Sehen von Art zu Art sehr unterschiedlich. Bei Artengruppen, die unter einem hohen Verfolgungsdruck durch Raubtiere stehen (z. B. Tauben und Hühnervögel), sind die Augen seitlich am Kopf angeordnet. Dies erlaubt einen fast vollständigen Rundblick um 360°, die Überlappung der Sichtfelder und damit die Fähigkeit zum räumlichen Sehen ist aber relativ gering. Das andere Extrem stellen Eulen dar. Bei ihnen sind die Augen nebeneinander an der Vorderseite des Kopfes angeordnet (also wie bei Menschen). Die Sichtfelder der Augen überlappen sehr stark, entsprechend gut ist daher auch das räumliche Sehvermögen. Die geringe seitliche Ausdehnung des Sichtfeldes wird durch eine sehr starke Beweglichkeit der Halswirbelsäule ausgeglichen. Eulen können ihren Kopf um bis zu 270° drehen.
Bewegungssehen
Viele Vögel wippen beim Gehen mit dem Kopf vor und zurück. Dabei dient die Rückwärtsbewegung dazu, den Kopf für einen Moment relativ zur Umgebung in Ruhe zu halten, sodass das Bild auf der Netzhaut sich nicht bewegt, zugunsten der besseren Erkennbarkeit bewegter Objekte – wie beispielsweise eines Raubtieres. Der Scharfsehbereich ist beim Menschen ca. 2,5°, bei den Vögeln ca. 20°. Auch das verbessert die Wahrnehmung von Bewegungen.
Magnetsinn
Bei einigen Arten, besonders bei Zugvögeln, aber auch bei Haustauben, wurde ein Sinn für das Erdmagnetfeld nachgewiesen. Dieser Magnetsinn besteht aus zwei unterschiedlichen, einander ergänzenden Mechanismen und ist zum einen im Auge, zum anderen im Oberschnabel lokalisiert.[12]
Der Magnetsinn im Auge funktioniert mit Hilfe der sogenannten Radikalpaarbildung. Hierbei lässt das ins Auge fallende Licht bestimmte Moleküle zu Radikalen zerfallen. Diese Reaktion könnte durch das Erdmagnetfeld beeinflusst werden. Der Magnetsinn im Schnabel funktioniert durch eingelagerte magnetische Teilchen, die sich nach dem Magnetfeld der Erde ausrichten und so einen Reiz auf das umliegende Nervengewebe ausüben. Im Unterschied zum technischen Kompass richtet sich der Magnetsinn im Auge der Vögel nicht nach der Polung des Erdmagnetfeldes, sondern basiert auf dem Erkennen des Neigungswinkels der Feldlinien des Magnetfeldes. Experimentell nachgewiesen wurde der Magnetsinn erstmals 1967 am Zoologischen Institut in Frankfurt am Main durch Wolfgang Wiltschko bei Rotkehlchen.
Hören
Vögel besitzen keine Ohrmuschel, die äußere Gehöröffnung ist von einem Kranz kleiner Federn umgeben. Zur Schallortung müssen Vögel daher intensive Kopfbewegungen ausführen. Das Mittelohr besitzt nur ein Gehörknöchelchen, die Columella, die dem Steigbügel der Säugetiere entspricht. Die Schnecke ist relativ kurz und nur leicht gewunden und wird als Papilla basilaris bezeichnet. Der Hörsinn ist bei Vögeln dennoch relativ gut entwickelt und hat beispielsweise bei Eulen große Bedeutung für die Jagd. Der untere wahrnehmbare Frequenzbereich ist ähnlich wie bei den Säugetieren, hohe Töne ab etwa 6.000 Hertz werden von den meisten Vögeln dagegen nicht wahrgenommen. Das zeitliche Auflösungsvermögen für Töne liegt über dem des Menschen. Ornithologen müssen sich bei vielen Lautäußerungen von Vögeln damit behelfen, dass sie sie aufnehmen und verlangsamt abspielen, um die Details hören zu können.
Gleichgewichtssinn
Vögel besitzen mehrere unabhängige Gleichgewichtsorgane. Neben einem Gleichgewichtsorgan im Ohr sitzt ein zweites Organ im Becken, das zum Beispiel beim Sitzen auf Ästen die Körperlage analysiert. Vögel, bei denen dieses Organ zerstört ist, können ohne Gesichtssinn auf Störungen wie zum Beispiel das Drehen des Sitzastes nicht mehr richtig reagieren.
Geruchssinn
Sehr lange Zeit ging man von der Vorstellung aus, dass Vögel nur einen gering entwickelten Geruchssinn besitzen (sog. Mikrosmatiker). Als Ausnahme galt der neuseeländische Kiwi, der seine Nasenöffnung an der Schnabelspitze hat und sich vorwiegend nach dem Geruch orientiert. Aber auch die Gruppe der amerikanischen Neuweltgeier besitzt nachweislich Geruchsvermögen. Das wird unter anderem auf die fehlende Nasenscheidewand zurückgeführt und ist in Freilandbeobachtungen und -versuchen bewiesen worden.
Geschmackssinn
Die Geschmacksknospen der Vögel liegen nicht wie beim Säuger auf der Zunge, sondern im Bereich des Zungengrunds und im Rachen. Die Anzahl der Geschmacksknospen ist deutlich geringer als bei Säugetieren (Ente etwa 200, Mensch 9000), dennoch ist der Geschmackssinn bei Vögeln nicht so untergeordnet wie man lange angenommen hat. Je nach Art spielt der Geschmack gegenüber dem Tastsinn des Schnabels und der Zunge jedoch bei der Nahrungsauswahl eine untergeordnete Rolle.
Tastsinn
Der Tastsinn ist für viele Vögel bei der Nahrungssuche und Nahrungsaufnahme von großer Bedeutung. Schnabel und Zunge der meisten Vögel sind sehr tastempfindlich. Insbesondere Watvögel suchen ihre Nahrung, indem sie mit meist langen Schnäbeln im Schlamm stochern. Vögel, die auf Bäumen herumklettern, orientieren sich auch tastend mit ihren Zehen.
Vogelintelligenz
Forschungsergebnisse seit den 1990er Jahren erbrachten, dass Vögel, zumindest aber die Vertreter bestimmter Großgruppen, intelligenter sind, als es bis dahin allgemein angenommen wurde.
Die traditionelle Nomenklatur der Anatomie des Vogelhirns beruhte auf der Annahme, dass es sich bei den Strukturen des Vogelhirns um Äquivalente des Gehirns der Säugetiere handelt. Da sich diese Annahme als falsch herausgestellt hat, wurde mittlerweile eine neue Nomenklatur für das Vogelgehirn ausgearbeitet[13] (siehe auch Gehirn und Kognition der Vögel).
Hirnorganisch scheint die Vogelintelligenz mit einer im Verhältnis zu Hirnvolumen und -masse sehr hohen Anzahl und Dichte von Neuronen in Zusammenhang zu stehen. So weisen die Gehirne von Sperlings- und Papageienvögeln, die als die intelligentesten gelten, bei gleicher Masse deutlich mehr Neuronen auf als Säugergehirne (einschließlich denen von Primaten). Zudem ist die Hirnmasse und die Neuronenanzahl bei diesen Vögeln im Verhältnis zu ihrer Körpermasse größer als bei Säugetieren (außer bei Primaten). So hat das Wintergoldhähnchen ein Neuntel des Körpergewichts einer Maus, aber sein Gehirn enthält mehr als doppelt so viele Neuronen. Hierbei konzentrieren sich die Neuronen im Großhirn und insbesondere in der Großhirnrinde, die bei Sperlingsvogel- und Papageiengehirnen eine drei- bis vierfach höhere Neuronendichte aufweist als bei Primaten. Das dennoch vergleichsweise geringe Gewicht und Volumen dieser Vogelgehirne resultiert aus einer Verkleinerung der einzelnen Neuronen, wohingegen die Großhirnrinde als größter Teil des Säugergehirns aus relativ wenigen aber relativ großen Nervenzellen aufgebaut ist. Deutliche Unterschiede im Enzephalisationsgrad bestehen aber auch innerhalb der Vögel. Demnach hat ein Bankivahuhn, als Vertreter einer „primitiveren“ Linie der Vögel, die gleiche Anzahl von Neuronen wie eine Kohlmeise, obwohl es ein rund 50-fach höheres Körpergewicht aufweist. Allerdings ist auch bei Hühner- und Straußenvögeln die Neuronendichte in der Großhirnrinde immer noch ungefähr genauso groß wie bei Primaten. Evolutionär könnte der hohe Enzephalisationsgrad der Sperlingsvögel (insbesondere der großen Rabenvögel) und Papageien unter anderem durch die intensive Brutpflege, die in diesen Gruppen betrieben wird, begünstigt worden sein, durch die den Jungvögeln ausreichend Zeit bleibt, um viele Hirnzellen auszubilden.[14]
Viele Vögel können sich im Spiegel nicht erkennen, weshalb sie als Spiegelfechter ihr eigenes Spiegelbild bekämpfen.[15]
Vögel und Menschen
Etymologie
Das gemeingermanische Substantiv mittelhochdeutsch vogel, althochdeutsch fogal hat keine außergermanischen Entsprechungen; seine Herkunft ist nicht sicher geklärt.[16]
Mythologie, Redewendungen, Motiv in der Kunst
Seit der Menschwerdung hat der Homo sapiens auch die Vögel aufmerksam beobachtet, um sich von ihnen orientieren oder warnen zu lassen, sie zu jagen oder einzufangen. Der afrikanische Honiganzeiger beispielsweise führt Menschen zu den Nestern von Wildbienen, um die Larven zu fressen, während der Mensch vom Honig profitiert.
In vielen Kulturkreisen spielen Vogelarten daher in der Mythologie eine Rolle, vom Adler bis zum Sperling. Zu den mythischen Vögeln zählen etwa Phönix, Greif, Ziz (Jüdisch), Roch (Arabisch), Feng (Chinesisch) oder Garuda (Indisch). Daneben seien die Harpyien der griechischen Sagenwelt genannt sowie Wotans Begleiter, die Raben Hugin und Munin. Die Heraldik ist reich an stilisierten mythischen Vögeln. Hier tritt insbesondere der Adler in Erscheinung, der das Wappentier vieler Staaten ist (Deutschland, Österreich, Polen, USA u. a.). In der christlichen Ikonographie spielt der Pelikan eine wichtige Rolle.
Vögel erscheinen in vielen Redensarten: Jemand „stinkt wie ein Wiedehopf“ oder ist „zänkisch wie eine Meise“ oder „ist einfach ein Gimpel“, wenn er „jedem auf den Leim geht“ (gemeint hier der Vogelleim). Gestisch kann man ihm „einen Vogel zeigen“.
Auch in den Künsten tauchen zahlreiche Vogelarten auf. Ein Beispiel ist Selma Lagerlöfs Roman Die wunderbare Reise des kleinen Nils Holgersson mit den Wildgänsen.
Vögel als Nutztiere und Haustiere
Der Mensch hält zahlreiche Vogelarten als Nutz- oder Haustiere. Der Verzehr von Geflügel erfüllt eine wichtige Rolle bei der menschlichen Ernährung, denn das Fleisch ist fettarm und liefert hochwertiges Protein. Die Nutzung einheimischer Vogelarten als Haustier nimmt in Deutschland seit dem 19. Jahrhundert jedoch stark ab. Waren es in den 1980er Jahren noch sieben Millionen Ziervögel (ohne Tauben), sind es im Jahre 2018 4,8 Millionen. Diese Zahlen erfassen jedoch nur die durch den Zoofachhandel verkauften Tiere. Eine nicht unerhebliche Anzahl dürften durch Hobby-Vogelzüchter zusätzlich gezüchtet/vermehrt werden.[17]
Zier- oder Heimvogel: Es gibt Heimvögel (Papageien), die keiner Haustierart angehören, sondern einer Wildtierart und dennoch in Haushalten gepflegt werden. Ziervögel werden hingegen vornehmlich artgerecht in Volieren, Vogelhäusern oder Zuchtanlagen im Garten gehalten. In der heutigen Zeit wird der Begriff Ziervogel jedoch für alle vom Menschen gehaltenen Vögel (außer Nutzgeflügel) verwendet.[18]
Einige Arten, wenn bereits als Jungvögel und im Käfig geduldig dazu angehalten, lernen es auch, Worte und kurze Sätze nachzusprechen, so besonders – nach absteigender Gelehrigkeit geordnet – Papageien und Stare sowie verschiedene Arten der Rabenvögel. Vielfach lassen sie sich abrichten, z. B. Körner oder Obst von den Lippen aufzunehmen bis hin zur Dressur als Nutztier, beispielsweise als Jagdfalke oder Brieftaube. Bis in die 1950er Jahre waren Kanarienvögel in Deutschland wichtige Nutztiere im Kohlebergbau, wo ihr Verhalten vor dem Auftreten giftiger Gase (Kohlenmonoxid) warnte, die beim Stollenvortrieb austreten können.
Das direkte Zusammenleben mit Vögeln, insbesondere durch intensiven Kontakt auf engstem Raum, birgt für den Menschen jedoch mitunter Gefahren in Form übertragbarer Krankheiten (Zoonosen). Unter nichthygienischen Bedingungen ist in Einzelfällen in Asien beispielsweise der Erreger der Geflügelpest bzw. der Vogelgrippe H5N1 und der Vogelgrippe H7N9 auf den Menschen übertragen worden, der Krankheitsverlauf endet meist tödlich.
Nichtsdestotrotz wird das Hobby der Vogelhaltung und Vogelzucht auch in der Neuzeit ausgeübt.
Vogelschutz
Viele der ausgestorbenen Arten gehörten zu Inselpopulationen; sie wurden durch den Menschen oder von ihm eingeführte andere Tierarten verdrängt, ein Prozess, der schon vor Beginn der Industrialisierung einsetzte und sich nun verstärkt hat. Gegenwärtig gelten über 10 Prozent der etwa 10.451[1] rezenten Vogelarten als gefährdet. Sie werden in sogenannten roten Listen aufgeführt. Neue Berechnungen ergaben sogar, dass bis 2100 etwa zwölf Prozent der Arten aussterben könnten.[19] Laut dem European Bird Census Council ist in Europa der Bestand bei den 146 gemessenen Arten zwischen 1960 und 2016 um 14 % zurückgegangen. Betrachtet man nur die Landwirtschaftliche Nutzfläche, belief sich der Rückgang bei den 39 gemessenen Arten gar auf 56 Prozent.[20] In Nordamerika ist die Vogelwelt zwischen 1970 und 2018 um 29 % zurückgegangen.[21][22]
„Wenn wir die ökologische Landwirtschaft nicht deutlich ausdehnen, können wir nicht sicher sagen, ob in zehn oder 20 Jahren von der jetzigen Vogelpopulation noch etwas übrig ist.“
Weltweit arbeitet die Vogelschutz-Organisation BirdLife International, mit ihren jeweils nationalen Partnerorganisationen für den Schutz der Vögel und ihrer Lebensräume. In Deutschland ist der Naturschutzbund Deutschland (NABU, der Landesbund für Vogelschutz LBV in Bayern ist angegliedert) der nationale Partner von BirdLife. In der Schweiz arbeiten der Schweizer Vogelschutz SVS, ASPO, ASPU und in Österreich Birdlife Österreich für den Erhalt der Artenvielfalt als BirdLife-Partner.
Siehe auch: Das gegenwärtige Massenaussterben
Rechtliches zur Haltung von Vögeln in Menschenobhut
In Deutschland sind gemäß dem Tierschutzgesetz die artgemäßen Bedürfnisse eines Vogels in der Haltung zu erfüllen. Die Haltung besonders und streng geschützter Arten, sowie die Haltung von Kreuzungen nicht geschützter Arten ist verboten. Greifvogelhybriden dürfen nur innerhalb besonders gesicherter Volieren gehalten werden. Für die Unterbringung von Vögeln, Kleinvögeln, Greifvögel, Laufvögel und Papageien gibt es jeweilige Mindestanforderungen, herausgegeben vom Bundesministerium für Ernährung und Landwirtschaft.[24]
In Österreich ist seit 1. Januar 2005 nach dem Bundestierschutzgesetz die Haltung von Vögeln in Menschenobhut strenger reglementiert als bisher. Insbesondere für nicht domestizierte Vögel wie Greifvögel, Papageien, und viele andere, ist nur mehr die artgerechte Haltung erlaubt, vielen arttypischen Bedürfnissen muss nun nachgekommen werden.[25][26] Unter anderem ist die Einzelhaltung sozialer Vögel und das Einschränken der Flugfähigkeit (Flügelstutzen) – bis auf wenige Ausnahmen – generell verboten, Mindestgrößen für Käfige, Volieren und Schutzräume sind für die verschiedenen Arten festgelegt, wobei im Wesentlichen für domestizierte Vögel wie Hühner oder Wellensittiche kleinere Gehege zulässig sind. Handaufzuchten sind nur mehr in Ausnahmefällen, und dann nur mit Nestlingen (zur Vermeidung von Fehlprägungen) zulässig.
In der Schweiz ist für die Vogelhaltung in Menschenobhut eine Bewilligung von kantonaler Ebene notwendig. Geschützte Vogel dürfen nur zu dem Zweck gehalten werden, wenn die Haltung der Wiederauswilderung dient. Bei Vogelgrippegefahr ist die Haltung von Vögeln im Freien verboten. Bei der Haltung von nicht gefährdeten Vögeln müssen die tatsächlichen Bedürfnisse der Tiere erfüllt werden.
Siehe auch
Literatur
- Der grosse Kosmos-Vogelatlas über 450 Arten in Wort, Bild und Ton,CD-ROM, United Soft Media Verlag, München 2001, Version 3.0, ISBN 3-8032-1742-3.
- William Geoffrey Arnott: Birds in the ancient world from A to Z. Routledge, London, New York 2007. – Rez. von Roberto Batisti in: Eikasmós 19, 2008, 517–519, (online).
- Einhard Bezzel, Roland Prinzinger: Ornithologie. Verlag Eugen Ulmer, Stuttgart 1990, ISBN 3-8252-8051-9.
- Colin Harrison: Jungvögel, Eier und Nester der Vögel Europas, Nordafrikas und des Mittleren Ostens. AULA Verlag, Wiebelsheim 2004, ISBN 3-89104-685-5.
- Roger Lederer und Carol Burr: Latein für Vogelbeobachter: über 3000 ornithologische Begriffe erklärt und erforscht, aus dem Englischen übersetzt von Susanne Kuhlmannn-Krieg, Verlag DuMont, Köln 2014, ISBN 978-3-8321-9491-8.
- Christoph Moning, Thomas Griesohn-Pflieger, Michael Horn: Grundkurs Vogelbestimmung. Eine Einführung zur Beobachtung und Bestimmung unserer heimischen Vögel. Yweite Auflage. Quelle & Meyer Verlag, Wiebelsheim 2013, ISBN 978-3-494-01535-4.
- Detlef Singer: Die Vögel Mitteleuropas. Kosmos Naturführer. Franckh Verlag, Stuttgart 1988, ISBN 3-440-05906-5.
- Hugo Suolahti: Die deutschen Vogelnamen. Eine wortgeschichtliche Untersuchung. Straßburg 1909.
- Lars Svensson: Der neue Kosmos Vogelführer: Alle Arten Europas, Nordafrikas und Vorderasiens. Franckh-Kosmos Verlag, Stuttgart 1999, ISBN 978-3-440-07720-7.
Weblinks
- Literatur von und über Vögel im Katalog der Deutschen Nationalbibliothek
- Avibase – Die Welt-Vogel-Datenbank (etwa 10.000 Arten)
- Handbook of the Birds of the World and BirdLife International digital checklist of the birds of the world: Version 9.1 (June 2017)
- Jungvogel gefunden – was tun? (PDF; 155 kB)
- Merkmale der Vögel – Aves (Memento vom 20. Juni 2008 im Internet Archive)
- Schweizerische Vogelwarte, alles rund um Vögel
- https://ebird.org/
- Vogelstimmen
Einzelnachweise
- Stand 22. Juli 2019 (v9.2), F. Gill, D. Donsker (Hrsg.): IOC World Bird Names
- George F. Barrowclough, Joel Cracraft, John Klicka, Robert M. Zink. How Many Kinds of Birds Are There and Why Does It Matter? PLOS ONE, 2016; 11 (11): e0166307 DOI: 10.1371/journal.pone.0166307
- Wilfried Westheide, Reinhard Rieger (Hrsg.): Spezielle Zoologie. Teil 2: Wirbel- oder Schädeltiere. Spektrum Akademischer Verlag, Heidelberg/Berlin 2004, ISBN 3-8274-0307-3, S. 403
- Darren Naish: The “Birds Are Not Dinosaurs” Movement. Tetrapod Zoology. blogs.scientificamerican.com, 15. November 2017
- Robert W. Meredith, Guojie Zhang, M. Thomas P. Gilbert, Erich D. Jarvis, Mark S. Springer: Evidence for a single loss of mineralized teeth in the common avian ancestor. In: Science. Bd. 346, Nr. 6215, 2014, doi:10.1126/science.1254390 (alternativer Volltextzugriff: Erich Jarvis Lab, inkl. Supplementum)
- J. A. Clarke, C. P. Tambussi, J. I. Noriega, G. M. Erickson, R. A. Ketcham: Definitive fossil evidence for the extent avian radiation in the Cretaceous. In: Nature Bd. 433, 2005, S. 305–308, doi:10.1038/nature03150 (alternativer Volltextzugriff: ResearchGate)
- Neil Brocklehurst, Paul Upchurch, Philip D. Mannion, Jingmai O’Connor: The Completeness of the Fossil Record of Mesozoic Birds: Implications for Early Avian Evolution. In: PLoS ONE. Bd. 7, Nr. 6, 2012, e39056 doi:10.1371/journal.pone.0039056
- Hai-lu You, Matthew C. Lamanna, Jerald D. Harris, Luis M. Chiappe, Jingmai O’Connor, Shu-an Ji, Jun-chang Lü, Chong-xi Yuan, Da-qing Li, Xing Zhang, Kenneth J. Lacovara, Peter Dodson, Qiang Ji: A Nearly Modern Amphibious Bird from the Early Cretaceous of Northwestern China. In: Science. Bd. 312, Nr. 5780, 2006, S. 1640–1643, doi:10.1126/science.1126377
- Physiologische Besonderheiten bei Vögeln und Reptilien. Powerpoint-Präsentation zur gleichnamigen Lehrveranstaltung, Universität Leipzig, 2010
- Mary H. Schweitzer, Jennifer L. Wittmeyer, John R. Horner: Gender-Specific Reproductive Tissue in Ratites and Tyrannosaurus rex. In: Science. Bd. 308, Nr. 5727, 2005, S. 1456–1460, doi:10.1126/science.1112158.
- Anusuya Chinsamy, Luis M. Chiappe, Jesús Marugán-Lobón, Gao Chunling, Zhang Fengjiao: Gender identification of the Mesozoic bird Confuciusornis sanctus. In: Nature Communications. Bd. 4, Artikel-Nr. 1381, 2012, doi:10.1038/ncomms2377
- Henrik Mouritsen: Long-distance navigation and magnetoreception in migratory animals. In: Nature. Band 558, Nr. 7708, Juni 2018, ISSN 1476-4687, S. 50–59, doi:10.1038/s41586-018-0176-1 (nature.com [abgerufen am 17. April 2021]).
- Anton Reiner, David J. Perkel, Claudio V. Mello, Erich D. Jarvis: Songbirds and the Revised Avian Brain Nomenclature. In: H. Philip Zeigler, Peter Marler (Hrsg.): Behavioral Neurobiology of Birdsong. Annals of the New York Academy of Sciences. Bd. 1016, 2004, S. 77–108, doi:10.1196/annals.1298.013, PMC 2481519 (freier Volltext)
- Gesamter Absatz nach: Seweryn Olkowicz, Martin Kocourek, Radek K. Lučan, Michal Porteš, W. Tecumseh Fitch, Suzana Herculano-Houzel, Pavel Němec: Birds have primate-like numbers of neurons in the forebrain. Proceedings of the National Academy of Sciences. Bd. 113, Nr. 26, 2016, S. 7255–7260, doi:10.1073/pnas.1517131113
- Joël Roerig: Shadow boxing by birds – a literature study and new data from southern Africa. In: Ornithological Observations, ISSN 2219-0341. Band 4, 4. Juni 2013, S. 39–68.
- Das Herkunftswörterbuch (= Der Duden in zwölf Bänden. Band 7). Nachdruck der 2. Auflage. Dudenverlag, Mannheim 1997 (S. 792). Siehe auch DWDS („Vogel“) und Friedrich Kluge: Etymologisches Wörterbuch der deutschen Sprache. 7. Auflage. Trübner, Straßburg 1910 (S. 477).
- Zentralverband Zoologischer Fachbetriebe Deutschlands e. V.: Der Deutsche Heimtiermarkt 2018. Hrsg.: IVH und ZZF. Wiesbaden Januar 2018.
- Bielfeld, Horst, fl. 1960-2008.: Ziervögel 120 Arten und ihre Haltung. 2., überarb. Auflage. Ulmer, Stuttgart (Hohenheim) 1998, ISBN 3-8001-7437-5, S. 9.
- Hochrechnung: Ein Viertel aller Vogelarten bedroht. Spiegel Online, 4. Juli 2006
- European wild bird indicators and Trends of common birds in Europe, 2018 update. In: ebcc.info. 21. Januar 2019, abgerufen am 31. Januar 2019.
- Kenneth V. Rosenberg et al.: Decline of the North American avifauna. In: Science. September 2019, doi:10.1126/science.aaw1313.
- Daniel Lingenhöhl: Artensterben: 3 Milliarden Vögel verschwunden. In: spektrum.de. 20. September 2019, abgerufen am 25. September 2019.
- Südkurier: Ornithologe Peter Berthold: "Unseren Vögeln geht's beschissen"
- BMEL: Mindestanforderungen an die Haltung von Kleinvögeln. Abgerufen im Januar 2019.
- 2. Tierhaltungsverordnung auf der Homepage des österreichischen Bundeskanzleramts
- Anlage 2: Mindestanforderungen für die Haltung von Vögeln der 2. Tierhaltungsverordnung, Homepage des österreichischen Bundeskanzleramts