Quartäre Aussterbewelle
Die quartäre Aussterbewelle war ein Massenaussterben der letzten Kaltzeit, bei dem zahlreiche Tierarten auf verschiedenen Kontinenten verschwanden. Der Prozess, bei dem vor allem große und sehr große Arten der eiszeitlichen Megafauna betroffen waren, war auf das Quartär-Zeitalter beschränkt und erreichte seinen Höhepunkt am Übergang vom Pleistozän zum Holozän (der Nacheiszeit). Die wissenschaftliche Diskussion des Phänomens ist bis heute in zwei Lager geteilt, da einerseits menschliche Einflüsse („Overkill“), andererseits Klimaveränderungen als Hauptursache gesehen werden. Auch eine Kombination aus beiden Faktoren könnte das Massenaussterben hervorgerufen haben.

Die ausgestorbenen Arten
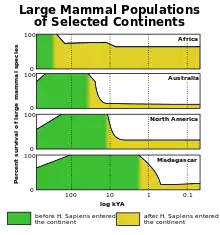
Bis zum Ende des Pleistozäns waren alle Kontinente von einer reichen Großtierfauna bevölkert, vergleichbar mit heutigen afrikanischen Lebensräumen. Während der letzten Kaltzeit (in Mitteleuropa Würmeiszeit in Süddeutschland bzw. Weichseleiszeit im Norden) und besonders an ihrem Ende verschwanden zahlreiche Großtierarten in einem relativ kurzen Zeitraum. Nur in Afrika und zum Teil in Südasien überlebten einige Tiergiganten wie Nashörner und Elefanten und auch wesentlich mehr andere Großsäugerarten als auf anderen Kontinenten. Mit Ausnahme Afrikas und des südlichen Asiens starben weltweit alle Arten mit über 1000 Kilogramm Gewicht und 80 % aller Arten mit 100 bis 1000 Kilogramm Gewicht, meist zeitgleich mit dem allmählichen Vordringen des modernen Menschen, aus. In einigen Regionen wie dem Norden Eurasiens oder in Amerika korreliert das Massensterben zusätzlich relativ gut mit klimatischen Veränderungen. Der Anteil ausgestorbener Arten unter 45 kg Gesamtgewicht ist verschwindend gering.
Afrika und Südasien
Die Tropen der Alten Welt stellen heute die einzigen Regionen der Erde dar, die über eine ausgeprägte Großtierfauna verfügen. Nur hier existieren noch heute Arten mit über 1000 kg Körpergewicht. Dazu zählen der Afrikanische Elefant, der Asiatische Elefant sowie das Flusspferd und verschiedene Nashörner. Als Grund dafür wird gelegentlich die Tatsache angeführt, dass der Mensch sich in Afrika entwickelte und die südlichen Teile Asiens bereits seit dem frühen Pleistozän erreichte. Aufgrund ihrer gemeinsamen Evolution hätten die Tiere dieser Regionen demzufolge die Gelegenheit gehabt, Fluchtreflexe zu entwickeln und sich so an die Bejagung durch den Menschen anzupassen. Allerdings verlor auch diese Region insbesondere im frühen oder mittleren Pleistozän relativ viele Großtierformen, ohne dass sie von Nachfolgearten vertreten worden wären. Daher wurde vermutet, dass bereits die frühen Hominiden des frühen oder mittleren Pleistozäns einige Arten der altweltlichen Tropen ausgerottet haben könnten.[1] Großsäuger, die im frühen und mittleren Pleistozän in Afrika und Südasien ausstarben, umfassen etwa die Dinotherien, die Chalicotherien, die Rindergiraffen und verschiedene Säbelzahnkatzen (Megantereon, Homotherium, Dinofelis). Das Verschwinden der Säbelzahnkatzen vor etwa 1,5 Millionen Jahren in Afrika korreliert stark mit dem erstmaligen Auftreten der Acheulkultur, so dass bisweilen ein Zusammenhang vermutet wird.[2] Die Verluste der afrikanischen Fauna im Spätpleistozän sind dagegen gering. Dazu zählen eine Riesenkuhantilope (Megalotragus) und der Riesenbüffel (Pelorovis (Syncerus) antiquus).
Australien
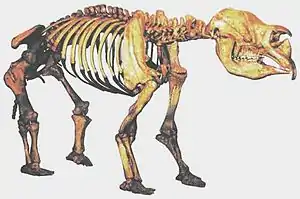
Der australische Kontinent einschließlich Tasmaniens und Neuguineas verlor im späten Pleistozän alle landlebenden Säugetier-, Vogel- und Reptilienarten mit einem Gewicht von über 100 kg. Darüber hinaus verschwanden alle Gattungen, die Arten zwischen 45 und 100 kg beinhalteten, mit Ausnahme einer einzigen. Diese einzige Großtiergattung, die in Australien das Pleistozän überlebte, war die Kängurugattung Macropus. Zu den Großtierarten Australiens, die verschwanden, zählten Diprotodon und Zygomaturus (nashorngroße Beuteltiere), der Beuteltapir Palorchestes, der Beutellöwe (Thylacoleo carnifex), ein Riesenmoschusrattenkänguru, riesige Wombats (Phascolonus) und bis zu drei Meter hohe Riesenkänguruhs (Gattung Procoptodon, Simosthenurus, Sthenurus, Protemnodon). Dazu kam der große, flugunfähige Vogel Genyornis und große Reptilien, wie der riesige Waran Megalania. Alle diese Arten scheinen vor etwa 40.000 bis 50.000 Jahren ausgestorben zu sein, was zeitlich sehr stark mit dem frühesten Auftreten des Menschen dort korreliert.[3] Auch wurden in einem der bisher ältesten bekannten menschlichen Lagerplätze Australiens Knochen eines Diprotodons gefunden.[4]
Das nördliche Eurasien

In Eurasien erstreckte sich dieser Vorgang über einen längeren Zeitraum, von vor 50.000 bis vor 12.000 Jahren, und erreichte mit dem Ende des Pleistozäns seinen Höhepunkt. Zu den in Europa bis spätestens vor etwa 12.000 Jahren ausgestorbenen Arten zählen Wollhaarmammut (Mammuthus primigenius), Wollnashorn (Coelodonta antiquitatis), Riesenhirsch (Megaloceros giganteus), Steppenwisent (Bos priscus), Höhlenlöwe (Panthera spelaea) und Höhlenhyäne (Crocuta crocuta spelaea).
Einige während der letzten Kaltzeit (Würmeiszeit bzw. Weichseleiszeit) ausgestorbene Arten verschwanden bereits vor dem Ende des Pleistozäns. Dies betrifft vor allem wärmeliebende Arten, wie den Europäischen Waldelefanten (Palaeoloxodon antiquus), das Waldnashorn (Stephanorhinus kirchbergensis) und das Steppennashorn (Stephanorhinus hemitoechus), die während der frühen Würmeiszeit vor etwa 100.000 Jahren zunächst aus Mittel- nach Südeuropa abgedrängt wurden und dort im Verlauf der nächsten Jahrzehntausende ausstarben. Der europäische Höhlenbär (Ursus spelaeus) verschwand nach heutigem Kenntnisstand vor dem letzten Kältemaximum der Würmeiszeit, während der archäologischen Kultur des Gravettien vor etwa 27.000 Jahren.[5][6] Etwa zur selben Zeit ist auch letztmals die Säbelzahnkatze (Homotherium) in Europa nachgewiesen.[7][8]
Nordamerika

In Nordamerika lag das Aussterben in einem engen Zeitrahmen (vor rund 12.000 Jahren) und erfolgte sehr plötzlich. Dort verschwanden unter anderem das Wollhaarmammut, das Präriemammut, das Amerikanische Mastodon, alle Kamele (Camelops, Hemiauchenia, Palaeolama) und Pferde, der Helm-Moschusochse, der Buschochse, sowie die Säbelzahnkatzen (Smilodon, Homotherium), vier Riesenfaultiere (Eremotherium, Megalonyx, Paramylodon, Nothroteriops) und Riesengürteltiere (Glyptotherium, Holmesina). Dazu verschwanden der Amerikanische Löwe, der Amerikanische Gepard, der große Wolf Aenocyon dirus, der Kurzschnauzenbär, Florida-Brillenbär, Wasserschweine und Tapire sowie verschiedene Hirsch-, Gabelbock- und Pekariarten. Mindestens 17 Gattungen der ausgestorbenen Megafauna Amerikas verschwanden in einem sehr engen Zeitfenster vor 11.400 bis 10.800 Radiokohlenstoffjahren.[9] Das Aussterben auf dem amerikanischen Kontinent korreliert mit dem Klimawandel am Ende des Pleistozäns sowie mit dem erstmaligen Auftreten des Menschen in der neuen Welt. Die größte Tierart, die das Massensterben überlebte, ist der Amerikanische Bison.
Südamerika
In Südamerika scheinen die meisten Arten um dieselbe Zeit oder wenig später als in Nordamerika ausgestorben zu sein. Hier ist die Fossildokumentation diesbezüglich allerdings weniger gesichert.[9] Südamerika verlor während dieses Massensterbens alle seine Rüsseltiere (Cuvieronius, Notiomastodon), sämtliche Riesenfaultiere (z. B. Megatherium, Glossotherium, Mylodon, Scelidotherium), Glyptodonten, Riesengürteltiere (Pampatherien) und Pferde (Equus, Hippidion), sowie die eigenartigen Macrauchenien und Toxodonten (Toxodon, Mixotoxodon). Dazu verschwanden um dieselbe Zeit das Flachkopfpekari, verschiedene Hirsche und Kamele (Palaeolama, Hemiauchenia). Unter den Raubtieren sind am Beginn des Holozäns die Säbelzahnkatze Smilodon, verschiedene Wildhunde (Aenocyon dirus, Protocyon) und Bären verschwunden. Insgesamt starben am Beginn des Holozäns 80 % aller Großsäuger über 44 kg sowie alle Arten mit einem Körpergewicht von über 300 kg in Südamerika aus.[10] Das größte überlebende Säugetier des Südamerikanischen Kontinents ist der Mittelamerikanische Tapir.

Abgelegene Inseln
Das Wollhaarmammut, das auf dem eurasischen und nordamerikanischen Festland am Ende des Pleistozäns verschwand, überlebte auf einigen abgelegenen Arktischen Inseln bis weit ins Holozän. Am längsten auf der sibirischen Wrangelinsel, wo eine Zwergform des Mammuts erst vor etwa 4000 Jahren ausstarb.[11] Ähnliches gilt für die Bodenfaultiere der karibischen Inseln. Während diese Tiere auf dem Festland am Ende des Pleistozäns ausstarben, überlebten größere Faultiere (Megalocnus, Parocnus) auf den Karibischen Inseln mindestens bis vor rund 5000 Jahren.[12] Auf den Mittelmeerinseln verschwanden im Verlauf des Holozäns verschiedene Flusspferde und Zwergelefanten sowie die Höhlenziege. Erst vor rund 2.000 Jahren starben die meisten Großtiere Madagaskars aus. Dazu zählten Madagassische Flusspferde, die Riesenfossa (Cryptoprocta spelea), riesige Lemuren wie Archaeoindris, Megaladapis oder Palaeopropithecus sowie Elefantenvögel. Madagaskar wurde vor rund 2.300 Jahren zum ersten Mal durch Menschen besiedelt.[13] In Neuseeland verschwanden vor wenigen Jahrhunderten unter anderem die Moas (Dinornis, Pachyornis, und Euryapteryx) und der Haastadler. Insgesamt korreliert das Verschwinden vieler Arten, insbesondere der größeren, auf abgelegenen Inseln mit dem jeweiligen erstmaligen Erscheinen des Menschen.
Der Mensch als Verursacher
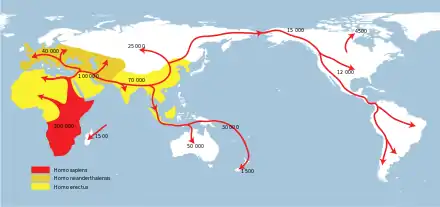
Seit langem wird für das Aussterben der eiszeitlichen Megafaunen der Mensch als Verursacher diskutiert. Für ein Aussterben dieser Arten durch menschliche Überjagung sprechen die Tatsachen, dass der Zeitpunkt des Aussterbens auffällig mit der weltweiten Ausbreitung des Menschen korreliert und dass bei keiner der früheren Aussterbephasen eine derartige Einschränkung hinsichtlich der Größe beobachtet werden konnte. Unter dem Namen „Overkill-Hypothese“ wurde das Konzept von Paul S. Martin in den 1960er Jahren ausgearbeitet.[14][15][16][17] Martin stellte sich das Aussterben in sehr kurzer Zeit vor, er sprach vom Over-Kill in Form eines Blitzkriegs. Heute sieht man Aussterbephasen von Jahrhunderten bis Jahrtausenden. Umstritten ist allerdings bis heute die Gewichtung der Wirkfaktoren Klimaveränderung und Überjagung.[18][19][20][21]

Besonders die Tatsache, dass die Aussterbewelle in Australien vor etwa 50.000 Jahren sehr gut mit dem Auftreten des Menschen, aber praktisch nicht mit auffälligen Klimawechseln korreliert, spricht dafür, dass menschliche Einflüsse die Hauptursache für das Verschwinden der Megafauna waren.[22] Auffälligerweise scheinen die letzten Vertreter der australischen Megafauna auf der Insel Tasmanien überlebt zu haben, die offenbar erst einige 1000 Jahre nach dem australischen Festland durch Menschen besiedelt wurde. Dort existierte das letzte der riesigen pleistozänen Kängurus Protemnodon anak bis vor 40.000 Jahren.[23]
Vertreter der Überjagungshypothese führen auch einen analogen Vorgang auf Inseln, die erst später besiedelt wurden, an. So sind auf Madagaskar, wo erst seit rund 2300 Jahren Menschen leben, in den darauffolgenden Jahrhunderten unter anderem die dortigen Flusspferde, die Elefantenvögel, zwei Arten endemischer Erdferkel, eine Krokodilart, die Riesenfossas und zahlreiche große Primatenarten, darunter die Riesenlemuren Megaladapis, verschwunden. In Neuseeland verschwanden neben den Moas auch viele andere flugunfähige Vögel und der Riesenadler Harpagornis bald nach der Besiedlung durch die Māori um etwa 800.[24]
Erörterung der Überjagungshypothese
Gegner der Überjagungshypothese verweisen auf die primitiven Jagdmethoden der frühen Menschen, die keinen so großen Einfluss auf die Populationsgröße haben können, und verweisen auf Afrika, wo es schon viel länger Menschen gegeben hat und wo es zu keinem nennenswerten Massenaussterben gekommen ist. Allerdings zeigen zum einen Modellrechnungen, dass gerade große und sich langsam reproduzierende Tiere selbst bei geringer Bejagung auf lange Sicht vom Aussterben bedroht sind, dass es also leicht zu einer Übernutzung des Naturangebots kommen kann. Zum anderen haben Versuche mit nachgebauten Speeren mit Feuersteinspitzen gezeigt, dass selbst so große Tiere wie Elefanten mit ihnen getötet werden können, die steinzeitlichen Jäger also durchaus in der Lage waren, praktisch jedes Tier zu erlegen. Dies gilt insbesondere dann, wenn die Tiere keinerlei Fluchtreflexe gegenüber dem Menschen besitzen.
Unklar ist, wieso einige große Arten, die in Größe und Lebensweise den ausgestorbenen Arten ähnlich waren und auch nachgewiesenermaßen Jagdbeute des Menschen waren, bis in die Gegenwart beziehungsweise in die historische Neuzeit, auch in Mitteleuropa, überlebten, zum Beispiel der Elch (entspricht der Größe von Megaloceros), die großen Rinderarten wie Bison, Wisent oder Ur (die der Größe des ausgestorbenen Buschochsen (Euceratherium) entsprachen).
Ein Hauptkritikpunkt der Gegner der Hypothese besteht außerdem darin, dass in Amerika nur relativ wenige Jagdplätze mit ausgestorbenen Arten bekannt sind, während man solche Plätze aus Eurasien (Nashörner, Pferde etc.) in großer Zahl kennt. Die Befürworter der Überjagungshypothese erklären diese Tatsache damit, dass die nordamerikanischen Faunen innerhalb nur weniger hundert Jahre ausstarben und somit kaum Potential für Fossilnachweise blieb.[9]
Einige Forscher nehmen an, dass sowohl klimatische als auch menschliche Einflüsse für das Aussterben der Großtierarten am Ende des Pleistozäns verantwortlich waren und erst im Zusammenspiel so viele Arten auslöschten. Demnach hätten die Steinzeitmenschen dann nur die ohnehin schon geschwächten Populationen ausgelöscht. Befürworter der Overkill-Hypothese bemerken hierzu aber, dass die meisten dieser Arten sich ohne den Einfluss des Menschen wohl wieder erholt hätten, wie nach den vorherigen Klimawechseln des Eiszeitalters auch. Im Grunde wäre also auch bei dieser abgeschwächten Form der Overkill-Hypothese der Mensch wohl der ausschlaggebende Faktor für das Massensterben gewesen.[25]

Auftreten auf Inseln und Kontinenten
Kaum in Frage gestellt wird, dass der Untergang endemischer Tierarten auf einigen abgelegenen Inseln wie Madagaskar (ca. 500 n. Chr.), Mauritius (ca. 1650 n. Chr.), Neuseeland (ca. 1300 n. Chr.) und Guam (ca. 1950 n. Chr.) in historischen Zeiten durch den Einfluss des Menschen verursacht wurde. Auch Inseln wie Tilos und Zypern oder die Wrangelinsel, wo die letzten, verzwergten Altelefanten bzw. Wollhaarmammute lebten, ereilte dieses Schicksal, als die ersten Menschen ankamen. Allerdings liegen weder für die Altelefanten von Tilos (um 1300 v. Chr.) und Zypern (um 9.500 v. Chr.) noch für die Mammute der Wrangelinsel (um 1700 v. Chr.) Belege eines direkten Einflusses des Menschen vor. Zumindest für das Wollhaarmammut der Wrangelinsel zeigen DNA-Befunde, dass die dortige Population bereits im Schwinden war und infolge von Inzucht unter zahlreichen Mutationen bis hin zu einer verminderten Fruchtbarkeit der Bullen litt.[26][27] Auf zahlreichen Inseln betraf das Aussterben sämtliche leicht zu erlegenden Riesentiere. Andere (kleinere) Tierarten starben durch die folgend aufkommende landwirtschaftliche Nutzung aus. Teilweise wurden sie auch durch die mitgebrachten Haustiere oder eingeschleppte Tiere wie Ratten oder Schlangen ausgerottet.
Verfechter der Überjagungshypothese stellen die Besiedelung dieser Inseln in eine Reihe mit der Besiedelung der Kontinente Amerika und Australien. Die Gegner der Überjagungsshypothese betonen, dass das Massensterben am Ende der Eiszeit nicht mit dem Aussterben von Tieren auf diesen Inseln zu vergleichen sei. Das Aussterben auf den Inseln wäre vermutlich vor allem dadurch bedingt gewesen, dass diese Tiere keine Fluchtreflexe entwickelten, da sie oft wenige natürliche Feinde gehabt oder Menschen nicht als solche erkannt hätten. Zudem hätten sie aus ihrem Lebensraum nicht fliehen können und wären für die jagenden Menschen somit eine leichte und ungefährliche Beute gewesen. Dies auf die Vorgänge am Ende des Pleistozäns zu übertragen sei jedoch schwierig. Inselpopulationen wären wegen ihres meist geringeren Verbreitungsgebiets und ihrer geringeren Populationsgröße anfälliger für Katastrophenereignisse.
Das Klima als Verursacher
Die Klimahypothese ist neben der Überjagungshypothese die mit Abstand am häufigsten genannte Erklärung des Massensterbens am Ende des Pleistozäns. Auf dem amerikanischen Doppelkontinent fallen das Auftreten des Menschen und die Klimaveränderungen am Ende des Pleistozäns in denselben Zeitraum und sind somit nur schwer zu trennen. Das Aussterben der Großtierfauna Australiens vor rund 45.000 Jahren korreliert überhaupt nicht mit bekannten Klimaschwankungen, wohl aber mit dem erstmaligen Auftreten des Menschen. Allerdings sind die Daten für die australischen Vorgänge weniger präzise, da sie länger zurückliegen. Problematisch ist weiter, dass es während des gesamten Pleistozäns zahlreiche Klimaschwankungen gab, die nicht zu Massenaussterben führten. Daten aus Eisbohrkernen belegen, dass frühere Klimaschwankungen jenen am Ende des Pleistozäns, die für das Aussterben der Megafauna verantwortlich gemacht werden, weder an Stärke noch Geschwindigkeit nachstanden.[9]
Clovis-Kometen-Hypothese
In einer Publikation von 2007 wurde vermutet, dass ein Asteroideneinschlag für das Artensterben insbesondere in Amerika verantwortlich sein soll. Die Hypothese wurde aufgestellt von James Kennett (University of California, Santa Barbara), Richard Firestone (Lawrence Berkeley National Laboratory) sowie Douglas Kennett und Jon Erlandson (University of Oregon) und besagt, dass vor ca. 12.900 Jahren im heutigen Kanada ein Asteroid eingeschlagen sei. Dieser Clovis-Komet sei der Grund für das Massensterben der Eiszeit-Megafauna und soll auch der zeitgleich existierenden steinzeitlichen Clovis-Kultur ein Ende bereitet haben.[28] Die Befunde konnten von einer unabhängigen Forschergruppe jedoch nicht bestätigt werden.[29] Eine im August 2008 veröffentlichte Untersuchung der demographischen Entwicklung der Paläoindianer für den fraglichen Zeitraum ergab darüber hinaus keinen Hinweis auf den in der Hypothese genannten Bevölkerungsrückgang.[30]
Im Januar 2009 berichtete das Magazin Science von der elektronenmikroskopischen Bestätigung der angeblichen Existenz von Nanodiamanten zur geologischen Zeit des fraglichen Ereignisses,[31] was auch in der International Herald Tribune rezipiert wurde[32]; laut Douglas Kennett sei dies der Beweis einer Kollision eines oder mehrerer Kometen mit der Erde am Ende des Pleistozäns, was gravierende Auswirkungen auf die Flora und Fauna Nordamerikas hatte.[33] Allerdings wurde diese Publikation von anderen Wissenschaftlern stark kritisiert.[34] Die Schlussfolgerungen konnten von anderer Seite nicht bestätigt werden und weitere Studien ergaben keinen Nachweis von Nanodiamanten[35] Im April 2010 erklärte Bill Napier (Professor der Astrobiologie an der Cardiff University) in „Monthly Notices“ (einem Journal der Royal Astronomical Society), dass die Erde mit Trümmern eines gewaltigen Kometen mit einer ursprünglichen Größe von 50 bis 100 Kilometern im Durchmesser, der vor 30.000 Jahren in unser Sonnensystem eingedrungen und dann zerbrochen sei, kollidiert sei, und dass der Einschlag der Trümmer einen Feuersturm ausgelöst habe, der die Atmosphäre mit Asche und Staub verdeckt habe und Wärme und Sonneneinstrahlung erheblich reduziert habe und eine plötzliche „Mini-Eiszeit“ der Jüngeren Dryas ausgelöst habe.[36] Unbefriedigend erscheint an dem angeblichen Kometeneinschlag als Erklärung für das Massensterben die Tatsache, dass den eiszeitlichen Tieren Nordamerikas, die vorher wesentlich kühlere Epochen überlebt haben, die mäßige Abkühlung der Jüngeren Dryas zum Verhängnis geworden sein soll.[9] Ein zusammenfassender Artikel von 2011 über die Impakt-Hypothese in Earth Science Reviews kam zu dem Schluss, dass keine Beweise für einen Impakt existieren (Proben mit terrestrischem Ursprung, Fehlinterpretation von Messergebnissen).[37]
Hingegen präsentierten in den darauffolgenden Jahren mehrere Studien eine Reihe von Indizien, darunter eine in Südafrika nachgewiesene Platinanomalie sowie in Nordsyrien entdeckte Schmelzgläser, die nach Ansicht der Autoren für die Impakthypothese sprechen.[38][39][40]
Weitere Erklärungshypothesen
Eine neuere Hypothese US-amerikanischer Wissenschaftler besagt, dass nicht die Bejagung allein, sondern durch den Menschen und seine Haustiere bzw. Kulturfolger eingeschleppte Seuchen für das Aussterben der Großtiere ausschlaggebend gewesen sei. Diese Tiere hätten ihr Immunsystem im Gegensatz zu den kleineren mit rascheren Generationsfolgen nicht rechtzeitig an die Krankheitserreger anpassen können.[41] Allerdings erscheint fraglich, ob Seuchen so viele verschiedene Tierarten ausrotten konnten (auch da viele ihrer oft nur unwesentlich kleineren Artgenossen überlebten), vor allem, wenn man bedenkt, dass Nordamerika immer wieder im Flora- und Faunenaustausch mit Eurasien stand und die meisten Großtiere (Rüsseltiere, Hunde, Katzen, Bisons, Bären, Kamele und Pferde) auch von dort stammten.
Ebenfalls eine Hypothese neueren Ursprungs ist die von LaViolette, dass es etwa im Jahre 12.837 BP zu einer Sonnen-Protuberanz bzw. einem koronalen Massenauswurf gekommen sei, zu der man im venezolanischen Cariaco-Becken entsprechende C14-Spuren gefunden hat. Diese Protuberanz sei etwa 125 mal so groß wie die 1956 bislang größte jemals direkt gemessene Protuberanz gewesen. Sie führte auf dem Boden zu Strahlungsdosen von bis zu 3 Sievert innerhalb der ersten drei Tage sowie zu einer jahrelangen Zerstörung der Ozonschicht, die somit weitere Strahlung auf Flora und Fauna durchließ. Auch diese Hypothese könnte somit den Teil der quartären Aussterbewelle, der sich vor rund 15.000 Jahren abspielte, erklären.[42] Dem entgegen steht, dass das Aussterben weltweit nicht zeitlich synchron ablief.[43]
In nördlichen Regionen könnte auch eine umfassende Vernässung und Versumpfung der Landschaft durch das Schmelzwasser der tauenden Gletscher und ein damit einhergehender Rückgang von Weidegebieten eine entscheidende Rolle beim Aussterben der eiszeitlichen Großsäuger gespielt haben.[44]
Literatur
- Göran Burenhult: Die ersten Menschen. Weltbild Verlag, 2000, ISBN 3-8289-0741-5.
- Peter D. Ward: Ausgerottet oder ausgestorben? Warum die Mammuts die Eiszeit nicht überleben konnten. Birkhäuser Verlag, 2001, ISBN 3-7643-5915-3.
- Ted Oakes: Land of Lost Monsters: Man Against Beast. The Prehistoric Battle for the Planet. 2003.
- Alan Weisman: Die Welt ohne uns. Reise über eine unbevölkerte Erde. Piper, München 2007, ISBN 978-3-492-05132-3 (deutsche Ausgabe).
- Bernisches Historisches Museum (Hrsg.): Jagen und Sammeln. Festschrift für Hans-Georg Bandi zum 65. Geburtstag (3. September 1985), gewidmet von den Mitarbeitern des bernischen historischen Museums, des Seminars für Urgeschichte der Universität Bern, sowie von Freunden und Fachkollegen im in- und Ausland, von Bernisches Historisches Museum, Hans-Georg Bandi, Rudolf Fellmann, Georg Germann, Karl Zimmermann; Veröffentlicht von Stämpfli & Cie
Einzelnachweise
- Martin, P., S., Wright, H., E., Jr. (1967). Pleistocene Extinctions. The Search for a Cause. New Haven and London, Yale University Press.
- Arribas, A., Palmqvist, P., 1999. On the Ecological Connection Between Sabre-tooths and Hominids: Faunal Dispersal Events in the Lower Pleistocene and a Review of the Evidence for the First Human Arrival in Europe. Journal of Archaeological Science 26, 571–585.
- Roberts, R. G., T. F. Flannery, L. A. Ayliffe, H. Yoshida, J. M. Olley, G. J. Prideaux, G. M. Laslett, A. Baynes, M. A. Smith, R. Jones, and B. L. Smith. 2001. New ages for the last Australian megafauna: continent-wide extinction about 46,000 years ago. Science 292: 1888–1892.
- Dani Cooper: Oldest known evidence of Aboriginal settlement in arid Australia found in Flinders Ranges rock shelter. ABC News, 3. November 2016, abgerufen am 3. November 2016 (englisch).
- Martina Pacher & Anthony J. Stuart: Extinction chronology and palaeobiology of the cave bear (Ursus spelaeus), 2009. Boreas, Volume 38 Issue 2, Pages 189–206. doi:10.1111/j.1502-3885.2008.00071.x
- Frühes Ende für eiszeitliche Riesen. In: Bild der Wissenschaft. 26. November 2008, abgerufen am 8. September 2019.
- Jelle W. F. Reumera et al.: Late Pleistocene Survival of the Saber-toothed Cat Homotherium in northwestern Europe. Journal of Vertebrate Paleontology, Volume 23, Issue 1 (March 2003).
- Dick Mol, Wilrie van Logchem, Kees van Hooijdonk & Remie Bakker (2007): The Saber-toothed Cat of the North Sea. Uitgeverij DrukWare, KS Norg. ISBN 90-78707-04-6.
- Stuart Fiedel, Gary Haynes: A premature burial: comments on Grayson and Meltzer’s “Requiem for overkill” Journal of Archaeological Science 31 (2004) 121–131, doi:10.1016/j.jas.2003.06.004.
- Cione, A. L., Tonni, E. P., Soibelzon, L. (2003). The Broken Zig-Zag: Late Cenozoic large mammal and tortoise extinction in South America. Rev. Mus. Argentino Cienc. Nat., 5(1), 1–19. ISSN 1514-5158.
- David L. Fox, Daniel C. Fisher, Sergey Vartanyan, Alexei N. Tikhonov, Dick Mol, Bernard Buigues: Paleoclimatic implications of oxygen isotopic variation in late Pleistocene and Holocene tusks of Mammuthus primigenius from northern Eurasia. In: Quaternary International 169–170, 2007, S. 154–165.
- Steadman, D. W., Martin, P. S., McPhee, R. D. E., Jull, A. J. T., McDonald, H. G., Woods, C. A., Iturralde-Vinent, M., Hodgins, G. W. L. (2005). Asynchronous extinction of late Quaternary sloths on continents and islands. PNAS vol. 102 no. 33 11763–11768
- Perez, V., R., Godfrey, L., R., Nowak-Kemp, M., Burney, D., A., Ratsimbazafy, J., Vasey, N. (2005): Evidence of early butchery of Giant lemurs in Madagaskar. Journal of Human Evolution, 49, S. 722–742.
- P.S. Martin, Prehistoric overkill. In: P.S. Martin and H.E. Wright (Hrsg.), Pleistocene Extinctions: The Search for a Cause. Yale University Press, New Haven (1967), ISBN 0-8165-1100-4, S. 75–120.
- S.L. Whittington and B. Dyke: Simulating overkill: experiments with the Mosimann and Martin model. In: P.S. Martin and R.G. Klein, Editors, Quaternary Extinctions: A Prehistoric Revolution, University of Arizona Press, Tucson (1984), S. 451–465.
- J. Alroy, A multispecies overkill simulation of the end-Pleistocene megafaunal mass extinction, Science 292 (2001), S. 1893–1896.
- D.C. Fisher: Season of death, growth rates, and life history of North American mammoths. In: D. West, Editor, Proceedings of the International Conference on Mammoth Site Studies. Publications in Anthropology 22, University of Kansas, Lawrence, KS (2001), pp. 121–135.
- P.S. Martin and R.G. Klein, Editors, Quaternary Extinctions: A Prehistoric Revolution, University of Arizona Press, Tucson (1984).
- R.D.E. MacPhee, Editor, Extinctions in Near Time: Causes, Contexts, and Consequences, Kluwer/Plenum, New York (1999).
- Anthony D. Barnosky, P.L. Koch, R.S. Feranec, S.L. Wing and A.B. Shabel, Assessing the causes of Late Pleistocene extinctions on the continents, Science 306 (2005), S. 70–75.
- D.K. Grayson, D.J. Meltzer, A requiem for North American overkill. Journal of Archaeological Science 30 (2003), S. 585–593.
- Sander van der Kaars, Gifford H. Miller, Chris S. M. Turney, Ellyn J. Cook, Dirk Nürnberg, Joachim Schönfeld, A. Peter Kershaw, Scott J. Lehman: Humans rather than climate the primary cause of Pleistocene megafaunal extinction in Australia. In: Nature Communications. 8, 2017, S. 14142, doi:10.1038/ncomms14142.
- Chris S. M. Turneya, Timothy F. Flannery, Richard G. Roberts, Craig Reid, L. Keith Fifield, Tom F. G. Higham, Zenobia Jacobs, Noel Kemp, Eric A. Colhoun, Robert M. Kalin, Neil Ogle: Late-surviving megafauna in Tasmania, Australia, implicate human involvement in their extinction. PNAS, vol. 105, no. 34, S. 12150–12153, August 26, 2008
- Morten Erik Allentoft, Rasmus Heller, Charlotte L. Oskam, Eline D. Lorenzen, Marie L. Hale, M. Thomas P. Gilbert, Christopher Jacomb, Richard N. Holdaway, Michael Bunce: Extinct New Zealand megafauna were not in decline before human colonization. In: Proceedings of the National Academy of Sciences. 111, 2014, S. 4922, doi:10.1073/pnas.1314972111.
- Barry W. Brook, David A. Burney, Timothy F. Flannery, Michael K. Gagan, Richard Gillespie, Christopher N. Johnson, Peter Kershaw, John W. Magee, Paul S. Martin, Gifford H. Miller, Benny Peiser, Richard G. Roberts: Would the Australian megafauna have become extinct if humans had never colonised the continent? Comments on ‘‘A review of the evidence for a human role in the extinction of Australian megafauna and an alternative explanation’’ by S. Wroe and J. Field. QSR Correspondence. Quaternary Science Reviews 26 (2007) 560–564
- Eleftheria Palkopoulou, Swapan Mallick, Pontus Skoglund, Jacob Enk, Nadin Rohland, Heng Li, Ayça Omrak, Sergey Vartanyan, Hendrik Poinar, Anders Götherström, David Reich und Love Dalén: Complete Genomes Reveal Signatures of Demographic and Genetic Declines in the Woolly Mammoth. Current Biology 25, 2015, S. 1395–1400, doi:10.1016/j.cub.2015.04.007.
- Erin Fry, Sun K. Kim, Sravanthi Chigurapti, Katelyn M. Mika, Aakrosh Ratan, Alexander Dammermann, Brian J. Mitchell, Webb Miller und Vincent J Lynch: Functional Architecture of Deleterious Genetic Variants in the Genome of a Wrangel Island Mammoth. Genome Biology and Evolution 12 (3), 2020, S. 48–58, doi:10.1093/gbe/evz279.
- Firestone, R. B., et al.: Evidence for an extraterrestrial impact 12,900 years ago that contributed to the megafaunal extinctions and Younger Dryas cooling. Proceedings of the National Academy of Sciences 104, 2007. S. 16016–16021. Abstract
- Surovell, T. A., Holliday, V. T., Gingerich, J. A. M., Ketron, C., Haynes, C. V. Jr., Hilman, I., Wagner, D. P., Johnson, E., Claeys, P. (2009). An independent evaluation of the Younger Dryas extraterrestrial impact hypothesis. PNAS. vol. 106, no. 43, pp. 18155–18158.
- Briggs Buchanan, Mark Collard, Kevan Edinborough: Paleoindian demography and the extraterrestrial impact hypothesis. Proceedings of the National Academy of Sciences 105, 2008. S. 11651–11654. Abstract
- Richard A. Kerr: Did the Mammoth Slayer Leave a Diamond Calling Card?. In: American Association for the Advancement of Science (Hrsg.): Science. 323., Nr. 5910,, Januar 2009, S. 26. doi:10.1126/science.323.5910.26.
- Kenneth Chang: Diamonds linked to quick cooling eons ago. In: Health and Science, International Herald Tribune, International Herald Tribune, 2. Januar 2009. Abgerufen am 3. Januar 2009.
- D. J. Kennett Kennett, J. P. Kennett, A. West, C. Mercer, S. S. Que Hee, L. Bement, T. E. Bunch, M. Sellers, W. S. Wolbach: Nanodiamonds in the Younger Dryas Boundary Sediment Layer. In: American Association for the Advancement of Science (Hrsg.): Science. 323., Nr. 5910, Januar 2009, S. 94. doi:10.1126/science.1162819.
- Rex Dalton: Comet Theory Comes Crashing to Earth (en) In: Pacific Standard Magazine. Sara Miller McCune. 2011. Abgerufen am 28. November 2018.
- T. L. Daulton, N. Pinter, A. C. Scott (30. August 2010): No evidence of nanodiamonds in Younger–Dryas sediments to support an impact event. Proc. Natl. Acad. Sci. U.S.A. 107 (37): 16043–7.
- Did comet smashup unleash Earth's mini Ice Age? auf: dawn.com 3. April 2010, abgerufen am 28. November 2018.
- Pinter, N. et al. (2011): The Younger Dryas impact hypothesis: A requiem. Earth-Science Reviews 106, 247–264, doi:10.1016/j.earscirev.2011.02.005.
- Mario Pino, Ana M. Abarzúa, Giselle Astorga, Alejandra Martel-Cea, Nathalie Cossio-Montecinos, R. Ximena Navarro, Maria Paz Lira, Rafael Labarca, Malcolm A. LeCompte, Victor Adedeji, Christopher R. Moore, Ted E. Bunch, Charles Mooney, Wendy S. Wolbach, Allen West, James P. Kennett: Sedimentary record from Patagonia, southern Chile supports cosmic-impact triggering of biomass burning, climate change, and megafaunal extinctions at 12.8 ka. In: Nature Scientific Reports. 9, März 2019. doi:10.1038/s41598-018-38089-y.
- Andrew M. T. Moore, James P. Kennett, William M. Napier, Ted E. Bunch, James C. Weaver, Malcolm LeCompte, A. Victor Adedeji, Paul Hackley, Gunther Kletetschka, Robert E. Hermes, James H. Wittke, Joshua J. Razink, Michael W. Gaultois, Allen West: Evidence of Cosmic Impact at Abu Hureyra, Syria at the Younger Dryas Onset (~12.8 ka): High-temperature melting at >2200 °C. In: Nature Scientific Reports. 10, März 2020. doi:10.1038/s41598-020-60867-w.
- Francis Thackeray, Louis Scott, P. Pieterse: The Younger Dryas interval at Wonderkrater (South Africa) in the context of a platinum anomaly. (PDF) In: Palaeontologia Africana. 54, Oktober 2019, S. 30–35. doi:10.5067/ASTER/ASTGTM.002.
- Ewe, Thorwald: Das große Sterben, in Focus-online.de (23. März 2008)
- Paul A. LaViolette: Evidence for a Solar Flare Cause of the Pleistocene Mass Extinction. In: Dept. of Geosciences, University of Arizona (Hrsg.): Radiocarbon. Nr. 53(2), Juni 2011, S. 303–323, doi:10.2458/azu_js_rc.53.3464.
- Anthony J. Stuart: Late Quaternary megafaunal extinctions on the continents: A short review. In: Liverpool Geological Society (Hrsg.): Geological Journal. Nr. 50(3), Dezember 2014, S. 342, doi:10.1002/gj.2633.
- M. Timothy Rabanus-Wallace, Matthew J. Wooller, Grant D. Zazula, Elen Shute, A. Hope Jahren, Pavel Kosintsev, James A. Burns, James Breen, Bastien Llamas, Alan Cooper. Megafaunal isotopes reveal role of increased moisture on rangeland during late Pleistocene extinctions. Nature Ecology & Evolution, 2017; 1 (5): 0125 DOI: 10.1038/s41559-017-0125