Pferde
Die Pferde (Equus) sind die einzige rezente Gattung der Familie der Equidae. Zur Gattung gehören die Wildpferde (das Przewalski-Pferd und der heute ausgestorbene Tarpan), die verschiedenen Wildeselformen (der Afrikanische und der Asiatische Esel beziehungsweise der Kiang) sowie wenigstens drei Zebra-Arten (das Steppen-, das Berg- und das Grevyzebra). Sie schließt auch die aus den Wildtieren domestizierten Hausformen ein. Die Anzahl der Arten und ihre Abgrenzung zueinander sind bis heute umstritten. Insgesamt werden häufig sieben oder acht Arten unterschieden, von denen die Mehrzahl in ihrem Bestand gefährdet ist. Die Tiere leben heute im Afrika südlich der Sahara und im südlichen sowie zentralen und östlichen Asien. Die bewohnten Habitate bestehen aus offenen, häufig grasbestandenen Landschaftsräumen, die mitunter auch sehr trocken bis wüstenartig sein können. An diese Regionen sind Pferde durch ihren kräftigen Körperbau und die langen, schlanken Gliedmaßen angepasst. An den Beinen findet sich auch das kennzeichnende Merkmal der Gattung, da sowohl die Vorder- als auch die Hinterfüße jeweils nur einen Zeh aufweisen, der von einem breiten Huf bedeckt wird. Der Rückgang der Zehenanzahl, der den Pferden auch die höherrangige Bezeichnung „Einhufer“ einbrachte, ermöglicht eine schnelle und reibungsarme Fortbewegung in den Steppen- und Savannengebieten.
| Pferde | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Das Grevyzebra (Equus grevyi), eine ostafrikanische Pferdeart | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Equus | ||||||||||||
| Linnaeus, 1758 |
Generell sind Pferde gesellig lebende Tiere. Es lassen sich zwei Gruppentypen in der Sozialgemeinschaft unterscheiden: einer mit Mutter-Jungtiergruppen und einzeln lebenden Hengsten und ein zweiter mit größeren Gruppen aus Stuten und Jungtieren, die auch ein oder mehrere männliche Tiere mit einschließen, die sogenannten „Harems“. Die Ausbildung des einen oder anderen Grundtyps ist in der Regel von äußeren Faktoren abhängig. Dazu zählt vor allem das Nahrungsangebot, das über das Jahr beständig oder – durch den stärkeren Einfluss von Jahreszeiten – auch wechselnd sein kann. Die Hauptnahrung der Tiere besteht aus Gräsern, gelegentlich fressen sie auch Blätter und Zweige. Zum Zerkauen der harten Grasnahrung bildeten sich bei den Pferden Backenzähne mit extrem hohen Zahnkronen heraus, was als ein weiteres typisches Kennzeichen der Gattung herangezogen wird. Der im Vergleich zu anderen Huftieren weniger effiziente Magen-Darm-Trakt bedingt, dass die Pferde den größten Teil ihrer aktiven Zeit mit der Nahrungsaufnahme verbringen. Die Hengste verpaaren sich mit mehreren Stuten, während die Stuten, abhängig von dem sozialen Gruppentyp, in dem sie leben, entweder einen oder mehrere Hengste als Paarungspartner haben. Zumeist wird ein einzelnes Fohlen geboren, das nach maximal sechs Jahren selbständig ist. Der männliche wie auch der weibliche Nachwuchs verlässt anschließend die elterliche Gruppe.
Die Pferde haben eine große Bedeutung in der menschlichen Entwicklungsgeschichte. In urgeschichtlicher Zeit wurden sie als eine wichtige Rohstoff- und Nahrungsquelle genutzt. Im Verlauf der Sesshaftwerdung gelang die Domestizierung von zwei Arten. Das Hauspferd entstand aus einer Form des Wildpferdes, der Hausesel aus dem Afrikanischen Esel. Beide Haustierformen spielen als Reit- und Lasttier eine wichtige Rolle und erlangten im Gefolge des Menschen eine weltweite Verbreitung. Die systematische Erforschung der Gattung begann im Jahr 1758 mit der Etablierung des Gattungsnamens Equus. In der nachfolgenden Zeit wurden verschiedene Untergliederungsvorschläge gemacht, die zumeist aber wenig Bestand hatten. Aus stammesgeschichtlicher Sicht sind die Pferde das jüngste Glied eines gut 56 Millionen Jahre währenden Entwicklungsprozesses der Familie. Die frühesten Vertreter der Gattung der Pferde traten im Pliozän vor rund dreieinhalb Millionen Jahren in Nordamerika auf. Nur wenig später hatten diese frühen Pferde Eurasien und Afrika besiedelt. Der amerikanische Zweig der Pferde starb vor rund 10.000 Jahren aus.
Merkmale
Habitus

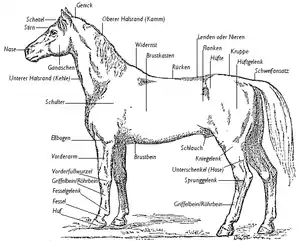
Pferde sind generell stämmige Tiere mit einem walzenförmigen Körper und langem Hals, einem vergleichsweise großen Kopf und langen Gliedmaßen. Größe und Gewicht variieren von Art zu Art: Insgesamt erreichen die Tiere Kopf-Rumpf-Längen von 200 bis 300 cm, der Schwanz wird 30 bis 60 cm lang. Die Schulterhöhe schwankt bei den kleineren Arten wie dem Asiatischen (Equus hemionus) und dem Afrikanischen Esel (Equus asinus) zwischen 110 und 140 cm bei einem Gewicht von 200 bis 275 kg,[1][2] die größte rezente Art, das Grevyzebra (Equus grevyi) wird am Widerrist bis zu 150 cm hoch und wiegt zwischen 350 und 430 kg, in Ausnahmefällen bis zu 450 kg.[3] Der Geschlechtsdimorphismus ist nur gering entwickelt, männliche Tiere übertreffen die weiblichen dadurch nur um rund 10 % an Körpergewicht. Am Kopf ist vor allem der Gesichtsbereich lang ausgezogen. Die Augen liegen seitlich am Kopf, die Ohren sind lang und beweglich. Das Fell ist dicht und häufig kurz, die meisten Arten haben am Nacken, am Schopf und am Schwanz längere Haare, Langhaar oder Mähne genannt. Die Fellfärbung ist bei einigen Arten grau oder braun an der Oberseite und weißlich-grau an der Unterseite. Streifen an Schultern und Gliedmaßen können bei mehreren Arten vorhanden sein. Die drei Zebraarten sind durch ihr auffälliges schwarz-weißes Fellkleid gekennzeichnet. Ein markantes Merkmal bilden die Kastanien (chestnuts) oder „Nachtaugen“, schwielenartige dunkle Flecken an den Beinen. Zebras und Wildesel haben diese vorwiegend an den Vorderbeinen, Wild- und Hauspferde häufig an allen vier Beinen. Größe und Form variieren individuell. Es handelt sich hierbei möglicherweise um verkümmerte Drüsen oder Reste eines Hand- und Fußgelenkpolsters. Die Vorder- und Hinterfüße enden in breiten Hufen, von denen Pferde jeweils nur einen je Gliedmaß besitzen („Einhufer“). Der „Hufschuh“ bedeckt das jeweils letzte Zehenglied vollständig.[4][5]
Schädel- und Gebissmerkmale
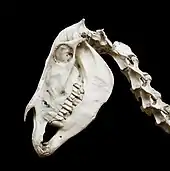
Pferde haben einen wuchtigen Kopf, der Gesichtsschädel ist auffallend langgestreckt und wird vorwiegend vom Oberkiefer gebildet. Auch das Zwischenkieferbein ist verlängert. Das Nasenbein besitzt eine lang-schmale Form. Die Augenhöhle ist weit nach hinten versetzt und liegt hinter den Zähnen. Sie wird vollständig von Knochen umschlossen. Die hintere Schädelabschnitt ist vergleichsweise kurz, die Gehirnkapsel aber relativ groß. Eine Besonderheit der Pferde findet sich in dem Luftsack, der eine Aussackung der Ohrtrompete unterhalb der Schädelbasis darstellt. Die paarige Öffnung besitzt jeweils ein Fassungsvermögen von 350 bis 500 ml. Ursprünglich als behilflich bei der Kühlung des Gehirns interpretiert, dient der Luftsack nach Meinung unter anderem von Horst Erich König und Klaus-Dieter Budras vermutlich – wie die Nasennebenhöhlen – eher zur Herabsetzung des spezifischen Gewichts des Schädels.[6] Der Unterkiefer zeigt sich ebenfalls massig gestaltet. Das Kiefergelenk liegt hoch, der Unterkieferast ist vergrößert. Am hinteren Ende tritt ein kräftiger Winkelfortsatz auf, an dem der Masseter-Muskel ankert.[4][5]

Pro Kieferhälfte besitzen Pferde jeweils drei Schneidezähne, einen Eckzahn und sechs bis sieben hintere Zähne. Die Zahnformel lautet: . Insgesamt besteht das Gebiss somit aus 36 bis 42 Zähnen. Die Schneidezähne sind meißelförmig ausgebildet. Im Inneren besitzen sie Einstülpungen, das sogenannte Infundibulum, das von Zahnschmelz umgeben ist und bei stärkerer Abkauung hervortritt. Es fehlt manchmal beim Steppenzebra (Equus quagga). Dahinter klafft eine als Diastema bezeichnete breite Lücke. In dieser Lücke steht bei männlichen Tieren der Eckzahn. Bei weiblichen Tieren ist er entweder sehr klein oder vollständig zurückgebildet. Die darauf folgenden Backenzähne setzen sich in der Regel aus drei Prämolaren (der vorderste, auch Wolfszahn genannt, ist nur selten vorhanden) und drei Molaren zusammen. Die vorderen Backenzähne ähneln im Bau stark den hinteren, sind also molariform. Auf der Kauoberfläche besteht ein Relief aus gewundenen Schmelzleisten, dazwischen befinden sich Lagen aus Zement und Dentin. Die Backenzähne haben eine säulenförmige Gestalt mit nahezu parallel verlaufenden Seitenlinien, wobei die einzelnen Seitenflächen durch Kanten und Rippeln strukturiert werden. Markant sind die extrem hohen (hypsodonten) Zahnkronen, Prinzipiell ragt aber nur der oberste, aktiv arbeitende Teil aus dem Zahnfach heraus. Der Rest liegt im Kiefer verborgen und wird bei Abnutzung nach und nach herausgeschoben. Die Zahnwurzel selbst ist klein und bleibt offen, bis der Zahn nahezu abgekaut ist. Erst dann schließt sie sich, zu diesem Zeitpunkt ist meist auch eine merkliche Größenzunahme der Wurzel zu verzeichnen. Dieses Wurzelwachstum wird offensichtlich durch die nun stärker wirkenden Scherkräfte beim Kauen als Resultat der niedrigeren Zahnkronen verursacht. Der durch das Herausschieben des Zahns freiwerdende Raum im Zahnfach füllt sich allmählich mit Spongiosa.[7] Die Bezahnung stellt eine ideale Anpassung an harte Grasnahrung dar.[4][5]

Gliedmaßen
Eines der charakteristischsten Merkmale der Pferde ist die Reduktion der Zehenanzahl. So haben alle heute lebenden Arten nur mehr eine funktionale Zehe (Monodaktylie). Es handelt sich dabei um die dritte Zehe, die restlichen Zehen sind zurückgebildet und am Skelett der Gliedmaßen als rudimentäre Griffelbeine erhalten. Die Griffelbeine sind allerdings nicht funktionslos, da sie eine wichtige Stützfunktion für die Sehnen ausüben, die die unteren Gliedmaßen mit den Vorder- beziehungsweise Hinterfüßen verbinden.[8][9] Die Mittelhandknochen sind kürzer als die Mittelfußknochen, was sich auch auf die Gesamtlänge der Vorder- und Hinterbeine auswirkt. Pferde haben wie alle Unpaarhufer ein sattelförmiges Talonaviculargelenk – das Sprunggelenk zwischen Sprungbein (Talus) und Kahnbein (Naviculare) –, das die Beweglichkeit deutlich einschränkt. Die Elle ist stark reduziert und in der unteren Hälfte mit der Speiche verschmolzen. Ebenso fusioniert das untere Ende des Wadenbeins vollständig mit dem Schienbein. Der Oberschenkelknochen ist vergleichsweise kurz, allerdings am oberen Schaftabschnitt unterhalb des Gelenkkopfes mit einem großen Knochenfortsatz (Trochanter tertius) versehen. Das Schlüsselbein fehlt.[4][5]
Innere Anatomie
.jpg.webp)
Das Herz der Pferde lässt wie bei allen Wirbeltieren als Muskelpumpe das Blut im ganzen Körper zirkulieren. Es ist in der Form mehr globoid als das menschliche Herz und besteht aus vier Kammern: dem linken und rechten Vorhof und dem linken und rechten Ventrikel. Das durchschnittliche erwachsene Pferd hat ein 3,4 kg schweres Herz, das entspricht etwa 0,6 bis 0,7 % des Körpergewichts.[10] Bei Hauspferden kann es etwas in der Größe als Antwort auf Konditionierung zunehmen.[11][12] Die Kreislaufkapazität wird zum Teil durch die funktionelle Masse des Herzens und der Milz bestimmt.[13] Die Herzfrequenz bei Tieren in Ruhe beträgt 15 bis 45 Schläge je Minute. Sie kann bei anstrengenden Aktivitäten um das Dreifache ansteigen. Untersuchungen an Zebras zeigen, dass Jungtiere generell eine höhere Herzfrequenz als Alttiere haben. Dagegen führt eine größere Muskelmasse zu langsameren Herzschlägen.[10][14][15]
Pferde sind wie alle Unpaarhufer Enddarmfermentierer, das heißt, die Verdauung findet größtenteils erst im Darmtrakt statt. Der Magen ist – im Gegensatz etwa zu dem der Wiederkäuer – stets einfach gebaut und einkammerig mit einer Länge von rund 74 beziehungsweise 14 cm (gemessen über die äußere und innere Krümmung) und einem Volumen von 0,8 l beim Hausesel. Die Fermentation findet im sehr großen Blinddarm und im doppelschlingigen, 2 bis 4 m langen aufsteigenden Grimmdarm (Colon ascendens) statt. Die pH-Werte im Dünndarm steigen von vorn nach hinten an und reichen von 6,3 bis 7,5. Im nachfolgenden Dickdarm fallen sie wieder auf rund 6,7. Ebenso nimmt der individuelle Anteil der Mikroflora im Dünndarm zu, im vorderen Abschnitt beträgt er etwa 2,9 × 106 je Gramm (Nassgewicht), im hinteren etwa 38,4 × 106. Im Blinddarm und im Dickdarm belaufen sie sich auf 25,9 beziehungsweise auf 6,1 × 106.[16] Der Blinddarm kann bis zu 33 l Füllmenge, der gesamte Dünndarm bis zu 64 l und der Dickdarm bis zu 96 l aufnehmen. Insgesamt wird der Darmtrakt beim Hausesel bis zu 18 m und beim Hauspferd bis zu 30 m lang.[10][17]
Pferde unterscheiden sich im Bau des Eierstocks von anderen Säugetieren: Das gewöhnlich als „Rinde“ bezeichnete Eierstockgewebe mit den Follikeln liegt bei ihnen im Inneren des Organs, das gefäßführende Eierstockmark dagegen außen. Die Eierstockrinde reicht nur an einer Stelle an die Oberfläche. Diese Stelle ist als Einziehung auch von außen sichtbar und wird als „Ovulationsgrube“ (Fossa ovarii) bezeichnet, nur an dieser Stelle kann der Eisprung (Ovulation) erfolgen. Der sprungreife Follikel erreicht einen Durchmesser von 5 cm und ist damit mehr als doppelt so groß wie der eines Rindes.[18][19] Männliche Tiere haben ein Skrotum, aber wie alle Unpaarhufer keinen Penisknochen. Der Penis selbst wird beim Hausesel im unerigierten Zustand rund 35 cm lang bei einem Durchmesser von etwa 5 cm. Die Hoden wiegen beim Hausesel jeweils zwischen 123 und 136 g. Ihr Gewicht steigt zur Paarungszeit hin deutlich an, beim Steppenzebra erhöht es sich so beispielsweise für beide Hoden kombiniert von rund 268 auf 345 g. Die Größenzunahme ist um so stärker, je strikter die Fortpflanzungsphase jahreszeitlich begrenzt ist.[20] Die Nieren liegen unterhalb der Lendenwirbel und werden beim Hausesel 240 bis 270 g schwer, beim Hauspferd erreichen sie das doppelte bis dreifache Gewicht.[21][22]
Chromosomenzahl
Die Chromosomenzahl der Pferdearten variiert von 2n = 66 bis 2n = 32:
|
2n = 66 |
|
2n = 64 |
|
2n = 62 |
|
2n = 54–56 |
|
2n = 50–52 |
|
2n = 46 |
|
2n = 44 |
|
2n = 32 |
Die Spanne bei E. hemionus wie auch bei E. kiang wird mit der Robertson-Translokation erklärt.[23]
Verbreitungsgebiet und Lebensraum
Die wilden Formen der rezenten Pferdearten leben heute noch im östlichen und südlichen Afrika und in den zentralen Regionen Asiens. Als Lebensraum bevorzugen Pferde offenes Gelände. Sie finden sich in Savannen und Steppen, aber auch in trockeneren Habitaten wie Halbwüsten und Wüsten. Die Lebensräume umfassen aber nicht nur einheitliche grasbestandene Gebiete, sondern schließen zum Teil auch Busch- und Waldlandschaften ein. Die Nutzung geschlossener Habitate ist insbesondere davon abhängig, wie gut die einzelnen Arten blatthaltige Nahrung verwerten können. Außerdem spielt der Einfluss von Beutegreifern eine Rolle. Des Weiteren unterliegen die verschiedenen Regionen im Verbreitungsgebiet der heutigen Pferde jahreszeitlichen Veränderungen durch variierenden Niederschlag (Regen in den eher tropischen und Schnee in den eher gemäßigten Klimazonen). Das Vorhandensein verfügbarer Wasserquellen ist darüber hinaus ein wichtiges Kriterium für die Anwesenheit der Pferde in einem bestimmten Gebiet.[4]
In den letzten Jahrtausenden ist das Verbreitungsgebiet der Pferde deutlich zurückgegangen. Bis zum Ende des Pleistozäns waren sie über weite Teile Eurasiens, Afrikas und Amerikas verbreitet. Auf dem amerikanischen Kontinent sind sie vor rund 10.000 Jahren aus nicht genau geklärten Gründen ausgestorben. Erklärungsversuche reichen von der Bejagung durch die neu eingewanderten Menschen über klimatische Veränderungen nach dem Ende der letzten Eiszeit bis hin zu Seuchen oder einer Kombination dieser Faktoren. Zumindest in Südamerika begannen die Verbreitungsgebiete der Pferde im ausgehenden Pleistozän bei Anwesenheit der ersten frühen menschlichen Besiedler rapide zu schrumpfen.[24] Auch in Teilen Europas dürften einzelne Bestände vor rund 10.000 Jahren ausgestorben sein. In Nordafrika und Westasien wurden sie vermutlich in der Antike ausgerottet – lediglich im Irak und im Iran hielt sich eine Population des Asiatischen Esels bis ins 20. Jahrhundert. Im östlichen Europa ist die letzte wildlebende Pferdeart – der Tarpan (Equus ferus) – im 19. Jahrhundert ausgestorben.
Im Gegensatz dazu wurden Hauspferd und Hausesel vom Menschen weltweit verbreitet, in einigen Ländern gibt es auch verwilderte Populationen beider Formen. Die größte Anzahl verwilderter Hauspferde und -esel lebt jeweils in Australien, aber auch in den USA und anderen Ländern sind sie zu finden.[4]
Lebensweise
Allgemein
Obwohl Pferde auch tagsüber auf Nahrungssuche gehen können, sind sie vorwiegend dämmerungs- und nachtaktiv. Dies gilt vor allem für die Arten der tropischen Regionen. In den gemäßigteren Breiten kann auch Tagesaktivität überwiegen. In der Regel verbringen die Tiere zwischen 60 und 70 % ihrer aktiven Zeit mit der Nahrungsaufnahme. Die übrige Zeit entfällt auf Wanderungsbewegungen und soziale Interaktionen wie Körperpflege, Spiel oder Kämpfe unter Hengsten. Bei der Fortbewegung setzen Pferde nur das letzte Zehenglied auf, sie sind also Spitzengänger. Es kommen mehrere natürliche Gangarten vor, die sich in der Geschwindigkeit sowie der Ausführung unterscheiden. Sie reichen von einem langsamen Gehen (Schritt) mit individueller Beinbewegung im Passgang über den schnelleren Trott (Trab), bei dem diagonal zwei Beine gleichzeitig bewegt werden, bis hin zum schnellen Rennen (Galopp). Bei letzterem heben alle vier Beine üblicherweise gleichzeitig vom Boden ab. Erreichte Geschwindigkeiten liegen beim Gehen bei 6 bis 10 km/h, beim Trott bei 6 bis 19 km/h und beim Galopp bei 26 bis 56 km/h und mehr.[10][9] Schlafphasen sind mit durchschnittlich 2,5 Stunden täglich eher kurz. Hierbei überwiegen kurze Schlafpausen von nur wenigen Minuten bis rund einer Viertelstunde. Ruhephasen insgesamt können aber länger sein. Zumeist stehen die Tiere dabei, lediglich Jungtiere legen sich hin. Die typische Schlafpose zeichnet sich durch schräggestellte Hinterbeine, einen herabgesenkten Hals und hängende Ohren aus. Die Augen sind häufig offen. Es handelt sich hierbei um charakteristische Merkmale von Fluchttieren. Das Schlafen oder Dösen im Stehen wird den Tieren dadurch ermöglicht, dass sie durch die Schrägstellung der Beine die Kniescheibe fest verankern und so ein Abknicken verhindern können.[4]
Sozial- und Territorialverhalten


Pferde verfügen über ein komplexes soziales Verhalten. Prinzipiell lassen sich zwei verschiedene Grundtypen der Gruppenbildung erkennen:
- Das Grevyzebra, der Afrikanische Esel, der Kiang sowie einige Populationen des Asiatischen Esels (unter anderem der Khur und der Onager) zeigen ein territoriales Verhalten. Die größte Bindung besteht zwischen dem Mutter- und dem Jungtier. Männliche Tiere etablieren Paarungsreviere, die mitunter über 10 km² groß sein können. Sie verpaaren sich mit Stuten, die deren jeweilige Territorien durchqueren. Obwohl sich manchmal Tiere zu Verbänden zusammenfinden, gibt es bei diesen Arten keine dauerhaften Beziehungen zwischen erwachsenen Tieren.[4]
- Das Berg- und Steppenzebra, verschiedene Populationen des Asiatischen Esels (etwa der Kulan und der Dschiggetai) sowie das Przewalski-Pferd und verwilderte Hauspferde wie zum Beispiel die Mustangs leben dagegen in größeren, stabilen Verbänden. Diese bestehen aus ein bis fünf Mutter-Jungtierpaaren und werden von einem Hengst überwachend und treibend begleitet. Das Gemeinschaftsgefüge wird als „Harem“ bezeichnet. Unter Umständen – wie bei einigen verwilderten Hauspferdgruppen – können auch mehrere Hengste einer Gruppe angehören.[25][26] Die Größe eines solchen Verbandes ist stark abhängig vom regionalen Nahrungsangebot. Die Gruppen durchstreifen ausgedehnte Aktionsräume, die sich mit denen anderer Gruppen überschneiden können. Innerhalb des Verbandes lässt sich eine gewisse Rangordnung feststellen, wobei diese nicht unbedingt identisch mit der Führungsrolle ist. So kann bei verwilderten Hauspferden der Aufbruch von allen Gruppenmitgliedern unabhängig vom Geschlecht initiiert werden, häufig folgen die Tiere aber höherrangigen Individuen.[27] Bei den Gruppen des Bergzebras wechselt die Führungsrolle, unter Umständen leitet der Hengst an (beim Erreichen von Nahrungs- oder Wasserquellen) oder eine ranghöhere Stute – meist jene mit dem aktuell jüngsten Fohlen – signalisiert den Aufbruch.[28][29] Männliche Tiere, die keinen Gruppenverband anführen, formieren sich häufig zu Junggesellengruppen. Gelegentlich bilden sich auch sehr große Herden, die sich dann aus mehreren Gruppenverbänden zusammensetzen. Diese haben jedoch nur einen temporären Charakter und zerfallen nach einer gewissen Zeit wieder. Beim Steppenzebra geschieht dies unter anderem in Regionen, in denen große Junggesellengruppen auftreten. Dadurch können die Hengste der einzelnen Gruppenverbände ihren Harem besser vor Übergriffen schützen.[4]
Der erstgenannte Typ mit Mutter-Jungtiergruppen und territorialen Hengsten kommt weitgehend in Gebieten mit einem über das Jahr mehr oder weniger gleichbleibenden Nahrungsangebot vor. Die Wanderungsbewegungen der Tiere sind wenig weitschweifend. Das Paarungsrecht liegt ausschließlich bei dem Hengst, durch dessen Territorium eine Mutter-Jungtiergruppe wandert. Der zweite Typ mit Haremsbildung und überwachendem Hengst ist typisch für Landschaften mit starken jahreszeitlichen Schwankungen und somit variierendem Nahrungsangebot im Verlauf eines Jahres. Die Gruppenverbände durchstreifen weite, offene Landschaften auf der Suche nach Nahrung, unterliegen dabei aber der andauernden Gefahr, von Fressfeinden bedroht zu werden. Eine größere Gruppenbildung verteilt die Wachsamkeit vor Beutegreifern auf mehrere Tiere, ohne dass ein einzelnes Individuum zu viel Zeit opfern muss, die es eigentlich für die Nahrungsaufnahme braucht. Ein anwesender Hengst in einer solchen Gruppe, der in der Regel beständig Ausschau nach potentiellen Konkurrenten hält, reduziert für die Stuten zusätzlich den Zeitaufwand der Wachsamkeit. Hier liegt das Paarungsrecht bei dem Hengst, der einen Harem anführt. Beide Grundtypen der Gruppenbildung ermöglichen es dabei dem Hengst, sich mit mehreren Stuten zu paaren und sich so polygyn zu vermehren. Stuten in Haremsverbänden leben dagegen monandrisch (verpaaren sich nur mit einem Hengst), solche in Mutter-Jungtierpaaren dagegen polyandrisch (verpaaren sich mit mehreren Hengsten). Häufig wird angenommen, dass der erste Grundtyp der ursprünglichere ist, der auch bei zahlreichen sehr frühen Vorfahren der heutigen Pferde ausgebildet war. Der zweite wäre dann ein abgewandelter Gruppentyp, der mit dem Aufkommen offener Landschaften und einer stärkeren Saisonalisierung des Klimas aufkam.[30][4]

Die Kommunikation der Pferde mit Artgenossen erfolgt auf vielfältigen Wegen über visuelle Zeichen und taktile und olfaktorische Signale und mittels Lauten. Das Repertoire an Gesten ist sehr umfangreich und drückt sich über die Haltung des Kopfes, der Ohren, des Kiefers oder des Schwanzes sowie Beinbewegungen wie das Aufstampfen aus. Ebenso gibt es ein variantenreiches Lautspektrum, das neben dem bekannten Wiehern auch verschiedene Bell-, Schnaub- und Blaslaute einschließt. Die Laute sind am vielfältigsten bei den verwilderten Hauspferden und bei den Wildpferden. Zebras und Wildesel sind meist mit Ausnahme der männlichen Tiere ruhiger. Nicht alle Gesten und Laute haben einen aggressiven Charakter, einige sind als Begrüßungsformel aufzufassen, drücken Wohlbefinden aus oder dienen bei Arten in Gruppenverbänden auch als Kontaktrufe. Von großer Bedeutung bei allen Pferden einschließlich des Hauspferds[31] ist die Kommunikation über Kot und Urin, die als Informationsträger über räumliche und zeitliche Distanzen fungieren. Dadurch erhalten die einzelnen Tiere Informationen über andere Individuen. In der Regel defäzieren weibliche Tiere an Ort und Stelle, während männliche überwiegend strategischer vorgehen und ihre Ausscheidungen neben denen der Weibchen ablegen. Bei einigen Arten bestehen feste Kotplätze, etwa an viel begangenen Pfaden oder an sozialen Sammelstellen. Zur taktilen Kommunikation gehört unter anderem das gegenseitige Kinnauflegen auf den Rücken des Partners, was allgemein die Aggressivität mindert und die gegenseitige soziale Bindung stärkt. Daneben lecken und putzen sich die Tiere gegenseitig oder knabbern Parasiten weg, was allerdings bei Zebras sehr selten vorkommt. Jede der genannten Kommunikationsformen übermittelt eigene Informationen. So geben Gerüche Hinweise auf das Individuum und seine Persönlichkeit selbst. Laute wiederum verraten den Status eines Tieres, da unter anderem dominante Hengste häufig ausdauerndere und höherenergetische Tonfolgen von sich geben als untergeordnete Tiere, die zudem guttural enden. All dies hilft den Tieren bei Begegnungen fremde Individuen einzuschätzen. Untersuchungen zufolge enden weniger als 15 % der Begegnungen von Einzeltieren oder Gruppen in physischen Konfrontationen. Lassen sich diese trotz der verschiedenen Signalgebungen nicht verhindern, werden sie mit Tritten der Vorder- und Hinterbeine und mit Bissen ausgeführt.[4]
Ernährung

Pferde sind ausschließlich Pflanzenfresser und nehmen in erster Linie Gräser zu sich. Aufgrund der harten Kieselsäure in diesen Pflanzen entwickelten sich bei den Pferden hochkronige Backenzähne mit einem hohen Zahnzementanteil, um dem verstärkten Abrieb beim Kauen entgegenzuwirken. In unterschiedlichem Ausmaß werden aber auch weichere Pflanzenteile wie Blätter oder Zweige gefressen,[32][33] einige ausgestorbene Equus-Arten waren auch an gemischte Pflanzenkost angepasst, wie beispielsweise das Kap-Zebra (Equus capensis).[34] In der Regel steht die qualitativ höherwertige Nahrung nur zu bestimmten Jahreszeiten zur Verfügung. Im Fall des Steppenzebras können sich dann bei ausreichendem Angebot verschiedene Gruppenverbände temporär zu großen Herden zusammenfinden. In der Regel wird die Nahrung mit den feinfühligen Lippen abgezupft und hinter die Schneidezähne geschoben. Als Enddarmfermentierer benötigen Pferde nahezu das Doppelte an Nahrung wie ähnlich große Wiederkäuer. Dies bedingt auch, dass der größte Teil ihrer Tagesaktivität der Nahrungsaufnahme gewidmet ist. Außerdem können sie stickstoffhaltige Bestandteile weniger gut abbauen und müssen sie über den Urin abführen. Dies hat eine größere Wasserabhängigkeit zum Ausgleich des Flüssigkeitsverlustes zur Folge, weswegen die Wanderungsbewegungen vor allem in den trockenen Jahresabschnitten begrenzt sind. Einige Arten wie das Grevyzebra oder der Afrikanische Esel können notfalls auch längere Zeit ohne Wasser auskommen, sie gleichen ihren Wasserhaushalt nach einer derartigen Phase aber mit einer Aufnahme von bis zu 30 l Wasser in kürzester Zeit wieder aus.[2][3] Dies betrifft aber weitgehend nur Tiere, die keinen Nachwuchs aufziehen. Das Verhalten ist auch vom Hauspferd bekannt und führte zur Redensart „saufen wie ein Pferd“.[35] Da das Verdauungssystem der Pferde insgesamt weniger effektiv ist als das von Wiederkäuern, ist auch der Kot entsprechend gröber ausgebildet. Jungtiere lernen in der Regel vom Muttertier, welche Nahrung nährstoffreich und für sie verwertbar ist. Unter Umständen fressen sie den Dung der Mutter, was möglicherweise den Lernprozess unterstützt. Umgekehrt vertilgen auch Muttertiere den Kot der Jungen. Hier wird vermutet, dass die Muttertiere dadurch Krankheitserreger ausfindig machen und ihrem Nachwuchs über die Muttermilch entsprechende T-Lymphozyten zur Abwehr verabreichen.[4]
Fortpflanzung

Die Pferdearten der eher temperierten Klimazonen haben eine jahreszeitlich begrenzte Fortpflanzungsphase, in den stärker warmklimatisch beeinflussten Gebieten kann sie das gesamte Jahr über andauern. Es gibt hier jedoch auch geburtenstarke Jahresabschnitte, die meist mit den Regenzeiten zusammenfallen. Alle Pferde sind polyöstrisch, sodass sich der Sexualzyklus bis zur Befruchtung wiederholt oder aber in den nördlicheren Breiten durch die kürzer werdenden Tage zum Winter hin zum Erliegen kommt. Der Zyklus selbst dauert rund eine Woche und startet bei ausbleibender Befruchtung nach zwei bis drei Wochen erneut. Die Zwischenphase ist bei Zebras länger als bei den Wildpferden und Wildeseln. Der Eisprung findet spontan statt. Fortpflanzungswillige Hengste beschnüffeln die Genitalien der Stuten und legen ihren Kopf auf deren Rumpf. Mit Hilfe des charakteristischen Flehmens können sie am Urin den Estrogenstatus einer Stute über das Jacobson-Organ bestimmen. Männliche Tiere führen untereinander einen Paarungswettstreit aus, der ritualisiert ist und mit einer Aufwölbung des Nackens beginnt, gefolgt von verschiedenen Lautäußerungen und einem gegenseitigen Beschnüffeln. Die letzte Stufe besteht in einem physischen Kampf. Ihre eigene Unwilligkeit drücken weibliche Tiere durch Ausschlagen mit den Hinterbeinen aus und verhindern so ein Aufsteigen des Hengstes. Empfangsbereite Stuten stehen dagegen meist still. Hengste in Haremsverbänden vollziehen je Stute nur einen Paarungsakt, wogegen territoriale Hengste alle 15 Minuten auf einer Stute aufsitzen und etwa jede Stunde ejakulieren. Nach dem Orgasmus benötigt der Hengst bis zu 20 Minuten Erholungszeit.[4]

Die Tragzeit der Pferde beträgt 330 bis 390 Tage, sie ist am längsten beim Grevyzebra und am kürzesten beim Hauspferd. In der Regel kommt ein einzelnes Jungtier zur Welt. Die Geburt erfolgt meist nachts. Das Neugeborene ist relativ schwer (es wiegt zwischen 25 und 40 kg, was etwa 9 bis 13 % des Gewichts der Mutter entspricht) und frühreif. Es kann der Mutter schon wenige Stunden nach der Geburt folgen und bleibt zumeist dicht bei ihr. Durchschnittlich saugt es etwa jede Stunde für eine Minute Milch. Vatertiere beteiligen sich nicht an der Aufzucht des Nachwuchses. Gelegentlich kommt aber Infantizid vor. Nach 7 bis 18 Monaten wird das Jungtier entwöhnt. Die Geschlechtsreife tritt mit zwei bis sechs Jahren ein, wobei sich Junghengste aufgrund der Sozialstrukturen meist erst in höherem Alter fortpflanzen können als Stuten. In Haremsgemeinschaften verlassen sowohl männliche als auch weibliche Tiere dann die elterliche Gruppe, was relativ selten bei Säugetieren vorkommt. Die Lebenserwartung von Pferden in freier Wildbahn liegt bei 21 bis 40 Jahren.[4][5]
Prinzipiell sind Stuten körperlich befähigt, sich jedes Jahr fortzupflanzen, häufig liegt aber ein Zeitraum von mehreren Jahren zwischen zwei Geburten. Der Brunstzyklus setzt schon drei bis vier Wochen nach der Geburt wieder ein. Das Geschlechtsverhältnis bei der Geburt liegt bei 1:1, individuell kann dies bei den Stuten aber sehr unterschiedlich sein. Bei einigen Populationen des Asiatischen Esels bringen junge und ältere Stuten eher weiblichen Nachwuchs zur Welt, mittelalte Tiere hingegen gebären häufiger männliche Fohlen. Gründe hierfür könnten in den stark variablen Fortpflanzungschancen der ausgewachsenen Hengste liegen. Muttertiere müssen daher mehr Zeit und Energie in die Aufzucht des männlichen Nachwuchses investieren, damit dieser sich später erfolgreich gegen Geschlechtsgenossen durchsetzen und anschließend verpaaren kann. Mittelalte Stuten besitzen in der Regel genug Erfahrung und haben die körperliche Voraussetzung für die intensivere Aufzucht männlicher Tiere. Junge Stuten sind demgegenüber meist unerfahren, während ältere vielfach eine nachlassende Konstitution aufweisen. Das überwiegende Gebären weiblichen Nachwuchses durch junge und alte Stuten fördert in diesem Fall die Vitalität einer Population und erhöht die Anzahl an weiblichen Tieren, die später wiederum Jungtiere austragen können.[4]
Feinde und Feindverhalten
Pferde haben eine Reihe natürlicher Feinde, dazu zählen in erster Linie große Raubtiere wie Hyänen, Wölfe, Wildhunde und Großkatzen. Sie sind wie viele Huftiere Fluchttiere. Der Körperbau der Pferde ist auf schnelles und ausdauerndes Laufen ausgelegt, daher flüchten sie bei Bedrohung. Wenn sie in die Enge getrieben werden, können Pferde auch mit den Hufen treten oder Angreifern schmerzhafte Bisswunden zufügen. Ihre wirkungsvollste Waffe sind die stark bemuskelten Hinterbeine. Bei Haremsgemeinschaften verteidigt der Hengst die Gruppe. Die weiblichen Tiere entfernen sich dabei mit rund der halben Fluchtgeschwindigkeit von der Gefahrenquelle, um dem Hengst die Möglichkeit zum Aufschließen zu geben.[4][5]
Systematik
Äußere Systematik
|
Die Pferde (Equus) bilden eine Gattung aus der im Deutschen gleichnamigen Familie der Pferde (Equidae). Die Familie entstand bereits im Unteren Eozän vor 56 Millionen Jahren und war seit dieser Zeit formenreich in Nordamerika und Eurasien verbreitet, im Übergang vom Unteren zum Mittleren Miozän vor rund 16 Millionen Jahren erreichten die ersten frühen Vertreter auch Afrika. Heute stellt Equus das einzige Mitglied seiner Familie dar, wodurch diese monotypisch ist. Die nächsten lebenden Verwandten sind die Tapire und die Nashörner, gemeinsam bilden sie die Ordnung der Unpaarhufer (Perissodactyla). Allerdings sind Tapire und Nashörner enger miteinander verwandt und formen zusammen die Unterordnung Ceratomorpha, denen auch zahlreiche, heute ausgestorbene Formen zugewiesen werden. Die Pferde stehen traditionell den Ceratomorpha gegenüber. Sie gehören zusammen mit ihren ausgestorbenen Vorfahren in die Unterordnung Hippomorpha (Pferdeverwandte). Innerhalb dieser Unterordnung wird die Überfamilie Equoidea unterschieden, die sich aus der Familie der Pferde und der ausgestorbenen Familie der Palaeotheriidae zusammensetzt. Manchmal werden auch die Brontotheriidae, eine ebenfalls fossile, teils sehr große Formen umfassende Gruppe aus dem Eozän in die Unterordnung Hippomorpha verwiesen und gelten damit auch als näher mit den Pferden verwandt.[37] Die Abtrennung der Linie der Pferde von jener der Nashörner und Tapire erfolgte laut molekulargenetischen Untersuchungen vor wenigstens 54 bis 56 Millionen Jahren.[38][39]
Innerhalb der Familie der Pferde wird die Gattung Equus in die Unterfamilie der Equinae gestellt. Deren Vertreter zeichnen sich durch eine bessere Anpassung an Grasnahrung aus und entwickelten daher hochkronige (hypsodonte) Zähne.[32] Hierin wiederum gehört Equus in die Tribus der Equini und die Untertribus der Pliohippina. Die Pliohippina umfassen die einhufigen (monodactylen) Pferde, ein Merkmal, das auch für alle heutigen Vertreter aus der Gattung Equus typisch ist. Sie stellen wiederum die Schwestergruppe der Protohippina dar, die als dreihufige (tridactyle) Tiere etwas urtümlicher gestaltet sind. Die Equini ihrerseits stehen den Hipparionini gegenüber (mitunter werden die Protohippina auch auf Tribusebene (Protohippini) innerhalb der Equinae geführt und bilden dann die Schwestergruppe zu den Hipparionini). Als nächster Verwandter zu Equus gilt aus anatomischen Gründen Dinohippus, das im Übergang vom Miozän zum Pliozän in Nordamerika lebte.[40][36][41][42]
Innere Systematik
Innere Systematik der Gattung Equus nach Vilstrup et al. 2013[43]
|
Innere Systematik der Gattung Equus nach Price et al. 2009[44]
|
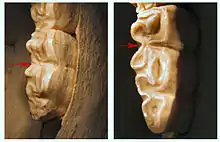
Die Anzahl der rezenten Pferdearten ist immer noch umstritten, meist werden sechs, sieben oder acht heutige Arten unterschieden. Ebenso sind die Verwandtschaftsverhältnisse zwischen den einzelnen Arten nicht restlos geklärt, so zeigen verschiedene molekulargenetische Untersuchungen teils widersprüchliche Ergebnisse.[44][43] Traditionell werden die modernen Pferde in zwei große Formengruppen geteilt: die caballine (auch caballoide) Gruppe, deren Name auf das heutige Hauspferd Equus caballus zurückgeht, und die stenonine (auch stenonide, zebroide oder non-caballine) Gruppe, benannt nach dem ausgestorbenen Equus stenonis aus dem Villafranchium. Der deutlichste Unterschied der beiden Formen besteht im Linguaflexid der unteren Molaren. Diese deutlich geschwungen verlaufende Zahnschmelzleiste an der hinteren (zungenseitigen) Zahnkante, gelegen zwischen zwei markanten Vorsprüngen (Metaconid und Metastylid), zeigt sich einerseits V-förmig (stenonin), andererseits U-förmig (caballin). Zu den stenoninen Pferden werden alle heutigen Zebras und Wildesel gestellt, während die caballinen die Wildpferde sowie das heutige Hauspferd einschließen und auch als „echte Pferde“ bezeichnet werden.[45][46]
Im Folgenden sind die allgemein akzeptierten, heutigen Arten aufgeführt. Sie verteilen sich auf drei Untergattungen:[47][4]
- Untergattung Equus Linnaeus, 1758
- Tarpan (Equus ferus Boddaert, 1785); im 19. Jahrhundert ausgestorben;
- Przewalski-Pferd (Equus przewalskii Poljakov, 1881); Bestand stark gefährdet;
- Untergattung Asinus Gray, 1822
- Afrikanischer Esel oder Wildesel beziehungsweise Echter Esel (Equus asinus Linnaeus, 1758); Eritrea, Äthiopien und Somalia; Stammform des Hausesels; Bestand in freier Wildbahn hochgradig gefährdet;
- Asiatischer Esel oder Halbesel beziehungsweise Pferdeesel (Equus hemionus Pallas, 1775); mehrere Unterarten (Onager, Kulan und andere) von Zentralasien über Südasien bis Ostasien; Bestand stark gefährdet;
- Kiang oder Tibet-Wildesel (Equus kiang Moorcroft, 1841); Hochland von Tibet und angrenzende Gebiete; größer und „pferdeähnlicher“ als andere Esel; Bestand nicht gefährdet;
- Untergattung Hippotigris H. Smith, 1842
- Grevyzebra (Equus grevyi Oustalet, 1882); Kenia, Somalia und Äthiopien; besonders enges Streifenmuster; Bestand stark gefährdet;
- Steppenzebra (Equus quagga Boddaert, 1785) mehrere Unterarten, darunter auch das im 19. Jahrhundert ausgestorbene Quagga, vom südlichen Sudan bis Südafrika; Bauch gestreift und zwischen den Streifen oft hellere „Schattenstreifen“; Bestand nicht gefährdet;
- Bergzebra (Equus zebra Linnaeus, 1758); zwei Unterarten in Namibia und Südafrika; kleinsten Zebraart mit Querstreifen an der Kruppe bis zur Schwanzwurzel; Bestand gefährdet;

Neben diesen allgemein anerkannten Arten hob eine Revision der Huftiere von Colin Peter Groves und Peter Grubb aus dem Jahr 2011 auch das Hartmann-Bergzebra (Equus hartmannae), den Khur (Equus khur) und den Syrischen Halbesel (Equus hemippus) in einen eigenständigen Artstatus.[48] Das Przewalski-Pferd galt lange Zeit als Unterart des Wildpferdes und wurde als Stammform des Hauspferdes angesehen, neuere genetische Untersuchungen interpretieren den Pferdevertreter aber als wieder verwilderte domestizierte Form.[49] Es ist in den 1960er Jahren in freier Wildbahn ausgestorben, mittlerweile laufen Auswilderungsversuche in der Mongolei, in China und in anderen Ländern. Der Bestand wird auf 2000 Tiere geschätzt. Ebenso wie Unstimmigkeiten bei der Anzahl der Arten bestehen, ist auch die Verteilung auf die Untergattungen in Diskussion. Relativ eindeutig werden das Hauspferd, der Tarpan und das Przewalski-Pferd als caballine Pferde der Untergattung Equus zugewiesen. Der Afrikanische und die asiatischen Esel (Asiatischer Esel, Kiang, Khur und Syrischer Halbesel) stehen manchmal in der Untergattung Asinus, mitunter werden die asiatischen Esel auch zur Untergattung Hemionus gezählt. Ähnliches gilt für die verschiedenen Zebraformen, für die einerseits die gemeinsame Untergattung Hippotigris existiert, andererseits wird das Grevyzebra auch in der Untergattung Dolichohippus geführt.[47][48]
Ursprünglich enthielt die Gattung Equus neben den rezenten Arten noch mehr als 230 beschriebene, nur fossil bekannte Taxa, davon allein 58 aus Nordamerika. Ein Großteil dieser ausgestorbenen Formen basierte allerdings auf nur fragmentiertem Fossilmaterial oder war mangelhaft beschrieben. Aus diesem Grund wurden 1985 bzw. 1989 zahlreiche Taxa synonymisiert.[50][30]
Stammesgeschichte
Ursprung
Die Familie der Pferde ist eine sehr alte Gruppe der Unpaarhufer, deren Stammesgeschichte rund 56 Millionen Jahre zurückreicht. Die Gattung Equus umfasst dabei die modernen Pferde und stellt das jüngste Glied in dieser Entwicklung dar. Sie bettet sich in eine Gruppe weiterer monodactyler Pferde ein, zu der unter anderem Pliohippus, Dinohippus, Astrohippus, Haringtonhippus und Hippidion gehören. Der Ursprung der modernen Pferde liegt in Nordamerika. Laut molekulargenetischen Analysen spaltete sich Equus vor rund 4,5 bis 4,0 Millionen Jahren im Pliozän von der Linie der anderen Pferde ab. Die genetisch nächsten Verwandten, Hippidion und Haringtonhippus, hatten sich bereits vor 5,2 bis 7,7 Millionen Jahren beziehungsweise vor 4,9 bis 5,7 Millionen Jahren abgetrennt.[51][52][9] Equus selber ging höchstwahrscheinlich aus Dinohippus hervor, wobei noch Unterscheidungsschwierigkeiten zwischen dieser Gattung und den frühesten modernen Pferden bestehen.[46] Vermutlich fand dieser Prozess im südlichen Teil Nordamerika durch Kladogenese statt.[53][54][55]
Die stenoninen Pferde

Die gewonnenen molekulargenetischen Daten zur Abspaltung der Pferde stimmen relativ gut mit den ältesten paläontologischen Nachweisen überein, die unter anderem aus der Ringold-Formation im US-Bundesstaat Washington stammen und älter als 3,4 Millionen Jahre sind. Diese werden der Art Equus simplicidens zugewiesen, einem frühen Vertreter der stenoninen Pferdegruppe, ähnlich wie die Funde aus der Hagerman-Fauna der Horse Quarry in Idaho, wo mehr als 150 Schädel dieser Pferdeart gefunden wurden, deren Alter auf etwa 3,7 Millionen Jahren geschätzt wird. Dieser früheste Vertreter von Equus wird manchmal auch der Untergattung Plesippus zugeordnet.[56][57] Weitere bedeutende Funde von Equus simplicidens sind unter anderem aus dem nördlichen und zentralen Mexiko überliefert, so etwa aus Jalisco.[54][55] Im späteren Verlauf des Pliozäns trat das relativ schlanke und feingliedrige Equus cumminsi auf, welches auf einzelnen Funden aus Texas basiert und häufig als den Wildeseln ähnlich bezeichnet wird.[58][59]

Etwa vor 3 bis 2,5 Millionen Jahren im ausgehenden Pliozän erreichten die stenoninen Pferde Eurasien und breiteten sich in den ursprünglich von Hipparion-Vertretern besiedelten Landschaften relativ schnell aus. Es waren damit die ersten einhufigen Pferde, die eurasischen Boden betraten.[60] Sehr frühe Funde aus dem Linxia-Becken in Ostasien, die um 2,6 Millionen Jahre datieren, werden zu Equus eisenmannae gestellt. Andere, nahezu zeitgleich auftretende stenonine Formen in Ostasien sind Equus huanghoensis, Equus qingyangensis und Equus yunnanensis. Da teilweise deutliche morphologische Unterschiede zwischen diesen Pferdeangehörigen bestehen, gehen einige Wissenschaftler auch von mehreren Einwanderungswellen aus.[61] Ähnlich alt ist Equus livenzovensis aus Montopoli im südlichen Europa.[62] Diese verschiedenen ursprünglichen Vertreter bilden möglicherweise die Basis für die nachfolgende Radiation der stenoninen Pferde, aus der bekannte Formen wie Equus stenonis, Equus sanmeniensis, Equus sivalensis und Equus suessenbornensis hervorgingen. Die Tiere wiesen häufig Merkmale der heutigen Zebras und Wildesel auf, weshalb sie ursprünglich zum Taxon Allohippus zusammengefasst wurden. Die Gattung ist zumeist nicht anerkannt, einige Wissenschaftler befürworten aber aufgrund der morphologischen Besonderheiten ihre Eigenständigkeit.[57] In Eurasien zeigen die stenoninen Pferde verschiedenste Verwandtschaftsverhältnisse untereinander. Im Allgemeinen lassen sich morphologisch ein besonders großwüchsiger Formenkreis um Equus suessenbornensis und ein eher variabler um Equus stenonis herausarbeiten.[63][64] Bemerkenswert ist, dass an vielen eurasischen Fundstellen stenoniner Pferde zwei sympatrische Arten auftreten, die sich hinsichtlich der Körpergröße unterscheiden. So kommt zum Beispiel an der eponymen mittelpleistozänen Fundstelle in Süßenborn, aber auch in Voigtstedt (beide Thüringen) neben dem großen, bis zu 590 kg schweren Equus suessenbornensis auch das kleinere Equus altidens vor, das nur rund 260 kg auf die Waage brachte.[63][65][46][47] Das gemeinsame Auftreten verschiedener stenoniner Pferde an einer Fundstelle ist höchstwahrscheinlich mit einer stärkeren Nischennutzung in Verbindung zu bringen.[66] Sowohl für Equus suessenbornensis als auch für Equus altidens wird aufgrund einiger besonderer Zahneigenschaften teilweise auch ein Verweis zur Untergattung Sussemionus vorgenommen, deren Charakterform Equus coliemensis vom Fluss Kolyma in Jakutien darstellt.[67][68] Der Großteil der stenoninen Pferde Eurasiens verschwand im Verlauf des Mittelpleistozäns wieder. Zu den wenigen Nachkommen zählt Equus ovodovi, welches anhand einiger Funde aus der Proskuriakova-Höhle im südwestlichen Sibirien beschrieben wurde. Die Fossilreste, die typische Merkmale der Untergattung Sussemionus aufzeigen, gehören mit einem Alter von 40.000 Jahren dem Jungpleistozän an.[69] Zusätzliche Funde der Art kamen im nordöstlichen China zu Tage.[70] Eine weitere späte Form ist Equus hydruntinus, der Europäische Wildesel, der im späten Mittelpleistozän und im Jungpleistozän über weite Bereiche des westlichen Eurasiens verbreitet auftrat und erst im Verlauf des Holozäns vor 5000 bis 3000 Jahren ausstarb.[71] Morphologisch zeigt er Mischmerkmale, die sowohl an den heutigen Asiatischen Esel als auch an die Angehörigen von Sussemionus erinnern. Aus genetischer Sicht steht Equus hydruntinus dem Asiatischen Esel näher und repräsentiert wohl einen ausgestorbenen Seitenzweig, während Sussemionus eher in den entfernten Verwandtschaftskreis innerhalb der Wildesel und Zebras gehört.[72][43][73][70]
Spätestens vor 2,5 bis 2 Millionen Jahren hatten die stenoninen Pferde auch Nordostafrika erreicht, wo sie mit Alterswerten um 2,3 Millionen Jahren erstmals in der Omo-Region nachgewiesen sind. Die als Equus oldowayensis benannte Form besaß etwa die Größe des heutigen Grevyzebras. Ähnlich wie in Eurasien verdrängten die Vertreter der Gattung Eqqus in Afrika frühe hipparionartige Pferde, die sich bereits im Mittleren Miozän auf dem Kontinent ausgebreitet hatten. Hier bildeten sich eigene Entwicklungslinien heraus, die Arten wie Equus koobiforensis, Equus numidicus, Equus tabeti oder Equus capensis hervorbrachten. Letzteres, unter anderem auch als „Kap-Zebra“ bekannt, ist relativ häufig im südlichen Afrika nachweisbar. Als vergleichsweise massives Tier wies es eine Schulterhöhe von rekonstruiert rund 150 cm auf, das Körpergewicht betrug schätzungsweise gut 400 kg.[74][65][59][40]
Die caballinen Pferde

Der Urahn der caballinen Pferde ist erstmals vor 2,5 bis 1,8 Millionen Jahren ebenfalls in Nordamerika fassbar und wird allgemein als Equus scotti bezeichnet. Frühe Funde dieses stämmigen, rund 550 kg schweren Pferdes sind mit mehreren Skeletten am Rock Creek in Texas geborgen worden ebenso wie in der Red-Cloud-Formation in Nebraska.[75] Neben den großen Pferden wurden für das Mittel- und Jungpleistozän Nordamerikas allgemein zwei Formengruppen kleinerer Vertreter beschrieben: einerseits eine robuste Form mit breiten Gliedmaßen, die morphologisch und genetisch den caballinen Pferden zuzurechnen ist und häufig mit Equus alaskae gleichgesetzt wird (teilweise auch mit Equus conversidens benannt, das Taxon ist aber umstritten), andererseits eine schlanke, feingliedrigere Art, die aufgrund ihrer markanten Fußanatomie im Englischen auch die Bezeichnung stilt-legged horse („stelzenbeiniges Pferd“) trägt. Letztere wurde teilweise unter dem Artnamen Equus francisci geführt. Ursprünglich für nahe Verwandte der Wildesel gehalten, blieben die genauen Beziehungen der stilt-legged horses zu anderen Pferden aber lange Zeit ungeklärt. Mehrere DNA-Untersuchungen zeigten dann, dass diese eine eigene endemische Gruppe in Amerika bilden mit einer möglicherweise engeren Bindung an die caballinen Formen.[76][77][72][43] Im Jahr 2017 wurden die stilt-legged horses basierend auf weiteren genetischen Untersuchungen zur eigenständigen Gattung Haringtonhippus verwiesen,[52] andere Autoren sehen diese Gattung aber als synonym zu Equus an.[57] Im Jungpleistozän sind verschiedene Pferdearten belegt. Relativ bedeutend ist das große Equus occidentalis, welches unter anderem in den Asphaltgruben von Rancho la Brea und in der Diamond Valley Lake Local Fauna, beide Kalifornien, recht zahlreich auftritt.[58] Deutlich kleiner hingegen waren Equus mexicanus und Equus cedralensis, die beide von Fundstellen im zentralen Mexiko beschrieben wurden.[78][59] Den Hohen Norden wiederum besiedelte das gleichfalls kleine und breitfüßige Equus lambei, von dem unter anderem ein teils mumifizierter Kadaver im Last Chance Creek bei Dawson City im kanadischen Territorium Yukon gefunden wurde.[79]
Mit der Entstehung der Landbrücke von Panama drangen die Pferde auch nach Südamerika vor. Zu den ersten Pferdevertretern dieses Kontinents zählte die Gattung Hippidion, die vor etwa 2,5 Millionen Jahren aus Nordamerika einwanderte. Vor etwa 2 Millionen Jahren erreichte dann auch Equus Südamerika, wo sich die Gattung bald darauf weit verbreitete. Einst mit wenigstens fünf Arten als recht formenreich in Südamerika angesehen, ist heute weitgehend nur Equus neogeus als valide Form anerkannt, die aber beträchtliche Größenvariationen aufzeigte.[80] Auf dem gesamten amerikanischen Kontinent starben die Pferde (sowohl Equus als auch Hippidion und Haringtonhippus) aus nicht genau bekannten Ursachen im Übergang vom Pleistozän zum Holozän vor etwa 10.000 Jahren aus.[81][77][59]
Das erste Erscheinen caballiner Pferde in Eurasien ist nicht gesichert, sehr alte Funde aus dem Villafranchium sind aus Berești (Moldawien) bekannt. Der älteste eindeutige Vertreter der caballinen Pferde in Eurasien wird in der Regel mit Equus mosbachensis in Verbindung gebracht. Es ist erstmals im frühen Mittelpleistozän in nennenswerter Fundanzahl belegt. Fossilreste sind dabei nicht nur von den namensgebenden Mosbacher Sanden in Hessen überliefert, sie verteilen sich über weite Teile Europas wie etwa Fontana Ranuccio in Italien[82] oder die Höhle von Arago in Frankreich.[83] In der Folgezeit ersetzten die caballinen Pferde die stenoninen in Eurasien weitgehend. Eine mögliche Erklärung dafür ist eine größere ökologische Bandbreite ersterer gegenüber letzteren. Bemerkenswert in diesem Zusammenhang ist auch, dass im Unterschied zu den stenoninen Pferden die caballinen sehr selten sympatrisch an einzelnen Fundstellen auftreten.[60] Vergleichbar den stenoninen Pferden spalteten sich auch die caballinen Formen in Eurasien in eine variantenreiche Gruppe mit zahlreichen Vertretern auf, bekannte Arten sind unter anderem Equus steinheimensis, Equus taubachensis und Equus chorsaricus. Die jungpleistozänen caballinen Pferde Mitteleuropas werden überwiegend als Equus germanicus bezeichnet, andere späte Vertreter sind etwa Equus latipes in Osteuropa oder Equus lenensis in Nordasien. Von letzterem blieben einige Eismumien überliefert, unter anderem das Jukagir-Pferd vom Fluss Kondratieva und das Selerikan-Pferd vom Indigirka, beide im nördlichen Jakutien.[84][85][86] Auffallend ist dabei eine merkliche Größenreduktion, die im ausgehenden Mittelpleistozän einsetzte. Das äußerst kräftige Equus mosbachensis erreichte so anfänglich noch ein Gewicht von 610 bis 740 kg bei einer Schulterhöhe von rund 165 cm, das jüngere Equus steinheimensis wog dem gegenüber rund 470 kg. Die im Vergleich zu den heutigen Arten damals dennoch deutlich größeren Ausmaße werden unter anderem mit einer höheren Wachstumsrate verursacht durch das umfangreichere Nahrungsangebot in den warmklimatischen Abschnitten des Pleistozäns erklärt.[87][88] Im folgenden Jungpleistozän bis hin zum Holozän setzte sich die Größenreduktion dann weiter fort. Für einen größeren Teil der jungpleistozänen Pferde werden Körperhöhen von 137 bis 145 cm rekonstruiert.[89] Allerdings ist für den Zeitraum nach der Maximalvereisung der letzten Kaltzeit (etwa ab 20.000 Jahre vor heute) mit einer starken Fragmentierung der Pferdepopulation zu rechnen, was unter anderem zu deutlich variierenden Größenformen abhängig von der geographischen Verbreitung führte. Es ist dabei in Diskussion, ob die Größenvariationen der caballinen Pferde des Mittel- und Jungpleistozäns jeweils eine eigenständige Artbildung ausdrücken. Mitunter sind zusätzlich unterschiedliche Skelettproportionen fassbar, erkennbar unter anderem an Tieren mit kräftigen oder schlanken Beinen sowie schmaleren oder breiteren Schnauzen. Einige Autoren vermuten in dieser deutlichen Plastizität des Pferdekörpers Anpassungen an entsprechende warm- oder kaltklimatische Bedingungen und damit an mehr geschlossene oder weitgehend offene Landschaften. Dieser Ansicht nach könnte es sich bei den unterschiedlichen Pferdformen Eurasiens eher um „Ökomorphotypen“ denn um eigenständige Arten handeln.[66][90][91][46][47]
In Afrika konnten sich die caballinen Pferde dagegen nie wirklich weit ausbreiten. Einzelne Funde sind aus dem Jungpleistozän von Allobroges in Algerien belegt und werden zu Equus algericus gestellt.[92][74]
Zur Herkunft der heutigen Pferde


Der stammesgeschichtliche Ursprung der heute noch lebenden Pferdearten ist aufgrund der vielfachen Artbenennungen pleistozäner Equus-Vertreter schwierig. In Eurasien bestanden im Jungpleistozän und Frühholozän verschiedene Linien an Wildpferden der caballinen Linie. Diese werden in westlichen Bereich zumeist mit Equus germanicus oder Equus latipes in Verbindung gebracht. Im nordasiatischen Raum sind sie meist unter der Bezeichnung Equus lenensis bekannt. Die bis in die Neuzeit überlebenden Formen sind aber höchstwahrscheinlich nicht als echte Wildpferde anzusehen. Das Przewalski-Pferd lässt sich genetischen Untersuchungen nach möglicherweise auf die Domestizierungsversuche der dem späten Neolithikum angehörenden Botai-Kultur Zentralasiens zurückführen. Genetisch könnte eine verwandtschaftliche Beziehung zu Equus lenensis bestehen.[49][93] Die Vorfahren des heutigen Hauspferdes und des Przewalski-Pferdes hatten sich bereits im Übergang vom Mittel- zum Jungpleistozän vor rund 117.000 Jahren getrennt.[94] Der Tarpan wiederum wird aus genetischer Sicht als eine Mischung aus westeurasischen Wildpferden und Hauspferden betrachtet.[95]
Über die Vorfahren der Wildesel und Zebras herrscht noch weitgehend Unklarheit. Molekulargenetisch liegt die Trennung zwischen den caballinen und stenoninen Pferden rund 3,7 bis 4,4 Millionen Jahre zurück. Die Wildesel und Zebras spalteten sich vor 1,7 bis 2 Millionen Jahren auf. Eine Diversifizierung der Zebras in das Steppen- (Equus quagga), Berg- (Equus zebra) und Grevyzebras (Equus grevyi) begann vor 1,6 Millionen Jahren zuerst mit dem Bergzebra. Die anderen Arten folgten in einem Zeitraum von rund 200.000 Jahren. Die afrikanischen und asiatischen Esel trennten sich zu einem annähernd ähnlichen Zeitpunkt vor 1,5 bis 1,8 Millionen Jahren. Zuletzt bildete sich der Kiang heraus, was wohl im Mittelpleistozän erfolgte.[51][43][96][52] Die angegebenen Daten sind deutlich jünger als ursprünglich vermutet, da eine Trennung der Wildesel und Zebras vor rund 2,8 Millionen Jahren angenommen wurde, wobei die Wildesellinie noch bis vor rund 3 Millionen Jahren zurückreichte.[45] Inwieweit Verwandtschaften zu einzelnen ausgestorbenen Formen bestehen, etwa der Wildesel zum sehr frühen nordamerikanischen Equus cumminsi wie früher häufig postuliert, bleibt unklar. Nach phylogenetischen Studien aus dem Jahr 2019 könnten sich die Zebras allerdings von Equus simplicidens Nordamerikas und den sehr frühen stenoninen Pferden Eurasiens herleiten. Dafür sprechen Übereinstimmungen im Bau der Zähne und in der vergleichenden Dimension der Mittelhand- und Mittelfußknochen.[40][97][98][99][100]
Taxonomiegeschichte, Nomenklatur und Etymologie

Die Gattung Equus wurde im Jahr 1758 von Linnaeus im Rahmen seines bedeutenden Werkes Systema Naturae wissenschaftlich benannt. Linnaeus definierte die Gattung nach ihren Zähnen und den Gliedmaßen mit dem jeweils nur einen Zeh. Die Bezeichnung Equus, die lateinischen Ursprungs ist und „Pferd“ bedeutet, wurde jedoch schon vorher verwendet. Sie findet sich unter anderem in John Rays Übersichtsarbeit zu den vierfüßigen und schlangenartigen Tieren aus dem Jahr 1693,[101] auf die sich Linnaeus in seinem Systema Naturae auch berief. Insgesamt unterschied Linnaeus drei Arten an Pferden: Equus caballus (Hauspferd), Equus asinus (Hausesel) und Equus zebra (Zebra).[102] Seine Untergliederung der Gattung wurde im Übergang vom 18. zum 19. Jahrhundert von den verschiedensten Naturforschern und Wissenschaftlern übernommen.[103][104] John Edward Gray propagiert dann 1824 in einer umfangreichen Revision der Pferde eine Gliederung in zwei Gattungen und trennte Asinus von Equus ab. In erstere schloss er auch die Zebras ein. Die Aufteilung in zwei Gattungen begründete Gray mit der Streifenbildung bei den Wildeseln und Zebras und der unterschiedlichen Verteilung der chestnuts an den Beinen.[105] Etwa anderthalb Jahrzehnte später, im Jahr 1841, hob Charles Hamilton Smith eine „asinine Gruppe“ mit den Wildeseln von einer „hippotigrinen Gruppe“ mit den Zebras ab. Er unterschied damit die Wildesel und Zebras auf Gattungsebene und vereinte letztere in dem neu geschaffenen Taxon Hippotigris.[106] Zwischenzeitlich waren bereits einige neue Arten beschrieben worden, allen voran der Asiatische Esel 1775, das Steppenzebra 1785 (als Quagga) und 1824 (als Burchell-Zebra) sowie der Tarpan ebenfalls 1785. Im weiteren Verlauf des 19. und im frühen 20. Jahrhundert wurden zusätzliche Gattungen kreiert. Von nennenswerter Wichtigkeit ist hier Dolichohippus für das Grevyzebra von Edmund Heller im Jahr 1912.[107] Bereits 1823 hatte Frédéric Cuvier die Bezeichnung Hemionus als höhere taxonomische Einheit genutzt, sie wird aber als Nomen nudum angesehen. Daher gilt Wilhelm Otto Dietrich als Erstbenenner der Gattung Hemionus, der im Jahr 1959 die asiatischen Esel unter ihr vereinte.[108] George Gaylord Simpson zweifelte dann 1945 in seiner generellen Taxonomie der Säugetiere an der generischen Eigenständigkeit der Gattungen Asinus, Hippotigris und Dolichohippus (sowie Hemionus nach F. Cuvier) und verschob sie aufgrund der deutlichen Ähnlichkeiten zu Equus auf den Rang von Untergattungen.[109] Die Ansicht wird weitgehend heute noch vertreten, die genaue Anzahl der Untergattungen – variable Angaben liegen zwischen drei und fünf – ist aber in Diskussion.[47][48][5]
Eine erste umfassende Beschreibung der Zähne der Pferde erbrachte im Jahr 1845 Richard Owen in seiner Odontography.[110] Darauf aufbauend erkannte dann 1899 Marcellin Boule unter Einbeziehung zahlreicher fossiler Formen anhand des Zahnbaus eine deutliche Zweiteilung innerhalb der Pferde mit dem Hauspferd auf der einen und den Zebras auf der anderen Seite. In dem umfangreich bebilderten Aufsatz führte Boule die Zebras auf Equus stenonis zurück.[111] Die Form war zuvor von Igino Cocchi im Jahr 1867 anhand eines Schädels aus Valdarno in der Toskana (Italien) eingeführt worden.[112] Spätere Autoren übernahmen diese Zweigliederung und arbeiteten sie detaillierter aus.[113] Vor allem Paul O. McGrew diskutierte 1944 in einem Beitrag die verschiedensten Zahnmerkmale fossiler und rezenter Pferde auf ihr Für und Wider. So kam er zu der Ansicht, dass unter anderem das pli caballin, eine enge Zahnschmelzschlaufe zwischen zwei Haupthöckern der Oberkiefermolare (Hypoconus und Protoconus) kein exklusives Merkmal des Hauspferds ist. Dagegen hob er das Linguaflexid der unteren Molaren mit dem an ein U (caballin) beziehungsweise V (stenonin) erinnernden Verlauf als unterstützend hervor.[114] Mit dem Aufkommen neuer naturwissenschaftlicher Untersuchungsmethoden im letzten Drittel des 20. Jahrhunderts fand diese bereits morphologisch ermittelte deutliche Dichotomie innerhalb der Gattung Equus auch auf genetischer und biochemischer Basis eine Bestätigung.[115] Unter anderem Ann Forstén, aber auch María T. Alberdi und andere führten daher die Teilung der Gattung in die informellen Gruppen der caballinen und stenoninen Pferde fort (nach Forstén „caballoid“ und „stenonid“), wobei Forstén auch die Wildesel in den stenoninen Formenkreis einschloss.[60][45][63]

Als Nominatform der Gattung Equus gilt Equus caballus bei Erstnennung in Linnaeus' Systema Naturae. Hierin zeichnete er mit Equus caballus das Hauspferd und mit Equus asinus den Hausesel aus. Im Jahr 1785 benannte Pieter Boddaert den Tarpan mit Equus ferus.[116] Ebenso führte Leopold Fitzinger im Jahr 1858 den Afrikanischen Esel unter der wissenschaftlichen Bezeichnung Asinus africanus.[117] Aus diesen verschiedenen Namensgebungen folgte im weiteren Verlauf eine inkonsistente Nutzung der Namen für die wilden und domestizierten Formen. Einige Forscher nutzen Equus caballus und Equus asinus deshalb auch als Artbezeichnungen für das Wildpferd und den Afrikanischen Esel (häufig dann unter Einbeziehung eines Unterartnamens für die Wildform, also E. c. ferus beziehungsweise E. a. africanus), andere verwendeten wiederum die später anhand von Wildpopulationen begründeten wissenschaftlichen Namen. Schwieriger als bei den gegenwärtig lebenden Haus- und Wildtieren wurde es allerdings bei den phylogenetischen Vorgängern oder Übergangsformen, bei denen eine sichere Zuweisung zu der einen oder anderen Gruppe nicht immer möglich ist. Generell fallen in der modernen zoologischen Systematik Haustiere nicht unter die bestehenden Namenskonventionen. Ausnahmen bilden hier aber die von Linnaeus vergebenen Artbezeichnungen, die mitunter schon seit über 200 Jahren in Gebrauch sind. Im Jahr 2003 beschloss daher die International Commission on Zoological Nomenclature auf Antrag einiger Wissenschaftler (Opinion 2027, Case 3010) die Namen Equus caballus und Equus asinus (gemeinsam mit 13 weiteren Namen domestizierter Säugetiere) zu konservieren und prinzipiell nutzbar zu machen. Wissenschaftler und Autoren können deshalb den Namen für eine wildlebende oder domestizierte Form wählen, sofern zwei Artbezeichnungen zur Verfügung stehen.[118] Die Entscheidung setzt aber wiederum nicht Opinion 271 von 1954 außer Kraft, in der die Typusart von Equus mit Equus caballus festgelegt wurde.[119] Auch steht sie nicht über der Prioritätsregel des International Code for Zoological Nomenclature, nach der der zuerst vergebene Artname auch der rechtmäßige ist. Demnach wäre bei Betrachtung des Wild- und des Hauspferdes als einzige Art ersteres letzterem beizuordnen und nicht umgekehrt. Gleiches kann zum Afrikanischen Esel und zum Hausesel gesagt werden.[120][121] Die Prioritätsregel greift in diesem Fall auch beim Steppenzebra, das 1824 von John Edward Gray mit Asinus burchelli wissenschaftlich etabliert worden war.[105] Fast 40 Jahre zuvor hatte aber Pieter Boddaert mit Equus quagga das ausgestorbene Quagga eingeführt.[116] Beide Arten galten lange Zeit als eigenständig. Jedoch zeigten genetische Untersuchungen eine enge Verwandtschaft auf.[122] Aufgrund der daraufhin erfolgten Synonymisierung beider Arten ist Equus quagga heute der korrekte Artname des Steppenzebras.[48][48][5]
Der deutsche Name „Pferd“ leitet sich von der mittellateinischen Bezeichnung paraveredus für ein Kurierpferd auf Nebenstrecken ab. Diese wiederum basiert auf dem keltisch-spätlateinischen Wort veredus für „Kurierpferd“ und der griechischen Vorsilbe παρά (pará) für „neben“ oder „bei“. Das Wort „Esel“ wurde über das althochdeutsche esil aus dem lateinischen asinus (oder asellus als Verkleinerungsform) vermittelt. Die Herkunft wird aus einer kleinasiatischen Sprache vermutet.[123] Der Ursprung des Wortes „Zebra“ ist unklar. Möglicherweise findet er sich im Wort zecora der Oromo-Sprache des nordöstlichen Afrikas. Erstmals benannten Portugiesen im 15. Jahrhundert gestreifte Tiere des zentralen Afrikas mit zebra. Später wurde dies auch auf ähnliche Tiere des südlichen Afrikas übertragen.[106][3]
Pferde und Menschen
Urgeschichte
Die Bedeutung der Pferde für den Menschen reicht bis in die Altsteinzeit zurück. Genutzt wurden die Tiere überwiegend als Rohstoff- und Nahrungsressource. Reste von Pferden finden sich an zahlreichen Fundstellen vom Alt- bis zum Jungpaläolithikum allein in Europa. Exemplarisch seien hier nur Miesenheim im Neuwieder Becken[124] oder Ehringsdorf bei Weimar in Thüringen[125] genannt. Auch an den mittel- bis jungpleistozänen Fundstellen des Geiseltals sind Pferde relativ häufig belegt.[126] Während sich die Nutzung der Tiere durch Schnittspuren und Schlagmarken an den Knochen vergleichsweise sicher feststellen lässt, sind Nachweise der direkten Jagd weitaus seltener in dieser Zeit zu finden. Einer der eindrucksvollsten stammt aus Schöningen in Niedersachsen, wo auf einer Fläche von rund 1200 m² neben acht flugtauglichen hölzernen, bis zu 250 cm langen Speeren auch unzählige Reste von Pferden entdeckt wurden. Fast alle dieser Pferdereste gehören Analysen zufolge zu Equus mosbachensis, untergeordnet auch zu Equus hydruntinus. Die Funde datieren in das späte Mittelpleistozän und dürften zwischen 300.000 und 400.000 Jahre alt sein.[127][128][129] Mit einem Alter von rund 50.000 Jahren bereits deutlich jünger ist ein Skelett eines Afrikanischen Esels, das in Umm el Tlel in Syrien aufgefunden wurde und in dessen Halswirbelsäule eine gesplitterte Levalloisspitze als Relikt eines ehemaligen Jagdereignisses steckte.[130]

Im Jungpaläolithikum zeigt sich die herausragende Stellung der Pferde vor allem in der Kleinkunst und in der Höhlenmalerei, die vor rund 35.000 bis 40.000 Jahren aufkamen. Allein in der Höhlenkunst des Frankokantabrischen Raumes sind wenigstens knapp zwei Dutzend Fundstellen mit Darstellungen von Pferden bekannt. Es kommen sowohl farbige Zeichnungen als auch Gravuren, Ritzungen und Reliefs vor. Die mitunter sehr individuell gestalteten Porträts lassen aber nicht nur frühe Vorfahren des Wildpferds erkennen, es wurden – wenn auch seltener – verschiedene Wildeselformen dargestellt, wobei die Interpretationen vom „Europäischen Wildesel“ (Equus hydruntinus) bis zum Asiatischen Esel reichen. Pferde repräsentieren die insgesamt am häufigsten abgebildeten Tiere. Auf sie entfallen rund 27 % aller Tierdarstellungen, womit sie noch vor den Hornträgern und den Hirschen rangieren. Die Häufigkeit von Pferden ist aber von Höhle zu Höhle recht unterschiedlich. Zu den ältesten Höhlenkunstwerken gehören jene der Grotte Chauvet mit rund 40 Wildpferd-Bildnissen aus der Zeit vor rund 32.000 bis 26.000 Jahren. In Lascaux wiederum wurden über 360 Darstellungen von Pferden gezählt, sie stellen damit rund 60 % aller Tierabbildungen. Mit etwa 17.500 Jahren sind sie nur etwa halb so alt wie die der Grotte Chauvet. Neben den Pferdedarstellungen der Frankokantabrischen Höhlenkunst sind auch einzelne Abbildungen aus der Kapova-Höhle im Ural beschrieben worden.[131][132] Einen ähnlich umfangreichen Anteil haben Pferde in der jungpaläolithischen Kleinkunst. Hier lassen sich Ritzungen in Stein oder Knochen beziehungsweise voll- und halbplastische Figuren unterscheiden. Einen besonderen Stellenwert nimmt beispielsweise die nur 4,8 cm lange Pferdefigur mit geschwungenem Hals aus der Vogelherdhöhle im Lonetal in der Schwäbischen Alb ein, da sie gemeinsam mit anderen Tierfiguren mit einem Alter von rund 35.000 Jahren zu den ältesten Kunstobjekten der Welt gehört.[133][134]
Domestizierung
Hausesellinie nach Kimura et al. 2011[135]
|

In ihrer domestizierten Form als Hauspferd und Hausesel leisteten die Pferde einen bedeutenden Beitrag in der Geschichte und Kulturentwicklung der Menschheit. Sie hatten dabei überwiegend die Funktion als Reit-, Arbeits- und Lasttiere inne. Der Zeitpunkt der Domestikation beider Arten ist in Diskussion und wird mit verschiedenen Ansätzen untersucht. Schätzungen zufolge geschah dies beim Hausesel rund 3000 v. Chr. im Alten Ägypten. Zu den ältesten eindeutigen Belegen aus dieser Zeit gehören einige gut erhaltene Skelettfunde aus Abydos. Nach anatomischen Untersuchungen durch ein Arbeitsteam um Stine Rossel und Fiona B. Marshall aus dem Jahr 2008 wurden die Tiere vorwiegend als Lastträger eingesetzt.[136] Möglicherweise nur wenig später erfuhren sie auch eine Verwendung als Zug- oder Reittier, was anhand von Eselresten aus Tell eṣ-Ṣâfi in Israel geschlussfolgert wird. Diese datieren in den Zeitraum um 2800 bis 2600 v. Chr., an den Zähnen finden sich charakteristische Abnutzungsspuren, die bei der Verwendung von Trensen entstehen.[137] Es gibt aber auch Hinweise, dass der Hausesel schon in prädynastischer Zeit um 4000 oder 4500 v. Chr. auftrat. Dafür sprechen einzelne Funde kleiner Tiere aus Tell el-Iswid beziehungsweise aus El Omari in Unter- und aus Nagada in Mittelägypten.[138][139][140] Genetischen Befunden zufolge ist der Afrikanische Esel die Wildform des Hausesels. In ersten Analysen aus dem Jahr 2004 ließ sich die nubische Unterart (Nubischer Wildesel) als wahrscheinliche Ausgangsform identifizieren. Allerdings wurde der Hausesel möglicherweise mehrfach domestiziert, wie diese frühen Studien bereits aufzeigten. In späteren Untersuchungen konnten wenigstens zwei Kladen des Hausesels herausgearbeitet werden, die auf jeweils eigenständige Domestikationsprozesse zurückgehen. Clade 1 repräsentiert weitgehend den heutigen Hausesel und hat ihren Ursprung wohl im nördlichen Afrika. In ihrer mitochondrialen DNA ist sie nicht vom Nubischen Wildesel zu unterscheiden. Dagegen steht Clade 2 dem Somali-Wildesel näher, ist aber nicht mit dieser Unterart identisch. Sie stellt die zweite Domestikationsform dar, basiert im Unterschied zu Clade 1 aber auf einer kleineren Ausgangsgruppe. Aufgrund ihrer Besonderheit konnten die Stammform und der Ursprungsort von Clade 2 bisher nicht genauer identifiziert werden.[141][135] Regional wie etwa in Mesopotamien wurde der frühe Hausesel teilweise auch mit dem Asiatischen Esel gekreuzt.[142]
|
.jpg.webp)
Domestizierte Pferde traten etwa im gleichen Zeitraum auf. Hauptsächlich in Mittel- und Westasien basierten einige der lokal bestehenden archäologische Kulturen des Spätneolithikums und der Frühbronzezeit weitgehend auf der Nutzung der Pferde als Rohstoffquelle, nicht nur zu Nahrungszwecken, sondern auch für die Werkzeugherstellung. Die Tiere fanden darüber hinaus auch Einzug in rituelle Handlungen, ihre große Bedeutung spiegelt sich unter anderem in zahlreichen Kunstgegenständen mit Pferdemotiven wider. Zu den bekanntesten Kulturgruppen gehören die Chwalynsk-Kultur in Russland, die Ockergrabkultur in der Ukraine und die Botai-Kultur in Kasachstan. Diskutiert wurde lange, ob die genutzten Pferde Wildfänge oder domestizierte Tiere repräsentierten. Zumindest die Pferde der sich um 3500 v. Chr. herausformenden Botai-Kultur weisen nach einigen Wissenschaftlern die für Trensen typischen Abnutzungsspuren an den Prämolaren auf. Demnach könnte es sich um gezähmte Tiere gehandelt haben, die neben der Verwendung als Nahrungsgrundlage einigen Überlegungen zufolge auch zum Reiten genutzt wurden, was möglicherweise die Mobilität der Steppenvölker erhöhte.[143][144][145][146] Der Ansicht wird zwar teilweise widersprochen,[147] eine Studie aus dem Jahr 2018 an Pferderesten aus der Botai-Kultur ergab aber, dass diese eventuell frühen domestizierten Pferde nicht in die Linie der heutigen Hauspferde gehören. Stattdessen bilden sie die Basis des Przewalski-Pferdes, das lange Zeit als ursprüngliche Wildform galt.[49] Das heutige Hauspferd muss demnach erneut domestiziert worden sein. Unterstützung findet diese Ansicht auch durch die lange genetische Trennung zwischen Przewalski-Pferd und Hauspferd, die bis in die Eem-Warmzeit zurückreicht.[94] Der Ursprungsort des heutigen Hauspferdes war lange Zeit ungeklärt. Als wahrscheinliche Region erwies sich nach genetischen Analysen aus dem Jahr 2021 das Schwarzmeergebiet. Demnach gehen die heutigen domestizierten Pferde alle auf eine Ursprungsgruppe zurück, die um rund 3000 v. Chr. entstand, eventuell im Zusammenhang stehend mit den spätneolithischen Komplexen der Maikop- oder Jamnaja-Kultur. Als Ausgangsgruppe kommen verschiedene Wildpferde in Betracht, im fraglichen Zeitraum lebten einzelne Populationen in Eurasien, darunter eine in Sibirien und eine auf der Iberischen Halbinsel, die beide aber nur wenig zum Genpool des Hauspferdes beitrugen.[93][95] Einige Wissenschaftler gehen bei der Domestikation von einem dynamischen Prozess aus, bei dem immer wieder regional Wildformen eingekreuzt wurden.[148][149] DNA-Analysen an Funden pleistozäner und frühholozäner Wildpferde und an Hauspferden des Neolithikums sowie der Bronze- und Eisenzeit erbrachten eine relativ hohe Farbvielfalt, die sich vermutlich erst in der Domestikation und Zucht herausgebildet hat.[150][151] Zu einem ähnlichen Ergebnis kommt eine weitere Studie aus dem Jahr 2019. Diese weist aber darauf hin, dass bei heutigen Hauspferden ein bedeutender Einfluss persischer Tiere besteht, der sich erst im letzten Jahrtausend durch die teils starke islamische Prägung einiger Regionen Europas herausgebildet habe. Außerdem führten moderne Zuchtpraktiken zum Rückgang der Diversität bei den Hauspferden.[93] Mitunter geschah dies aber auch schon in fernerer Vergangenheit, wie dies am Beispiel der „leopardfleckigen“ Pferde (hauptsächlich weiße Tiere mit schwarzen Flecken, Typ Tobiano) gezeigt werden konnte. Diese sind genetisch seit dem ausgehenden Pleistozän bekannt und fanden auch Einzug in den Genpool früher Hauspferdpopulationen seit dem mittleren Neolithikum. In der Folgezeit verschwanden sie aber mehrfach und wurden offensichtlich wieder erneut eingeführt. Ein Grund für das mehrfache Wegzüchten dieses Merkmals könnte darin liegen, dass der Nachwuchs unter Umständen nachtblind ist und dadurch eventuell leichter Prädatoren zum Opfer fällt.[152][151]

Vergleichbar dem Hausesel wurden die ersten Hauspferde im westlichen Teil Eurasiens wohl zunächst als Trag- und Zugtiere eingesetzt. Seit der Bronzezeit fanden sie dann auch zunehmend als Reittiere Verwendung, wie dies beispielsweise einige Felsbilder in Schweden, unter anderem das Reiterfelsbild von Tegneby, annehmen lassen.[153] Wann das Hauspferd im westlichen Eurasien erschien, ist nicht eindeutig. Allgemein geht man von der frühen Bronzezeit aus, worauf unter anderem Funde von Trensen hindeuten. Andererseits führen einzelne Befunde aus dem mittleren Neolithikum zu der Vermutung, dass Hauspferde möglicherweise hier schon weitaus früher genutzt wurden. Dazu gehört etwa ein Pferdeschädel, der im trichterbecherzeitlichen Erdwerk von Salzmünde in Sachsen-Anhalt intentionell niedergelegt worden war. Er datiert auf etwa 3400 bis 3100 v. Chr. Weitere sehr frühe Hinweise auf domestizierte Pferde in Mitteleuropa wurden unter anderem aus Vyškov in Südmähren berichtet. Hier lagen in einem Grab mit menschlichem Leichenbrand aus der Zeit der Glockenbecherkultur zwei Pferdeschädel.[154]

Sowohl der Hausesel als auch das Hauspferd erlangten als Begleiter und Nutztier des Menschen eine weltweite Verbreitung. Sie erreichten dadurch Gebiete, in denen Wildpferde zuvor ausgestorben waren (Amerika) oder die sie nie besiedelt hatten (Australien und zahlreiche abgelegene Inseln). So stammen etwa die wildlebenden Pferde Amerikas weitgehend von europäischen Züchtungen. Die ersten modernen Pferde kamen im Gefolge von Christoph Kolumbus im Jahr 1492 nach Amerika. Die Tiere stammten aus der Provinz Sevilla, vornehmlich aus den Salzmarschen des Flusses Guadalquivir. Historischen Berichten zufolge war bereits 1503 eine Gruppe von 70 Pferden auf Hispaniola stationiert. Im Zuge der Kolonialisierung Amerikas führten die Spanier iberische Pferde in zahlreichen Regionen ein. Im Jahr 1553 lebten dann schon rund 10.000 verwilderte Exemplare allein im mexikanischen Bundesstaat Querétaro. Die Abstammung der ersten Hauspferde in Amerika lässt sich auch genetisch belegen, wie dies Untersuchungen an Mustangs sowie europäischen und arabischen Zuchtlinien zeigen. Demnach weist fast ein Drittel aller untersuchten Mustangs genetische Verbindungen zu iberischen Pferden auf.[155] Auch einige amerikanische Zuchtlinien haben ihren Ursprung bei iberischen Pferden, da sich beispielsweise die nordamerikanischen Sulphur- und Spanischen Mustangs gemeinsame und teils sehr ursprüngliche Haplotypen mit den Menorquinern und Sorraias teilen. Die nordamerikanischen Rassepferde besitzen dabei generell eine geringere Variabilität als ihre europäischen Verwandten, was auf die nur kleinen Gründergruppen zurückzuführen ist. Ähnliches kann für einige südamerikanische Linien ausgesagt werden. Andererseits bilden auch die iberischen Pferde keine geschlossene Einheit, da sie dem Einfluss verschiedenster Zuchtlinien unterlagen, etwa durch nordafrikanische Pferde während der Herrschaft der Mauren.[156] Nachfolgend verbrachten die Europäer dann auch andere Zuchtlinien nach Amerika, die dort teilweise ebenfalls verwilderten.[157][158]
Aufgrund der Motorisierung der Landwirtschaft und der Verbreitung des Automobilverkehrs ist die Nutzung von Pferden und Eseln in den westlichen Industrieländern im Personen- und Güterverkehr stark zurückgegangen, das Reiten wird meist nur mehr als Hobby oder Sport betrieben.[4] In den unterentwickelten Regionen der Erde ist der Einsatz von Tieren als Verkehrsmittel aber immer noch weit verbreitet. Ein weiterer wichtiger Bereich der Nutzung ist das Pferdefleisch als Nahrungsmittel. Auch die Stuten- und Eselsmilch werden verwendet, und die Haut beider Arten wird zu Leder verarbeitet, wobei dem Pferdeleder bei der Herstellung aufwendiger Schuhe eine besondere Bedeutung zukam. Im Gegensatz zu anderen Nutztieren spielten diese Zwecke jedoch stets eine untergeordnete Rolle. Daneben gibt es für Rosshaar vielfältige Anwendungsmöglichkeiten.
Im Unterschied zu den Wildpferden und Wildeseln wurden Zebras nie domestiziert, lediglich die Zähmung einzelner Tiere fand statt. Gründe mögen in der höheren Aggressivität der Hengste liegen, die häufiger Kämpfe untereinander mit unter Umständen auch schwereren Verletzungen führen. Dies trifft vor allem für das gruppenbildende Steppenzebra zu, das sich zu größeren Gemeinschaften für längere Wanderungen im Jahresrhythmus zusammenfindet. Darüber hinaus haben Zebras eine bessere Seitensicht als die anderen Vertreter der Gattung, so dass ein Einfangen sich schwieriger gestaltet.[159][4]
Hybride
Nachkommen eines Eselhengstes und einer Pferdestute werden als Maultiere bezeichnet; der umgekehrte Fall, Nachkommen von Pferdehengst und Eselstute, als Maulesel. Kreuzungen zwischen Pferd und Zebra oder Esel und Zebra nennt man Zebroide. Die meisten Hybride der Gattung Equus werden in menschlicher Gefangenschaft geboren und teils gezielt gezüchtet, da der Nachwuchs kräftiger und mitunter widerstandsfähiger ist als die Elternformen.[4] Sie können allerdings auch in freier Wildbahn vorkommen, wenn sich die Verbreitungsgebiete zweier Pferdearten überschneiden. Eine natürliche Hybridisierung ist gegenwärtig nur beim Zebra und dem Afrikanischen Wildesel der Fall.[160] In der stammesgeschichtlichen Vergangenheit war es verschiedentlich zum Genaustausch zwischen den einzelnen Arten gekommen.[96]
Gefährdung
Die meisten Pferdearten sind gefährdet. Durch Bejagung und Einschränkung des Lebensraumes wurden viele Arten an den Rand der Ausrottung gedrängt. Das Quagga, eine Unterart des Steppenzebras, wurde Ende des 19. Jahrhunderts ausgerottet. Das Przewalski-Pferd gilt als in freier Wildbahn ausgerottet und lebt nur mehr dank Auswilderungsprojekten in der Natur. Vom wildlebenden Afrikanischen Esel gibt es nur noch wenige hundert Exemplare, die IUCN listet ihn als vom Aussterben bedroht (critically endangered). Der Asiatische Esel und das Grevyzebra gelten als stark gefährdet (endangered), das Bergzebra als gefährdet (vulnerable). Das Steppenzebra und der Kiang sind in ihrem Bestand momentan nicht gefährdet (least concern).[161] Bezüglich der beiden Haustierformen ist der Bestand des Hausesels stark gefährdet. Vor allem die Haut der Tiere findet zu Pulver oder Gelee zermahlen (ejiao) Verwendung in der traditionellen chinesischen Medizin, wofür jährlich Schätzungen zufolge bis zu 4,8 Millionen Individuen geschlachtet werden. Allein in China ging der Bestand von 11 Millionen Hauseseln im Jahr 1992 auf 2,6 bis 4,6 Millionen im Jahr 2017 zurück. Einige Experten vermuten, dass der weltweite Bestand innerhalb von fünf Jahren auf die Hälfte zusammenbrechen könnte (Stand 2019).[162]
Literatur
- Stephen Budiansky: The Nature of Horses: Exploring Equine Evolution, Intelligence, and Behavior. The Free Press, New York 1997, ISBN 0-684-82768-9.
- Jens Lorenz Franzen: Die Urpferde der Morgenröte. Elsevier, Spektrum Akademischer Verlag, München 2007, ISBN 3-8274-1680-9.
- Ronald M. Nowak: Walker’s Mammals of the World. The Johns Hopkins University Press, Baltimore 1999, ISBN 0-8018-5789-9.
- Dan I. Rubenstein: Family Equidae (Horses and relatives). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 2: Hooved Mammals. Lynx Edicions, Barcelona 2011, ISBN 978-84-96553-77-4, S. 106–143.
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. 3. Ausgabe. The Johns Hopkins University Press, Baltimore 2005, ISBN 0-8018-8221-4.
Einzelnachweise
- Colin Peter Groves und V. Mazák: On some taxonomic problems of Asiatic wild asses; with the description of a new subspecies (Perissodactyla; Equidae). Zeitschrift für Säugetierkunde 32, 1967, S. 321–355.
- Martha I. Grinder, Paul R. Krausman und Robert S. Hoffmann. Equus asinus. Mammalian Species 794, 2006, S. 1–9.
- C. S. Churcher: Equus grevyi Mammalian Species 453, 1993, S. 1–9.
- Dan I. Rubenstein: Family Equidae (Horses and relatives). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 2: Hooved Mammals. Lynx Edicions, Barcelona 2011, ISBN 978-84-96553-77-4, S. 106–143.
- Jonathan Kingdon, David Happold, Michael Hoffmann, Thomas Butynski, Meredith Happold und Jan Kalina (Hrsg.): Mammals of Africa Volume V. Carnivores, Pangolins, Equids and Rhinoceroses. Bloomsbury, London, 2013, S. 410–443 (verschiedene Autoren).
- Horst Erich König, Klaus-Dieter Budras, Johannes Seeger und Mircea-Constantin Sora: Zur topographisch-klinischen Anatomie des Luftsackes (Diverticulum tubae auditivae) beim Pferd. Pferdeheilkunde 26, 2010, S. 152–156.
- Nikos Solounias, Melinda Danowitz, Irvind Buttar und Zachary Couppee: Hypsodont crowns as additional roots: A new explanation for hypsodonty. Frontiers in Ecology and Evolution 7, 2019, S. 135, doi:10.3389/fevo.2019.00135.
- Nikos Solounias, Melinda Danowitz, Elizabeth Stachtiaris, Abhilasha Khurana, Marwan Araim, Marc Sayegh und Jessica Natale: The evolution and anatomy of the horse manus with an emphasis on digit reduction. Royal Society Open Science 5 (1), 2018, S. 171782, doi:10.1098/rsos.171782.
- Christine M. Janis und Raymond l Bernor: The Evolution of Equid Monodactyly: A Review Including a New Hypothesis. Frontiers in Ecology and Evolution 7, 2019, S. 119, doi:10.3389/fevo.2019.00119.
- Deb Bennett und Robert S. Hoffmann: Equus caballus. Mammalian Species 628, 1999, S. 1–14.
- Rikke Buhl, Annette K. Ersbøll, Lis Eriksen und Jørgen Koch: Changes over time in echocardiographic measurements in young standardbred racehorses undergoing training and racing and association with racing performance. Journal of the American Veterinary Medical Association 226 (11), 2005, S. 1881–1887.
- Meg M. Sleeper, Mary M. Durando, Todd C. Holbrook, Mark E. Payton und Eric K. Birks: Comparison of echocardiographic measurements in elite and nonelite Arabian endurance horses. American Journal of Veterinary Research 75 (10), 2014, S. 893–898.
- S. G. B. Persson: Blood Volume, State of Training and Working Capacity of Race Horses. Equine Veterinary Journal 1 (2), 1968, S. 52–62.
- Girma Gebresenbet, Samuel Aradom und Pascal Gitari Kaumbutho: Performance and Welfare Status of Working Donkeys. Journal of Agricultural Science and Technology A 6, 2016, S. 108–115, doi:10.17265/2161-6256/2016.02.004.
- Debbie A. Myers, Scott Citino und Mark A. Mitchell: Electrocardiography of Grevy's zebra (Equus grevyi). Journal of Zoo and Wildlife Medicine 39 (3), 2008, S. 298–304.
- Roderick I. MacKie und Clive A. Wilkins: Enumeration of Anaerobic Bacterial Microflora of the Equine Gastrointestinal Tract. Applied and Environmental Microbiology 54 (9), 1988, S. 2155–2160.
- H. Jerbi, A. Rejeb, S. Erdoğan und W. Pérez: Anatomical and morphometric study of gastrointestinal tract of donkey (Equus africanus asinus). Journal of Morphological Sciences 31 (1), 2014, S. 18–22.
- H. E. König: Zur Anatomie des Eierstocks der Stute. Acta Veterinaria Brno 64 (1), 1995, S. 13–16.
- T. F. P. Renner-Martin, G. Forstenpointner, G. E. Weissengruber und L. Eberhardt: Gross Anatomy of the Female Genital Organs of the Domestic Donkey (Equus asinus Linné, 1758). Anatomia, Histologia, Embryologie 38, 2009, S. 133–138.
- Peter Grubb: Equus burchelli. Mammalian Species 157, 1981, S. 1–9.
- Ramiro E. Toribio: Essentials of Equine Renal and Urinary Tract Physiology. Veterinary clinics of North America. Equine practice 23, 2007, S. 533–561.
- H. M. Mai: Gross Anatomical Study of the Urogenital System of the Indigenous Nigerian Male Donkey (Equus africanus africanus) in Comparison with the Stallion. Anatomy & Physiology 4 (3), 2014, S. 1000145, doi:10.4172/2161-0940.1000145.
- O. A. Ryder und L. G. Chemnick: Chromosomal and molecular evolution in Asiatic wild asses. Genetica 83 (1), 1990, S. 67–72.
- Natalia A. Villavicencio, Derek Corcoran und Pablo A. Marquet: Assessing the Causes Behind the Late Quaternary Extinction of Horses in South America Using Species Distribution Models. Frontiers in Ecology and Evolution 7, 2019, S. 226, doi:10.3389/fevo.2019.00226.
- James D. Feist und Dale R. McCullough: Behavior patterns and communication in feral horses. Zeitschrift für Tierpsychologie 41, 1976, S. 337–371, doi:10.1111/j.1439-0310.1976.tb00947.x.
- M. Andreina Pacheco und Emilio A. Herrera: Social Structure of Feral Horses in the Llanos of Venezuela. Journal of Mammalogy 78, 1977, S. 15–22, doi:10.2307/1382634.
- Konstanze Krüger, Birgit Flauger, Kate Farmer und Charlotte Hemelrijk: Movement initiation in groups of feral horses. Behavioural Processes, 103, 2014, 91–101, doi:10.1016/j.beproc.2013.10.007 (Volltext PDF).
- Hans Klingel: Soziale Organisation und Verhaltensweisen von Hartmann- und Bergzebras (Equus zebra hartmannae und E. z. zebra). Zeitschrift für Säugetierpsychologie 25 (1), 1968, S. 76–88, doi:10.1111/j.1439-0310.1968.tb00004.x.
- L. B. Penzhorn: Equus zebra. Mammalian Species 314, 1988, S. 1–7.
- Bruce J. MacFadden: Fossil Horses. Systematics, Paleobiology, and Evolution of the Family Equidae. Cambridge University Press, 1992 (hier S. 113 und S. 264–268).
- Konstanze Krüger und Brigitte Flauger: Olfactory Recognition of Individual Competitors by Means of Faeces in Horse (Equus caballus). Animal Cognition 14, 2011, S. 245–257, doi:10.1007/s10071-010-0358-1 (Volltext, PDF).
- Matthew C. Mihlbachler, Florent Rivals, Nikos Solounias und Gina M. Semprebon: Dietary Change and Evolution of Horses in North America. Science 331, 2011, S. 1178–1181.
- Caroline A. E. Strömberg: Evolution of hypsodonty in equids: testing a hypothesis of adaptation. Paleobiology 32 (2), 2006, S. 236–258.
- Thomas M. Kaiser und Tamara A. Franz-Odendaal: A mixed-feeding Equus species from the Middle Pleistocene of South Africa. Quaternary Research 62 (3), 2004, S. 316–323.
- Ingolf Bender: Wasser – ein Treibstoff für den Körper. Reiter Revue Juni 2007, S. 69–74.
- José L. Prado und María T. Alberdi: A cladistic analysis of the horses of the tribe Equini. Journal of Palaeontology 39 (3), 1996, S. 663–680.
- Luke T. Holbrook: Comparative osteology of early Tertiary tapiromorphs (Mammalia, Perissodactyla). Zoological Journal of the Linnean Society 132, 2001, S. 1–54.
- Christelle Tougard, Thomas Delefosse, Catherine Hänni und Claudine Montgelard: Phylogenetic Relationships of the Five Extant Rhinoceros Species (Rhinocerotidae, Perissodactyla) Based on Mitochondrial Cytochrome b and 12S rRNA Genes. Molecular Phylogenetics and Evolution 19, 2001, S. 34–44.
- Cynthia C. Steiner und Oliver A. Ryder: Molecular phylogeny and evolution of the Perissodactyla. Zoological Journal of the Linnean Society 163, 2011, S. 1289–1303.
- Raymond L. Bernor, Omar Cirilli, Advait M. Jukar, Richard Potts, Maia Buskianidze und Lorenzo Rook: Evolution of Early Equus in Italy, Georgia, the Indian Subcontinent, East Africa, and the Origins of African Zebras. Frontiers in Ecology and Evolution 7, 2019, S. 166, doi:10.3389/fevo.2019.00166.
- Bruce J. MacFadden: Astrohippus and Dinohippus from the Yepomera Local Fauna (Hemphillian, Mexico) and Implications for the Phylogeny of One-Toed Horses. Journal of Vertebrate Paleontology 4 (2), 1984, S. 273–283.
- José Luis Prado und María Teresa Alberdi: Fossil Horses of South America. Phylogeny, Systemics and Ecology. Springer International Publishing, 2017, S. 73–84.
- Julia T. Vilstrup, Andaine Seguin-Orlando, Mathias Stiller, Aurelien Ginolhac, Maanasa Raghavan, Sandra C. A. Nielsen, Jacobo Weinstock, Duane Froese, Sergei K. Vasiliev, Nikolai D. Ovodov, Joel Clary, Kristofer M. Helgen, Robert C. Fleischer, Alan Cooper, Beth Shapiro und Ludovic Orlando: Mitochondrial Phylogenomics of Modern and Ancient Equids. PLoS ONE 8 (2), 2013, S. e55950.
- Samantha A. Price und Olaf R. P. Bininda-Emonds: A comprehensive phylogeny of extant horses, rhinos and tapirs (Perissodactyla) through data combination. Zoosystematics and Evolution 85 (2), 2009, S. 277–292.
- Ann Forstén: Mitochondrial-DNA time-table and the evolution of Equus: comparison of molecular and paleontological evidence. Annales Zoologici Fennici 28, 1992, S. 301–309.
- Ralf-Dietrich Kahlke: Die Entstehungs-, Entwicklungs- und Verbreitungsgeschichte des oberpleistozönen Mammuthus-Coelodonta-Faunenkomplexes in Eurasien (Großsäuger). Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft 546 Frankfurt am Main, 1994.
- Rudolf Musil: Evolutionäre Trends bei den Pferden des europäischen Quartärs. Praehistoria Thuringica 11, 2006, S. 125–138.
- Colin Groves und Peter Grubb: Ungulate Taxonomy. Johns Hopkins University Press, 2011, S. 1–317 (S. 13–17).
- Charleen Gaunitz, Antoine Fages, Kristian Hanghøj, Anders Albrechtsen, Naveed Khan, Mikkel Schubert, Andaine Seguin-Orlando, Ivy J. Owens, Sabine Felkel, Olivier Bignon-Lau, Peter de Barros Damgaard, Alissa Mittnik, Azadeh F. Mohaseb, Hossein Davoudi, Saleh Alquraishi, Ahmed H. Alfarhan, Khaled A. S. Al-Rasheid, Eric Crubézy, Norbert Benecke, Sandra Olsen, Dorcas Brown, David Anthony, Ken Massy, Vladimir Pitulko, Aleksei Kasparov, Gottfried Brem, Michael Hofreiter, Gulmira Mukhtarova, Nurbol Baimukhanov, Lembi Lõugas, Vedat Onar, Philipp W. Stockhammer, Johannes Krause, Bazartseren Boldgiv, Sainbileg Undrakhbold, Diimaajav Erdenebaatar, Sébastien Lepetz, Marjan Mashkour, Arne Ludwig, Barbara Wallner, Victor Merz, Ilja Merz, Viktor Zaibert, Eske Willerslev, Pablo Librado, Alan K. Outram und Ludovic Orlando: Ancient genomes revisit the ancestry of domestic and Przewalski’s horses. Science 360 (6384), 2018, S. 111–114, doi:10.1126/science.aao3297.
- Melissa C. Winans: A quantitative study of North American fossil species of the genus Equus. In: Donald R. Prothero und R. Schoch (Hrsg.): The evolution of Perissodactyls. New York, Oxford University Press, 1989, S. 262–297.
- Ludovic Orlando, Aurélien Ginolhac, Guojie Zhang, Duane Froese, Anders Albrechtsen, Mathias Stiller, Mikkel Schubert, Enrico Cappellini, Bent Petersen, Ida Moltke, Philip L. F. Johnson, Matteo Fumagalli, Julia T. Vilstrup, Maanasa Raghavan, Thorfinn Korneliussen, Anna-Sapfo Malaspinas, Josef Vogt, Damian Szklarczyk, Christian D. Kelstrup, Jakob Vinther, Andrei Dolocan, Jesper Stenderup, Ahmed M. V. Velazquez, James Cahill, Morten Rasmussen, Xiaoli Wang, Jiumeng Min, Grant D. Zazula, Andaine Seguin-Orlando, Cecilie Mortensen, Kim Magnussen, John F. Thompson, Jacobo Weinstock, Kristian Gregersen, Knut H. Røed, Véra Eisenmann, Carl J. Rubin, Donald C. Miller, Douglas F. Antczak, Mads F. Bertelsen, Søren Brunak, Khaled A. S. Al-Rasheid, Oliver Ryder, Leif Andersson, John Mundy, Anders Krogh, M. Thomas P. Gilbert, Kurt Kjær, Thomas Sicheritz-Ponten, Lars Juhl Jensen, Jesper V. Olsen, Michael Hofreiter, Rasmus Nielsen, Beth Shapiro, Jun Wang und Eske Willerslev: Recalibrating Equus evolution using the genome sequence of an early Middle Pleistocene horse. Nature 499, 2013, S. 74–78.
- Peter D. Heintzman, Grant D. Zazula, Ross D. E. MacPhee, Eric Scott, James A. Cahill, Brianna K. McHorse, Joshua D. Kapp, Mathias Stiller, Matthew J. Wooller, Ludovic Orlando, John Southon, Duane G. Froese und Beth Shapiro: A new genus of horse from Pleistocene North America. eLife 6, 2017, S. e29944, doi:10.7554/eLife.29944.
- Bruce J. MacFadden und Oscar Carranza-Casteñeda: Cranium of Dinohippus mexicanus (Mammalia: Equidae) from the Early Pliocene (Latest Hemiphillian) of Central Mexico, and the Origin of Equus. Bulletin of the Florida Museum of Natural History 43, 2002, S. 163–185 ().
- Oscar Carranza-Castañeda: Dinohippus mexicanus (Early-Late, Late, and Latest Hemphillian) and the Transition to Genus Equus, in Central Mexico Faunas. Frontiers in Earth Science 7, 2019, S. 89, doi:10.3389/feart.2019.00089.
- Victor Manuel Bravo-Cuevas und Eduardo Jiménez-Hidalgo: Evolutionary Significance of Equinae From the Mexican Neogene. Frontiers in Ecology and Evolution 7, 2019, S. 287, doi:10.3389/fevo.2019.00287.
- Ann Forstén und V. Eisenmann: Equus (Plesippus) simplicidens (Cope), not Dolichohippus. Mammalia 59, 1995, S. 85–89.
- Christina I. Barrón-Ortiz, Leonardo S. Avilla, Christopher N. Jass, Victor M. Bravo-Cuevas, Helena Machado und Dimila Mothé: What Is Equus? Reconciling Taxonomy and Phylogenetic Analyses. Frontiers in Ecology and Evolution 7, 2019, S. 343, doi:10.3389/fevo.2019.00343.
- Eric Scott: Pliocene and Pleistocene Horses from Porcupine Cave. In: Anthony Barnosky (Hrsg.): Biodiversity Response to Climate Change in the Middle Pleistocene: The Porcupine Cave Fauna from Colorado. University of California Press, 2004, S. 264–279.
- Lorenzo Rook, Raymond L. Bernor, Leonardo S. Avilla, Omar Cirilli, Lawrence Flynn, Advait Jukar, William Sanders, Eric Scott und Xiaoming Wang: Mammal Biochronology (Land Mammal Ages) Around the World From Late Miocene to Middle Pleistocene and Major Events in Horse Evolutionary History. Frontiers in Ecology and Evolution 7, 2019, S. 278, doi:10.3389/fevo.2019.00278.
- Ann Forstén: Middle Pleistocene replacement of stenonid horses by caballoid horses – ecological implications. Palaeogeography, Palaeoclimatology, Palaeoecology 65, 1988, S. 23–33.
- Boyan Sun und Tao Deng: The Equus datum and the early radiation of Equus in China. Frontiers in Ecology and Evolution 7, 2019, S. 429, doi:10.3389/fevo.2019.00429.
- Raymond Louis Bernor, Omar Cirilli, Shi-Qi Wang und Lorenzo Rook: Equus cf. livenzovensis from Montopoli, Italy (early Pleistocene; MN16b; ca. 2.6 Ma). Bollettino della Società Paleontologica Italiana 57 (3), 2018, S. 203–216, doi:10.4435/BSPI.2018.13.
- María Teresa Alberdi, E. Ortiz-Jaureguizar und J. L. Prado: A quantitative review of European stenonoid horses. Journal of Paleontology 72 (2), 1998, S. 371–387.
- Maria Rita Palombo und Maria Teresa Alberdi: Light and shadows in the evolution of South European stenonoid horses. Fossil Imprint, 73 (1–2), 2017, S. 115–140.
- María Teresa Alberdi und Maria Rita Palombo: The late Early to early Middle Pleistocene stenonoid horses from Italy. Quaternary International 288, 2013, S. 25–44.
- Nicolas Boulbes und Eline N. van Asperen: Biostratigraphy and Palaeoecology of European Equus. Frontiers in Ecology and Evolution 7, 2019, S. 301, doi:10.3389/fevo.2019.00301.
- Véra Eisenmann: Pliocene and Pleistocene Equids: Paleontology versus Molecular Biology. In: Ralf-Dietrich Kahlke, Lutz C. Maul und P. Mazza (Hrsg.): Late Neogene and Quaternary Biodiversity and Evolution: Regional Developments and Interregional Correlations. Proceedings Volume of the 18th International Senckenberg Conference (VI International Palaeontological Colloquium in Weimar), 25th–20th April 2004. Courier Forschungsinstitut Senckenberg 256, 2006, S. 71–89.
- Véra Eisenmann: Sussemionus, a new subgenus of Equus (Perissodactyla, Mammalia). Comptes Rendus Biologies 333, 2010, S. 235–240.
- Véra Eisenmann und Vasiliev Sergej: Unexpected finding of a new Equus species (Mammalia, Perissodactyla) belonging to a supposedly extinct subgenus in late Pleistocene deposits of Khakassia (southwestern Siberia). Geodiversitas 33 (3), 2011, S. 519–530.
- Jun-Xia Yuan, Xin-Dong Hou, Axel Barlow, Michaela Preick, Ulrike H. Taron, Federica Alberti, Nikolas Basler, Tao Deng, Xu-Long Lai, Michael Hofreiter und Gui Lian Sheng: Molecular identification of late and terminal Pleistocene Equus ovodovi from northeastern China. PLoS ONE 14 (5), 2019, S. e0216883, doi:10.1371/journal.pone.0216883.
- Jennifer J. Crees und Samuel T. Turvey: Holocene extinction dynamics of Equus hydruntinus, a late-surviving European megafaunal mammal. Quaternary Science Reviews 91, 2014, S. 16–29.
- Ludovic Orlando, Jessica L. Metcalf, Maria T. Alberdi, Miguel Telles-Antunes, Dominique Bonjean, Marcel Otte, Fabiana Martin, Véra Eisenmann, Marjan Mashkour, Flavia Morello, Jose L. Prado, Rodolfo Salas-Gismondi, Bruce J. Shockey, Patrick J. Wrinn, Sergei K. Vasil’ev, Nikolai D. Ovodov, Michael I. Cherry Blair Hopwood, Dean Male, Jeremy J. Austin, Catherine Hänni und Alan Cooper: Revising the recent evolutionary history of equids using ancient DNA. PNAS 106, 2009, S. 21754–21759.
- E. Andrew Bennett, Sophie Champlot, Joris Peters, Benjamin S. Arbuckle, Silvia Guimaraes, Mélanie Pruvost, Shirli Bar-David, Simon J. M. Davis, Mathieu Gautier, Petra Kaczensky, Ralph Kuehn, Marjan Mashkour, Arturo Morales-Muñiz, Erich Pucher, Jean-François Tournepiche, Hans Peter Uerpmann, Adrian Bălăşescu, Mietje Germonpré, Can Y. Gündem, Mahmoud-Reza Hemami, Pierre-Elie Moullé, Aliye Ötzan, Margarete Uerpmann, Chris Walzer, Thierry Grange und Eva-Maria Geigl: Taming the late Quaternary phylogeography of the Eurasiatic wild ass through ancient and modern DNA. PLoS ONE 12 (4), 2017, S. e0174216, doi:10.1371/journal.pone.0174216.
- Raymond L. Bernor, Miranda J. Armour-Chelu, Henry Gilbert, Thomas M. Kaiser und Ellen Schulz: Equidae. In: Lars Werdelin und William L. Sanders (Hrsg.): Cenozoic Mammals of Africa. University of California Press, 2010, S. 685–721.
- Véra Eisenmann: Origins, dispersals, and migrations of Equus (Mammalia, Perissodactyla). Courier Forschungsinstitut Senckenberg 153, 1992, S. 161–170.
- Jaco Weinstock, Eske Willerslev, Andrei Sher, Wenfei Tong, Simon Y.W. Ho, Dan Rubenstein, John Storer, James Burns, Larry Martin, Claudio Bravi, Alfredo Prieto, Duane Froese, Eric Scott, Lai Xulong, Alan Cooper: Evolution, Systematics, and Phylogeography of Pleistocene Horses in the New World: A Molecular Perspective. PLoS Biology 3 (8), 2005, S. 1373–1379 e241.
- Ludovic Orlando, Dean Male, Maria Teresa Alberdi, Jose Luis Prado, Alfredo Prieto, Alan Cooper und Catherine Hänni: Ancient DNA Clarifies the Evolutionary History of American Late Pleistocene Equids. Journal of Molecular Evolution 2008, S. 1–6.
- María Teresa Alberdi, Joaquín Arroyo-Cabrales, Alejandro H. Marín-Leyva und Oscar J. Polaco: Study of Cedral Horses and their place in the Mexican Quaternary. Revista Mexicana de Ciencias Geológicas 31 (2), 2014, S. 221–237.
- C. R. Harington und Marcia Eggleston-Stott: Partial carcass of a small Pleistocene horse from Last Chance Creek near Dawson City, Yukon. Current Research in the Pleistocene 13, 1996, S. 105–107.
- Helena Machado und Leonardo Avilla: The Diversity of South American Equus: Did Size Really Matter? Frontiers in Ecology and Evolution 7, 2019, S. 235, doi:10.3389/fevo.2019.00235.
- A. Azzaroli: Ascent and decline of monodactyl equids: a case for praehistoric overkill. Annales Zoologici Fennici 28, 1992, S. 151–163.
- Flavia Strani, Diana Pushkina, Hervé Bocherens, Luca Bellucci, Raffaela Sardella und Daniel DeMiguel: Dietary Adaptations of Early and Middle Pleistocene Equids From the Anagni Basin (Frosinone, Central Italy). Frontiers in Ecology and Evolution 7, 2019, S. 176, doi:10.3389/fevo.2019.00176.
- Véra Eisenmann, Evelyne Crégut-Bonnoure und Anne-Marie Moigne: Equus mosbachensis et les grands chevaux de la Caune l'Arago et de Lunel-Viel: craniologie comparèe. Bulletin du Museum national d'histoire naturelle Section C 4 (7), 1985, S. 157–173.
- R. Dale Guthrie: Frozen Fauna of the Mammoth Steppe: The Story of Blue Babe. University of Chicago Press, 1990, S. 1–324 (Kapitel 1: The curse of the frozen mammoths., S. 1–44).
- Gennady G. Boeskorov: Arctic Siberia: refuge of the Mammoth fauna in the Holocene. Quaternary International 142–143, 2006, S. 119–123.
- O. F. Chernova und G. G. Boeskorov: Identification of the Hair of a Holocene “Yukagir Horse” (Equus spp.) Mummy. Doklady Biological Sciences 462, 2015, S. 141–143.
- Véra Eisenmann: Gigantic horses. Advances in Vertebrate Paleontology „Hen to Panta“, 2003, S. 31–40.
- Carmen Nacarino-Meneses und Orlandi-Oliveras: The life history of European Middle Pleistocene equids: first insights from bone histology. Historical Biology: An International Journal of Paleobiology, 2019, doi:10.1080/08912963.2019.1655011.
- Irina V. Foronova: Late quaternary equids (genus Equus) of South-western and South-central Siberia. In: Marjan Mashkour (Hrsg.): Equids in Time and Space. Papers in Honour of Véra Eisenmann. Durham, 2002, S. 20–30.
- Ann Forstén: Size decrease in Late Pleistocene–Holocene caballoid horses (Genus Equus), intra- or interspecific evolution? A discussion of alternatives. Quaternary International 19, 1993, S. 71–75.
- Maria Teresa Alberdi, José L. Prado und Edgardo Ortiz-Jaureguizar: Patterns of body size changes in fossil and living Equini (Perissodactyla). Biological Journal of the Linnean Society 54, 1995, S. 349–370.
- Belcasem Bagtache, Djillali Hadjounis und Véra Eisenmann: Presence d'un Equus caballin (E. algericus n. sp.) et d'un autre espèce nouvelle d'Equus (E. melchiensis n. sp.) dans l'Atérien des Allobroges, Algérie. Comptes rendus de l'Académie des sciences 298, 1984, S. 609–612.
- Antoine Fages, Kristian Hanghøj, Naveed Khan, Charleen Gaunitz, Andaine Seguin-Orlando, Michela Leonardi, Christian McCrory Constantz, Cristina Gamba, Khaled A. S. Al-Rasheid, Silvia Albizuri, Ahmed H. Alfarhan, Morten Allentoft, Saleh Alquraishi, David Anthony, Nurbol Baimukhanov, James H. Barrett, Jamsranjav Bayarsaikhan, Norbert Benecke, Eloísa Bernáldez-Sánchez, Luis Berrocal-Rangel, Fereidoun Biglari, Sanne Boessenkool, Bazartseren Boldgiv, Gottfried Brem, Dorcas Brown, Joachim Burger, Eric Crubézy, Linas Daugnora, Hossein Davoudi, Peter de Barros Damgaard, María de los Ángeles de Chorro y de Villa-Ceballos, Sabine Deschler-Erb, Cleia Detry, Nadine Dill, Maria do Mar Oom, Anna Dohr, Sturla Ellingvåg, Diimaajav Erdenebaatar, Homa Fathi, Sabine Felkel, Carlos Fernández-Rodríguez, Esteban García-Viñas, Mietje Germonpré, José D. Granado, Jón H. Hallsson, Helmut Hemmer, Michael Hofreiter, Aleksei Kasparov, Mutalib Khasanov, Roya Khazaeli, Pavel Kosintsev, Kristian Kristiansen, Tabaldiev Kubatbek, Lukas Kuderna, Pavel Kuznetsov, Haeedeh Laleh, Jennifer A. Leonard, Johanna Lhuillier, Corina Liesau von Lettow-Vorbeck, Andrey Logvin, Lembi Lõugas, Arne Ludwig, Cristina Luis, Ana Margarida Arruda, Tomas Marques-Bonet, Raquel Matoso Silva, Victor Merz, Enkhbayar Mijiddorj, Bryan K. Miller, Oleg Monchalov, Fatemeh A. Mohaseb, Arturo Morales, Ariadna Nieto-Espinet, Heidi Nistelberger, Vedat Onar, Albína H. Pálsdóttir, Vladimir Pitulko, Konstantin Pitskhelauri, Mélanie Pruvost, Petra Rajic Sikanjic, Anita Rapan Papěsa, Natalia Roslyakova, Alireza Sardari, Eberhard Sauer, Renate Schafberg, Amelie Scheu, Jörg Schibler, Angela Schlumbaum, Nathalie Serrand, Aitor Serres-Armero, Beth Shapiro, Shiva Sheikhi Seno, Irina Shevnina, Sonia Shidrang, John Southon, Bastiaan Star, Naomi Sykes, Kamal Taheri, William Taylor, Wolf-Rüdiger Teegen, Tajana Trbojević Vukičević, Simon Trixl, Dashzeveg Tumen, Sainbileg Undrakhbold, Emma Usmanova, Ali Vahdati, Silvia Valenzuela-Lamas, Catarina Viegas, Barbara Wallner, Jaco Weinstock, Victor Zaibert, Benoit Clavel, Sébastien Lepetz, Marjan Mashkour, Agnar Helgason, Kári Stefánsson, Eric Barrey, Eske Willerslev, Alan K. Outram, Pablo Librado und Ludovic Orlando: Tracking Five Millennia of Horse Management with Extensive Ancient Genome Time Series. Cell 177, 2019, S. 1419–1435, doi:10.1016/j.cell.2019.03.049.
- Hiroki Goto, Oliver A. Ryder, Allison R. Fisher, Bryant Schultz, Sergei L. Kosakovsky Pond, Anton Nekrutenko und Kateryna D. Makova: A Massively Parallel Sequencing Approach Uncovers Ancient Origins and High Genetic Variability of Endangered Przewalski’s Horses. Genome Biology and Evolution 3, 2011, S. 1096–1106, doi:10.1093/gbe/evr067.
- Pablo Librado, Naveed Khan, Antoine Fages, Mariya A. Kusliy, Tomasz Suchan, Laure Tonasso-Calvière, Stéphanie Schiavinato, Duha Alioglu, Aurore Fromentier, Aude Perdereau, Jean-Marc Aury, Charleen Gaunitz, Lorelei Chauvey, Andaine Seguin-Orlando, Clio Der Sarkissian, John Southon, Beth Shapiro, Alexey A. Tishkin, Alexey A. Kovalev, Saleh Alquraishi, Ahmed H. Alfarhan, Khaled A. S. Al-Rasheid, Timo Seregély, Lutz Klassen, Rune Iversen, Olivier Bignon-Lau, Pierre Bodu, Monique Olive, Jean-Christophe Castel, Myriam Boudadi-Maligne, Nadir Alvarez, Mietje Germonpré, Magdalena Moskal-del Hoyo, Jarosław Wilczyński, Sylwia Pospuła, Anna Lasota-Kuś, Krzysztof Tunia, Marek Nowak, Eve Rannamäe, Urmas Saarma, Gennady Boeskorov, Lembi Lōugas, René Kyselý, Lubomír Peške, Adrian Bălășescu, Valentin Dumitrașcu, Roxana Dobrescu, Daniel Gerber, Viktória Kiss, Anna Szécsényi-Nagy, Balázs G. Mende, Zsolt Gallina, Krisztina Somogyi, Gabriella Kulcsár, Erika Gál, Robin Bendrey, Morten E. Allentoft, Ghenadie Sirbu, Valentin Dergachev, Henry Shephard, Noémie Tomadini, Sandrine Grouard, Aleksei Kasparov, Alexander E. Basilyan, Mikhail A. Anisimov, Pavel A. Nikolskiy, Elena Y. Pavlova, Vladimir Pitulko, Gottfried Brem, Barbara Wallner, Christoph Schwall, Marcel Keller, Keiko Kitagawa, Alexander N. Bessudnov, Alexander Bessudnov, William Taylor, Jérome Magail, Jamiyan-Ombo Gantulga, Jamsranjav Bayarsaikhan, Diimaajav Erdenebaatar, Kubatbeek Tabaldiev, Enkhbayar Mijiddorj, Bazartseren Boldgiv, Turbat Tsagaan, Mélanie Pruvost, Sandra Olsen, Cheryl A. Makarewicz, Silvia Valenzuela Lamas, Silvia Albizuri Canadell, Ariadna Nieto Espinet, Ma Pilar Iborra, Jaime Lira Garrido, Esther Rodríguez González, Sebastián Celestino, Carmen Olària, Juan Luis Arsuaga, Nadiia Kotova, Alexander Pryor, Pam Crabtree, Rinat Zhumatayev, Abdesh Toleubaev, Nina L. Morgunova, Tatiana Kuznetsova, David Lordkipanidze, Matilde Marzullo, Ornella Prato, Giovanna Bagnasco Gianni, Umberto Tecchiati, Benoit Clavel, Sébastien Lepetz, Hossein Davoudi, Marjan Mashkour, Natalia Ya. Berezina, Philipp W. Stockhammer, Johannes Krause, Wolfgang Haak, Arturo Morales-Muñiz, Norbert Benecke, Michael Hofreiter, Arne Ludwig, Alexander S. Graphodatsky, Joris Peters, Kirill Yu. Kiryushin, Tumur-Ochir Iderkhangai, Nikolay A. Bokovenko, Sergey K. Vasiliev, Nikolai N. Seregin, Konstantin V. Chugunov, Natalya A. Plasteeva, Gennady F. Baryshnikov, Ekaterina Petrova, Mikhail Sablin, Elina Ananyevskaya, Andrey Logvin, Irina Shevnina, Victor Logvin, Saule Kalieva, Valeriy Loman, Igor Kukushkin, Ilya Merz, Victor Merz, Sergazy Sakenov, Victor Varfolomeyev, Emma Usmanova, Viktor Zaibert, Benjamin Arbuckle, Andrey B. Belinskiy, Alexej Kalmykov, Sabine Reinhold, Svend Hansen, Aleksandr I. Yudin, Alekandr A. Vybornov, Andrey Epimakhov, Natalia S. Berezina, Natalia Roslyakova, Pavel A. Kosintsev, Pavel F. Kuznetsov, David Anthony, Guus J. Kroonen, Kristian Kristiansen, Patrick Wincker, Alan Outram und Ludovic Orlando: The origins and spread of domestic horses from the Western Eurasian steppes. Nature, 2021, doi:10.1038/s41586-021-04018-9.
- Hákon Jónsson, Mikkel Schubert, Andaine Seguin-Orlando, Aurélien Ginolhac, Lillian Petersen, Matteo Fumagallic, Anders Albrechtsen, Bent Petersen, Thorfinn S. Korneliussen, Julia T. Vilstrup, Teri Lear, Jennifer Leigh Myka, Judith Lundquist, Donald C. Miller, Ahmed H. Alfarhan, Saleh A. Alquraishi, Khaled A. S. Al-Rasheid, Julia Stagegaard, Günter Strauss, Mads Frost Bertelsen, Thomas Sicheritz-Ponten, Douglas F. Antczak, Ernest Bailey, Rasmus Nielsen, Eske Willerslev und Ludovic Orlando: Speciation with gene flow in equids despite extensive chromosomal plasticity. PNAS 111 (52), 2014, S. 18655–18660.
- Ludovic Orlando, Marjan Mashkour, Ariane Burke, Christophe J. Douady, Véra Eisenmann und Catherine Hänni: Geographic distribution of an extinct equid (Equus hydruntinus: Mammalia, Equidae) revealed bymorphological and genetical analyses of fossils. Molecular Ecology 15, 2006, S. 2083–2093
- Raymond L. Bernor und Miranda Armour-Chelu: Family Equidae. In: Gertrud E. Rössner und Kurt Heissig: The Miocene land mammals of Europe. München, 1999, S. 193–202.
- Wighart von Koenigswald: Lebendige Eiszeit. Klima und Tierwelt im Wandel. Stuttgart, 2002, S. 62–66.
- Omar Cirilli, Luca Pandolfi, Lorenzo Rook und Raymond L. Bernor: Evolution of Old World Equus and origin of the zebra‑ass clade. Scientific Reports 11, 2021, S. 10156, doi:10.1038/s41598-021-89440-9
- John Ray: Synopsis methodica animalium quadrupedium et serpentini generis. London, 1693, S. 1–336 (S. 57) ().
- Carl von Linné: Systema naturae. 10. Auflage, 1758, Band 1, S. 73–74 ().
- Johann Friedrich Blumenbach: Handbuch der Naturgeschichte., Göttingen, 1797, S. 1–714 (S. 106–109) ().
- Johann Karl Wilhelm Illiger: Prodromus systematis mammalium et avium additis terminis zoographicis utriudque classis. Berlin, 1811, S. 1–301 (S. 101) ().
- John Edward Gray: A revision of the family Equidae. Zoological Journal, 1825, S. 241–248 ().
- Charles Hamilton Smith: The natural History of horses. London, Dublin, 1841, S. 1–352 (S. 321) ().
- Edmund Heller: New genera and races of African ungulates. Smithsonian Miscellaneous Collections 60 (8), 1912, S. 1–16 ().
- Wolfgang Otto Dietrich: Hemionus Pallas im Pleistozän von Berlin. Vertebrata Palasiatica 3 (1), 1959, S. 13–22.
- George Gaylord Simpson: The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History 85, 1945, S. 1–350 (S. 137 und 253–254).
- Richard Owen: Odontography; or, A treatise on the comparative anatomy of the teeth; their physiological relations, mode of development, and microscopic structure, in the vertebrate animals. Hippolyte Bailliere, London 1840–1845, S. 572–582 ().
- Marcellin Boule: Observations sur quelques équidés fossiles. Bulletin de la Société géologique de France 27 (2), 1899, S. 531–542 ().
- Igino Cocchi: L'uomo fossile dell'Italia Centrale. Memorie Societa Italiana Science Naturale 2 (N7), 1867, S. 1–180 (S. 18, 20) ().
- A. Tendell Hopwood: The Former Distribution of Caballine and Zebrine Horses in Europe and Asia. Proceedings of the Zoological Society of London 106 (4), 1936, S. 897–912.
- Paul O. McGrew: An Early Pleistocene (Blancan) fauna from Nebraska. Geological Series, Field Museum of Natural History 9 (2), 1944, S. 33–66 ().
- Matthew George Jr., und Oliver A. Ryder: Mitochondrial DNA Evolution in the Genus Equus. Molecular Biology and Evolution 3 (6), 1986, S. 535–546.
- Pieter Boddaert: Elenchus animalium. Volume I. Rotterdam, 1785, S. 1–174 (S. 159–161) ().
- Leopold Fitzinger: Wissenschaftlich-populäre Naturgeschichte der Säugethiere in ihren sämmtlichen Hauptformen. Nebst einer Einleitung in die Naturgeschichte überhaupt und in die Lehre von den Thieren insbesondere. III. Band. Wien, 1860, S. 1–380 (S. 434) ().
- International Commission on Zoological Nomenclature: Opinion 2027 (Case 3010). Usage of 17 specific names based on wild species which are pre-dated by or contemporary with those based on domestic animals (Lepidoptera, Osteichthyes, Mammalia): conserved. Bulletin of the Zoological Nomenclature 60 (1), 2003, S. 81–84 ().
- International Commission on Zoological Nomenclature: Opinion 271. Addition to the „Official list of generic names in zoology“ of the generic names „Equus“ Linnaeus, 1758 (Class Mammalia) and „Alca“ Linnaeus, 1758 (Class Aves) („Opinion“ supplementary to „Opinion“ 16). Opinions and declarations rendered by the International Commission on Zoological Nomenclature 6, 1954–-1955, S. 43–50 ().
- Anthea Gentry, Juliet Clutton-Brock und Colin P. Groves: The naming of wild animal species and their domestic derivatives. Journal of Archaeological Science 31, 2004, S. 645–651.
- Wolfgang Zessin, Elke Gröning und Carsten Brauckmann: Bemerkungen zur Systematik rezenter Equidae (Mammalia). Ursus, Mitteilungsblatt des Zoovereins und des Zoos Schwerin, 15 (1), 2009, S. 20–31.
- Jennifer A. Leonard, Nadin Rohland, Scott Glaberman, Robert C. Fleischer, Adalgisa Caccone und Michael Hofreiter: A rapid loss of stripes: the evolutionary history of the extinct quagga. Biological Letters 1, 2005, S. 291–295.
- Einträge „Pferd“ () und „Esel“ () bei Duden.de.
- Elaine Turner: Miesenheim I. Excavations at a Lower Palaeolithic site in the Central Rhineland of Germany. Römisch-Germanisches Zentralmuseum Monographien 44, 2000, S. 1–151 (S. 69–78).
- Walter Steiner: Der Travertin von Ehringsdorf und seine Fossilien. Lutherstadt Wittenberg, 1981, S. 1–200 (S. 127–132).
- Thomas Laurat und Enrico Brühl: Zum Stand der archäologischen Untersuchungen im Tagebau Neumark-Nord, Kr. Merseburg-Querfurt (Sachsen-Anhalt) – Vorbericht zu den Ausgrabungen 2003-2005. Jahresschrift für Mitteldeutsche Vorgeschichte 90, 2006, S. 9–69.
- Hartmut Thieme: Lower Palaeolithic hunting spears from Germany. Nature 385, 1997, S. 807–810.
- Hartmut Thieme: The Lower Palaeolithic art of hunting. The case of Schöningen 13 II-4, Lower Saxony, Germany. In: Clive Gamble und Martin Porr (Hrsg.): The hominid individual in context. London, New York, 2005, S. 115–132.
- Jordi Serangeli, Utz Böhner, Thijs Van Kolfschoten und Nicholas J. Conard: Overview and new results from large-scale excavations in Schöningen.Journal of Human Evolution 89 (2015) 27–45.
- Eric Boёda, J. M. Geneste und C. Griggo: A Levallois point embedded in the vertebra of a wild ass (Equus africanus): hafting, projectiles and Mousterian hunting weapons. Antiquity 73, 1999, S. 394–402.
- Ingmar M Braun und Wolfgang Zessin: Pferdedarstellungen in der paläolithischen Wandkunst und der Versuch ihrer zoologisch-ethologischen Interpretation. Ursus, Mitteilungsblatt des Zoovereins und des Zoos Schwerin 17 (1), 2011, S. 4–26.
- Michel Lorblanchet: Höhlenmalerei. Ein Handbuch. Sigmaringen 1997, S. 1–340.
- Gerhard Bosinski: Die große Zeit der Eiszeitjäger. Europa zwischen 40.000 und 10.000 v. Chr. Jahrbuch des Römisch-Germanischen Zentralmuseums Mainz 34, 1987, S. 3–139.
- Gerhard Bosinski: Die Kunst der Eiszeit in Deutschland und der Schweiz. Kataloge Vor- und Frühgeschichtlicher Altertümer 20, Bonn, 1982, S. 1–91.
- Birgitta Kimura, Fiona B. Marshall, Shanyuan Chen, Sónia Rosenbom, Patricia D. Moehlman, Noreen Tuross, Richard C. Sabin, Joris Peters, Barbara Barich, Hagos Yohannes, Fanuel Kebede, Redae Teclai, Albano Beja-Pereira und Connie J. Mulligan: Ancient DNA from Nubian and Somali wild ass provides insights into donkey ancestry and domestication. Proceedings of the Royal Society B 278, 2011, S. 50–57, doi:10.1098/rspb.2010.0708.
- Stine Rossel, Fiona B. Marshall, Joris Peters, Tom Pilgram, Matthew D. Adams und David O’Connor: Domestication of the donkey: Timing, processes, and indicators. PNAS 105 (1), 2008, S. 3715–3720.
- Haskel J. Greenfield, Itzhaq Shai, Tina L. Greenfield, Elizabeth R. Arnold, Annie Brown, Adi Eliyahu und Aren M. Maeir: Earliest evidence for equid bit wear in the ancient Near East: The "ass" from Early Bronze Age Tell eṣ-Ṣâfi/Gath, Israel. PLoS ONE 13 (5), 2018, S. e0196335, doi:10.1371/journal.pone.0196335.
- Achilles Gautier und Wim van Neer: Animal remains from predynastic sites in the Nagada region, Middle Egypt. Archaeofauna 18, 2009, S. 27–50.
- Peter Mitchell: The Donkey in Human History: An Archaeological Perspective. Oxford University Press, 2018, S. 1–305 (S. 14–71)
- Joséphine Lesur: Fishing and herding in the Nile Delta during the Predynastic (4th Millenium BC). In: C. Çakırlar, J. Chahoud, R. Berthon und S. Pilaar Birch: Archaeozoology of the Near East XII: Proceedings of the 12th International Symposium of the ICAZ Archaeozoology of Southwest Asia and Adjacent Areas Working Group, Groningen Institute of Archaeology, June 14-15 2015, University of Groningen, the Netherlands. Groningen, 2018, S. 59–72.
- Albano Beja-Pereira, Phillip R. England, Nuno Ferrand, Steve Jordan, Amel O. Bakhiet, Mohammed A. Abdalla, Marjan Mashkour, Jordi Jordana, Pierre Taberlet, Gordon Luikart: African Origins of the Domestic Donkey. Science 304, 2004, S. 1781.
- E. Andrew Bennett, Jill Weber, Wejden Bendhafer, Sophie Champlot, Joris Peters, Glenn M. Schwartz, Thierry Grange und Eva-Maria Geigl: The genetic identity of the earliest human-made hybrid animals, the kungas of Syro-Mesopotamia. Science Advances 8, 2022, S. eabm0218, doi:10.1126/sciadv.abm0218.
- Dorcas Brown und David Anthony: Bit wear, horseback riding and the Botai site in Kazakstan. Journal of Archaeological Science 25, 1998, S. 331–347.
- David W. Anthony und Dorcas R. Brown: Eneolithic horse exploitation in the Eurasian steppes: diet, ritual and riding. Antiquity 74, 2000, S. 75–387.
- David W. Anthony: The Horse, the Wheel, and Language. Princeton University Press, 2007, S. 1–553 (S. 193–224) ().
- Alan K. Outram, Natalie A. Stear, Robin Bendrey, Sandra Olsen, Alexei Kasparov, Victor Zaibert, Nick Thorpe und Richard P. Evershed: The Earliest Horse Harnessing and Milking. Science 323, 2009, S. 1332–1335.
- William Timothy Treal Taylor und Christina Isabelle Barrón‑Ortiz: Rethinking the evidence for early horse domestication at Botai. Scientific Reports 11, 2021, S. 7440, doi:10.1038/s41598-021-86832-9.
- Vera Warmuth, Anders Eriksson, Mim Ann Bower, Graeme Barker, Elizabeth Barrett, Bryan Kent Hanks, Shuicheng Li, David Lomitashvili, Maria Ochir-Goryaeva, Grigory V. Sizonov, Vasiliy Soyonov und Andrea Manica: Reconstructing the origin and spread of horse domestication in the Eurasian steppe. PNAS 109 (21), 2012, S. doi:10.1073/pnas.1111122109.
- Pablo Librado, Antoine Fages, Charleen Gaunitz, Michela Leonardi, Stefanie Wagner, Naveed Khan, Kristian Hanghøj, Saleh A. Alquraishi, Ahmed H. Alfarhan, Khaled A. Al-Rasheid, Clio Der Sarkissian, Mikkel Schubert und Ludovic Orlando.The Evolutionary Origin and Genetic Makeup of Domestic Horses. Genetics 204, 2016, S. 423–434, doi:10.1534/genetics.116.194860.
- Arne Ludwig, Melanie Pruvost, Monika Reissmann, Norbert Benecke, Gudrun A. Brockmann, Pedro Castaños, Michael Cieslak, Sebastian Lippold, Laura Llorente, Anna-Sapfo Malaspinas, Montgomery Slatkin und Michael Hofreiter: Coat Color Variation at the Beginning of Horse Domestication. Science 324, 2009, S. 485, doi:10.1126/science.1172750.
- Saskia Wutke, Norbert Benecke, Edson Sandoval-Castellanos, Hans-Jürgen Döhle, Susanne Friederich, Javier Gonzalez, Jón Hallsteinn Hallsson, Michael Hofreiter, Lembi Lõugas, Ola Magnell, Arturo Morales-Muniz, Ludovic Orlando, Albína Hulda Pálsdóttir, Monika Reissmann, Matej Ruttkay, Alexandra Trinks und Arne Ludwig: Spotted phenotypes in horses lost attractiveness in the Middle Ages. Scientific Reports 6, 2016, S. 38548, doi:10.1038/srep38548.
- Arne Ludwig, Monika Reissmann, Norbert Benecke, Rebecca Bellone, Edson Sandoval-Castellanos, Michael Cieslak, Gloria G. Fortes, Arturo Morales-Muñiz, Michael Hofreiter und Melanie Pruvost: Twenty-five thousand years of fluctuating selection on leopard complex spotting and congenital night blindness in horses. Philosophical Transactions of the Royal Society B 370, 2015, S. 20130386, doi:10.1098/rstb.2013.0386.
- Burchard Brentjes: Das älteste Reittier des Menschen. Berlin 1960.
- Hans-Jürgen Döhle und Torsten Schunke: Der erste neolithische Pferdeschädel Mitteldeutschlands – ein frühes Hauspferd? In: Harald Meller und Susanne Friederich (Hrsg.): Salzmünde-Schiepzig – ein Ort, zwei Kulturen. Ausgrabungen an der Westumfahrung Halle (A 143) Teil I. Sonderband Archäologie in Sachsen-Anhalt 21/I, 2014, S. 257–261.
- Thomas Jansen, Peter Forster, Marsha A. Levine, Hardy Oelke, Matthew Hurles, Colin Renfrew, Jürgen Weber und Klaus Olek: Mitochondrial DNA and the origin of the domestic horse. PNAS 99 (16), 2002, S. 10905–10910.
- Cristina Luís, Cristiane Bastos-Silveira, E. Gus Cothran und Maria do Mar Oom: Iberian Origins of New World Horse Breeds. Journal of Heredity 97 (2), 2006, S. 107–113, doi:10.1093/jhered/esj020.
- Anas Khanshour, Rytis Juras, Rick Blackburn und E. Gus Cothran: The Legend of the Canadian Horse: Genetic Diversity and Breed Origin. Journal of Heredity 106 (1), 2015, S. 37–44, doi:10.1093/jhered/esu074.
- Igor V. Ovchinnikov, Taryn Dahms, Billie Herauf, Blake McCann, Rytis Juras, Caitlin Castaneda und E. Gus Cothran: Genetic diversity and origin of the feral horses in Theodore Roosevelt National Park. PLoS ONE 13 (8), 2018, S. e0200795, doi:10.1371/journal.pone.0200795.
- Jared Diamond: Evolution, consequences and future of plant and animal domestication. Nature 418, 2002, S. 700–707.
- B. Megersa, D. Biffa und B. Kumsa: A mysterious zebra-donkey hybrid (zedonk or zonkey) produced under natural mating: A case report from Borana, southern Ethiopia. Animal Production Research Advances 2 (3), 2006, S. 148–154.
- IUCN Red List of Threatened Species, abgerufen am 24. Februar 2013.
- The Donkey Sanctuary: Under the skin. Update on the global crisis for donkeys and the people who depend on them. Bericht des Donkey Sanctuary November 2019, online ().
Weblinks
- Gefährdungsgrad der einzelnen Arten in der Roten Liste gefährdeter Arten der IUCN.
- Literatur von und über Pferde im Katalog der Deutschen Nationalbibliothek