Oligozän
Das Oligozän ist in der Erdgeschichte eine chronostratigraphische Serie (also ein Zeitintervall) innerhalb des Systems des Paläogens, früher des Tertiärs. Der Beginn des Oligozäns liegt bei etwa 33,9 Millionen Jahren, es endete vor etwa 23,03 Millionen Jahren. Dem Oligozän folgte das Miozän und ihm ging das Eozän voraus.
| System | Serie | Stufe | ≈ Alter (mya) |
|---|---|---|---|
| später | später | später | jünger |
| Paläogen | Oligozän | Chattium | 23,03 ⬍ 28,1 |
| Rupelium | 28,1 ⬍ 33,9 | ||
| Eozän | Priabonium | 33,9 ⬍ 38 | |
| Bartonium | 38 ⬍ 41,3 | ||
| Lutetium | 41,3 ⬍ 47,8 | ||
| Ypresium | 47,8 ⬍ 56 | ||
| Paläozän | Thanetium | 56 ⬍ 59,2 | |
| Seelandium | 59,2 ⬍ 61,6 | ||
| Danium | 61,6 ⬍ 66 | ||
| früher | früher | früher | älter |
Namensgebung und Geschichte
Der Name Oligozän (von Griech. ὀλίγος = „wenig, gering, schwach“ und καινός = „neu, ungewöhnlich“) wurde von Heinrich Ernst Beyrich 1854 vorgeschlagen.[1]
Definition und GSSP
Der Beginn des Oligozäns (und damit auch der Rupelium-Stufe) ist durch das Aussterben der Foraminiferen-Gattung Hantkenina definiert. Die Obergrenze (und damit die Untergrenze von Miozän und des Aquitaniums) wird durch mehrere Ereignisse gut definiert. Kennzeichnend ist die Basis der Magnetischen Polaritäts-Chronozone C6Cn.2n, das Erstauftreten der Foraminiferen-Art Paragloborotalia kugleri und das Aussterben der kalkigen Nannoplankton-Art Reticulofenestra bisecta (Basis der Nannoplankton-Zone NN1). Der GSSP (= globaler Eichpunkt) für die Basis des Oligozäns (und die Basis der Rupelium-Stufe) ist das Massignano-Profil in einem aufgelassenen Steinbruch auf der Ostseite der Straße von Ancona nach Sirolo an der Adriaküste nahe dem Dorf Massignano (Ancona, Italien).
Untergliederung
Das Oligozän wird in zwei chronostratigraphische Stufen unterteilt:
Regional und in älteren Publikationen werden auch noch weitere Stufennamen verwendet. Sie eigneten sich aus verschiedensten Gründen nicht als internationale Stufen und werden nur noch regional benutzt oder sind wegen mangelhafter Definition außer Gebrauch gekommen.
Geographie und Klima
Bis in das Eozän existierte mit den verbundenen Landflächen von Australien, Antarktika und Südamerika noch ein relativ umfangreicher Rest des früheren südlichen Großkontinents Gondwana. Damals war der zirkumpolare Meeresstrom, der heute Antarktika im Uhrzeigersinn umfließt, noch nicht vorhanden. Stattdessen drang wärmeres Meerwasser entlang der kontinentalen Küstenbereiche weit nach Süden vor und gestaltete das antarktische Klima deutlich milder. Im Mittleren Eozän vor etwa 45 Millionen Jahren erfolgte die endgültige Trennung von Antarktika und Australien, und während des Übergangs zum Oligozän (34 mya) zerbrach auch die Landbrücke zu Südamerika. Dies führte zur Bildung der heute etwa 480 Seemeilen breiten Drakestraße, und als unmittelbare Folge davon entstand der Antarktische Zirkumpolarstrom, der Antarktika nunmehr thermisch isolierte und wahrscheinlich – unter Mitwirkung verschiedener Rückkopplungen – den Anstoß für einen weltweiten Abkühlungsprozess gab. Auf globaler Ebene sank die Temperatur der Ozeane bis in tiefere Schichten um 4 bis 5 °C, und der Meeresspiegel fiel innerhalb einer geologisch sehr kurzen Zeitspanne um etwa 30 Meter. Auffällig in dem Zusammenhang ist die abrupte Reduzierung der atmosphärischen CO2-Konzentration. Bewegte sich diese gegen Ende des Eozäns in einem Bereich von 700 bis 1.000 ppm, nahm dieser Wert um etwa 40 Prozent ab (und lag eventuell für einige Jahrtausende noch tiefer).[2] Die bei einem CO2-Schwellenwert um 600 ppm einsetzende Vereisung des südpolaren Festlands, in der Anfangsphase zum Teil gesteuert von den sich zyklisch verändernden Erdbahnparametern und der dadurch variierenden Sonneneinstrahlung, markiert den Beginn des Känozoischen Eiszeitalters.[3][4] Parallel dazu verzeichneten auch die arktischen Regionen und besonders Grönland eine deutliche Abkühlung. Unter anderem belegen Funde von Dropstones die zeitweilige Existenz von grönländischem Kontinentaleis.[5][6]
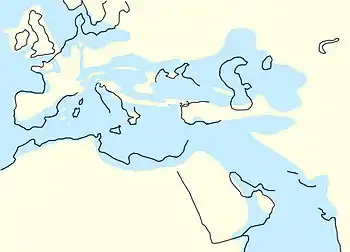
Die zunehmende Ausdehnung der Eiskappen und Gletscher führte dazu, dass zahlreiche Schelfmeere verlandeten, wodurch verschiedene neue Landverbindungen entstanden. So fiel am Beginn des Oligozäns die Turgaistraße trocken, die über Millionen Jahre als Flachmeer die Grenze zwischen Asien und Europa bildete. Auch das einst überflutete Ebro-Becken südwestlich der Pyrenäen wurde Festland, was zur vollständigen Verbindung der Iberischen Halbinsel mit Europa führte. Tektonische Verschiebungen bewirkten eine Teilung der Tethys in das Mittelmeer im Westen und der Paratethys im Osten. Die Paratethys befand sich im Osten des heutigen Europas und stand nur zeitweilig mit dem Mittelmeer und dem Indischen Ozean in Verbindung. Das Mittelmeer war aber zum Atlantik hin geöffnet. Zusätzlich wird angenommen, dass der lange Zeit isolierte Arktische Ozean nach einem Übergangsstadium als Brackwassermeer im frühen Oligozän (≈ 32 mya) mit dem Einströmen von salzhaltigem Nordatlantikwasser Anschluss an die globale Meereszirkulation fand.[7] Der Afrikanische Kontinent blieb nach wie vor weitgehend von Europa und Asien isoliert. Dies sollte sich erst im Miozän grundlegend ändern.[8] Daneben erreichte die Auffaltung der Alpen[9] und der Rocky Mountains einen Höhepunkt.
Für große Teile des späteren Oligozäns gilt eine durchschnittliche CO2-Konzentration im Bereich von 400 bis 450 ppm als wahrscheinlich.[10] Vor 27,8 Millionen Jahren kam es zu einer etwa 3 Millionen Jahre dauernden Periode mit wärmeren Temperaturen, etwas schwächer ausgeprägt als das Klimaoptimum des Miozäns, aber stark genug, um bereits in mittleren Breiten subtropische Bedingungen zu etablieren.[11] Diese Klimasignale kamen jedoch besonders in der Südpolaregion kaum zu Geltung. Es deutet somit Vieles darauf hin, dass der Kernbereich des Antarktischen Eisschilds zu dieser Zeit bereits eine Ausdehnung erreicht hatte, die mit verminderter Empfindlichkeit gegenüber globalen Klimaeinflüssen einherging.[11]
Im zeitlichen Umkreis des Oligozän-Miozän-Übergangs begann erneut eine signifikante, aber relativ kurzzeitige Abkühlungsphase, verbunden mit einem weiteren Wachstum der antarktischen Eisbedeckung und entsprechender Absenkung des Meeresspiegels.[12]
Paläogeographie in Mitteleuropa
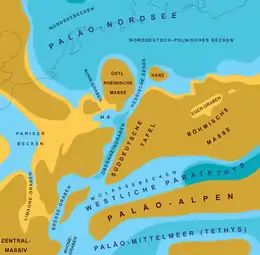
In Deutschland zur Zeit des Oligozäns (vor etwa 30 Millionen Jahren) reichte die Nordsee bis zum Niederrhein und in den Raum von Kassel (Hessen). Ostdeutschland war bis zum Harz und bis Leipzig (Sachsen) vom Meer überflutet. Zwischen der Nordsee und dem Tethys-Restmeer im heutigen Alpenraum erstreckte sich eine etwa 300 Kilometer lange und maximal 40 Kilometer breite Meeresstraße. Sie verlief von Kassel ausgehend über die Wetterau-Senke und den Oberrheingraben bis nach Basel (Schweiz). Am Nordende des Oberrheingrabens lag zwischen Odenwald, Spessart, Taunus, Hunsrück und Pfälzer Bergland ein Binnenmeer, das fast zehnmal so groß war wie der heutige Bodensee. In dieser Meeresstraße lebten Haie, Rochen und Seekühe.
Faunenentwicklung
Frühes Oligozän
Durch den Trockenfall der Turgaistraße entstand am Beginn des Oligozäns eine Landverbindung zwischen Europa und Asien. Dies ermöglichte es zahlreichen Tierformen, vom Osten nach Europa vorzudringen, was zusammen mit gravierenden klimatischen Veränderungen zu größeren Verlusten unter der in Europa heimischen Fauna führte. Davon betroffen waren etwa 60 Prozent der europäischen Säugetiergattungen. Dieser Faunenaustausch am Beginn des Oligozäns wurde vom schweizerischen Paläontologen Hans Georg Stehlin im Jahr 1909 erstmals als Grande Coupure („Großer Einschnitt“) bezeichnet. Zu den Opfern dieses Faunenaustauschs zählten die meisten der Palaeotherien. Nur wenige Gattungen, wie Palaeotherium und Plagiolophus, überlebten die Grande Coupure. Auch die Primaten verschwanden damals fast vollständig aus Europa. Lediglich die Gattung Pseudoloris überlebte etwas länger im heutigen Spanien, starb aber noch im Oligozän aus, ebenso wie die Leptictida, die Apatemyiden und die letzten Ischromyiden mit der Gattung Plesiarctomys. Das Aussterben zahlreicher Baum- oder Waldbewohner deutet darauf hin, dass sich am Beginn des Oligozäns offenere Lebensräume weiter ausbreiteten. Im Gegensatz dazu überlebten die Beuteltiere in Europa ohne größere Verluste.[8]
Bei den Neuankömmlingen aus Asien fallen besonders die ersten Nashörner Europas auf. Diese waren im Oligozän Europas durch die hornlosen Hyracodonten und Amynodonten sowie durch die gehörnten Nashörner der Diceratheriinae und die Menoceratherien vertreten. Unter den Hyracodonten wanderte das schafsgroße Eggysodon am Beginn des Oligozäns nach Europa ein. Die Amynodonten waren vom Beginn bis zum Ende der Epoche mit Cadurcotherium in Europa verbreitet. Weitere Nashörner des europäischen Oligozäns waren Ronzotherium und Epiaceratherium. Diese Tiere begründeten den später erfolgreichen Stamm der Aceratheriinae. Die westlichen Teile Europas wurden im Verlauf des Oligozäns darüber hinaus auch vom Paraceratherium erreicht, dem größten landlebenden Säugetier aller Zeiten, das vor allem in Asien verbreitet war.[8] Neben den Nashörnern und ihren näheren Verwandten wanderte am Beginn des Oligozäns mit den Chalicotherien eine weitere äußerst erfolgreiche Gruppe von Unpaarhufern aus Asien ein. Die erste Gattung, die Europa erreichte, war Schizotherium.

Die Paarhufer waren vom großen Faunenaustausch am Beginn des Oligozäns weniger betroffen. Zahlreichen europäischen Familien und Gattungen gelang der Übergang ins Oligozän, wie zum Beispiel den Dichobuniden mit Dichobune, Tapirulus und Diplobune. Auch die Cainotheriidae überlebten mit der hasengroßen Form Plesiomeryx und dem größeren Caenomeryx. Allerdings starben auch unter den Paarhufern einige Linien aus, wie die Dacrytheriden, Xiphodontiden und Anoplotheriden. Ursache dafür könnte die Überlegenheit der gelociden Wiederkäuer gewesen sein, die im Oligozän mit Formen wie Gelocus, Lophiomeryx und Bachitherium eine große Formenfülle erreichten. Auffällige Paarhufer des Oligozäns waren die großen Anthracotherien, die mit der Gattung Anthracotherium riesige, flusspferdgroße Formen hervorbrachten. Andere, wie Bothriodon, Elomeryx und Methriotherium, waren kleiner und ähnlich groß wie heutige Schweine. Im Oligozän erreichten auch erstmals die riesigen schweineähnlichen Entelodonten mit Entelodon Europa. Allein der Schädel dieser Tiere war bis zu 1 m lang. Außerdem erschienen die ersten näheren Verwandten der Schweine mit den Gattungen Palaeochoerus und Doliochoerus.[8]

Auch die Räuber unter den Säugetieren waren am Beginn des europäischen Oligozäns einem drastischen Wandel unterworfen. Relativ wenig betroffen waren lediglich die archaischen Creodonten. Einige Formen, wie etwa Pterodon, starben aus, doch wurden etwa die Vertreter der Gattung Hyaenodon durch asiatische Arten ersetzt. Weitaus stärkeren Wandlungen unterlagen die Vertreter der Ordnung Carnivora (Raubtiere). So traten mit Eusmilus die ersten Nimraviden auf, katzenähnliche Räuber, die gewaltige Säbelzähne ausbildeten. Eine weitere Gattung der Familie, Nimravus, besaß kleinere Zähne. Sie erschien geringfügig später, aber ebenfalls im frühen Oligozän. Ein weiterer Angehöriger der Nimraviden war Quercylurus, das größte Raubtier seiner Epoche. Es erreichte die Ausmaße eines Bären und scheint ein Sohlengänger gewesen zu sein. Von den Arctoiden verschwand Cynodictis am Übergang zum Oligozän und wurde von Aphicyonodon ersetzt. Unter den kleineren Raubtieren des Eozäns waren Tiere, die an heutige Zibetkatzen erinnerten, wie Stenoplescitis und die nah verwandte Gattung Palaeoprinodon.
Bei den Kleinsäugern fällt im frühen Oligozän das erstmalige Erscheinen der Lagomorpha (Hasenartige) in Europa auf. Unter den Insektenfressern wurden die Maulwürfe (Eotalpa) und Spitzmäuse dominant. Die Nager waren wie bereits im Eozän vor allem durch Theridomyiden und Bilche (Gliridae) vertreten. Als Neuerscheinungen traten erstmals moderne Familien wie Wühler, Hörnchen, Biber und die heute auf Nordamerika beschränkten Aplodontia auf. Die Hörnchen waren etwa durch Palaeosciurus vertreten, die Biber durch Stenofiber. Daneben traten erstmals die Eomyiden auf, die im Oligozän und Miozän typisch waren, aber schließlich ausstarben. Der kennzeichnendste Neuankömmling unter den Kleinsäugern war allerdings Atavocricetodon, der erste Vertreter der Wühler (Cricetidae).
Ein oligozänes Krokodil Europas war Hispanochampsa,[8] auch bekannt als Diplocynodon muelleri.
Spätes Oligozän

Der ausgeprägte Wechsel klimatischer Bedingungen während des Oligozäns führte zur Ausbreitung von Trockengebieten, was vermutlich den Untergang der Palaeotherien mit Plagiolophus, der meisten Anthracotheriden und der Gattung Entelodon in Europa besiegelte. Im Verlauf des späten Oligozäns verschwanden auch die hyracodonten Nashörner sowie das Nashorn Ronzotherium. Dagegen wurden die Menoceratherien dominant und aus der früholigozänen Gattung Epiceratherium entstanden die Gattungen Menoceras und Protaceratherium. Aus diesem Stamm der Menoceratherien entwickelten sich bis zum frühen Miozän die drei Nashorngruppen der Rhinocerotinen, Anthracerotherinen und Teleoceratherinen. Die älteste bekannte Gattung der Aceratherien, Mesaceratherium entstand noch im späten Oligozän, ebenso wie Brachydiceratherium, die erste Gattung der Teleoceratherien. Als früher Rhinocerotheride gilt Menoceras. Außerdem tauchten im späten Oligozän die ersten Tapire in Europa auf. Diese gehörten zur Gattung Protapirus, die offenbar bereits einen Rüssel, ganz ähnlich wie heutige Tapire, besaß.
Unter den Paarhufern war für das späte Oligozän die Diversifizierung der moschoiden Wiederkäuer typisch. Darunter sind die Gattungen Prodremotherium und Bedenomeryx, die aus Gelocus hervorgegangen waren. Sie hatten im Gegensatz zu diesem längere Beine und ein verändertes Kanonenbein.[8]
Unter den Kleintieren mussten die theridomyden Nager herbe Verluste hinnehmen. Zu den überlebenden Gattungen zählten Blainvillimys und Issidoromys, eine Gattung, die hypsodonte Zähne entwickelte. Die Bilche und die Eomyiden brachten verschiedene neue Gattungen hervor. Deutlich mehr verschiedene Formen entstanden allerdings unter den Cricetiden, die in zahlreichen neuen Gattungen mit komplexen Backenzähnen erschienen. Eine darunter, Melissiodon, überlebte bis ins frühe Miozän und scheint baumlebend gewesen zu sein.[8]
In Europa verschwanden unter den Raubtieren im Verlauf des späten Oligozäns die großen Nimraviden, wie Eusmilus, Nimravus und Quercylurus. Dies scheint allerdings ein lokaler Vorgang gewesen zu sein. Auch die Creodonten waren im Niedergang begriffen. Die letzten verschwanden in Europa mit Hyaenodon am Ende des Oligozäns. In Afrika überlebten sie jedoch länger und gelangten von dort aus im Miozän sogar kurzzeitig nach Europa zurück. Im Gegenzug verbreiteten sich die Raubtiere (Carnivora) weiter. So entwickelten sich aus den Gattungen Stenolescitis und Palaeoprionodon verschiedene andere Räuber wie Haplogale, Stenogale, und Proailurus. Proailurus war etwa so groß wie ein Ozelot und gilt als erste Gattung der Katzen. Außerdem verbreiteten sich die Amphicyoniden. Dies waren Raubtiere, die wie Mischwesen aus Hunden und Bären ausgesehen haben dürften. Die ersten Formen des Oligozäns waren recht kleine unspezialisierte Gattungen wie Pseudocyonopsis und Cynelos, deren Gebiss an die heutiger Hunde erinnerte. Eine andere Gattung, Ysengrinia hatte dagegen ein spezialisiertes Fleischfressergebiss. Spätere Amphicyoniden erreichten die Ausmaße von Tigern und dürften ähnlich gejagt haben. Allerdings waren sie wohl langsamer aber ausdauernder. Zeitgleich mit den ersten Amphicyoniden tauchten mit Cephalogale die ersten Bären auf. Diese Gattung scheint sich aus dem früholigozänen Amphicyonodon entwickelt zu haben. Mit verschiedenen Gattungen verbreiteten sich im späten Oligozän auch die Musteliden.[8]
Asien
Das größte Landtier des asiatischen Oligozäns und zugleich das größte Landsäugetier aller Zeiten war Paraceratherium, ein riesiges hornloses, mit den Nashörnern verwandtes Tier von bis zu 5 m Schulterhöhe. Allein der Schädel maß 1,3 m. Die ausgewachsenen Männchen dürften etwa 15 Tonnen gewogen haben. Daneben waren auch die Amynodonten als hornlose Nashörner des Oligozäns typisch für Asien.
Nordamerika

Katzenähnliche Raubtiere des nordamerikanischen Oligozäns waren Hoplophoneus und Nimravus.[13] Charakteristisch für die Raubtierfauna des Oligozäns in Nordamerika waren insbesondere die Hunde. Hesperocyon war eine überlebende Gattung aus dem Eozän und scheint im frühen Oligozän der einzige Canide mit beständiger Fossilüberlieferung gewesen zu sein. Aus dieser Form bildeten sich im Verlauf des Oligozäns die drei Unterfamilien der Hunde, die Hesperocyoninae, die Borophaginae und die noch heute existierenden Canidae. Die Hesperocyoninae sind zunächst durch Mesocyon, Enhydrocyon und Osbornodon repräsentiert, die erste Gattung der Borophaginae ist Otarocyon. Die Canidae sind nur durch ein Unterkieferfragment bekannt, das von einem Vorfahren der Gattung Leptocyon stammen dürfte. Im späteren Verlauf des Oligozäns entstanden größere Hunde aus dem Stamm der Hesperocyoninen, die teilweise die Größe kleiner Wölfe erreichten. Dies waren Sunkahetanka, Philotrox, Enhydrocyon und Paraenhydrocyon. Während die Hesperocyoninae im Oligozän die dominierenden Hunde darstellten, waren die Borophaginae damals noch eher kleinwüchsig. Sie waren etwa durch Archaeocyon, Cynarctoides und Phlaocyon vertreten. Die Canidae waren mit Leptocyon vertreten. Im späteren Oligozän erreichten die Caniden in Nordamerika mit 25 Arten eine nie wieder erreichte Vielfalt. Andere Säuger des späten Oligozäns Nordamerikas waren Protoceras, der oreodontide Leptauchenia, und das Pferd Miohippus.[14]
Afrika

Die Landverbindung zwischen Afrika und Eurasien war im Oligozän noch nicht vollständig ausgebildet. So bewahrte der Kontinent bis zum Miozän eine sehr eigentümliche Säugerfauna mit unverwechselbaren Elementen. Die dominanten größeren Pflanzenfresser des oligozänen Afrikas waren die Schliefer. Sie entwickelten eine große Formenfülle, darunter einige sehr große Formen. Die Paarhufer traten im Oligozän Afrikas noch spärlich auf. Allerdings kamen bereits große Formen wie die Anthracotherien vor. Die Rüsseltiere waren noch deutlich kleiner und erinnerten nur entfernt an heutige Elefanten. Die Rüsseltiere waren damals noch auf ihre Urheimat Afrika beschränkt. Daneben existierte mit den Embrithopoda eine weitere Tiergruppe, die große Pflanzenfresser hervorbrachte. Deren bekanntester Vertreter, das gewaltige Arsinoitherium, erinnert entfernt an ein Nashorn. Allerdings besaß es zwei paarige Hörner auf der Stirn und war nicht mit den Nashörnern verwandt, die zur gleichen Zeit bereits auf anderen Kontinenten lebten. Große Fleischfresser waren die Hyaenodontidae. Allerdings sind sie im Fossilbericht des oligozänen Afrikas relativ selten. Die Primaten waren im Oligozän Afrikas durch Apidium vertreten.[15]
Oligozäne Aufschlüsse und Fundstellen in Deutschland
Marine Fossillagerstätten

Der Doberg bei Bünde ist kein Berg, sondern ein ehemaliges Abbaugebiet von marinen Sedimenten (Kalkmergel) des Oligozän-Meeres. Zahlreiche Fossilfunde können im Dobergmuseum in Bünde besichtigt werden.
- Kasseler Meeressand
Die unter marinen Verhältnissen im Raum Kassel abgelagerten Sedimente des Kasseler Meeressandes gehören mit ihren Aufschlüssen zu den klassischen geologischen Fundorten des Oligozän in Deutschland. Nennenswerte Sammlungsbestände befinden sich noch im Naturkundemuseum im Ottoneum in Kassel. Ein Großteil der alten Sammlung des Naturkundemuseums ist allerdings 1943 bei einem Bombenangriff verbrannt.
Das Mainzer Becken, das als Ausbuchtung (Mainzer Bucht) im Oberrheingraben eine Sedimentfalle wurde, ist die bekannteste Fundregion für marine oligozäne Fossilien in Deutschland. Exponate zeigen u. a. das Heimatmuseum in Alzey, das Naturhistorische Museum in Mainz, das Paläontologische Museum Nierstein und das Museum Wiesbaden. An der Küste im westlichsten Teil des Beckens kam es bei Steinhardt in der Nähe von Bad Sobernheim im Nahetal zur Bildung von fossilführenden Barytkonkretionen, die als Steinhardter Erbsen Bekanntheit erlangten. Eine größere Sammlung von etwa 500 Exponaten besitzt das Heimatmuseum von Bad Sobernheim.

Das nach der mecklenburgischen Kleinstadt Sternberg benannte, meist rötlichbraune, Sternberger Gestein, auch bekannt als Sternberger Kuchen, wurde durch die Gletscher der Eiszeit vom Untergrund Mecklenburgs als Geschiebe an die Oberfläche verlagert. Das Heimatmuseum Sternberg beherbergt eine umfangreiche Sammlung von Gesteinen dieses Nahgeschiebes, in dem bisher etwa 600 Arten von Pflanzen und Tieren nachgewiesen werden konnten.
Limnische Fossillagerstätten
- Kunkskopf-See
Der Kunkskopf-See südlich von Burgbrohl etwa 3,5 km nördlich vom Laacher See war ein Süßwassersee des späten Oligozäns mit oberoligozänen Sedimenten (Schwarzpelite), die Wilhelm Ahrens 1929 bekannt machte. Das Landesamt für Denkmalpflege Rheinland-Pfalz führte in den Jahren 1998 und 1999 paläontologische Grabungen an dieser Fundstelle durch. Am Fundort konnten bisher fossile Blätter, Samen, Frösche, Fische, Vogelfedern und Insekten nachgewiesen werden.
- Rotter See
Der Rotter See (Fossillagerstätte Rott) ist eine wegen des Reichtums und der außergewöhnlich guten Erhaltung von fossilen Pflanzen und Tieren bereits seit Anfang des 19. Jahrhunderts auch außerhalb von Deutschland bekannte limnische Lagerstätte von Fossilien aus dem Oligozän beim heutigen Ortsteil Rott der Stadt Hennef (Sieg). Bis zum Jahr 1997 wurden von diesem Fundort unter anderem 250 Pflanzenarten, 630 Insektenarten und 20 Amphibien- und Reptilienarten in etwa 470 Publikationen, die sich mit der Fossillagerstätte oder den einzelnen Fossilien selbst beschäftigen, beschrieben.
- Stöffel-See
Der Stöffel-See bei Enspel im Westerwald war nach den bisherigen Erkenntnissen ein dem Spät-Oligozän zuzuordnender Süßwassersee, dessen feine Fundschichten von einer Basaltdecke geschützt wurden und von 1990 bis 2015 durch gezielte Grabungen untersucht wurden. Bisher wurden ca. 36.000 unterschiedlichste Fossilien geborgen, präpariert und inventarisiert. Die 1992 gefundene Enspelmaus Eomys quercyi Storch, Engesser & Wuttke, 1996, ist der bisher älteste Beleg für den Gleitflug bei Nagetieren. Das Fossil war zuvor nur durch isolierte Zahnfunde aus Frankreich bekannt. Vor diesem Hintergrund wurde der Tertiär- und Industrie-Erlebnispark Stöffel als Freilichtmuseum und Bestandteil des Geoparks Westerwald-Lahn-Taunus eingerichtet.
- Siebloser See
Der Siebloser See (Fossillagerstätte Sieblos) war ein waldumstandener Süßwassersee mit einer vegetationsreichen Uferzone, dessen Sedimente dagegen mit einem Alter von etwa 33 Millionen Jahren (Frühes Oligozän) einige Millionen Jahre älter sind und die Fauna und Flora dieses Fundortes somit entwicklungsgeschichtlich noch vor den Fossilien vom Stöffel-See wie auch denen vom Rotter See einzuordnen ist. Die Sammlung vom Sieblos-Museum in Poppenhausen (Wasserkuppe) umfasst etwa 2600 Exponate und kann nach vorheriger Anmeldung auch im Archiv direkt begutachtet werden. Die ehemalige Sammlung Ernst Hassencamp ging an das Geologisch-Paläontologische Institut in Würzburg und ist heute Teil der Bayerische Staatssammlung für Paläontologie und Geologie.
- Münzenberger Blättersandstein
Der Münzenberger Blättersandstein in der Wetterau (Hessen) im Steinbruch am nordöstlichen Ortsrand von Münzenberg schließt die jüngsten (= spätesten) oligozänen Sedimente in rosa bis kräftig rotbraunen Farben, überlagert von mächtigen Konglomeraten des Miozäns, auf. Die Sammlung der Paläobotanischen Sektion des Forschungsinstitutes Senckenberg am Standort Frankfurt umfasst eine größere Anzahl von Pflanzenresten aus dem „Münzenberger Blättersandstein oder Blätterquarzit“.
- Rockenberg in der Wetterau
.jpg.webp)
In einem Naturschutzgebiet in der Höll am nördlichen Ortsrand von Rockenberg sind oligozäne Sande aufgeschlossen, die teilweise durch Kieselsäureausfällungen zu stabilen Sedimentquarziten erhärtet sind. Eindrucksvolle Sandrosen aus diesen Sanden, die in der Literatur auch als Rockenberger Röschen bekannt sind, zeigt das Sandrosenmuseum im Untertor in Büdingen.
Weblinks
- Deutsche Stratigraphische Kommission, Manfred Menning (Hrsg.): Stratigraphische Tabelle von Deutschland 2002. Potsdam 2002, ISBN 3-00-010197-7 (1 Blatt, Stratigraphie.de [PDF; 6,6 MB]).
- Kommission für die paläontologische und stratigraphische Erforschung Österreichs der Österreichischen Akademie der Wissenschaften (Hrsg.): Die Stratigraphische Tabelle von Österreich (sedimentäre Schichtfolgen). Wien 2004 (PDF; 376 kB)
- International Chronostratigraphic Chart 2020/03
- Weltkarte für das Oligozän (Blakey)
- Oligozäne Mikrofossilien: mehr als 300 Bilder von Foraminiferen
{kind=link}
Literatur
- Peter Prinz-Grimm: Subtropisches Leben in einem langen, schmalen Meer. 18 Tertiär: Oligozän. In: Peter Rothe, Volker Storch und Claudia von See (Hrsg.): Lebensspuren im Stein. Ausflüge in die Erdgeschichte Mitteleuropas. Wiley/VCH 2014, S. 199–207
- Fritz F. Steininger, M. P. Aubry, W. A. Berggren, M. Biolzi, A. M. Borsetti, J. E. Cartlidge, F. Cati, R. Corfield, R. Gelati, S. Iaccarino, C. Napoleone, F. Ottner, F. Rögl, R. Roetzel, S. Spezzaferri, F. Tateo, G. Villa, D. Zevenboom: The Global Stratotype Section and Point (GSSP) for the base of the Neogene. In: Episodes. Band 20(1), 1997, ISSN 0705-3797, S. 23–28.
- Isabella Premoli-Silva, David G. Jenkins: Decision on the Eocene-Oligocene boundary stratotype. In: Episodes. Band 16(3), 1993, ISSN 0705-3797, S. 379–382.
Einzelnachweise
- Ernst Beyrich: Über die Stellung der hessischen Tertiärbildungen. In: Bericht über die zur Bekanntmachung geeigneten Verhandlungen der Königlich Preussischen Akademie der Wissenschaften zu Berlin, Berlin 1854, S. 664 (Digitalisat)
- Mark Pagani, Matthew Huber, Zhonghui Liu, Steven M. Bohaty, Jorijntje Henderiks, Willem Sijp, Srinath Krishnan, Robert M. DeConton: The Role of Carbon Dioxide During the Onset of Antarctic Glaciation. (PDF) In: Science. 334, Nr. 6060, Dezember 2011, S. 1261–1264. doi:10.1126/science.1203909.
- Simone Galeotti, Robert DeConto, Timothy Naish, Paolo Stocchi, Fabio Florindo, Mark Pagani, Peter Barrett, Steven M. Bohaty, Luca Lanci, David Pollard, Sonia Sandroni, Franco M. Talarico, James C. Zachos: Antarctic Ice Sheet variability across the Eocene-Oligocene boundary climate transition. (PDF) In: Science. 352, Nr. 6281, April 2016, S. 76–80. doi:10.1126/science.aab0669.
- James Hansen, Makiko Sato, Gary Russell, Pushker Kharecha: Climate sensitivity, sea level and atmospheric carbon dioxide. (PDF) In: Philosophical Transactions A (The Royal Society). 371, Nr. 2001, Oktober 2013. doi:10.1098/rsta.2012.0294.
- James S. Eldrett, Ian C. Harding, Paul A. Wilson, Emily Butler, Andrew P. Roberts: Continental ice in Greenland during the Eocene and Oligocene. (PDF) In: Nature. 446, März 2007, S. 176–179. doi:10.1038/nature05591.
- Aradhna Tripati, Dennis Darby: Evidence for ephemeral middle Eocene to early Oligocene Greenland glacial ice and pan-Arctic sea ice. (PDF) In: Nature Communications. 9, März 2018. doi:10.1038/s41467-018-03180-5.
- Michael Stärz, Wilfried Jokat, Gregor Knorr, Gerrit Lohmann: Threshold in North Atlantic-Arctic Ocean circulation controlled by the subsidence of the Greenland-Scotland Ridge. In: Nature Communications (online). 8, Juni 2017. doi:10.1038/ncomms15681.
- Jordi Augusti: Mammoths, Sabertooths and Hominids 65 Million Years of Mammalian Evolution in Europe. Columbia University Press, 2002, ISBN 0-231-11640-3.
- Stefan M. Schmid, Daniel Bernoulli, Bernhard Fügenschuh, Liviu Matenco, Senecio Schefer, Ralf Schuster, Matthias Tischler, Kamil Ustaszewski: The Alpine-Carpathian-Dinaridic orogenic system: correlation and evolution of tectonic units. (PDF) In: Swiss Journal of Geosciences. 101, März 2008, S. 139–183. doi:10.1007/s00015-008-1247-3.
- Michaela Grein, Christoph Oehm, Wilfried Konrad, Torsten Utescher, Lutz Kunzmann, Anita Roth-Nebelsick: Atmospheric CO2 from the late Oligocene to early Miocene based on photosynthesis data and fossil leaf characteristics. (PDF) In: Palaeogeography, Palaeoclimatology, Palaeoecology. 374, März 2013, S. 41–51. doi:10.1016/j.palaeo.2012.12.025.
- D. W. Hauptvogel, S. F. Pekar, V. Pinca: Evidence for a heavily glaciated Antarctica during the late Oligocene “warming” (27.8–24.5 Ma): Stable isotope records from ODP Site 690. In: Paleoceanography and Paleoclimatology. 32, Nr. 4, April 2017, S. 384–396. doi:10.1002/2016PA002972.
- Helen M. Beddow, Diederik Liebrand, Appy Sluijs, Bridget S. Wade, Lucas J. Louren: Global change across the Oligocene-Miocene transition: High-resolution stable isotope records from IODP Site U1334 (equatorial Pacific Ocean). (PDF) In: Paleoceanography (AGU Publications). 31, Nr. 1, Januar 2016, S. 81–97. doi:10.1002/2015PA002820.
- A. Turner, M. Antón: The big cats and their fossil relatives. Columbia University Press, New York 1997, ISBN 0-231-10229-1.
- Xiaoming Wang, Richard H. Tedford, Mauricio Antón: Dogs, their fossil relatives & evolutionary history. Columbia University Press, New York 2008, ISBN 978-0-231-13528-3.
- Alan Turner, Mauricio Anton: Evolving Eden: An Illustrated Guide to the Evolution of the African Large Mammal Fauna. Columbia University Press, 2004, ISBN 0-231-11944-5.