Reptilien
Die Reptilien oder Kriechtiere (Taxon: Reptilia, lat. reptilis „kriechend“) sind eine unterschiedlich definierte Gruppe von Tetrapoden, die – je nach Systematik (Klasse oder Klade) – unterschiedliche Gruppen der Amnioten umfasst.[1]
| Reptilien | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Veraltete systematische Gruppe Das hier behandelte Taxon ist nicht Teil der in der deutschsprachigen Wikipedia dargestellten Systematik. Näheres hierzu findet sich im Artikeltext. | ||||||||||||||

Vertreter von vier rezenten Reptiliengroßgruppen: | ||||||||||||||
| Systematik | ||||||||||||||
| ||||||||||||||
| Wissenschaftlicher Name | ||||||||||||||
| Reptilia | ||||||||||||||
| Laurenti, 1768 | ||||||||||||||
Nach traditioneller Auffassung sind die Reptilien (Reptilia) eine Klasse der Wirbeltiere am Übergang von den „niederen“ (Anamnia) zu den „höheren“ Wirbeltieren (Säugetiere und Vögel). Nach moderner Auffassung sind sie als solche jedoch keine natürliche Gruppe, sondern ein paraphyletisches Taxon, weil sie nicht alle Nachkommen ihres letzten gemeinsamen Vorfahren enthalten. Das klassische Taxon Reptilia gilt daher als veraltet und wird in der zoologischen und paläontologischen Systematik kaum noch verwendet. Der Taxonname dient heute meist als informeller Sammelbegriff für Landwirbeltiere mit ähnlicher Morphologie und Physiologie (siehe Merkmale). In diesem Sinne werden aktuell 11.690[2] rezente Reptilienarten unterschieden.
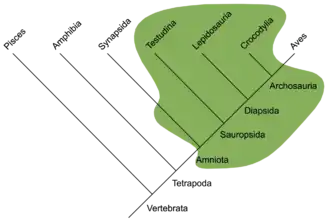
Aus kladistischer Sicht, die heute wissenschaftlicher Standard ist, müssten die Reptilien als monophyletisches Taxon, also als natürliche (vollständige) Abstammungsgruppe (Reptilia als Klade), mindestens auch die Vögel enthalten, unter Berücksichtigung bestimmter ausgestorbener Formen („säugetierähnliche Reptilien“ wie Dinocephalia) sogar auch die Säugetiere. Um diese Verhältnisse abzubilden, wird das bereits 1866 eingeführte und heute als Klade definierte[3] Taxon Amniota genutzt, das alle rezenten Reptilien einschließlich der Säuger und Vögel sowie alle mittlerweile ausgestorbenen Nachfahren ihres letzten gemeinsamen Vorfahren einschließt. Die 1864 eingeführten und heute ebenfalls als Klade definierten Sauropsida umfassen alle rezenten Reptilien einschließlich der Vögel sowie alle ausgestorbenen Formen, die näher mit den heutigen Reptilien und Vögeln verwandt sind als mit den Säugetieren. Tatsächlich sind alle rezenten Reptilien näher mit den Vögeln verwandt als mit den Säugetieren, d. h. alle Reptilien der Entwicklungslinie, die zu den Säugetieren führt, sind heute ausgestorben. Verschiedene Wissenschaftler versuchten sich ab Ende der 1980er Jahre an einer phylogenetischen Definition des Taxons Reptilia, um dieses wissenschaftlich weiter nutzbar zu machen.[1] Insbesondere in wissenschaftlichen Veröffentlichungen aus dem englischsprachigen Raum wird Reptilia heute teils synonym für die Klade Sauropsida verwendet.[1][4]
Die wissenschaftliche Beschäftigung mit Reptilien fällt in das Gebiet der Herpetologie. Das Wissen um ihre Pflege und Zucht in Terrarien bezeichnet man als Terraristik oder Terrarienkunde, die ein Teil der Vivaristik ist.
Merkmale


Das kennzeichnendste Merkmal der rezenten Reptilien (hier gemeint als Nicht-Vogel-Sauropsiden) ist ihre trockene, schleimlose, aus Hornschuppen bestehende Körperbedeckung. Von Vögeln und Säugetieren unterscheiden sie sich durch das Fehlen von Federn bzw. Haaren. Bei Schuppenkriechtieren überlappen sich die Hornschuppen in der Regel dachziegelartig, bei Schildkröten und Krokodilen tun sie dies nicht. Eine „echte“ Häutung (Ecdysis), das periodische Abstreifen größerer zusammenhängender Partien der Oberhaut, tritt prinzipiell nur bei Schuppenkriechtieren und besonders ausgeprägt bei Schlangen auf.[5]
Die meisten heute lebenden Reptilien besitzen einen typischen echsenartigen Habitus, das heißt, sie haben einen langen Schwanz, laufen auf vier Beinen (Quadrupedie), und bewegen sich im Spreizgang. Dies ist der ursprüngliche Habitus der Landwirbeltiere, der bereits bei den Vorfahren der Reptilien vorhanden war. Alle Schlangen und einige Echsen weichen von diesem urtümlichen Bauplan ab, indem ihre Beine und Extremitätengürtel zurückgebildet sind und Hals, Rumpf und Schwanz ansatzlos ineinander übergehen. Ebenfalls relativ stark abgeleitete rezente Reptilien sind die Schildkröten, bei denen der Rippenkorb und die Rumpfbeschuppung insbesondere bei den Landschildkröten eine Art Gehäuse bilden, in das sie sich zurückziehen können. Der echsenartige Habitus der Krokodile ist jedoch nicht von deren Vorfahren ererbt, sondern sekundär erworben (siehe Abstammungsgeschichte). Dies zeigt sich u. a. daran, dass Krokodile, wenn sie schnell laufen, ihren Rumpf, anders als Echsen, nicht in der horizontalen Ebene winden und ihre Beine unter den Körper stellen.
Im Gegensatz zu den Amphibien sind alle Reptilien, wie auch Vögel und Säugetiere, zeit ihres Lebens Lungenatmer, sie durchlaufen also kein aquatisches, kiemenatmendes Larvenstadium.
Die meisten rezenten Formen legen Eier (Oviparie), nur einige wenige gebären lebende Junge (Viviparie) oder sind eierlebendgebärend (Ovoviviparie). Die Eier sind bei den meisten Schuppenkriechtieren mit einer pergamentartigen, flexiblen Schale umhüllt. Die Eier vieler Schildkröten und aller Krokodile besitzen hingegen eine relativ feste Kalkschale. Der Grad der Verkalkung gilt als Grad der Anpassung an schwankende Bedingungen hinsichtlich der Feuchtigkeit des Milieus, in dem die Eiablage erfolgt: Die Eier mit der am stärksten verkalkten Schale sind am besten sowohl gegen das Eindringen von Wasser als auch gegen Austrocknung geschützt.[6][7]
Die rezenten Reptilien sind ektotherme und wechselwarme (poikilotherme) Tiere, die ihre Körpertemperatur so weit wie möglich durch Verhalten regulieren (z. B. Sonnenbaden). Des Weiteren weist der Blutkreislauf aller rezenten Reptilien keine vollständige Trennung von Lungen- und Körperkreislauf auf. Bei den meisten Formen ist dies durch eine nicht durchgehende Herzscheidewand verwirklicht. Krokodile hingegen haben eine geschlossene Herzscheidewand und der Blutaustausch erfolgt im Aortenstamm über eine Öffnung in der Trennwand zwischen linker und rechter Aorta (Foramen Panizzae).
Abstammungsgeschichte


Die stammesgeschichtlich ersten Reptilien sind zugleich auch die stammesgeschichtlich ersten Amnioten bzw. sind die frühesten Amnioten durchweg reptilienartige Formen. Sie sind aus dem frühen Oberkarbon, einer Zeit vor etwa 315 Mio. Jahren, erstmals fossil überliefert. Alle Amnioten und damit auch alle Reptilien stammen von ursprünglichen Landwirbeltieren („Amphibien“ im weiteren Sinn) ab. Im Unterschied zu den Reptilien pflanzten sich diese Amphibien nicht über ein amniotisches Ei fort, eine Art autonome Überlebenskapsel,[8] die den sich entwickelnden Embryo bzw. Fötus mit Nährstoffen versorgt und vor Austrocknung schützt. Die Amnioten sind daher im Gegensatz zu den Amphibien zur Fortpflanzung nicht auf Gewässer angewiesen und damit generell besser an trockene Lebensräume angepasst. Mit den modernen Amphibien hat sich mindestens eine Linie der ursprünglichen Landwirbeltiere bis heute gehalten, jedoch sind diese überwiegend auf feuchte Lebensräume spezialisiert und mit den unmittelbaren Vorfahren der Amnioten bzw. Reptilien nicht zu vergleichen, die bereits relativ unabhängig vom Wasser gewesen sein müssen (siehe → Reptiliomorpha).
Während die Skelettreste des ersten „echten“ Reptils Hylonomus in einem fossilen tropischen Feuchtwald („Steinkohlenwald“) überliefert sind,[9] zeigen etwa gleich alte (ca. 315 Mio. Jahre) Spurenfossilien die Existenz früher Amnioten in einer zumindest saisonal wasserarmen Umwelt, in der das amniotische Ei sehr wahrscheinlich einen Fortpflanzungsvorteil bedeutet hat.[10]
Die Amniota waren schon im Oberkarbon in zwei Hauptlinien aufgespalten: Die eine, Synapsida genannte Linie (siehe Systematik) führte zu den Säugetieren und die andere, Sauropsida genannte Linie führte zu den rezenten Reptilien und den Vögeln. Im traditionellen paläontologischen Verständnis werden auch die frühen Vertreter der Synapsidenlinie („Pelycosaurier“ und frühe Therapsiden) zu den Reptilien gezählt.
Die Sauropsiden waren wiederum bereits im Oberkarbon in zwei Hauptlinien gespalten: Parareptilien und Eureptilien, wobei das älteste bekannte Parareptil aus dem höchsten Oberkarbon stammt[11] und damit rund 15 Millionen Jahre jünger ist als Hylonomus, das erste Eureptil. Die Parareptilien haben keine rezenten Vertreter, denn mit den Procolophonida stirbt in der späten Trias die letzte ihrer Untergruppen aus. Zu den populärsten Parareptilien zählen die schon im Perm wieder ausgestorbenen Pareiasaurier.
Folglich sind die meisten postpermischen und alle rezenten Reptilien (sowie die Vögel) Vertreter der Eureptilia. Fast alle Eureptilien und alle rezenten Vertreter gehören einer Großgruppe an, die Diapsida (siehe Systematik) genannt wird. Zwar erscheinen deren früheste Vertreter ebenfalls schon im Oberkarbon, aber zur Blüte kommen sie erst im Mesozoikum, was dieser erdgeschichtlichen Ära den Beinamen „Reptilienzeitalter“ einbringt.
Bei vielen mesozoischen Diapsiden unterlag der ursprünglich echsenartige Habitus einem deutlichen Wandel: Die Dinosaurier gingen zu einer zweibeinigen Fortbewegung über, die Flugsaurier entwickelten Flügel, und mehrere Gruppen passten sich einem Leben im Meer an und bildeten ihre Gliedmaßen in Flossen um. Die dahingehend stärkste Anpassung erfolgte bei den Ichthyosauriern, die einen fischartigen Habitus entwickelten, ähnlich wie die heutigen Delfine.
Die Vertreter der unter anderem zu den Dinosauriern und Flugsauriern führenden Diapsiden-Linie werden Archosaurier (Archosauria) genannt. Ihre Wurzeln liegen im Perm. Im Mesozoikum vollzieht sich bei ihnen eine Entwicklung, im Zuge derer diese Formen immer mehr Merkmale verlieren, die heute als typisch reptilienhaft angesehen werden. Sie entwickeln eine Trennung von Lungen- und Körperkreislauf, werden endotherm und bedecken ihre Haut mit isolierendem Material, um die selbsterzeugte Körperwärme besser halten zu können (siehe auch gefiederte Dinosaurier). Während die Dinosaurier im traditionellen Verständnis einmal mehr eine paraphyletische Gruppe mit überwiegend reptilienartigen Vertretern und somit komplett ausgestorben sind, schließen sie nach moderner Auffassung als monophyletisches Taxon die Vögel, den aktuellen Endpunkt des beschriebenen „Entreptilisierungstrends“, mit ein. Eine ähnliche Entwicklung lief in Perm und Trias bereits bei den Synapsiden ab und führte letztlich zu den Säugetieren. In der Entwicklungslinie der Krokodile, einer weiteren Linie der Archosaurier, kehrt sich dieser Trend jedoch um und deren Vertreter werden im Vergleich zu ihren Vorfahren (siehe → Crocodylomorpha) wieder zunehmend reptilienhaft.
Den Archosauriern gegenüber stehen die Schuppenechsen (Lepidosauria), die mit den Brückenechsen, Eidechsen, Waranen, Geckos, Chamäleons, Schlangen usw. den mit Abstand größten Teil der rezenten Reptilien umfassen. Obwohl sie viele „primitive“, das heißt die heute als reptilientypisch angesehenen, Merkmale aufweisen, sind die meisten rezenten Lepidosauriergruppen stammesgeschichtlich relativ jung und erscheinen frühestens in der Kreidezeit.
Lange Zeit unklar war die systematische Stellung der Schildkröten (Testudinata): Ihr Schädel weist keine Schläfenfenster auf, weshalb diese Gruppe traditionell den Anapsiden (siehe Systematik) zugeordnet wird. Mittlerweile setzt sich jedoch zunehmend die Ansicht durch, dass die Schildkröten Nachfahren diapsider Reptilien sind, welche ihre Schläfenöffnungen sekundär verschlossen haben. Innerhalb der Diapsiden wird sowohl ein engeres Verwandtschaftsverhältnis mit den Archosauriern als auch ein engeres Verwandtschaftsverhältnis mit den Schuppenkriechtieren diskutiert.[12] Die bislang ältesten bekannten Schildkrötenfossilien, die aus der Obertrias stammen, erlauben dahingehend keine Klärung.
Systematik
Klassische Systematik
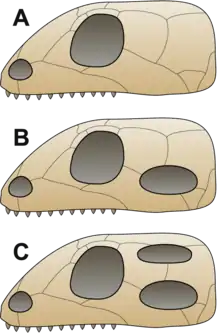
Nach der charakteristischen Anzahl und Lage von Öffnungen im hinteren Teil des Schädeldachs, den Schläfen- oder Temporalfenstern, werden innerhalb der Reptilia klassisch drei Großgruppen unterschieden:
- die Anapsida (kein Schläfenfenster), die im traditionellen Sinn als „Stammamnioten“ gelten und sowohl fensterlose Vertreter der Eureptilia als auch zahlreiche Parareptilien sowie die Schildkröten einschließen
- die Synapsida (ein unteres Schläfenfenster), die im traditionellen Sinn nicht die Säugetiere mit einschließen und damit ein rein fossiles Taxon sind,
- sowie die Diapsida (zwei Schläfenfenster), die im traditionellen Sinn nicht die Vögel mit einschließen.
Im Folgenden wird die Systematik der rezenten Reptilien nach klassischem Verständnis wiedergegeben.
- Unterklasse Schildkröten (Testudines) (→ Systematik der Schildkröten)
- Unterklasse Diapsida
- Infraklasse Schuppenechsen (Lepidosauria)
- Ordnung Brückenechsen (Sphenodon)
- Ordnung Schuppenkriechtiere (Squamata)
- Unterordnung Echsen (Lacertilia, Sauria)
- Infraordnung Leguanartige (Iguania)
- Infraordnung Geckoartige (Gekkota)
- Infraordnung Skinkartige (Scincomorpha)
- Infraordnung Schleichenartige i. w. S. (Anguimorpha)
- Unterordnung Schlangen (Serpentes) (→ Systematik der Schlangen)
- Unterordnung Echsen (Lacertilia, Sauria)
- Infraklasse Archosaurier (Archosauria)
- Ordnung Krokodile (Crocodylia)
- Infraklasse Schuppenechsen (Lepidosauria)
Moderne Systematik (Kladistik)
Das folgende Kladogramm stellt die Gruppe der Reptilien (Reptilia) gemäß der Definition von Modesto und Anderson (2004) vor[1] (Synonym mit Sauropsida) und umfasst die heutigen Reptiliengruppen (rezente Nicht-Vogel-Sauropsiden) sowie alle Tiere, die mit ihnen näher verwandt sind als mit den Säugetieren. Die Untergliederung der Parareptilien folgt Tsuji und Müller (2009),[13] die Untergliederung der Eureptilien folgt Laurin und Reisz (1995).[14]
| Amniota |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Gefährdung
Die IUCN stuft zurzeit (2021) von den gelisteten 8.492 Arten, 2 Arten als in der Natur ausgestorben (Extinct in the Wild), 332 Arten als vom Aussterben bedroht (Critically Endangered), 588 Arten als stark gefährdet (endangered) und 538 Arten als gefährdet (vulnerable), insgesamt 1.460 Arten. 30 Arten gelten bereits als ausgestorben (Extinct). 1.220 Arten können aktuell nicht bewertet werden (data deficient)[15]
Klimawandel
Bei vielen Reptilien ist das Geschlecht der Nachkommen temperaturgesteuert. Bei Geckos schlüpfen Männchen bei mittleren Temperaturen, bei niedrigen oder hohen Temperaturen nur Weibchen. Bei Krokodilen schlüpfen unter 30 °C nur Weibchen, über 34 °C nur Männchen, bei Schildkröten ist die Verteilung ähnlich, aber reziprok. Durch den Klimawandel besteht hier die Gefahr einer einseitigen Geschlechterausprägung die letztlich zum Aussterben führen muss.
Quellen
- Robert L. Carroll: Paläontologie und Evolution der Wirbeltiere. Thieme, Stuttgart (1993), ISBN 3-13-774401-6.
- Wilfried Westheide / Reinhard Rieger: Spezielle Zoologie Teil 2: Wirbel und Schädeltiere, 1. Auflage, Spektrum Akademischer Verlag Heidelberg • Berlin, 2004, ISBN 3-8274-0307-3.
- Laurie J. Vitt, Janalee P. Caldwell: Herpetology – An Introductory Biology of Amphibians and Reptiles. 4. Auflage. Academic Press (Elsevier), 2014, ISBN 978-0-12-386919-7.
- Michel Laurin, Jacques A. Gauthier: Amniota. Mammals, reptiles (turtles, lizards, Sphenodon, crocodiles, birds) and their extinct relatives. Version vom 30. Januar 2012 (The Tree of Life Web Project).
Einzelnachweise
- Sean P. Modesto, Jason S. Anderson: The phylogenetic definition of Reptilia. Systematic Biology. Bd. 53, Nr. 5, 2004, S. 815–821, doi:10.1080/10635150490503026 (alternativer Volltextzugriff: IUCN/SSC Tortoise and Freshwater Turtle Specialist Group PDF 552 kB).
- Stand: November 2021. Quelle: The Reptile Database..
- Michael S. Y. Lee, Patrick S. Spencer: Crown-clades, key characters and taxonomic stability: When is an amniote not an amniote? In: Stuart S. Sumida, Karen L. M. Martin (Hrsg.): Amniote Origins – Completing the Transition to Land. Academic Press, 1997, ISBN 978-0-12-676460-4, S. 61–84.
- P. Martin Sander: Reproduction in early amniotes. Science. Bd. 337, Nr. 6096, 2012, S. 806–808, doi:10.1126/science.1224301
- L. J. Vitt, J. P. Caldwell: Herpetology. 2014 (siehe oben), S. 48 f.
- Mary J. Packard, Gary C. Packard, Thomas J. Boardman: Structure of eggshells and water relations of reptilian eggs. Herpetologica. Bd. 38, Nr. 1 (Sonderausgabe Reproductive Biology of Reptiles), 1982, S. 136–155 (JSTOR 3892368, frei online lesbar).
- Knut Schmidt-Nielsen: Animal Physiology: Adaptation and Environment. 5. Auflage. Cambridge University Press, Cambridge (UK) 1997, ISBN 0-521-57098-0, S. 49.
- auch „persönlicher Teich“ (engl. private pond) genannt, siehe Michael J. Benton: Vertebrate Paleontology. 3. Auflage. Blackwell, Malden MA 2005, ISBN 0-632-05637-1, S. 111.
- Robert L. Carroll: The earliest reptiles. Journal of the Linnean Society (Zoology). Bd. 45, Nr. 304, 1964, S. 61–83, doi:10.1111/j.1096-3642.1964.tb00488.x.
- Howard J. Falcon-Lang, Michael J. Benton, Matthew Stimson: Ecology of earliest reptiles inferred from basal Pennsylvanian trackways. Journal of the Geological Society. Bd .164, Nr. 6, 2007, S. 1113–1118, doi:10.1144/0016-76492007-015 (alternativer Volltextzugriff: The Palaeobiology Research Group PDF 394 kB).
- Sean P. Modesto, Diane M. Scott, Mark J. MacDougall, Hans-Dieter Sues, David C. Evans, Robert R. Reisz: The oldest parareptile and the early diversification of reptiles. Proceedings of the Royal Society B. Bd. 282, Nr. 1801, 2015, doi:10.1098/rspb.2014.1912.
- Rosemary E. Becker, Roldan A. Valverde, Brian I. Crother: Proopiomelanocortin (POMC) and testing the phylogenetic position of turtles (Testudines). Journal of Zoological Systematics and Evolutionary Research. Bd. 49, Nr. 2, 2011, S. 148–159, doi:10.1111/j.1439-0469.2010.00589.x.
- Johannes Müller, Linda A. Tsuji: Impedance-matching hearing in Paleozoic reptiles: evidence of advanced sensory perception at an early stage of amniote evolution. In: PLoS ONE. Bd. 2, Nr. 9, 2007, e889, doi:10.1371/journal.pone.0000889.
- Michel Laurin, Robert R. Reisz: A reevaluation of early amniote phylogeny. In: Zoological Journal of the Linnean Society. Bd. 113, Nr. 2, 1995, ISSN 0024-4082, S. 165–223, doi:10.1111/j.1096-3642.1995.tb00932.x.
- Table 4a: number of animal species in class Reptilia (reptiles) in each IUCN Red List Category by order, abgerufen am 16. Mai 2021 (englisch)
Weblinks
- Deutsche Gesellschaft für Herpetologie und Terrarienkunde (DGHT)
- Österreichische Gesellschaft für Herpetologie (ÖGH)
- naturfilme.com – Videos von einigen Echsen und Schlangen
- amphibien-reptilien.com – Fotos von europäischen Reptilien
- herpetofauna.at – Die Reptilien Österreichs
- reptile-database.org – Umfangreiche Datenbank zur Systematik der Reptilien (engl.)