Ouled-Abdoun-Becken
Das Ouled-Abdoun-Becken ist eine Region sowie eine wichtige Rohstoff- und Fossillagerstätte in Marokko. Es befindet sich im zentralen Teil des Landes im nördlichen Vorland des Hohen Atlas, etwa 100 km südöstlich von Casablanca. Eine herausragende überregionale Bedeutung erlangte das Becken neben seinem Fossilreichtum durch immense Phosphatvorräte, die zu den größten der Welt zählen. Die Phosphate lagern in einer kompakten Sedimenteinheit bestehend aus Kalk- und Mergelsteinen von teilweise nur 15 bis 30 m Mächtigkeit und sind in mehreren Lagen ausgebildet. Die Schichtenfolge wird als „Phosphatserie“ bezeichnet. Sie setzt sich nach Südwesten über einzelne weitere Becken bis in den Norden der Westsahara fort und findet sich zusätzlich noch am Südrand des Hohen Atlas. Dabei nimmt sie teilweise an Mächtigkeit zu, die Phosphate verlieren aber mitunter an Qualität. Die Entstehung der Phosphatlagen erstreckte sich über einen Zeitraum von der Oberkreide bis zum Mittleren Paläozän vor rund 72 bis 48 Millionen Jahren. Ihren Ursprung haben sie in einem damals flachgründigen Meer, das den Südrand des ehemaligen Tethys-Ozeans bildete. Heute werden die Phosphate sowohl im Tage- als auch im Untertagebau in einer Größenordnung von mehreren Millionen Tonnen jährlich wirtschaftlich gefördert und weltweit exportiert.
Fossilien finden sich in allen Abschnitten der Phosphatserie, sie treten aber teilweise konzentriert auf. Aufgrund des langen Bildungszeitraumes der Ablagerungen von über 25 Millionen Jahren schließen sie die Kreide-Paläogen-Grenze vor rund 66 Millionen Jahren und das Paläozän/Eozän-Temperaturmaximum vor rund 56 Millionen Jahren ein. Der bei weitem überwiegende Anteil der Fossilfunde umfasst Tiere, Pflanzen spielen nur eine untergeordnete Rolle. Es dominieren dem Bildungsmilieu der Phosphatserie entsprechend Meeresbewohner. Unter diesen haben die Hai- und Rochenartigen eine herausragende Stellung, da sie einerseits die häufigste Fundkategorie bilden, andererseits über die Biostratigraphie zur zeitlichen Einordnung der einzelnen Schichten herangezogen werden. Des Weiteren ist auch eine größere Anzahl an Knochenfischen belegt. Zusätzlich tritt eine sehr vielfältige Reptiliengemeinschaft auf. Diese setzt sich aus Schildkröten, Krokodilen, Mosasauriern und Plesiosauriern zusammen, schließt aber auch Schlangen und Warane mit ein. Als bemerkenswert kann das Vorkommen verschiedener Flugsaurier und Vögel angesehen werden, ebenso wie das von landbewohnenden Tieren, die sowohl Dinosaurier als auch Säugetiere beinhalten. Vor allem diese Reste verweisen darauf, dass die Bildung der Phosphatserie relativ küstennah erfolgte. Im Gegensatz zu den Funden der meeresbewohnenden Reptilien, die teilweise als vollständige Skelette vorliegen, ist das Material der landlebenden Formen stärker fragmentiert.
Die erste Erwähnung von Phosphatlagen im Ouled-Abdoun-Becken erfolgte im Jahr 1917, nur wenige Jahre späte begann die kommerzielle Förderung der Rohstoffe. Die bereits zu diesem Zeitpunkt bekannten reichhaltigen Fossillagen führten ab Mitte der 1930er Jahre zu einer intensiven Untersuchung durch Camille Arambourg. Er erarbeitete in den nächsten zwanzig Jahren eine detaillierte stratigraphische Gliederung und zeitliche Einstufung der Phosphatserie, die bis heute Bestand hat. Nach seinem Ausscheiden in den 1950er Jahren kam die wissenschaftliche Untersuchung im Ouled-Abdoun-Becken weitgehend zum Erliegen. Erst die Entdeckung einzelner Reste landbewohnender Säugetiere in den 1990er Jahren schob den Fokus des wissenschaftlichen Interesses wieder auf die Region, der bis heute ununterbrochen ist.
Geographische Lage
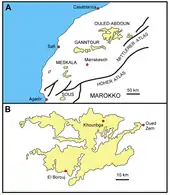
Das Ouled-Abdoun-Becken liegt im zentralen Marokko etwa 100 km südöstlich von Casablanca. Es befindet sich im westlichen Abschnitt der marokkanischen Meseta. Des Weiteren gehört es zu mehreren bedeutenden Phosphatbecken des Landes, die wiederum einen Teil der Mediterranean (Tethyan) phosphogenic Province formen, welche sich als breiter Ablagerungsgürtel vom Nahen Osten bis in den Osten von Brasilien erstreckt. In Marokko werden verschiedene Becken dieser Provinz zugesprochen. Im Nordosten bildet das Ouled-Abdoun-Becken den Anfang, nach Südwest folgen das Ganntour-, Meskala- und Sous-, während das Oued-Eddahab-Becken die Reihe in der Westsahara abschließt. Die einzelnen Becken sind durch Auffaltungsprozesse des Atlas voneinander getrennt.[1][2][3]
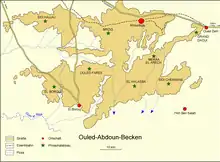
Von den fünf Phosphatbecken Marokkos stellt das Ouled-Abdoun-Becken das größte dar. Es erstreckt sich über 80 km in Ost-West- sowie 60 km in Nord-Süd-Richtung und nimmt eine Fläche von rund 9000 km² ein. Im Osten begrenzt der Hohe Atlas das Becken, im Westen das Rhamna-Massiv und im Süden das Beni-Amir-Plateau. Außerdem wird es im Süden und Westen vom Oum er-Rbia umflossen. Als größere Städte befinden sich Khouribga im Norden, Oued Zem im Nordosten und El Borouj im Südwesten des Ouled-Abdoun-Beckens. Die Höhenlagen reichen von 500 m im Südwesten bis 875 m im Norden. Die überwiegend ebene Landschaft wird durch trockenes Wüstenklima bestimmt. Der stärkste Niederschlag fällt von November bis Mai, er überschreitet aber kaum 400 mm auf das Jahr verteilt. Die Vegetation ist eher spärlich. Die örtliche Bevölkerung lebt von Viehzucht und jahreszeitlich beschränkter Landwirtschaft. Sie verteilt sich auf mehrere Ortschaften. Grundwasser ist generell selten und wird aus zum Teil über 100 m tiefen Brunnen geschöpft.[1][4][5][6]
Geologie
Geologische Abfolge im Ouled-Abdoun-Becken

Die Basis der Sedimentabfolge bilden paläozoische Ablagerungen bestehend aus Glimmerschiefer und Quarziten. Darauf folgen die Schichten der Oberkreide und des Paläogens, so dass zwischen diesen und den basalen Lagen ein großer zeitlicher Hiatus existiert. Die unteren Einheiten der Oberkreide setzen sich weitgehend aus Mergeln, Kalk- sowie Ton- und Schluffsteinen mit einer Mächtigkeit von bis zu 200 m zusammen. Sie werden in den Zeitraum vom Cenomanium bis Campanium eingestuft. Den oberen Abschluss bildet eine Lage teils intensiv verwitterter Gipse.[5][7][8]
Erst im Hangenden dieser Schichtenreihe findet sich die eigentliche „Phosphatserie“ (série phosphatée). Sie ist im gesamten Gebiet relativ einheitlich aufgebaut und horizontal gelagert. Innerhalb der Sedimentabfolge lassen sich vier Gesteinstypen unterscheiden, die sich so in der Regel auch in anderen Phosphatlagerstätten wiederfinden. Der wichtigste wird durch die Phosphatgesteine repräsentiert. Diese treten entweder als lockere, sandig-mergelige pseudo-oolithische Phosphorite (Phospharenit) oder als feste phosphathaltige Kalksteine (Phospharudit) auf. Letztere sind wiederum einerseits mit primärem Dolomit- oder mit sekundärem Calcitzement verbacken. Neben diesem hauptsächlichen Bestandteil kommen als weitere Gesteinstypen Ton- und Mergelsteine sowie Kalksteine und vorwiegend als Knollen oder plattig auftretende Silikatgesteine vor. Die Abfolge wird in vier Ablagerungseinheiten oder Megasequenzen unterschieden, die dem Maastrichtium (ausgehende Oberkreide), dem Danium/Thanetium (Paläozän), dem Ypresium (Unteres Eozän) und dem Lutetium (Mittleres Eozän) angehören. Die drei unteren enthalten dabei die hauptsächlichen Phosphatablagerungen. Diese bedeutendsten Schichteinheiten werden jeweils durch eine Hauptdiskontinuität voneinander getrennt. Generell setzt sich die Phosphatserie im Ouled-Abdoun-Becken folgendermaßen zusammen:[1][4][5][7][9]
- Maastrichtium (Megasequenz A): vorwiegend aus sandigen Phosphoriten aufgebaut, die oft auf Tonen und Mergeln gebettet sind und mitunter von Kalksteinen unterbrochen werden; typisch sind linsenförmige Silikatgesteine; teils gelblich gefärbt;
- Danium/Thanetium (Megasequenz B): besteht aus sandigen und mergeligen Phosphoriten und zwischengebetteten silikatischen Mergeln; das Danium zeichnet sich durch das Fehlen von tonigen und mergeligen Zwischenschichten aus, sein oberer Abschluss wird durch die Anhäufung der von Cardita coquandi-Muschelschalen angezeigt;
- Ypresium (Megasequenz C): bildet eine Wechsellagerung aus gelblichen bis gräulichen sandigen Phosphoriten, gräulichen phosphatischen Kalksteinen sowie weißlichen bis grünlichen mergeligen Kalksteinen und kompakten grünlichen bis bräunlichen Tonmergel mit Silikatknollen;
- Lutetium (Megasequenz D): ist als Wechsellagerung aus silikatischen und calcitischen Mergeln sowie mergeligen Kalksteinen ausgebildet, in denen Silikatknollen eingelagert sind und gelegentlich auch dünne Phosphathorizonte auftreten.
Die Phosphatserie weist eine schwankende Mächtigkeit auf. Im Norden und Nordosten des Ouled-Abdoun-Beckens wird sie nur etwa 15 bis 30 m dick. Die einzelnen Ablagerungseinheiten erreichen mit dem Maastrichtium eine vertikale Ausdehnung von 3 bis 5 m, mit dem Danium/Thanetium von etwa 3 m und mit dem Ypresium von gut 12 m, während das Lutetium noch sehr geringmächtig ist. Nach Süden hin mit abfallender Geländeoberfläche nimmt die Phosphatserie deutlich an Dicke zu. Am Südrand des Ouled-Abdoun-Beckens liegen die Schichtmächtigkeiten für das Maastrichtium bei 8 bis 10 m, für das Danium/Thanetium bei 8 m und für das Ypresium bei 12 m. Das Lutetium erreicht dagegen gut 25 bis 30 m. Mit eingerechnet ist hier der sogenannte Thersiteenkalk, auch Dalle à Thersitées („Thersitea-Platte“) genannt. Der feste bankige Kalkstein bildet den oberen Abschluss der Folge und wird durch das häufige Auftreten von Resten der Schnecke Hemithersitea gekennzeichnet.[1][4][7][9]
Die einzelnen Phosphatlagen werden als couche („Bett“) oder sillon („Furche“ oder „Horizont“) bezeichnet und sind der stratigraphischen Abfolge nach durchnummeriert. Die beste Aufschlüsselung findet sich im Nordostteil des Ouled-Abdoun-Beckens, in dem Bereich mit dem zur Zeit intensivsten Abbau. Die Phosphatlage des Maastrichtiums trägt hier die Bezeichnung couche III, ist aber manchmal zweigeteilt in einen unteren und einen oberen Abschnitt. Das Danium/Thanetium entspricht den Phosphatlagen couche IIb und couche IIa. Das Ypresium besteht aus wenigstens vier Phosphatlagen, die vom Liegenden zum Hangenden mit couche I, couche 0, sillon A und sillon B benannt sind. Unter Umständen setzt sich zwischen dem couche I und dem couche 0 zusätzlich eine dünne, mit couche 0' ausgewiesene Phosphatschicht ab, ebenso wie einzelne Phosphatbereiche oberhalb von sillon B fortlaufende Buchstaben erhalten (sillon C und sillon D). Die Schichten zwischen den verschiedenen Phosphatlagen werden als intercalaire („Trennschicht“) angesprochen. Demnach weist intercalaire II/I auf die Trennschicht zwischen couche II und couche I hin, intercalaire III/II auf die zwischen couche III und couche II. Die Trennschichten variieren in ihrer Dicke sehr und enthalten mitunter Taschen oder Linsen an Phosphaten.[4][7]
Überregionaler Kontext
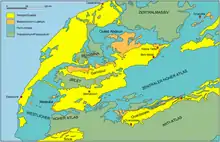
Die gesamte Phosphatserie ist marinen Ursprungs und findet sich in einem ähnlichen Aufbau auch in den anderen Phosphatbecken Marokkos wieder. Die starken Variationen in der Mächtigkeit zwischen dem Nord- und dem Südrand des Ouled-Abdoun-Beckens erklären sich durch die Entstehungsbedingungen der Phosphatlagen in einem einst buchtartigen Meereseinschnitt. Die nördlichen Gebiete lagen dabei in ehemaliger Küstennähe und zeichnen sich durch einen hohen Phosphatreichtum aus. Die Meeresrandlage führte aber zu einem stark kondensierten Aufbau mit mehreren Schichtlücken und zudem scharfen Schichtübergängen.[10] Nach Süden hin zum ehemals offenen Meer löst sich diese Kondensierung weitgehend auf und wechselt zu einer vollständigen Schichtenabfolge. Diese ist dann aus dem südlich benachbarten Ganntour-Becken belegt. Die Unterschiede zwischen den küstennahen und küstenfernen Ablagerungen verdeutlichen sich etwa in den obersten Abschnitten der Oberkreide, dem Maastrichtium. Dieses weist im Norden des Ouled-Abdoun-Beckens nur eine Mächtigkeit von 3 bis 5 m auf, im Ganntour-Becken hingegen von gut 45 m. Dadurch sind hier allein im Maastrichtium sieben Phosphatlagen unterscheidbar, die sich teilweise aber auch schon am Südwestrand des Ouled-Abdoun-Beckens abzeichnen. Auch im auflagernden Danium/Thanetium und im Ypresium können durch zunehmende Schichtmächtigkeit und -differenzierung jeweils vier bis sechs einzelne Phosphatlager im Ganntour-Becken auseinander gehalten werden. Sowohl die Zunahme der Schichtmächtigkeit als auch der Rückgang der Kondensierung sind mit einer Abnahme des Phosphatgehalts und einer Anreicherung an organischem Material verbunden. Die Phosphatlagen wechseln dadurch von einer überwiegend gelblich-gräulichen Färbung im Norden des Ouled-Abdoun-Beckens zu einer dunklen Tönung und nehmen zudem einen stechenden Geruch an.[4][7][11][2]
Die sich wiederholenden Sedimentabfolgen der Phosphatserie sind Resultat eines zyklischen Vordringens und nachfolgenden Rückzugs des Meeres. Die Transgressionsphasen werden durch die kalkigen Abschnitte angezeigt, die Regressionsphasen durch die sandig-schluffigen. Die marinen Ablagerungen der Oberkreide und des Paläogens setzen sich über die Phosphatbecken hinaus nach Süden fort und können ebenfalls im Ouarzazate-Becken am Südrand des Hohen Atlas beobachtet werden. Dort sind die Phosphatlager allerdings vergleichsweise marginal ausgebildet. Sie kommen aber noch reichhaltig unter einer mächtigen Deckschichtenlage in der Tadla-Ebene vor und finden sich gleichfalls in Relikten in Aufschlüssen in der Ben-Cherro-Mulde bei Beni Mellal, beide jeweils am Nordrand des Hohen Atlas gelegen. Die Meeresablagerungen erstrecken sich somit über die gesamte Breite des Atlas hin, wurden später aber durch die Auffaltung des Gebirges überschleiert. Den variierenden Sedimentmächtigkeiten der marinen Ablagerungen im Ouarzazate-Becken zufolge entstand die Gesteinsabfolge auf einer einst nach Süden ansteigenden Geländefläche. Dadurch war das südliche Ende des Meeres vermutlich im Bereich des heutigen Anti-Atlas zu finden.[12][13][14][15]
Ursprung der Phosphatablagerungen
Phosphorite bilden in der Regel rundliche Gesteine, deren einzelnen Phosphatkörner durch sekundären Zement mehr oder weniger fest miteinander verbacken sind. Die Phosphatkörner bestehen zumeist aus organischem Abfall, Kleinkonkretionen mit konzentrischer Struktur, Mikroonkolithen, ummantelten Körnern („pellets“), Kotkügelchen und verschiedenen phosphatiserten Mikroorganismen wie Algen, Bakterien oder Pilzen. Teilweise kommen auch größere organische Bestandteile wie Knochen und Zähne vor. Dagegen ist nicht-phosphathaltiger organischer Schill eher selten. Unabhängig von ihrem Ursprung bestehen alle Phosphatkörner aus Carbonat-Fluorapatit, das jeweils als strukturiertes Kristall auftritt.[10][4]
Die Entstehung von phosphathaltigen Sedimentlagen, die sogenannte Phosphatogenese, ist komplex und vielfach diskutiert. Sie schließt tektonische, geomorphologische, klimatische, biochemische und biologische Faktoren ein. Die Phosphatablagerungen der Mediterranean (Tethyan) phosphogenic Province entstanden in einem langandauernden Prozess, der geologisch gesehen vom Turonium in der Oberkreide bis zum Lutetium im Eozän reicht. Er setzte somit vor rund 94 Millionen Jahren ein und kam vor mehr als 41 Millionen Jahren zum Erliegen, verlief also über rund 50 Millionen Jahre. Die Bildungsregion entspricht den Randlagen der damaligen Tethys. Dort erfolgte die Phosphatogenese aber nicht überall gleichzeitig. Vielmehr begann sie im nordöstlichen Abschnitt, in der heutigen Türkei und verlagerte sich sukzessive in andere Regionen. Die marokkanischen Phosphatablagerungen nahmen dabei einen der längsten Bildungszeiträume ein, der sich über gut 25 Millionen Jahre vom Maastrichtium bis zum Ypresium hinzog.[10][7]
Die Phosphatogenese stellt eine organische Sedimentation (Bioproduktit) dar, die an Meeresbedingungen gekoppelt ist. Hier kann sie an verschiedensten Orten erfolgen, die neben Kontinentalhängen auch Inseln, Tiefseeberge oder Guyots einschließen. Neben einigen anderen Prozessen wird sie hauptsächlich durch aufsteigende Kaltwasserströmungen begünstigt. Derartige vertikale Wasserbewegungen sind im nördlichen Atlantik erstmals seit der frühen Oberkreide belegt und intensivierten sich zum Ende der Oberkreide hin.[16] Die aufsteigenden Strömungen bringen unter anderem mit Phosphaten angereichertes Detritusmaterial aus den Meerestiefen in die Schelfgebiete mit. Hierbei kommt es dann durch Übersättigung zu einer primären Ausfällung von Apatit. In einer späteren, durch hydrologische und biologische Prozesse gesteuerten Aufarbeitung entstehen daraus die typischen, aus rundlichen Körnern aufgebauten Phosphorite. Einen bedeutenden Einfluss haben hierbei offensichtlich Mikroorganismen,[17][18] die in Form von gramnegativen Bakterien auch fossil aus dem Ouled-Abdoun-Becken überliefert sind.[19] Vor allem für die zweite Phase der Phospatogenese sind günstige Lagerungsbedingungen in einem Becken von Bedeutung, da diese stabile Umweltbedingungen voraussetzt, etwa stratifizierte Wassersäulen mit nur einem geringen Austausch zwischen den einzelnen Wasserschichten. Diese sekundäre Stufe der Phosphatogenese ist für die Schelfgebiete typisch. Dazu gehören auch die marokkanischen Phosphatbecken, welche sich am Rand eines epikontinentalen Meereseinschnittes bildeten.[20][10][7][21]
Eigenschaften und Bedeutung der Phosphatablagerungen

Im Ouled-Abdoun-Becken kommen überwiegend zwei Typen an Phosphatkörnern vor: Zum Einen als schlecht sortierte Ansammlung von Körnern mit einem Durchmesser von mehr als 500 μm, in die zahlreiches organisches Material eingearbeitet ist, zum Anderen als gut ausgebildete Körner von kugeliger Gestalt mit einem Durchmesser von 100 bis 500 μm. Die letzte Variante dominiert in den Ablagerungen, die erste hat einen Anteil von weniger als 10 %. Generell ist das Phosphat der Phosphatbecken Marokkos arm an Silicium-, Aluminium- oder Eisengehalt und vergleichsweise hochgradig mit einem Phosphatanteil von 78 bis hin zu 82 %, was 35,7 % bis 38 % an Phosphorpentoxid entspricht. Ausnahmen bilden die Phosphatablageerungen des Maastrichtiums, bei denen der Gehalt von Phosphorpentoxid bei 25 bis 28 % liegt. Die Phosphatablagerungen mit den besten Eigenschaften finden sich in den Schichten des Ypresiums im Grand Daoui. Sie enthalten weitgehend noch 14 bis 16 % an Wasser, sind allerdings leicht verwittert.[22][23][4]
Die Phosphatvorkommen Marokkos gelten mit schätzungsweise 51 Milliarden Tonnen an Phosphoriten als die weltweit reichsten Rohstoffquellen dieser Art, sie repräsentieren nach einigen Angaben etwa drei Viertel der Weltressourcen.[24] Die Zahlen werden teils kritisch gesehen, allerdings sind sowohl das Ouled-Abdoun-Becken als auch die weiteren Phosphatlagerstätten Marokkos bisher nicht vollständig erkundet.[25][5][26] Jährlich werden in dem Land rund 19 Millionen Tonnen an Phosphaten gefördert, von denen zwischen 13 und 15 Millionen Tonnen in den weltweiten Handel gelangen. Ein großer Anteil davon stammt aus dem Ouled-Abdoun-Becken, dessen Reserven noch Mitte der 1990er Jahre auf rund 28 Milliarden Tonnen an Phosphoriten beziffert wurden.[4] Im Ouled-Abdoun-Becken sind mehr als ein Dutzend Lagerstätten erkundet, die wichtigsten davon finden sich mit Mrzig, Khouribga und Grand Daoui im nördlichen, mit Meraâ El Arech und Ouled Farès im zentralen sowie mit El Borouj, El Halassa und Sidi Chennane im südlichen Abschnitt.[9] Der Großteil des Abbaus der Phosphorite erfolgt auf Grund der harten Deckschichten unter Tage. Lediglich im Norden des Ouled-Abdoun-Beckens, wo die auflagernden Kalksteine teils stärker verwittert sind, ist seit der Mitte des 20. Jahrhunderts auch ein Tagebaubetrieb verbreitet. Er wurde zusätzlich begünstigt durch die Entwicklung großer Abraummaschinen wie unter anderem der Schürfkübelbagger „Marion 7900“, der bei seinem Dienstantritt 1960 im Grand Daoui-Abbaugebiet mit einem Gewicht von 1870 t, einem Kranausleger von 84 m Länge, einem Aktionsradius von 59 m und einem Schaufelvolumen von 39 m³ der größte seiner Art war.[22][27][4] Teilweise erschwert werden die Arbeiten durch sterile Abschnitte aus silikatischem Kalkstein, die aus dem Hangenden in die Phosphatlager eindringen und zwischen 10 und 150 m im Durchmesser aufweisen. Möglicherweise gehen sie auf die Evaporitlage am oberen Abschluss der Campanium-Folge zurück, die durch Karstverwitterung Hohlräume ausbildete. Diese brachen folgend zusammen, was zum Nachrutschen der auflagernden Sedimente führte.[28][29][26][8]
Fossilfundstellen
Fossilreste von Wirbeltieren treten in der gesamten Sequenz der Phosphatserie auf, sowohl in den Phosphatlagen als auch in den Zwischenschichten. Sie unterscheiden sich aber in ihrem Reichtum, ihrer Zusammensetzung und ihrer Erhaltung. In den Ablagerungen des Maastrichtiums dominieren Meeresbewohner von Fischen über Krokodile, Schuppenkriechtiere bis hin zu Schildkröten. Darüber hinaus sind Flugsaurier und gelegentlich Reste von Dinosauriern dokumentiert. Neben Einzelfunden von Knochen und Zähnen kommen hier auch teils gut erhaltene Skelette vor. Ein Großteil der Fossilien konzentriert sich in einem Knochenbett (bone bed), markiert durch seinen Reichtum an Fischwirbeln und Haizähnen, das etwa in der Mitte des couche III ausgebildet ist.[30][31] In den Schichten des Paläogens überwiegen wiederum neben Fischen vor allem Krokodile und Schildkröten. Vögel und Säugetiere hingegen sind eher rar und zudem durchgehend deutlich stärker fragmentiert. Das Auftreten von landbewohnenden Säugetieren in den marinen Sedimenten des Paläogens, vergleichbar mit den Dinosaurierresten aus der Oberkreide, setzt einen längeren Transport von den Küstengebieten und dem kontinentalen Hinterland voraus. Da aber zum Teil gut erhaltene Schädel mit gelegentlich vollständigen Zahnreihen vorliegen, fand diese Umlagerung wohl unter eher niedrigenergetischen Bedingungen statt. Funde von Säugetieren stammen überwiegend aus mehreren angereicherten Knochenschichten, die sich einerseits an der Basis und innerhalb des couche IIa (F1 oder das sogenannte Eritherium bone bed sowie F2 oder das big coprolite bone bed), andererseits an der Basis des intercalaire II/I (F3 oder das sogenannte Otodus obliquus bone bed beziehungsweise Phosphatherium bone bed) befinden. Die Knochenkonzentrationen zeichnen sich durch eine große Fülle an disartikulierten Wirbeltierresten aus, die in einer Lage aus Silikatgeröllen und grünlichen Tonen, möglicherweise Smektiten, eingebettet sind. Bedingt durch den Phosphatabbau befinden sich die meisten Fundstellen an Wirbeltierresten im nordöstlichen Ouled-Abdoun-Becken, hauptsächlich im Bereich von Grand Daoui, allein für die Säugetiere können hier acht verschiedene Lokalitäten benannt werden. Des Weiteren sind auch im Abschnitt Sidi Chennane im Südosten mehrere Fundplätze mit Wirbeltierresten dokumentiert.[7][32][33]
Funde
Zusammensetzung
Der Fossilreichtum des Ouled-Abdoun-Beckens ist enorm. Es liegen Reste von mehr als 330 Arten an Wirbeltieren vor, die sich auf über 190 Gattungen in über 85 Familien aufteilen. Ein letzter Gesamtüberblick zum Fossilgehalt der Region ist im Jahr 2017 von Nathalie Bardet und Kollegen erstellt worden,[34] seitdem sind jedoch mehrere Taxa neu eingeführt worden.[30][31][35] Den größten Anteil an der Zusammensetzung des paläontologischen Fundmaterials haben die Fische, hierunter wiederum vor allem die Hai- und Rochenartigen, die allein mehr als 250 Arten und 120 Gattungen stellen. Sie treten in allen Fundlagen auf. Ihre große Vielfalt und ihr häufiges Vorkommen haben unschätzbare Bedeutung für die Biostratigraphie und damit für die zeitliche Einordnung der fossilführenden Schichten. Sonstige Fische, vor allem die Knochenfische, sind dagegen mit rund 20 Arten aus fast ebenso vielen Gattungen und Familien deutlich seltener. Einige Gruppen wie die Eidechsenfischartigen bleiben bisher vollständig auf die Oberkreide beschränkt, andere wie die Barschartigen hingegen auf das Paläogen. Eine sehr heterogene Gruppe bilden die Sauropsiden, die übertragen die Reptilien einschließlich der Dinosaurier und die Vögel umfassen. Sie können auf über 55 Arten in mehr als 40 Gattungen und beinahe 20 Familien verweisen. Das Fundmaterial verteilt sich relativ einheitlich über die verschiedenen Zeitebenen der Ablagerungen, die Vögel sind allerdings bisher weitgehend nur aus dem Paläogen und die Dinosaurier aus der Oberkreide überliefert. Spärlich treten bisher Säugetiere in Erscheinung. Sie kommen nur im Paläozän und Eozän vor und repräsentieren rund ein Dutzend Arten in über neun Gattungen aus einer Handvoll Familien. Gegenüber den Wirbeltieren haben Wirbellose und Pflanzen nur einen geringen Anteil im Fossilbericht.[34][36]
Flora
Makrofossilien von Pflanzen sind im Ouled-Abdoun-Becken äußerst selten erhalten. Bisher wurde lediglich der silifizierte Holzrest von Pterocarpus aus der Gruppe der Hülsenfrüchtler berichtet, der womöglich in das Untere Eozän datiert. Die immergrünen Bäume sind heute im tropischen Afrika und Asien verbreitet.[37] Sehr reichhaltig vertreten und intensiv studiert wurde dagegen das Phytoplankton in Form von Dinoflagellaten, Mikroalgen mit Größen von 50 bis 500 μm. Bei Analysen einer Bohrung aus dem südlichen Teil des Beckens ließen sich allein 90 Arten aus 50 Gattungen feststellen. Diese können in vier Abfolgen gegliedert werden, die von Beginn bis zum Ende des Maastrichtiums reichen. In der untersten dominiert Trichodinium und stellt fast 50 % der Funde, daneben kommen Palaeohystrichophora und Chatangiella relativ häufig vor. Das hohe Alter dieser Folge wird vor allem durch Odontochitina bestätigt. Die zweite Folge charakterisiert sich wiederum über sehr hohe Anteile an Isabelidinium, gefolgt von Svalbardella und Trithyrodinium. Die dritte Folge wird durch Deflandrea, Fibradinium und Muratodinium angezeigt, wobei Trithyrodinium noch weitgehend erhalten bleibt. Sehr artenarm zeigt sich die vierte Folge. Hier treten vor allem Isabelidinium, Spiniferite und lmpagidinium auf. Dass die gesamte Dinoflagellaten-Folge hier dem Maastrichtium angehört, lässt sich unter anderem an dem beständigen Auftreten von Dinogymnium ableiten.[38] In den höheren Lagen des Paläogens konnten in weiteren Untersuchungen unter anderem Isabelidinium, Liesbergia, Spinidinium und Manumiella beobachtet werden. Alle vier Formen sind im Paläozän anwesend, aus dem Eozän ist dagegen Bitubericysta registriert.[39][40][36]
Wirbellose
Die Wirbellosen werden hauptsächlich durch die Weichtiere vertreten. Unter diesen sind die Muscheln besonders häufig und verteilen sich in ihrem größten Anteil auf die Autolamellibranchiata. Dazu gehören etwa die Cucullaeidae, die mit Cucullaea bereits in der frühen Oberkreide im Ouled-Abdoun-Becken vorkommen. In der eigentlichen Phosphatserie tritt Cucullaea nicht auf, ist aber später im Lutetium wieder nachweisbar. Flügelmuscheln werden im Maastrichtium unter anderem durch Heligmopsis angezeigt. Austern bilden wiederum mit Liostrea teils dichte Lagen im Paläozän. Gleiches gilt auch für Venericardia aus der Gruppe der Trapezmuscheln. Schnecken dem gegenüber sind im Maastrichtium und im frühen Paläogen eher selten belegt, kamen aber noch im Beginn der Oberkreide sehr vielfältig und häufig vor. Möglicherweise hängt die Verarmung der Schneckenfauna mit den Besonderheiten der Phosphatogenese und dem hohen Anteil an Kalkphosphaten zusammen, da einige Formen dieser Zeit ein extrem dickes Gehäuse ausgebildet haben. Nachgewiesen wurden unter anderem die Ampullinidae mit Ampullinopsis und die Turmschnecken mit Turritella. Im späteren Lutetium haben vor allem die Thersiteidae mit Thersitea und Hemithersitea eine wichtige stratigraphische Bedeutung. Unter den Kopffüßern können die Perlboote hervorgehoben werden, die beispielsweise mit Cimomia als Vertreter der Hercoglossidae im Unteren Eozän präsent sind.[36][41]
Weitere Wirbellose sind mit den Strahlentierchen, den Foraminiferen und den Muschelkrebsen anwesend. Diese bilden sehr häufig den Kernbestand der Phosphatkörner. Dadurch ist ihre Überlieferung aber weitgehend schlecht und sie entziehen sich so weitgehend einer genauen Bestimmung.[36][41]
Knorpelfische


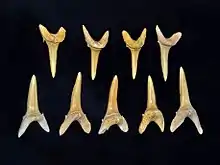

Den weitaus größten Anteil aller Fossilfunde haben die Knorpelfische mit den Hai- und Rochenartigen. Deren Fundmaterial beschränkt sich aber naturgegebenermaßen auf Zähne und vereinzelt Wirbel. Unter den Haien sind wenigstens acht größere Linien nachweisbar. Hierzu gehören ursprüngliche Gruppen wie die Grauhaiartigen, die Nagelhaie, die Sägehaiartigen, die Dornhaiartigen und die Engelhaiartigen, die allesamt zu den Squalomorphii zusammengefasst werden. Bis auf die Nagelhaie, die im Ouled-Abdoun-Becken bisher nur aus dem Paläogen nachgewiesen sind, kommen alle anderen Formen auch im Maastrichtium vor. Zu den Nagelhaien zählt unter anderem Heptranchias. Die Gattung schließt heute noch den Spitzkopf-Siebenkiemerhai ein, welcher den Meeresgrund bewohnt. Fossil ist sie über etwa 1,2 cm lange Zähne belegt, die typischerweise eine Hauptspitze und mehrere in Reihe angeordneter kleinerer Nebenspitzen aufweisen.[42] Die Stierkopfhaiartigen, die Ammenhaiartigen, die Makrelenhaiartigen und die Grundhaiartigen fassen die moderneren Haie der Galeomorphii zusammen. Von diesen sind aber nur die letzten drei sehr häufig fossil im Ouled-Abdoun-Becken anzutreffen. Allein 14 Gattungen stellen die Ammenhaiartigen. Palaeorhincodon kam dabei aus den Paläogen-Schichten zu Tage. Er wird als eine Vorgängerform des Walhais angesehen, dem größten Fisch der Gegenwart und heute einziges Mitglied der Rhincodontidae. Funde sind aber eher selten, bisher wurden etwas mehr als 40 Zähne entdeckt. Diese sind an der Hauptspitze bis zu 4 mm hoch. Die Hauptspitze wiederum ist stark nach hinten oder zur Seite gekrümmt, seitlich flankiert sie je eine kleinere Spitze.[43] Wesentlich häufiger treten die Ammenhaie selbst auf, unter anderem mit Ginglymostoma.[44] Andere Formen verweisen auf die Bambushaie, die Blindhaie beziehungsweise auf die Teppichhaie. Bei letzteren wurde Squatiscyllium rund 1,5 m lang und war damit etwa halb so groß wie heutige Formen. Seine Reste datieren in das Paläozän. Fast doppelt so große Ausmaße erreichten die Sandhaie, die im Paläozän eine hohe Diversität entfalteten, unter anderem mit der Gattung Odontaspis. Carcharias wurde ursprünglich ebenfalls in die Gruppe gestellt, zählt aber heute zu den Sandtigerhaien. Die Gattung ist nur im Maastrichtium dokumentiert. Sowohl die Sandhaie als auch die Sandtigerhaie stehen innerhalb der Makrelenhaiartigen, die mit insgesamt fast zwei Dutzend Gattungen die umfangreichste Gruppe der Haie im Ouled-Abdoun-Becken darstellen. Sie schließen mit den Koboldhaien und den ausgestorbenen Otodontidae die größten räuberischen Fische ein. So erreichte wohl der im Maastrichtium auftretende Koboldhai Scapanorhynchus aufgrund seiner rund 7 cm hohen vorderen Zähne eine Länge von rund 5 m. Vergleichbare Körperausmaße werden für den Otodontidae-Vertreter Otodus aus dem Paläogen angenommen. Ebenfalls überlieferte Wirbel besitzen mitunter über 10 cm Durchmesser. Sie bestehen aus ringförmigen Schichten, die laut Isotopenuntersuchungen jährliche Wachstumsraten angeben, wodurch ein Individuum zur Zeit seines Todes auf rund 19 Jahre bestimmt werden konnte.[45] Die Gattung enthält auch den deutlich jüngeren und riesigen Megalodon. Darüber hinaus sind die Fuchshaie präsent, dessen weniger als 1 cm langen Zähne seitlich neben der Hauptspitze mit zusätzlichen kleinen, niedrigen Spitzen ausgestattet waren.[42] Als sehr häufig erwiesen sich die ausgestorbenen Anacoracidae. Hier ist Squalicorax hervorzuheben, ein 3 bis 4 m langes Tier, dessen seitlich gepressten und am Rand fein gezackten Zähne etwas denen des nicht näher verwandten Tigerhais ähneln und auf einen effektiven Räuber deuten lassen. Vermutlich ernährte er sich von Schildkröten. Die mehrere Arten einschließende Gattung ist auf das Maastrichtium beschränkt.[46] Gleiches gilt für die ebenfalls nicht mehr bestehenden Cretoxyrhinidae mit Cretolamna. In der Größe Squalicorax ähnelnd, finden sich dessen Zähne häufig in Resten von Meeresreptilien, so dass diese Form wohl aasfressend war. Eine weitere erloschene Gruppe sind die Serratolamnidae, deren Typusform Serratolamna wiederum aus dem Maastrichtium vorliegt. Zum Schluss seien noch die Grundhaiartigen erwähnt, die hauptsächlich im Paläogen sehr häufig auftreten. Von ihnen sind fast genauso viele Gattungen wie bei den Makrelenhaiartigen dokumentiert. Zwei Charakterformen bilden hier Abdounia und Physogaleus aus der Gruppe der Requiemhaie. Äußerst divers erscheinen die Katzenhaie, die allerdings meist weniger als 2 m lang wurden. Belegt sind unter anderem Pteroscyllium, Scyliorhinus und Premontreia. Andere Formen wie Palaeogaleus und Triakis verweisen auf das Vorkommen der Glatthaie.[1][43][34]


Die Rochen als zweitgrößte Fossilgruppe des Ouled-Abdoun-Beckens sind mit den Rajiformes, den Rhinopristiformes, den Zitterrochenartigen und den Stechrochenartigen nachweisbar. Unter diesen vier Großgruppen nehmen letztere mit über 30 beschriebenen Gattungen und mehr als doppelt so vielen Arten eine dominante Stellung ein und machen gut die Hälfte des Fundmaterials aus. Die Stechrochen treten bereits im Maastrichtium auf, sind aber vor allem im Unteren Paläogen gut vertreten. Viele Formen aus dem Ouled-Abdoun-Becken existieren heute nicht mehr und gehören wahrscheinlich in ein weiteres Beziehungsumfeld. Darunter fällt etwa Coupatezia, dessen kleine Zähne mit niedrigen und flachen Zahnkronen ausgestattet waren.[47] Etwa im gleichen Zeitraum entwickelten die Schmetterlingsrochen eine hohe Vielfalt. Aufgefundene Formen werden durch die heute noch bestehende Gattung Gymnura und der ausgestorbenen Form Ouledia repräsentiert. Die spitzen Zähne waren teilweise nur rund 1 bis 2 mm hoch, was einen auffallenden Kontrast zu ihrer Körperspannweite von 1,5 m und mehr bildet. Gymnura ist zumeist aus dem frühesten Paläozän belegt, Ouledia trat etwas später auf.[48][49] Zu den Myliobatidae gehören Formen wie Lophobatis, Leidybatis oder Pseudaetobatus. Sie waren mit hohen, flachen Zähnen bestückt und ernährten sich wohl überwiegend von hartschaliger Beute wie Weichtieren oder Krebsen. Rhombodus und Dasyrhombodus bilden die Gruppe der Rhombodontidae, welche bereits zum Ende der Kreidezeit verschwand. Sie sind dadurch nur im Maastrichtium verbreitet und kommen dort mit hoher Anzahl vor. In ihrer Zahnstruktur ähneln sie ein wenig den Myliobatidae, weisen aber in der Regel rautenförmiger Zahnflächen auf. Auch die Ernährungsweise dürfte vergleichbar gewesen sein.[50][51] Sehr selten wurden bisher Teufelsrochen aufgefunden, aus dem Paläogen stammt lediglich Rhinoptera. Häufiger treten die Kuhnasenrochen in Erscheinung, hauptsächlich mit Burnhamia, das im Verlauf des Paläogens eine deutliche Größenreduktion durchläuft. Aus der Gruppe der Rhinopristiformes sind die Sägerochen zu nennen, die im Ouled-Abdoun-Becken erst im entwickelten Paläogen auftauchen, aber nur von ihren vordersten Zähnen dokumentiert sind. Eine äußerliche Übereinstimmung mit den Sägerochen, aber keine verwandtschaftliche Nähe, besitzen die ausgestorbenen Sclerorhynchidae, im Ouled-Abdoun-Becken etwa durch Gebissteile von Schizorhiza dokumentiert. Andere erloschene Gruppen wie die Hypsobatidae und die Parapalaeobatidae sind nur über einzelne Zähne belegt.[52] Bei den Rhinobatidae und den Rhynchobatidae, erstere mit Funden aus dem Maastrichtium, letztere im Paläogen anwesend, bestehen vor allem noch systematische Schwierigkeiten, die momentan keine genaue Zuweisung erlauben. Ebenfalls selten treten Zitterrochenartige in Erscheinung. Beschrieben wurden bisher Narcine aus der Gruppe der Narcinidae und Eotorpedo aus der Gruppe der Zitterrochen. Die geringe Anzahl der Zähne beider Vertreter datiert in das Paläogen.[1][43][34]
Keiner speziellen Gruppe der Hai- und Rochenartigen zuordenbar ist Odontorhytis, eine kleine Form, die ursprünglich aus Südwestafrika beschrieben wurde, seit dem Beginn der 1980er Jahre jedoch auch aus dem Ouled-Abdoun-Becken vorliegt. Die gleichförmigen Zähne waren stark abgeplattet.[42] Einer dritten Gruppe innerhalb der Knorpelfirsche ist mit den Seekatzen zu verzeichnen. Ihr Nachweis ist aber äußerst selten. Neben einzelnen Zähnen von Edaphodon tritt ein Rest der stachelartigen Rückenflosse auf, die am Rand mit feinen Zacken besetzt ist. Beide Funde gehören den Paläogen an.[44][53][34]
Knochenfische

Die Knochenfische sind weitaus weniger umfangreich als die Knorpelfische. Es kommen acht Großgruppen vor, von denen die Knochenzünglerartigen, die Kugelfischartigen und die Ichthyodectiformes weitgehend nur eine Gattung vorweisen können. Als umfangreicher erwiesen sich die Pflasterzahnfische. Zu nennen sind hier die Serrasalmimidae, unter denen mit Eoserrasalmimus und Serrasalmimus zwei piranhaartige Formen beschrieben wurden, beide jeweils über Gebissreste. Sie sind mit den eigentlichen Piranhas nicht näher verwandt. Ihre Zähne wiesen scharfe Schneidkanten auf und dienten eventuell zur Erbeutung von weicher Beute, etwa kleinen Fischen und Kopffüßern. Sowohl Eoserrasalmimus als auch Serrasalmimus gehören dem frühen Paläozän an.[54] Eine dritte Form der Pflasterzahnfische wird mit Phacodus bereits aus dem Maastrichtium angegeben. Die Eidechsenfischartigen sind hingegen mit Enchodus und Stratodus belegt, die jeweils nur im Maastrichtium auftreten. Von Enchodus wurden Zahn- und Schädelreste gefunden. Zu den Grätenfischen zählen Albula und Phosphonatator. Erstere Form besteht noch heute, letztere ist nur fossil nachgewiesen. Aus dem Ouled-Abdoun-Becken wurde aus den Maastrichtium-Lagen ein zerquetschter Schädel von gut 36 cm Länge geborgen.[55] Die variantenreichste Gruppe findet sich in den Barschartigen und ihrer unmittelbaren Verwandtschaft. Unter ihnen treten Sägebarsche, Meerbrassen, Barrakudas, Makrelen und Thunfische, Haarschwänze und Schwertfische auf. Das Belegmaterial der meisten Formen setzt sich aus einzelnen Zähnen zusammen. In seltenen Fällen wie bei Phosphichthys aus der Gruppe der Sägebarsche sind Schädelreste erhalten oder wie bei Xiphiorhynchus und Cylindracanthus als Vertreter der Schwertfische kommen Teile des verlängerten Oberkiefers vor. Die letzte Gruppe bilden die Tarpunartigen. Die Tarpune selbst werden durch Protarpon repräsentiert, dessen Fundmaterial aus zwei unvollständigen Schädeln besteht, die jeweils mit dem Hirnabschnitt erhalten sind. Der größere der beiden ist rund 19 cm lang.[56] Weitere Gruppenmitglieder gehören zu den Phyllodontidae. Sowohl die Barschartigen als auch die Tarpunartigen sind mit wenigen Ausnahmen aus dem Paläogen überliefert.[1][34]
Schuppenkriechtiere und Plesiosaurier

Die Schuppenkriechtiere treten relativ zahlreich auf, von den heute noch bestehenden Linien sind aber nur die Schlangen und Warane belegt. Zu ersteren zählt Palaeophis, von der mehrere Teilskelette aufgefunden wurde, einige davon mit mehr als 40 zusammenhängenden Wirbeln. Vollständige Exemplare sind nicht bekannt, aber es wird vermutet, dass sich das Skelett aus mehreren hundert Wirbeln zusammensetzt. Die Ausmaße der einzelnen Wirbel, die jene heutiger Netzpythons um fast das Doppelte übertreffen, verweisen auf ein extrem großes Tier von 6 m Länge und mehr. Die kaum gepressten Wirbel und fehlende Pachyostose lassen an ein an Flachwasserbedingungen und küstennahe Lebensräume angepasstes Tier denken. Funde kamen sowohl in den Lagen des Maastrichtiums als auch des Paläogens zum Vorschein.[57][58] Dem gegenüber bleibt Pachyvaranus aus der näheren Verwandtschaft der Warane auf die ältesten Ablagerungen beschränkt. Auch von diesem sind einzelne Teilskelette mit Wirbeln und Rippen geborgen worden. Rekonstruiert besaß das Tier möglicherweise eine Körperlänge von rund 1,5 m. Auffallende Knochenverdichtungen bei Pachyvaranus sind als deutliche Anzeichen an ein Leben im offenen Meer zu interpretieren.[59][60][34]



Die umfangreichste und vielfältigste Gruppe der Schuppenkriechtiere bilden jedoch die Mosasaurier, teils riesige Tiere mit aalartig verlängertem Warankörper und großem Schädel. Alle im Ouled-Abdoun-Becken aufgefundenen Mosasaurier gehören der jüngsten Entwicklungslinie an und waren vollständig an das Wasserleben angepasst. Ihre Kennzeichen sind ausgebildete Flossen mit überzähligen Gliedern und ein Becken, das nicht mit der Wirbelsäule verbunden war. Mit 3 bis 4 m Gesamtlänge einen der kleinsten und zudem den urtümlichsten Vertreter repräsentiert Halisaurus. Wie bei den stammesgeschichtlich jüngeren Mosasauriern nahm die Wirbelsäule im Schwanzbereich einen nach unten gekrümmten Verlauf an. Eine dort ausgebildete halbmondartige Schwanzflosse diente dabei als Antrieb im Wasser, wodurch die Tiere bereits reine Meeresbewohner darstellen. Von Halisaurus sind mehrere Teilskelette und Schädel dokumentiert. Das Gebiss setzt sich aus schlanken, nadelspitzen und nach hinten gekrümmten Zähnen zusammen.[61][62] In enger Beziehung zu Halisaurus steht Pluridens, von dem mehrere Schädel mit assoziierten Unterkiefern bekannt sind. An der langen Schnauze waren mehrere Öffnungen für Nervenbahnen ausgebildet. In Verbindung mit den deutlich kleinen Augenfenstern orientierten sich die 5 bis 6 m langen Tiere wohl eher tastend und riechend im Wasser, vergleichbar heutigen wasserlebenden Schlangen. Die zahlreichen kleinen und spitzen Zähne implizieren eher kleine Beute als Fang.[63] Einer anderen Verwandtschaftsgruppe innerhalb der Mosasaurier gehört der rund 6 m lange Gavialimimus an, dem Schädel und Skelettteile sowie zahlreiche Einzelzähne zugeordnet werden. Der 90 cm messende Schädel wies eine extrem ausgedehnte Schnauze auf, die gut zwei Drittel der Gesamtlänge ausmachte. Dies erinnert stark an die Gaviale, worauf auch der Gattungsname hinweist.[64] Alle anderen Formen aus dem Ouled-Abdoun-Becken sind den Mosasaurinae zuzurechnen, deren Schnauze zumeist kürzer war und etwa die Hälfte der Schädellänge einnahm. Ihr bekanntester Angehöriger ist Mosasaurus, der hier zwar mit wenigstens zwei Arten, insgesamt aber eher selten in Erscheinung tritt. Er ist über Einzelzähne und zwei fragmentierte Schädel überliefert. Dessen Ausmaße von 100 cm verweisen auf eine Gesamtlänge der Tiere von knapp 10 m.[65][66] Nur halb so groß wurde Eremiasaurus, eine vergleichsweise häufige Form. Ihre Beschreibung beruht auf einem nahezu vollständigen, allerdings in acht Teilblöcke zerfallenen Skelett. Wie Mosasaurus besitzt Eremiasaurus hohe spitze und scharfe Zähne, die aber bei letzterem eine variierenden Gestalt haben.[67] Den größten Vertreter der Mosasaurier stellte Prognathodon, der mit mehreren Arten im Ouled-Abdoun-Becken ausgewiesen ist. Sie können anhand der Form der Zähne unterschieden werden, die von leicht spitz bis deutlich abgerundet reichen. Allein der Schädel misst bis zu 150 cm in der Länge. Daneben kommen noch zwei weitere, näher miteinander verwandte Gattungen vor, einerseits Globidens, andererseits Carinodens. Bei beiden handelt es sich um sehr kleine Mitglieder der Mosasaurier, die weitgehend von isolierten Zähnen und einzelnen Schädelfragmenten bekannt sind. Sie heben sich von den anderen Mosasaurieren des Ouled-Abdoun-Beckens durch ihre charakteristisch stumpf endenden Zähne ab, die bei ersterer Form kugelig, bei letzterer hingegen seitlich abgeplattet sind. Die Gestaltung der Zähne befürwortet somit eine spezialisierte, auf Weichtiere basierende Ernährung. Während Carinodens weitgehend rar ist, kommt Globidens wesentlich häufiger vor.[11][68][69] Innerhalb der Mosasaurier zeigt Xenodens als weitere und wohl Carinodens näher stehende Form eine ungewöhnliche Anpassung. Belegt ist die Form mit einem Oberkieferfragment, dem noch vier Zähne anhaften, gut doppelt so viele Zahnfächer zeigen weitere Zahnpositionen an. Die Zähne der möglicherweise nur rund 1,6 m langen Tiere sind seitlich stark gepresst, asymmetrisch spitz und stehen eng im Kiefer, wodurch im Gebiss eine durchgehende Schneidkante entsteht. Diese eher an einige Haie erinnernde und von Schuppenkriechtieren sonst nicht bekannte Gebissstruktur ermöglichte es Xenodens, Fleischbrocken aus größeren Beutetieren herauszubeißen, die entweder aktiv gejagt oder aasfressend gesucht wurden.[70][71][34]

Im Vergleich zu den Mosasauriern sind die Plesiosaurier sehr selten im Ouled-Abdoun-Becken. Beide Gruppen sind nicht näher verwandt, worauf der abweichende Körperbauplan mit den großen paddelartigen Gliedmaßen und der kurze Rumpf hindeuten. Sie teilen jedoch die marine Lebensweise, allerdings bewegten sich die Plesiosaurier eher wie heutige Meeresschildkröten im Wasser gleitend fort mit den Flossen als Hauptantrieb und weniger undulierend über die Wirbelsäule wie andere aquatische Reptilien. Ein vollständiges Skelett und einzelne Schädelteile gehen auf Zarafasaura aus der Gruppe der Elasmosauridae zurück. Wie alle späten Plesiosaurier besaß auch dieser einen extrem langen Hals, der über die Hälfte der gesamten Körperlänge einnahm. Darauf spielt auch der Gattungsname an. Dieser ist arabischen Ursprungs und leitet sich von der Bezeichnung für die Giraffen ab. Die Wirbelsäule umfasst mehr als 120 Wirbel, davon beansprucht der Hals allein 56. Der Schädel hingegen war ausgesprochen kurz.[72][73] Weitere postcraniale Skelettelemente gehören zu einer noch unbeschriebenen Form, deren Hals aus 64 Wirbeln besteht. Das Material schließt Jung- und ausgewachsene Tiere ein und stammt jeweils von der gleichen Fundstelle. Der Umstand führt zu der Annahme, dass die unterschiedlichen Altersstadien der Plesiosaurier offensichtlich die gleichen Nahrungsressourcen nutzten.[74][75][34]
Schildkröten und Krokodile
Das umfangreiche Material der Schildkröten des Ouled-Abdoun-Beckens reicht stratigraphisch vom Maastrichtium bis zum Paläogen. Es verteilt sich auf rund ein Dutzend Gattungen aus vier Familien. Dabei kommen sowohl Halswender- als auch Halsberger-Schildkröten vor. Erstere werden lediglich durch die ausgestorbene Gruppe der Bothremydidae repräsentiert. Hierbei handelt es sich um eine kosmopolitische und sehr variantenreiche Gruppe, die von der Unterkreide bis zum Miozän vorkam und zahlreiche ökologische Nischen besetzte. Sie schließen kleine bis große Arten von 30 bis 150 cm Körperlänge ein. Im Ouled-Abdoun-Becken verfällt gut die Hälfte der beschriebenen Gattungen auf die Bothremydidae, die fast ausschließlich auf Schädelfunden basieren. Alle diese Formen sind paläogenen Alters, allerdings liegen auch oberkreidezeitliche Reste vor, wurden bisher aber noch nicht näher untersucht.[76] Innerhalb der Bothremydidae bilden Ummulisani, Phosphatochelys und Taphrosphys eine engere Verwandtschaftsgruppe. Erstere Gattung kann auf drei, letztere beiden auf je zwei dokumentierte Schädel verweisen. Als Besonderheit trägt Ummulisani eine paarige Hornbildung auf dem Präfrontale, die ursprünglich wohl mit Hornschuppen überzogen war. Das Merkmal ist einzigartig unter den Schildkröten und kommt auch bei den unmittelbar verwandten Formen nicht vor. Weitere Gattungen aus dem Beziehungsumfeld finden sich mit Rhothonemys und mit Labrostochelys, letztere war im Vergleich zu den anderen sehr schmalschnauzig. Die Schädellängen aller Gattungen variieren zwischen 6,8 und 20 cm mit der kleinsten bei Phosphatochelys und der größten bei Ummulisani. In eine andere verwandtschaftliche Gemeinschaft gehören Bothremys und Araiochelys. Deren Schädelgrößen liegen im Bereich der Maße von Phosphatochelys. Lediglich von Araiochelys sind als einzige Form der Bothremydidae des Ouled-Abdoun-Beckens auch mehrere Teile des Rückenpanzers geborgen worden, von Ummulisani ist darüber hinaus auch ein Bauchpanzer dokumentiert.[77][78][79][34]
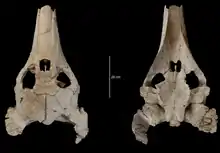
Fast genauso vielfältig sind die Halsberger-Schildkröten, die wohl erst im Verlauf der Oberkreide aus den nördlichen Festlandsbereichen des heutigen Eurasiens über die Tethys in Afrika einwanderten.[80] Im Ouled-Abdoun-Becken sind sie sowohl aus dem Maastrichtium als auch aus dem Paläogen überliefert und ähnlich wie bei den Halswender-Schildkröten lediglich anhand der Schädel bekannt. Die Dermochelyoidae werden heute noch durch die Lederschildkröte repräsentiert. Im Verlauf ihrer Stammesgeschichte brachten sie einige ungewöhnliche Formen hervor. Dazu gehört Ocepechelon aus dem Maastrichtium, die möglicherweise eine der größten Schildkröten der Erdgeschichte war. Ihr etwa 70 cm langer Schädel weist eine röhrenartig verlängerte Schnauze auf, mit der die Tiere wohl, vergleichbar zu heutigen Buckelwalen, ihre Nahrung aus dem Meerwasser saugten.[76] Anders hingegen das etwa gleich alte Alienochelys. An dessen breiten und gut 55 cm langen Schädel ist ein massiver, halbkreisförmig gebogener und platter Unterkiefer ausgebildet. Dieser erfüllte wohl eine besondere Brechfunktion während der Nahrungsaufnahme.[81] Eine weitere Form der Lederschildkröten ist nur über einzelne Fragmente des Bauchpanzers belegt und wurde bisher noch nicht benannt.[82] Verschiedene andere Fossilreste lassen sich den Meeresschildkröten zuordnen. Es handelt sich überwiegend um basale Vertreter, die als besonderes Kennzeichen über einen zweiten Gaumen verfügten. Zudem waren sie alle relativ klein mit Schädellängen zwischen 9 und 23 cm. Gemäß ihrem häufig dreieckigen Unterkiefer mit verbreiterter Schabefläche ernährten sie sich weitgehend von Weichtieren. Einige Formen besaßen vergleichsweise kurze Gliedmaßen, wodurch sie nur bedingt an das Meeresleben angepasst waren und wahrscheinlich eher küstennah lebten. Hierzu zählt Euclastes, eine mittelgroße Meeresschildkröte, die sowohl in den Lagen des Maastrichtiums wie des Paläogens anwesend ist und mehrere Arten einschließt (darunter eine mit einer ursprünglichen Stellung in der Gattung Osteopygis,[83] die sich aber später als Chimäre herausstellte[84]). Sie zeichnet sich durch einen breit-dreieckigen und flachen Schädel aus.[85] Des Weiteren wurde Argillochelys beschrieben, die bisher größte bekannte Meeresschildkröte aus dem Ouled-Abdoun-Becken. Ihre stumpfe Schnauze setzt sich leicht vom Schädel ab.[86] Dagegen zeigt sich bei Tasbacka und Puppigerus, zwei sehr kleinen Angehörigen der Gruppe, der Schädel deutlich langgestreckt und die Schnauze dadurch zugespitzter. Erstere könnte bereits stärker an ein Leben im offenen Meer angepasst gewesen sein.[87][88] Als weitere Gruppe ist mit Brachyopsemys eine Gattung der Sandownidae dokumentiert. Die Sandownidae sind ausgestorben, stehen aber in relativer verwandtschaftlicher Nähe zu den Meeresschildkröten. Der nur 11 cm lange und sehr breite Schädel ist ebenfalls flach, aber vorn deutlich gerundet und mit einem sekundären Gaumen ausgestattet.[89][34]


Die Krokodile sind ebenfalls mit rund einem Dutzend Gattungen vertreten, die sich auf mehrere Linien verteilen. Die umfangreichste bilden die heute erloschenen Dyrosauridae, eine basale Formengruppe. Sie gehören zu den ersten Funden, die im Ouled-Abdoun-Becken wissenschaftlich bekannt wurden. Es handelt sich um ausschließlich meeresbewohnende Formen, die bereits im Maastrichtium anwesend waren, aber im Paläogen ihre größte Vielfalt erreichten. So auch in Marokko, wo alle dyrosauriden Krokodile des Ouled-Abdoun-Beckens dem Paläogen angehören. Als Besonderheit der Gruppe kann die teils extrem lange Schnauze hervorgehoben werden, die als Anpassung an fischhaltige Nahrung anzusehen ist. In ihrer weitgehend gleichförmigen Bezahnung, die pro Seite jeweils gut zwei Dutzend Zähne umfasste, war der siebente Zahn des Unterkiefers extrem verkleinert und dicht am achten positioniert. Der vierte hingegen zeigte sich deutlich vergrößert und passte in eine entsprechend große Zahnlücke im Oberkiefer. Die auffällig lange Schnauze zeigt sich unter anderem bei der Typusform Dyrosaurus sowie bei Atlantosuchus, bei denen sie gut 75 beziehungsweise 79 % der Schädellänge beansprucht. Von Dyrosaurus sind mehrere vollständige Individuen aufgefunden worden, darunter aber nur wenigen Jungtieren. Von der relativ langbeinigen Form erhielten sich zudem Teile der Schuppenpanzerung des Rückens. Deren seitlich glatten Osteoderme besaßen zahlreiche typische Eintiefungen auf der Oberfläche. Atlantosuchus wiederum wird durch einen aufgefundenen Schädel mit anhaftenden Teilen der Halswirbelsäule repräsentiert.[90][91] Im Vergleich zu diesen beiden wirkte Chenanisuchus, dokumentiert über zwei Schädel, mit rund 63 % Schnauzenanteil bezogen auf die Schädellänge eher kurzschnauzig. Hinsichtlich der Körpergröße war Chenanisuchus auch deutlich kleiner als Dyrosaurus und Atlantosuchus. Während bei diesen der Schädel jeweils 90 bis 100 cm lang ist, misst er bei Chenanisuchus nur gut 58 cm. Seine Körperlänge lag daher schätzungsweise bei rund 4,5 m.[92] In der Schnauzengestaltung steht Arambourgisuchus eher intermediär zwischen diesen Extremformen. Von der Gattung liegt ebenfalls weitgehend Schädelmaterial vor.[93] Andere dyrosauride Krokodile aus dem Ouled-Abdoun-Becken sind Hyposaurus und Rhabdognathus. In einer verwandtschaftlichen Nähe finden sich die Pholidosauridae, in die die Dyrosauridae ursprünglich eingeschlossen waren. Zu diesen wird ein Individuum geordnet, welches nur von Körperskelett bekannt ist und noch nicht wissenschaftlich benannt wurde. Bei diesem kleinen, wohl nur rund 2 m langen Tier handelt es sich unter Ausschluss der Dyrosauridae aus den Pholidosauridae um deren jüngsten Vertreter, da die meisten anderen Formen bereits zu Beginn der Oberkreide verschwanden. Wahrscheinlich war die Form an ein Leben in Flüssen angepasst.[94]
Neben diesen ausgestorbenen Linien sind auch Reste von Angehörigen heute noch bestehender Gruppen entdeckt worden. Hierbei handelt es sich weitgehend um den Verwandtschaftskreis der Gaviale. Diese ebenfalls langschnauzigen Krokodile, weisen zuzüglich einiger anderer Merkmalen aber Unterschiede im Aufbau des Gaumens auf. Hervorzuheben ist auch ihr im Vergleich mit anderen modernen Krokodilen weitgehend homodontes Gebiss aus gleich großen Zähnen. Die frühesten Gaviale sind im Ouled-Abdoun-Becken mit Ocepesuchus bereits im Maastrichtium präsent, dessen Belegmaterial aus einem stark zerquetschten Schädel besteht. Es ist damit einer der ältesten Fossilnachweise moderner Krokodile in Afrika. Es wird angenommen, dass die Tiere eventuell teilweise oder vollständig auf Flüsse beschränkt waren, da in den Meeren zu dieser Zeit andere Reptilien dominierten.[95] Weitere Gaviale traten dann erst im Paläogen auf. Aufgefundene Formen werden zu Agrochampsa und Thoracosaurus geordnet. Von beiden ist erstere mit wenigstens vier Schädeln überliefert, das Holotyp-Exemplar misst rund 43 cm, die Schnauze davon allein 31 cm. Als auffällige Eigenschaft der Tiere knickt der vordere Teil der Schnauze deutlich nach unten ab.[96][97] Innerhalb der Gaviale können die Tomistominae abgesetzt werden, deren Bezahnung variabler ausfällt. Maroccosuchus gehört zu den ältesten Repräsentanten dieser Linie, die heute noch den Sunda-Gavial stellt. Die Ausweisung der Form basiert auf zahlreichem Schädelmaterial aus dem Ouled-Abdoun-Becken. Im Vergleich zum Sunda-Gavial ist sein Schädel aber deutlich robuster gebaut, wodurch die Schnauze insgesamt breiter wirkt.[98][34]
Flugsaurier, Dinosaurier und Vögel
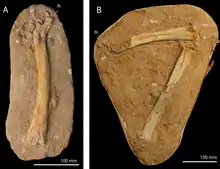

Flugsaurier brachten die größten flugfähigen Tiere der Erdgeschichte hervor. Sie sind äußerst rar im Ouled-Abdoun-Becken und waren bis Anfang der 2000er Jahre gar nicht belegt. Seitdem kamen mehr als 200 Fundstücke zum Vorschein. Die ersten Funde, einzelne Wirbel, wurden zu Phosphatodraco gestellt, ein vergleichsweise großes Tier mit rekonstruiert rund 5 m Flügelspannweite.[99] Später ließen sich noch weitere Wirbel der Gattung zuordnen. Die röhrenförmigen und stark verlängerten Wirbelkörper stellen die Form zu den Azhdarchidae. Zur gleichen Familie gehört ein bisher unbenannter Vertreter, dessen Fragment der Elle auf ein deutlich größeres Tier mit 9 m Flügelspannweite verweist. Weiterhin sind mit Alcione, Simurghia und Barbaridactylus auch drei Vertreter der Nyctosauridae dokumentiert. Die Gruppe kennzeichnen vierfingrige Hände und ein extrem kräftiger, beilartig aufragender Muskelkamm am Oberarmknochen. Von den drei Formen weist erstere das umfangreichste Material auf. Es besteht aus einem Teilskelett und zusätzlichen Flügelelementen. Basierend auf dem 9,3 cm langen Oberarm, handelt es sich um einen kleinen Angehörigen der Familie. Auffallend sind die verkürzten Flügelknochen. Sie könnten eine Anpassung an einen kräftigen, schnellen Flügelschlag sein oder für ausgedehntere Tauchgänge sprechen. Von den beiden anderen Vertretern sind weitgehend nur einzelne Gliedmaßenknochen bekannt. Aufgrund ihrer deutlich größeren Oberarmknochen übertrafen sie Alcione teilweise um mehr als das Fünffache an Größe. Tethydraco wiederum repräsentiert die Pteranodontidae, welche sich durch einen langen Hinterhauptskamm auszeichnen. Ebenfalls nur durch einzelne Arm- und Beinknochen überliefert, besaß das Tier eine Flügelspannweite von rund 5 m.[31]


Vergleichbar den Flugsauriern finden sich landbewohnende Dinosaurier nur selten im Fossilbericht des Ouled-Abdoun-Beckens, auch hier stammen die ersten Funde aus den Beginn der 2000er Jahre. Zu den wenigen Hinweisen gehört ein Hinterbein eines kleinen Sauropoden, das aber bisher keiner bestimmten Form zugewiesen werden konnten. Möglicherweise ist sie innerhalb dieser teils riesigen, vierfüßigen und langhalsigen Pflanzenfresser als ein basaler Vertreter der Titanosauria eingrenzbar.[100] Darüber hinaus zeigen ein vorderes, hoch gestaltetes Unterkieferfragment und mehrere Einzelzähne das Auftreten der Theropoda an, zweibeinig laufenden Fleischfressern. Die Zähne, von denen allein zehn noch im Unterkiefer stecken, sind dreieckig-spitz geformt und mit scharfen, fein gezackten Seitenkanten ausgestattet. Die Reste wurden als zu Chenanisaurus gehörig beschrieben, einem rund 7 bis 8 m langen Vertreter der Abelisauridae.[101][30] Mit Ajnabia ließ sich auch der erste Vertreter der Hadrosauridae aus Afrika belegen. Diese durch ihre entenschnabelartige Schnauzenstruktur charakterisierte Gruppe ist sonst nur von den nördlichen Festlandsmassen von Laurasia bekannt. Ihr Auftreten im Ouled-Abdoun-Becken und somit im nördlichen Gondwana setzt eine Einwanderung über die Tethys voraus, weswegen die neue afrikanische Form auch mit dem Artepitheton odysseus nach dem gleichnamigen Held der griechischen Mythologie bedacht wurde. Bisher kam ein rund 16 cm langes Oberkieferfragment zu Tage, das rund 23 Zahnfächer besitzt und auf ein etwa 3 m langes Tier schließen lässt.[102][34]

Alle bisher aus dem Ouled-Abdoun-Becken vorgestellten Vögel datieren in das Paläogen und stehen innerhalb der Neukiefervögel, es handelt sich um die derzeit ältesten Belege aus Afrika. Hierzu zählt unter anderem Dasornis aus der ausgestorbenen Gruppe der Pelagornithidae. Diese großen Seevögel mit gezackten Schnabelrändern, was an Zähne erinnert und ihnen auch die Bezeichnung als „Pseudozahnvögel“ einbrachte, wurden ursprünglich als den Pelikanen nahestehend angesehen, gehören aber mittlerweile einer eigenständigen Gruppe an. Dasornis ist im Ouled-Abdoun-Becken mit wenigstens drei Arten überliefert, deren Flügelspannweite zwischen 1,5 und 4,5 m variierte. Das aufgefundene Material ist recht umfangreich, aber insgesamt sehr stark fragmentiert, was der Natur des fragilen Skelettbaus entspricht. Im Gegensatz zu anderen Angehörigen der Gruppe war Dasornis eventuell kein so stark spezialisierter Gleitflieger und aufgrund der längeren Hinterbeine auch an Land agiler.[103] Zwei weitere Vogelvertreter reihen sich in das engere Umfeld der Tropikvögel ein. Bei der einen handelt es sich um Lithoptila, eine Form, die ursprünglich basierend auf einem Hirnschädel aufgestellt wurde. Nachfolgend kamen auch zahlreiche Gliedmaßenknochen zum Vorschein, die das Tier als einen geschickten Flieger mit schlanken Flügeln auszeichnen, deren Spitzen gespreizt etwa 1 m auseinanderstanden.[104][105] Die zweite Form wird durch Phaetusavis repräsentiert, von der aber lediglich ein Oberarmknochen zu Tage gefördert wurde. In den Körperausmaßen entsprach sie weitgehend Lithoptila, allerdings war bei Phaetusavis das obere Gelenkende massiver gestaltet.[106][34]
Säugetiere



Die paläogenen Ablagerungen enthalten auch eine nennenswerte Anzahl an Resten von Säugetieren, bei denen es sich ausnahmslos um landlebende Formen handelt. Es kommen Vertreter von zwei der vier großen Linien der Höheren Säugetiere mit weniger als ein Dutzend Gattungen vor. Das varianten- und umfangreichste Material gehört zu den Afrotheria. Hier repräsentieren Ocepeia und Abdounodus zwei Vertreter aus dem älteren Paläogen, die in einem näheren Verwandtschaftsverhältnis zu den Paenungulata stehen ohne diesen direkt eingegliedert zu sein. Ersterer kann umfangreiches Schädelmaterial vorweisen. Es zeigt ein kleines Tier mit 8 bis 20 kg Körpergewicht und kurzer Schnauze an, das sich von weicher Pflanzenkost ernährte. Letzterer ist von einigen Ober- und Unterkiefern belegt und kann gemäß den Zahnmerkmalen als etwas moderner eingestuft werden.[107][108][109][110] Die Schliefer, eine in ihrer Vergangenheit extrem variable, heute aber auf einige wenige murmeltierartige Formen zusammengeschrumpfte Gruppe der Paenungulata, sind mit einem einzigen Mahlzahn dokumentiert, der eventuell der urtümlichen Gattung Seggeurius entspricht. Mit einer Altersdatierung in den Beginn des mittleren Abschnitts der Paläogens stellt er den Erstnachweis der Schliefer dar.[7] Gleiches ist für die Rüsseltiere zutreffend. Sie kommen mit mehreren Formen vor. Als die ursprünglichste wird Eritherium aus dem älteren Paläogen angesehen, von dem eine Teilschädel und einzelne Unterkieferfragmente zu Tage kamen. Abweichend von allen späteren Rüsseltieren wies der Unterkiefer noch die vollständige Bezahnung der Höheren Säugetiere auf.[111][112] Das etwas jüngere und durch die beginnende Reduktion der vorderen Zähne auch modernere Phosphatherium war die erste im Ouled-Abdoun-Becken nachgewiesene Säugetierform.[113][114][115] Seit dem Erstfund eines Schädels sind mehrere weitere Exemplare hinzugekommen.[116] Der jüngste Vertreter der Rüsseltiere im Ouled-Abdoun-Becken wird Daouitherium zugewiesen und lässt sich in das mittlere Paläogen einstufen. Es liegen verschiedene Unterkieferreste vor.[117] Alle Rüsseltiere des Ouled-Abdoun-Beckens waren im Vergleich zu den heutigen Elefanten sehr klein. Für Eritherium ist ein Körpergewicht von 3 bis 8 kg rekonstruierbar, Phosphatherium war dem gegenüber schon fast doüppelt so schwer. Die starke Körpergrößenzunahme der Rüsseltiere zeichnet sich schon bei Daouitherium ab, das 80 bis 170 kg auf die Waage brachte. Sie besitzen weitgehend Zähne mit niedrigen Zahnkronen und ernährten sich daher wohl von weicher Pflanzenkost. Bei keiner Form waren, soweit es das Fundmaterial aussagen lässt, Rüssel oder Stoßzähne ausgebildet. Mit den Rüsseltieren nahe verwandt sind die ausgestorbenen Embrithopoda, deren Charakterform Arsinoitherium aus dem Unteren Oligozän des Fayyum in Ägypten riesige Körpermaße ausbildete und durch knöcherne Hörner auf dem Schädel charakterisiert ist. Im Ouled-Abdoun-Becken kommt mit Stylolophus, belegt über mehrere zerquetschte Schädel, ein früher Vorfahre vor, der zwischen 30 und 80 kg wog und somit deutlich kleiner war. Auch bezüglich der Zähne zeigt er sich ursprünglicher, da diese noch nicht so extrem hochkronig sind.[35]
Neben den Afrotherien sind die Laurasiatheria die zweite registrierte Säugetierlinie des Ouled-Abdoun-Beckens. Es wurden bisher ausschließlich Angehörige der Hyaenodonta gefunden, eine Gruppe ausgestorbener fleischfressender Formen, die den Raubtieren nahe stehen, sich von diesen aber durch eine im Gebiss weiter nach hinten verlagerte Brechschere unterscheiden. Aus dem älteren Paläogen stammt Lahimia, jünger ist dagegen Boualitomus, von beiden wurden mehrere Unterkiefer aufgedeckt. Im Gegensatz zu anderen sehr frühen Hyaenodonten ist das Gebiss bei Lahimia und Boualitomus schon deutlicher spezialisiert und zeigt neben der Reduktion der vorderen Backenzähne eine Tendenz zu einer überwiegend schneidenden Funktion. Wahrscheinlich gehören beide Gattungen einer ursprünglich afrikanischen Radiationsgruppe der Hyaenodonten an.[118][119] Darüber hinaus kann hier noch ein Unterkieferfragment mit einem einzigen erhaltenen Zahn mit zwei quergestellten Leisten genannt werden. Seine genaue Zuweisung ist unklar, eventuell ist es der Überrest eines ursprünglichen Huftiers.[7]
Altersstellung

Die Ablagerungen der Phosphatserie erlauben bisher keine absoluten Altersdatierungen. Die zeitliche Zuordnung der einzelnen Schichten und ihrer Funde basiert weitgehend auf der Biostratigraphie, wofür hauptsächlich die artenreiche Hai- und Rochenfauna herangezogen wird. Die Grundlagen hierfür wurden bereits in der ersten Hälfte des 20. Jahrhunderts herausgearbeitet und später verfeinert, etwa durch die Berücksichtigung der Microfauna. Allgemein datieren die Plattenkiemer die Phosphatserie in den Zeitraum von Maastrichtium als letzten Abschnitt der Oberkreide bis zum Ausgang des Ypresiums, also dem Übergang vom Unteren zum Mittleren Eozän innerhalb der Paläogens. Die absoluten Altersdaten liegen damit zwischen 72 und knapp 48 Millionen Jahren, was etwa 24 Millionen Jahre umspannt. Innerhalb dieses Zeitraums sind zwei bedeutende geologische Ereignisse eingebettet: einerseits die Kreide-Paläogen-Grenze vor rund 66 Millionen Jahren verbunden mit dem Verschwinden der Dinosaurier und zahlreicher verwandter Gruppen und dem Aufstieg der Säugetiere, andererseits das Paläozän/Eozän-Temperaturmaximum vor gut 56 Millionen Jahren, welches zu einem kurzfristigen starken globalen Temperaturanstieg führte. Einige Haiformen wie Squalicorax, Plicatoscyllium oder Serratolamna erreichen dabei eine sehr große stratigraphische Reichweite, andere wie Ginglymostoma, Anomotodon beziehungsweise Ganntouria treten nur kurzfristig in Erscheinung. Gleiches gilt für die Rochen wie Rhombodus oder Ganopristis im Vergleich zu Youssoubatis oder Duwibatis. Sie ermöglichen mit ihren sich teils abwechselnden einzelnen Artvertretern eine recht präzise relative Alterseinstufung der Schichten, die für nahezu alle Bereiche der Phosphatablagerungen gelten. Nachteilig für die Biostratigraphie wirkt sich die starke Komprimierung und Kondensierung der Schichtenfolge im Ouled-Abdoun-Becken aus. Dadurch lässt sich hier beispielsweise das Maastrichtium weniger gut auflösen und feiner untergliedern als es das Ganntour-Becken mit seiner deutlich mächtigeren und vollständigeren Ablagerungsserie bietet.[51][43][120] Problematisch ist auch die Paläozän-Folge, da hier unter anderem das Seelandium als dessen mittlerer Abschnitt nicht vom Danium und Thanetium abtrennbar ist und innerhalb dieser beiden Stufen verborgen bleibt.[33]
Neben dieser stratigraphischen Auflösung mittels der marinen Faunengemeinschaft werden seit den 2010er Jahren auch geochemische Analyseverfahren eingesetzt. Hervorzuheben sind hier Isotopenuntersuchungen basierend auf Sauerstoff und Kohlenstoff, mit denen vorerst aber nur die Abfolgen des Paläogens feiner differenziert werden konnten. Grundlage bildeten Fossilreste, vor allem aus den bone beds. Die Untersuchungen konnten die biostratigraphischen Ansätze weitgehend bestätigen und sind besonders in Bezug auf die Kohlenstoffanalysen aufschlussreich. Demnach umfasst das untere bone bed (F1) innerhalb des Couche IIa, von wo die Reste von Eritherium und Ocepeia stammen, aufgrund einiger positiver δ13C-Schwankungen das beginnende Thanetium und schließt das Seelandium in Teilen mit ein. Hier befindet sich demnach die Grenze zwischen Seelandium und Thanetium, die auf etwa 59 Millionen Jahren datiert ist. Das obere bone bed (F2) aus dem couche IIa gehört in den weiteren Verlauf des Thanetiums. Dagegen markiert das dritte bone bed (F3) an der Basis des intercalaire II/I mit den Fossilfunden von Phosphatherium den Beginn des Ypresiums vor rund 56 Millionen Jahren. Allerdings lässt sich das hier ebenfalls einsetzende Paläozän/Eozän-Temperaturmaximum, welches sich weltweit durch ein sehr niedriges δ13C-Verhältnis abzeichnet, nur schwer nachweisen. Die höheren Lagen gehören dann ebenfalls zum Ypresium. Hier zeigen sich im Abschnitt von couche 0 bis zum sillon A sehr niedrige Kohlenstoff-Isotopenwerte, was höchstwahrscheinlich mit zwei weiteren Wärmeschwankungen korreliert, die als ETM-2 und ETM-3 bezeichnet werden und den Zeitraum vor 54 und 53 Millionen Jahren abdecken. Aufgrund von Auflösungsschwierigkeiten bei den Isotopenanalysen ist unklar, ob die obersten Schichten der Phosphatserie auch das ausgehende Ypresium einschließen, jedoch können ein Teil der höheren sillon-Lagen mit dem Früheozänen Klimaoptimum vor rund 52 bis 50 Millionen Jahren gleichgesetzt werden, das sich wiederum durch positive δ13C-Werte absetzt. Das Lutetium scheint aber nicht mit eingeschlossen zu sein. Im Vergleich mit der Magnetostratigraphie, welche den Wechsel der Polarität des Erdmagnetfeldes dokumentiert, reicht die gesamte analysierte Sektion von Chron 26 bis höchstwahrscheinlich Chron 22, welche zwischen 61 und 49 Millionen Jahren datieren.[33][32]
Landschaftsrekonstruktion
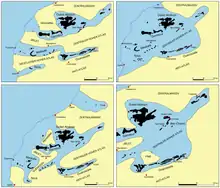


Die Phosphatlager Marokkos sowie die allgemein marinen Ablagerungen zeugen von einem ehemaligen Meer, das von der Oberkreide bis zum Eozän bestand. Darauf deuten auch die zahlreichen Fossilreste wie die Foraminiferen und die Hai- und Rochenartigen. Letzterer verweisen zudem auf variierende Meerestiefen, teilweise von Schelfgebieten mit 100 bis 150 m Tiefe bis hin zu Tiefseeregionen. Es existieren verschiedene Rekonstruktionen über die damalige Landschaft der heutigen Phosphatlager. In einer klassischen Ansicht, maßgeblich entwickelt von Georges Choubert und Henri Gauthier in den 1950er Jahren, erstreckten sich von der westlichen Atlantikküste drei enge Golfe landeinwärts. Der „nördliche Golf“ begann mit einer breiten Mündung zwischen Safi sowie El Jadida und verlief zwischen den Festlandsblöcken des Jbilets im Norden und der Meseta im Süden ostwärts über das Ganntour-Becken bis zum Ouled-Abdoun-Becken und umgab das Rhamna-Massiv, das dadurch als Insel hervortrat. Nach Süden schloss sich der „Golf von Essaouira“ an, der bei der gleichnamigen Stadt mündete und das Meskala-Becken und das Ouarzazate-Becken einnahm. Eingerahmt wurde er durch den Jbilet im Norden und den westlichen Hohen Atlas im Süden. Den südlichen Abschluss bildete der „Golf von Sous“ bei Agadir mit dem Sous-Becken als Zentrum. Bei diesem Modell blieben vor allem die Mündungsbereiche unklar, da spätere Hebungen im Küstengebiet die dortigen Ablagerungen überprägten. Es wurde aber mehrfach abgewandelt, so unter anderem von Armand Boujo in den 1970er Jahren. Er betrachtete unter anderem das Rhamna-Massiv als halbinselartig, wodurch der „nördliche Golf“ bei Youssoufia sehr schmal auf den Atlantik traf. Alternativ dazu sahen einzelne Autoren eine zweite Atlantikverbindung des „nördlichen Golfes“ über Settat.[12][13][121]
Ein weiteres Modell wurde in den 1980er Jahren von H. Salvan erarbeitet. In seiner Rekonstruktion vereinigten sich die beiden nördlichen engen Golfe zu einem breiten Epikontinentalmeer, das „Phosphatmeer“, dessen Atlantikzugang sich zwischen Casablanca und Essaouira erstreckte. Das Rhemna-Massiv bestand als Insel analog zur Ansicht von Boujo, während vom Jbilet aus eine Landverbindung nach Süden ausgebildet war. Die Existenz eines südlichen Ǵolfes bei Sous lehnte Salvan weitgehend ab, vielmehr verband sich das Sous-Becken nach Norden mit dem Meskala-Becken. Etwa im gleichen Zeitraum entwarf Hans-Georg Herbig ein ähnliches Bild, allerdings nahm er eine weitflächige Überflutung des gesamten Bereiches an. Nach Osten hin überdeckte das Meer zusätzlich das Ouarzazate-Becken und hatte damit auch das Gebiet des heutigen Hohen Atlas eingenommen. Die Existenz einzelner Inseln schloss Herbig nicht aus. Als Variation davon kann die Rekonstruktion von Jörg Trappe aus dem Beginn der 1990er Jahre angesehen werden. Hier dehnte sich die Meeresverbindung des Epikontinentalmeeres von Casablanca bis weit nach Süden nach Agadir aus.[122][12][13]
Die Gestaltung des damaligen Meeres ist noch nicht eindeutig geklärt. So sehen einzelne Wissenschaftler aufgrund der Zusammensetzung der Fischfauna eine vollständige Meeresbedeckung eher kritisch,[7] andere befürworten wiederum ein einheitliches Meer und beziehen sich dabei auf die Charophyten und Einsiedlerkrebse. Lediglich lokal wird dann der einstige Küstenverlauf modifiziert, etwa am Rand des Hohen Atlas.[123] Auch besteht mitunter aus sedimentologischer Sicht die Annahme eines sich teilweise verengenden Meereszuganges von der Oberkreide hin zum Lutetium. Dem widersprechen aber Analysen an den Metallen der Seltenen Erden aus den Fossilien quer durch alle Schichten. Hierbei ließ sich keine Verknappung dieser Elemente ermitteln, so dass von einer kontinuierlichen und gleichbleibenden Zufuhr von frischem Meereswasser auszugehen ist. Außerdem scheint das damalige „Phosphatmeer“ von dem zunehmenden Wasseraustausch zwischen dem nördlichen und südlichen Atlantik der damaligen Zeit im Zuge der weiteren Ozeanöffnung profitiert zu haben.[124] Generell werden sowohl das „Phosphatmeer“ als auch die engen Golfgebiete als relativ warm angesehen, deren Temperaturen in der Oberkreide bei rund 27 °C gelegen haben dürften, was Analysen an Sauerstoffisotopen aus Fischfossilien ergaben. Im Paläozän kam es zu einer rapiden Abkühlung des „Phosphatmeeres“ auf rund 19 °C, dem sich dann wieder eine Erwärmung im Eozän anschloss. Die Ursachen sind nicht eindeutig und können sowohl mit einer zunehmenden Meerestiefe als auch mit Änderungen in der Erdatmosphäre zusammenhängen, wobei der auffällige Abfall der Temperatur an der Kreide-Paläogen-Grenze letzteres befürwortet.[125][126]
Die reichhaltige marine Fauna der Oberkreide ist geprägt durch eine Dominanz an Spitzenprädatoren, die große Haie, Mosasaurier und Plesiosaurier einschlossen.[75] In der Regel nutzen solche einflussreichen Räuber in modernen Ökosystemen unterschiedliche Nahrungsquellen. Dies wird auch für die diversen Mosasaurier angenommen und mit der variierenden Zahngestaltung erklärt, die für eine vielfältige Nischennutzung spricht. Demnach kamen sowohl fischfressende Tiere mit scharfen und spitzen Zähnen als auch auf hartschalige Muscheln spezialisierte Formen mit rundlich-stumpfen Zähnen vor.[71] Untersuchungen an Kalziumisotopen erbrachten, dass sich der größte Teil der Räuber des „Phosphatmeeres“ allerdings von Fischen ernährte, wodurch sich das Beutespektrum stark einengte und ein „Wespentaillen“-Ökosystem bestand, in dem die Existenz der Beutegreifer von wenigen Beutetiere abhängt. Unter heutigen Bedingungen sind derartige Gemeinschaften sehr instabil, da ein Verschwinden der Beute auch eine Dezimierung der Räuber nach sich zieht, wofür nur einzelne Änderungen im ökologischen Umfeld ausschlaggebend sein können. Möglicherweise führten die drastischen Umwälzungen während des Kreide-Paläogen-Übergangs damit auch zum lokalen Verschwinden der großen räuberischen Reptilien, aber auch der Fische. Im frühen Paläogen übernahmen teilweise die dyrosauriden Krokodilsverwandten und weiterhin Haie die Rolle der großen marinen Räuber. Sie verschwanden dann aber auch weitgehend im weiteren Verlauf. Hier zeigt der Beginn des Ypresiums ein stärkeres Verschwinden der Haie an.[127][121]
Das Vorkommen von landbewohnenden Dinosauriern und Säugetieren im Ouled-Abdoun-Becken spricht für ein nur wenig entferntes Küstengebiet. Ihre häufig stärkere Zersetzung verweist auf länger zurückgelegte Transportwege in die Meeresumgebung. Dieses Hinterland war vermutlich mit einem tropischen Regenwald oder einem Mangrovenwald bewachsen, der von langsam fließenden Gewässern gequert wurde. Als Anzeiger dafür können neben den Pterocarpus-Funden aus dem Ouled-Abdoun-Becken direkt[37] unter anderem auch Früchte und Stammreste der Nipapalme herangezogen werden, die vom Südrand des Hohen Atlas im Ouarzazate-Becken stammen und in den Zeitraum des Übergangs vom Paläozän zum Eozän datieren.[128][129] Mit einer Stellung im ausgehenden Unteren Eozän etwas jünger sind Pollenansammlungen aus der gleichen Region, die auf Pflanzen mit Anpassungen an Brackwasser zurückgehen. Auch vereinzelte Makroreste von Blättern deuten darauf hin, so beispielsweise von Wassersalat.[128] Allgemein ähnelt die Pollenflora aus dem südlichen Atlasgebiet jener aus dem Ganntour-Becken der gleichen Zeit, doch fehlen hier eindeutige Mangroven- und Salzmarschanzeiger.[130] Auf ein tropisches bis subtropisches Klima verweisen des Weiteren verschiedene Wirbeltierreste wie Krokodile und Schildkröten. Mitunter ausgebildete Lagen aus Smektiten und Gips im Ouarzazate-Becken legen dabei ein möglicherweise stärker saisonalisiertes Klima mit sich abwechselnden Trocken- und Regenzeiten nahe. Isotopenmessungen an einzelnen Knochen von Säugetieren bestätigten diese Annahme und geben jährliche Temperaturwerte von über 20 °C für den Zeitraum des Paläogens an. Der ermittelte Jahresniederschlag von unter 500 mm verweist auf eher trockene Bingungen.[131] Geographisch befand sich die Landschaft damals im Bereich des 20. bis 23. nördlichen Breitengrades.[128][129]
Vergleich mit regional und überregional bedeutenden Fundstellen
Die formal nächsten Vergleiche bilden ähnliche phosphathaltige Beckenstrukturen in Nordafrika wie das benachbarte Ganntour-Becken oder aber auch das Meskala- und das Sous-Becken. Alle diese Becken weisen eine sehr umfassende sedimentäre Abfolge des Maastrichtiums auf, die sich teils höher auflösend gliedern lässt als im Vergleich zum Ouled-Abdoun-Becken. Dagegen sind die Ablagerungen des Paläogens bis auf das Ganntour-Becken eher spärlich ausgebildet. Übereinstimmend mit dem Ouled-Abdoun-Becken kommt eine reichhaltige Knorpel- und Knochenfischfauna vor, die als Grundlage der biostratigraphischen Gliederung dient.[43] Aus dem Meskala- und Sous-Becken sind Funde von Reptilien bisher nur selten berichtet worden und beschränken sich weitgehend auf Krokodile und Schildkröten.[132][1] Bedeutende Vorkommen liegen hingegen aus dem Ganntour-Becken mit rund 60 Arten an Hai- und Rochenartigen, über einem halben Dutzend Arten an Knochenfischen und mehr als ein Dutzend Arten an Reptilien. Die Fauna spiegelt jene des Ouled-Abdoun-Beckens wieder, gelegentlich treten aber lokale Eigenheiten auf.[11][46] Unter den Reptilien dominieren vor allem die Mosasaurier gegenüber den Plesiosauriern. Des Weiteren sind neben Krokodilen, hier hauptsächlich die Dyrosauridae auch Schildkröten und Warane belegt. Mit Ausnahme des Ouled-Abdoun-Beckens brachte keines der anderen Phosphatbeckens Marokkos bisher Reste von Vögeln und Säugetieren hervor.[2][120][71]
Einen teils deutlich älteren Ursprung als die marokkanischen Phosphatablagerungen haben jene Ägyptens, die sich in der Oase von Dachla im westlichen Landesteil befinden. Hier sind vor allem die Sedimentfolgen der Quseir-, Duwi- und der Dakhla-Formation aufgeschlossen. Ihr Entstehungszeitraum reicht vom Cenomanium über das Maastrichtium bis zum Paläozän. Die phosphathaltigen Bildungen beschränken sich auf die Duwi-Formation und sind hier in einer Sequenz aus Ton-, Sand- und Schluffsteinen sowie Konglomeraten eingeschlossen. Die liegende Quseir-Formation ist weitgehend aus geschichteten Schiefern aufgebaut, die hangende Dakhla-Formation wiederum charakterisiert sich durch Schiefer, Sand- und Schluffsteine sowie Mergel. Fossilfunde fanden sich in allen drei Gesteinseinheiten, wobei einschränkend gesagt werden muss, dass paläontologische Arbeiten hier bisher nur sporadisch stattfanden. Aus der Quseir-Formation wurden verschiedene Haie und Lungenfische, dyrosauride und eventuell gavialoide Krokodile, Schildkröten und Dinosaurier, möglicherweise Spinosaurus, berichtet, ohne das für den Großteil der Formen bisher eine genauere Beschreibung vorliegt. Für die Duwi-Formation sind Fische sowie Plesiosaurier und Mosasaurier, hier Globidens, dokumentiert. Die Dakhla-Formation lieferte Wirbeltierreste und Pflanzenmaterial zuzüglich eines bisher nicht genauer bestimmten Gliedmaßenknochens eines mittelgroßen Sauropoden.[133][134][135][136] Das Phosphatbecken von Gafsa in Tunesien umfasst ebenfalls einen langen Bildungszeitraum, die hauptsächlichen Ablagerungen datieren jedoch in das Untere und Mittlere Eozän und beinhalten neben zahlreichen Hai- und Rochenzähnen gelegentlich Reste von Schildkröten und Krokodilen. Unter anderem wurde von hier im Jahr 1893 der erste bekannte Schädel eines Dyrosauriden vorgestellt.[137][138]
Außerhalb dieses speziellen Bildungsmilieus sind Fundstellen der Oberkreide in Nordafrika recht gut belegt, genannt seien hier nur die Bahariya-Formation in Ägypten oder die Kem Kem Beds in Marokko,[139] die aber beide mit einer Datierung in das Cenomanium in einen deutlich früheren Abschnitt der geologischen Serie gehören. Zeitlich näher stehende Lokalitäten sind dagegen weitaus seltener. Bedeutend ist die Wadi-Milk-Formation im Sudan, die ursprünglich als ähnlich alt wie die Bahariya-Formation angesehen wurde, neueren absoluten Datierungen zufolge aber mit etwa 79,2 Millionen Jahren im Campanium entstand.[140] Ein großer Unterschied zu den Phosphatbecken ist das Überwiegen terrestrischer Faunenelemente, auch wenn die Bildung der Gesteinseinheit in relativer Küstennähe erfolgte. Angezeigt wird dies durch verschiedene Schleichenlurche, Schildkröten und den Dinosauriern, die sowohl große Sauropoden wie die Titanosauridae oder die Carcharodontosauridae als auch Theropoden wie die Dromaeosauridae oder Ornithopoden wie die Iguanodontidae oder die Hypsilophodontidae einschließen.[141][142][143][144]
Als zeitliches Äquivalent zu den paläogenen Ablagerungen des Ouled-Abdoun-Beckens ist das Ouarzazate-Becken am Südrand des Hohen Atlas in Marokko aufzufassen. Die Ablagerungen hier enthalten teilweise auch Phosphate und sind mit einer sehr reichhaltigen Fossilfauna ausgestattet, die sich über mehrere Fundstellen verteilt. Diese decken mit Adrar Mgorn, Ilimzi oder Talazit aus dem ausgehenden Paläozän, N’Tagourt aus dem Unteren und Aznag aus dem Mittleren Eozän einen Zeitraum von mehr als 10 Millionen Jahren ab. Die Datierung der einzelnen Fundstellen erfolgt dabei sowohl magneto- als auch biostratigraphisch, letzteres wie in den Phosphatbecken nördlich des Hohen Atlas über die umfassende Hai- und Rochengemeinschaft.[145] Auffallend am Fundmaterial des Ouarzazate-Beckens ist der hohe Fragmentierungsgrad der Funde sowie der große Anteil an landbewohnenden Formen, was nicht nur die Säugetier-, sondern auch die Herpetofauna einschließt, letztere beispielsweise vertreten mit Fröschen, Geckos, Skinkartigen und Doppelschleichen.[146] Unter den Säugetieren sind zahlreiche Kleinsäuger präsent, gezeigt unter anderem an insektenfresserartigen Formen wie Afrodon, Todralestes oder Garatherium. Hierbei handelt es sich wohl um eine endemische Faunengemeinschaft, die mit zeitgleich im nördlichen Tethys-Randgebiet, dem heutigen Eurasien, auftretenden Gattungen weniger verwandt sein dürfte als zu den späteren Afroinsectiphilia. Daneben kommen bereits in den ältesten Fundschichten frühe Primaten vor, so Altiatlasius, anfänglich als ein den Koboldmakis nahestehendes Tier interpretiert, das aber nach heutiger Auffassung eventuell als Basalform die Entwicklung der Affen einläutet. Analog zum Ouled-Abdoun-Becken repräsentiert hier Tinerhodon die fleischfressenden Hyaenodonta.[147][148][149][150] In den jüngeren Fundschichten lassen sich dann auch erstmals Funde von Fledertieren und Soricomorpha, der Verwandtschaftsgruppe der Spitzmäuse, Maulwürfe und ähnlicher Formen, sowie möglicherweise auch der Rüsselspringer belegen.[151]
In das Untere und Mittlere Eozän fallen dann Fundstellen wie El-Kohol und Gour Lazib im nördlichen beziehungsweise im westlichen Algerien und Chambi in Tunesien. Bezüglich der Entwicklung der Säugetierfauna können noch einzelne Gemeinsamkeiten mit dem Ouled-Abdoun-Becken und Ouarzazate festgestellt werden, etwa in El-Kohol im Auftreten früher insektenfresserartiger Formen wie Garatherium oder dem ursprünglichen Schliefer Seggeurius. Allerdings sind hier auch schon teils stammesgeschichtlich deutlich entwickeltere Formen belegt. Erkenntlich wird dies unter anderem am Rüsseltier Numidotherium, das nicht nur gegenüber den Frühformen des Ouled-Abdoun-Beckens eine markante Größenzunahme durchlief, sondern zusätzlich ein weitaus stärker reduziertes Gebiss besitzt und schon Anzeichen einer Rüsselbildung aufweist.[152] Die zunehmende Herausbildung stammesgeschichtlich entwickelterer Formen wird in Gour Lazib ersichtlich, wo wiederum Reste von Titanohyrax, Megalohyrax und Microhyrax vorliegen und damit Schliefer, die nicht mehr der alten Seggeurius-Gruppe angehören, sondern modernere und ökologisch deutlich diverse Formen repräsentieren. Zudem sind hier mit Azibius und Algeripithecus frühe Primaten nachgewiesen ebenso wie auch Nagetiere dokumentiert wurden.[153][154] Chambi wartet mit frühen Rüsselspringern wie Chambius sowie ebenfalls Schliefern und Primaten auf.[155][156] Die Fundstellen leiten zur überaus fossilreichen Lagerstätte des Fayyum im nördlichen Ägypten über. Das Fayyum zählt zu den bedeutendsten Fossilregionen des Paläogens Afrikas und datiert in das Obere Eozän und Untere Oligozän. Die extrem reichhaltige Fauna wird geprägt durch Fische, Reptilien, Vögel und Säugetiere. Unter letzteren befinden sich Rüsseltiere, die an der Basis der Entwicklung elefantenartiger Formen stehen, verschiedenste Affen in der Vorfahrenlinie zu den Altweltaffen und Angehörige einer frühen Aufsplitterung der Wale sowie der Seekühe. Des Weiteren kommen Tenrekartige, Schliefer, Paarhufer, Fledertiere und Nagetiere wie auch einige heute erloschene Linien vor.[157] Ähnlich zum Fayyum erbrachte der obereozäne Abschnitt der Samlat-Formation bei Ad-Dakhla in der Westsahara neben Fischen, Schildkröten, Krokodilen und einzelnen Vögeln auch Meeres- und Landsäugetiere.[158][159][160]
Forschungsgeschichte
Anfänge
Die Entdeckung phosphathaltiger Ablagerungen im nördlichen Afrika reicht bis in das ausgehende 19. Jahrhundert zurück. Im Jahr 1885 berichtete Philippe Thomas über eozäne Vorkommen bei Gafsa in Tunesien. Nur wenige Zeit später wurden die ersten Fossilien beschrieben, während der kommerzielle Abbau um 1899 einsetzte.[161][137] Im Vergleich dazu erreichten Hinweise über die marokkanischen Phosphatlager deutlich später die Öffentlichkeit. Die erste Erwähnung von Sedimentgesteinen des Eozäns in Marokko zwischen Essaouira und Marrakesch wurde von Abel Brives im Jahr 1905 formuliert. Entdeckt hatte er sie im Jahr zuvor auf seiner Reise durch Nordafrika.[162] Nur drei Jahre später verkündete er nach einer erneuten Reise in das Gebiet das Vorkommen von Phosphaten in diesen Schichten, die auch Reste von Muschelschalen enthielten.[163] Die Aufschlüsse stießen anfangs auf nur wenig Interesse, was sich erst 1917 änderte, als Brives über bedeutende Phosphatvorkommen zwischen den Ortschaften Oued Zem und El Borouj berichtete. In einer zwei Jahre später erschienenen Veröffentlichung beschrieb Brives erstmals die geologische Situation genauer und unterteilte die Abfolge in drei Abschnitte. Die unterste aus Tonen und Gipsen war von dunkler Farbgebung, gefolgt von einer Wechsellage aus Mergel und Kalksteinen mit eingebetteten Silexlagen sowie einer nicht-silikatischen, phosphathaltigen Sandsteinbank, abgeschlossen durch eine feste Kalksteinlage. Hierbei legte er auch eine erste Fossilliste vor, die vor allem verschiedene Haie berücksichtigte. Seinen Beobachtungen zufolge barg die untere Lage nur schlecht erhaltene Fossilien, während sich die mittlere als sehr reich an Funden kennzeichnete und die obere durch zahlreiche Weichtierreste charakterisiert war. Nach Brives gehörten alle drei Abschnitte dem Eozän an.[164] Die durch die Phosphatlagen sprunghaft angestiegene strategische und wirtschaftliche Bedeutung des Ouled-Abdoun-Beckens führte im Jahr 1920 zur Gründung des Office Chérifien des Phosphates (OCP), ein großes Unternehmen, das bereits im Jahr darauf die Förderung der Phosphate in Marokko, zerst im Ouled-Abdoun-Becken, später auch in den benachbarten Gebieten wie dem Ganntour-Becken koordinierte.[2][34]
Im Jahr 1922 stellte Léonce Joleaud eine Reihe von Wirbeltierfossilien vor, darunter unter anderem auch das Krokodil Dyrosaurus und verschiedene fossile Haiformen. Seiner Einschätzung nach gehörten die Ablagerungen des Ouled-Abdoun-Beckens einem zeitlich weiter umfassenden geologischen Rahmen an, der von der ausgehenden Kreide bis zum Eozän reichte. Damit stand er im Widerspruch zu Brives, schloss sich aber der Meinung von Louis Gentil an. Dieser hatte im gleichen Jahr eine vergleichbare Alterseinschätzung abgegeben und sich dabei auf biostratigraphisch aussagekräftige Fossilien und deren Verbreitung in den Ablagerungen berufen.[165][166] Weitere frühe Funde gehen auf Charles Depéret and P. Russo aus der Mitte der 1920er Jahre zurück. Sie vermeldeten Reste von Haien, Krokodilen und Mosasauriern aus der Region um Melgou im nordöstlichen Teil des Ouled-Abdoun-Beckens.[2][34]
Die Zeit Arambourgs

Auf Anfrage des damaligen Generaldirektors des OCP zur Klärung der bestehenden stratigraphischen Probleme im Ouled-Abdoun-Becken begann 1934 Camille Arambourg mit seinen Forschungsarbeiten. Sie sollten für die nächsten zwei Dekaden bestimmend sein. Arambourg, Mitarbeiter des Muséum national d’histoire naturelle in Paris, wählte dafür die am häufigsten in den Phosphatlagen auftretenden Wirbeltiere, namentlich die Haie, Rochen und Reptilien. Er führte eine Untersuchung der einzelnen geologischen Schichten durch und sortierte beziehungsweise siebte das entsprechende Fundmaterial. Nach einer Untersuchungsperiode von knapp zehn Jahren hatte Arambourg rund 100.000 Zahn- und Knochenreste gesichtet. Eine erste grobe Schichtgliederung stellte er bereits 1935 vor. Seine Hauptpublikation erschien im Jahr 1952 unter dem Titel Les Vertébrés fossiles des gisements de phosphates (Maroc-Algérie-Tunisie). In dieser unterteilte Arambourg die Schichtenfolge des Ouled-Abdoun-Beckens in drei hauptsächliche phosphathaltige Glieder, von ihm couches (französisch für „Schicht“ oder „Bett“) genannt, mit dem jüngsten (Couche I) aus dem Eozän und dem ältesten (Couche III) aus dem Maastrichtium. Zudem erkannte er eine zunehmende Schichtmächtigkeit von Nordost nach Südwest, was sich weiter in das südwestlich anschließende Ganntour-Becken fortsetzt. Ihm gelang dabei auch eine Korrelierung der Schichten des Ouled-Abdoun-Beckens mit der des Ganntour-Beckens. Die von Arambourg festgelegte Schichtenabfolge für das Ouled-Abdoun-Becken hat weitgehend bis heute Bestand. Lediglich in der Feinstratigraphie erfolgten Korrekturen, da unter anderem das älteste Paläozän von Arambourg nicht abgetrennt werden konnte. Die Beschäftigung mit dem Fossilmaterial führte des Weiteren zur Einführung zahlreicher neuer Taxa. Auffallend hierbei ist jedoch, dass Arambourg trotz der Reichhaltigkeit im Fossilbericht so gut wie keine Vogel- oder Schildkrötenreste berücksichtigte.[1] Seine Ersterwähnung von Schildkröten fällt in das Jahr 1951, die Funde gehören aber nicht dem Ouled-Abdoun-Becken, sondern dem weit südlich gelegenen Sous-Becken an und datieren in die Oberkreide.[132][2][34] Gemeinsam mit Arambourg war Henri Salvan in den Phosphatbecken tätig. Er stellt 1955 in einer umfangreichen Monographie betitelt mit Les invertébrés fossiles des phosphates marocains ausführlich die Weichtiere der Region vor.[41]
Moderne Forschungen
Nach Arambourgs Pionierarbeit kam die Erforschung des Ouled-Abdoun-Beckens in den nächsten Jahrzehnten weitgehend zum Stillstand. Lediglich die verschiedensten Hai- und Rochenreste fanden Beachtung, denen sich unter anderem Henri Cappetta und Abdelmajid Noubhani in den 1980er und 1990er Jahren widmeten. Punktuell wurden darüber hinaus einzelne Reptilien berücksichtigt, etwa durch Éric Buffetaut. Der Zustand änderte sich erst Mitte der 1990er Jahre wieder. Zu diesem Zeitpunkt erlangte das Ouled-Abdoun-Becken durch die relativ unerwartete Entdeckung von Phosphatherium, dem bis dato ältesten Vertreter der Rüsseltiere, neue Aufmerksamkeit.[113] Dies führte dazu, dass im Jahr 1997 ein offizieller Vertrag zwischen dem OCP, dem Pariser Muséum national d’histoire naturelle und dem marokkanischen Ministerium für Energie, Bergbau, Wasser und Umwelt unterzeichnet wurde, der die Untersuchung der Fossilfunde aus den Phosphatablagerungen befördern sollte. Involviert waren darin auch verschiedenste Universitäten wie jener von Marrakesch, El Jadida, Montpellier und Bilbao. Im Ergebnis dieser Zusammenarbeit konnte in der Folgezeit eine Vielzahl neuer Fossilien geborgen und untersucht werden. Diese schlossen neben den bekannten Formen auch Tiergruppen ein, die zur Zeit Arambourgs noch völlig unbekannt im Ouled-Abdoun-Becken waren, etwa Pterosaurier und landbewohnende Säugetiere. Bedeutende, an den Untersuchungen beteiligte Wissenschaftler sind unter anderem Nathalie Bardet (Schildkröten und Mosasaurier), Haiyan Tong (Schildkröten), Stéphane Jouve (Krokodile), Nicholas R. Longrich (Dinosaurier und Pterosaurier), Xabier Pereda Suberbiola (Dinosaurier und Pterosaurier), Peggy Vincent (Plesiosaurier), Alexandra Houssaye (Schuppenkriechtiere), Estelle Bourdon (Vögel) und Emmanuel Gheerbrant (frühe Säugetiere). Die meisten Funde werden beim OCP oder im Muséum national d’histoire naturelle aufbewahrt. Daneben kam es aber auch zu größeren privaten Aufsammlungen und Verkäufen von Fossilfunden, die dadurch in die verschiedensten Institutionen in Europa, Nordamerika oder Asien gelangten.[2][34]
Literatur
- Nathalie Bardet, Emmanuel Gheerbrant, Abdelmajid Noubhani, Henri Cappetta, Stéphane Jouve, Estelle Bourdon, Xabier Pereda Suberbiola, Nour-Eddine Jalil, Peggy Vincent, Alexandra Houssaye, Floréal Solé, Khadija El Houssaini, Sylvain Adnet, Jean-Claude Rage, France de Lapparent de Broin, Jean Sudre, Baâdi Bouya, Mbarek Amaghzaz und Saïd Meslouh: Les Vertébrés des Phosphates cretacés-paléogènes (70,6 – 46,6 Ma) du Maroc. In: Samir Zouhri (Hrsg.): La Paléontologie des Vertébrés du Maroc. Paris, Mémoires de la Société Géologique de France, nouvelle série, 180, 2017, S. 351–452
- E. Jourani, F. Khaldoune und N.-E. Aoudjehane (Hrsg.): Promoting Geological Heritage: Challenges and Issues. La Conférence Internationale RALI 2015, Marrakech 05–11 octobre 2015. Notes et Mémoires du Service Géologique du Maroc, Numéro Spécial dédié au Patrimoine Géologique du Maroc, 2018, S. 103–136 ()
Einzelnachweise
- Camille Arambourg: Les Vertébrés fossiles des gisements de phosphates (Maroc-Algérie-Tunisie). Paris, 1952, S. 1–396
- Nathalie Bardet, Xabier Pereda Suberbiola, Stéphane Jouve, Estelle Bourdon, Peggy Vincent, Alexandra Houssaye, Jean-Claude Rage, Nour-Eddine Jalil, Baâdi Bouya und Mbarek Amaghzaz: Reptilian assemblages from the latest Cretaceous – Palaeogene phosphates of Morocco: from Arambourg to present time. Historical Biology 22 (1), 2010, S. 186–199, doi:10.1080/08912961003754945
- Aaron R. H. Leblanc, Michael W. Caldwell und Nathalie Bardet: A new mosasaurine from the Maastrichtian (Upper Cretaceous) phosphates of Morocco and its implications for mosasaurine systematics. Journal of Vertebrate Paleontology 32 (1), 2012, S. 82–104
- Office Chérifien des Phosphates: The phosphates basins of Morocco. In: A. J. G. Northolt, R. P. Sheldon und D. F. Davidson (Hrsg.): Phosphate deposits of the world, Vol. 2 – Phosphate rock resources. Cambridge University Press, 1989, S. 301–311
- Saad Bakkali: A resistivity survey of phosphate deposits containing hardpan pockets in Oulad Abdoun, Morocco. Geofísica Internacional 45 (1), 2006, S. 73–82
- Hamid El Haddia, Abdelmajid Benbouziane, Mustapha Mouflih, Es-said Jourani und M’barek Amaghzaz: Siliceous forms of phosphate deposits of Cretaceous age in Oulad Abdoun basin (Central Morocco). Mineralogy, geochemistry and diagenetic phenomena. Procedia Engineering 83, 2014, S. 60–69
- Emmanuel Gheerbrant, Jean Sudre, Henri Cappetta, Cécile Mourer-Chauviré, Estelle Bourdon, Mohamed Iarochène, Mbarek Amaghzaz und Baâdi Bouya: Les localités à mammifères des carrières de Grand Daoui, bassin des Ouled Abdoun, Maroc, Yprésien: premier état des lieux. Bulletin de la Societe Geologique de France 174 (3), 2003, S. 279–293
- N. El Assel, A. Kchikach, C. Durlet, N. AlFedy, K. El Hariri, M. Charroud, M. Jaffal, E. Jourani und M. Amaghzaz: Mise en évidence d’un Sénonien gypseux sous la série phosphatée du bassin des Ouled Abdoun: Un nouveau point de départ pour l’origine des zones dérangées dans les mines à ciel ouvert de Khouribga, Maroc. Estudios Geológicos 69 (1), 2013, S. 47–70, doi:10.3989/egeol.40781.168
- H. El Haddi, A. Benbouziane und M. Mouflih: Geochemical Siliceous and Silicified Facies of Phosphate Series of Ouled Abdoun Basin (Morocco). Open Journal of Geology 4, 2014, S. 295–302, doi:10.4236/ojg.2014.47022
- J. Lucas und L. Prévôt-Lucas: Tethyan phosphates and bioproductites. In: A. E. M. Nairn,L.-E. Ricou, B. Vrielynck und J. Dercourt (Hrsg.): The Ocean Basins and Margins, the Tethys Ocean. Plenum Press, New York, 1995, S. 367–391
- N. Bardet, X. Pereda Suberbiola, M. Iarochène, M. Amalik und B. Bouya: Durophagous Mosasauridae (Squamata) from the Upper Cretaceous phosphates of Morocco, with description of a new species of Globidens. Netherlands Journal of Geosciences – Geologie en Mijnbouw 84 (3), 2005, S. 167–175
- Hans-Georg Herbig: Das Paläogen am Südrand des zentralen Hohen Atlas und im Mittleren Atlas Marokkos. Stratigraphie, Fazies, Paläogeographie und Paläotektonik. Berliner Geowissenschaftliche Abhandlungen Reihe A 135, 1991, S. 1–289
- Hans-Georg Herbig und Jörg Trappe: Stratigraphy of the Subatlas Group (Maestrichtian – Middle Eocene, Morocco). Newsletter on Stratigraphy 30 (3), 1994, S. 125–165
- Nadia El Kiram, Mohammed Jaffal, Azzouz Kchikach, Driss El Azzab, Mustapha El Ghorfi, Oussama Khadiri, Es-Said Jourani, Ahmed Manar und Mohamed Nahim: Phosphatic series under Plio-Quaternary cover of Tadla Plain, Morocco: Gravity and seismic data. Comptes Rendus – Geoscience 351, 2019, S. 420–429, doi:10.1016/j.crte.2019.05.002
- Radouan El Bamiki, Michel Séranne, El Hassane Chellaï, Gilles Merzeraud, Mohamed Marzoqi und Mihaela Carmen Melinte‐Dobrinescu: The Moroccan High Atlas phosphate-rich sediments: Unraveling the accumulation and differentiation processes. Sedimentary Geology, 2020, doi:10.1016/j.sedgeo.2020.105655
- Gerhard Einsele and Jost Wiedmann: Turonian black shales in the Moroccan Coastal basins: first upwelling in the Atlantic Ocean. In: U. von Rad, K. Hinz, M. Sarnthein und E. Seibold (Hrsg.): Geology of the Nordwest African Continental Margin. Springer, 1982, S. 396–414
- Krzysztof P. Krajewski, Philippe Van Cappellen, Jean Trichet, Oliver Kuhn, Jacques Lucas, Agustín Martín-Algarra, Liliane Prévôt, Vinod C. Tewari, Luis Gaspar, Robin I. Knight und Michel Lamboy: Biological processes and apatite formation in sedimentary environments. Eclogae geologicae Helveticae 87 (3), 1994, S. 701–745
- Heide N. Schulz und Horst D. Schulz: Large Sulfur Bacteria and the Formation of Phosphorite. Science 307, 2005, S. 416–418
- J. Cosmidis, K. Benzerara, E. Gheerbrant, I. Estève, B. Bouya und M. Amaghzaz: Nanometer‐scale characterization of exceptionally preserved bacterial fossils in Paleocene phosphorites from Ouled Abdoun (Morocco). Geobiology 11 (2), 2013, S. 139–153, doi:10.1111/gbi.12022
- Craig R. Glenn, Karl B. Föllmi, Stanley R. Riggs, Gleb N. Baturin, Kurt A. Grimm, Jörg Trappe, Abdulkader M. Abed, Carlos Galli-Olivier, Robert E. Garrison, Andrei V. Ilyin, Caroline Jehl, Vera Rohrlich, Rushdi M. Y. Sadaoah, Manfred Schidlowski, Richard E. Sheldon und Hendrik Siegmund: Phosphorus and phosphorites: Sedimentology and environments of formation. Eclogae geologicae Helveticae 87 (3), 1994, S. 747–788
- Gabriel M. Filippelli: Phosphate rock formation and marine phosphorus geochemistry: The deep time perspective. Chemosphere 84, 2011, S. 759–766, doi:10.1016/j.chemosphere.2011.02.019
- Oskar Bär: Die Phosphate Marokkos. Geographica Helvetica 23 (2), 1968, S. 89–94 ()
- Jacques Lucas, Liliane Prévôt und Mohamed El Mountassir: Les phosphorites rubéfiées de Sidi Daoui, transformation météorique locale du gisement de phosphate des Ouled Abdoun (Maroc). Sciences Géologiques. Bulletin 32 (1–2), 1979, S. 21–37
- Steven J. Van Kauwenbergh: World Phosphate Rock Reserves and Resources. International Fertilizer Development Center, Muscle Shoals, Alabama, 2010, S. 1–48 ()
- J. D. Edixhoven, J. Gupta und H. H. G. Savenije: Recent revisions of phosphate rock reserves and resources: a critique. Earth System Dynamics 5, 2014, S. 491–507
- Azzouz Kchikach, Pierre Andrieux, Mohammed Jaffal, Mostafa Amrhar, Mohammed Mchichid, Baadi Boya, Mbarek Amaghzaz, Thierry Veyrieras und Khadija Iqizou: Les sondages électromagnétiques temporels comme outil de reconnaissance du gisement phosphaté de Sidi Chennane (Maroc): apport à la résolution d’un problème d’exploitation. Comptes Rendus – Geoscience 338, 2006, S. 289–296
- Bruce A. Kennerdy: Surface Mining, Second Edition. Littleton, Colorado, 1990, S. 1–1206 (S. 622)
- Azzouz Kchikach, Mohammed Jaffal, Tahar Aïfa und Lahcen Bahi: Cartographie de corps stériles sous couverture quaternaire par méthode de résistivités électriques dans le gisement phosphaté de Sidi Chennane (Maroc). Comptes Rendus – Geoscience 334, 2002, S. 379–386
- Armand Boujo: About shape and development of sterile bodies in phosphatic deposits. Comptes Rendus – Geoscience 334, 2002, S. 1113–1114
- Nicholas R. Longrich, Xabier Pereda Suberbiola, Nour-Eddine Jalil, Fatima Khaldoune und Essaid Jourani: An abelisaurid from the latest Cretaceous (late Maastrichtian) of Morocco, North Africa. Cretaceous Research 76, 2017, S. 40–52, doi:10.1016/j.cretres.2017.03.021
- Nicholas R. Longrich, David M. Martill und Brian Andres: Late Maastrichtian pterosaurs from North Africa and mass extinction of Pterosauria at the Cretaceous-Paleogene boundary. PLoS Biology 16 (3), 2018, S. e2001663, doi:10.1371/journal.pbio.2001663
- Lászlό Kocsis, Emmanuel Gheerbrant, Mustapha Mouflih, Henri Cappetta, Johan Yans und Mbarek Amaghzaz: Comprehensive stable isotope investigation of marine biogenic apatite from the late Cretaceous–early Eocene phosphate series of Morocco. Palaeogeography, Palaeoclimatology, Palaeoecology 394, 2014, S. 74–88
- Johan Yans, M'Barek Amaghzaz, Baâdi Bouya, Henri Cappetta, Paola Iacumin, László Kocsis, Mustapha Mouflih, Omar Selloum, Sevket Sen, Jean-Yves Storme und Emmanuel Gheerbrant: First carbon isotope chemostratigraphy of the Ouled Abdoun phosphate Basin, Morocco; implications for dating and evolution of earliest African placental mammals. Gondwana Research 25, 2014, S. 257–269
- Nathalie Bardet, Emmanuel Gheerbrant, Abdelmajid Noubhani, Henri Cappetta, Stéphane Jouve, Estelle Bourdon, Xabier Pereda Suberbiola, Nour-Eddine Jalil, Peggy Vincent, Alexandra Houssaye, Floréal Solé, Khadija El Houssaini, Sylvain Adnet, Jean-Claude Rage, France de Lapparent de Broin, Jean Sudre, Baâdi Bouya, Mbarek Amaghzaz und Saïd Meslouh: Les Vertébrés des Phosphates cretacés-paléogènes (70,6 – 46,6 Ma) du Maroc. In: Samir Zouhri (Hrsg.): La Paléontologie des Vertébrés du Maroc. Paris, Mémoires de la Société Géologique de France, nouvelle série, 180, 2017, S. 351–452
- Emmanuel Gheerbrant, Arnaud Schmitt und László Kocsis: Early African Fossils Elucidate the Origin of Embrithopod Mammals. Current Biology 28 (13), 2018, S. 2167–2173, doi:10.1016/j.cub.2018.05.032
- E. Jourani, F. Khaldoune und N.-E. Jalil: Les fossiles des phosphates du Maroc, une mémoire de 24 millions d’années, valorisation et préservation par l’OCP. In: K. El Hariri und H. Chennaoui Aoudjehane (Hrsg.): Promoting Geological Heritage: Challenges and Issues. La Conférence Internationale RALI 2015, Marrakech 05-11 octobre 2015. Notes et Mémoires du Service Géologique du Maroc, Numéro Spécial dédié au Patrimoine Géologique du Maroc, 2018, S. 103–136
- Edouard Boureau: Contribution à l’étude paléoxylologique de l’Afrique du Nord (III): Pterocarpoxylon arambourgii n. gen., n. sp., bois silicifié de Leguminoseae–Papilioneae découvert dans les phosphates yprésiens de Khouribga (Maroc). Bulletin du Muséum National d’Histoire Naturelle 2 (23), 1951, S. 552–557 ()
- Raymond Rauscher und Jeanne Doubinger: Les dinokystes du Maestrichtien phosphaté du Maroc. Sciences Géologiques. Bulletin 35 (3), 1982, S. 97–116
- Marie-Jo Soncini und Raymond Rauscher: Morphologies particuliéres chez les dinokystes des genres lsabelidinium, Manumiella et Dinogymnium dans les phosphates maastrichtiens et paléocénes du Maroc. Bulletin des Centres de Recherches Exploration-Production Elf-Aquitaine 14 (2), 1990, S. 583–596
- Marie-Jo Soncini: Three new dinoflagellate cysts from the Moroccan Paleocene-Eocene phosphates. Review of Palaeobotany and Palynology 70, 1992, S. 325–338
- Jean-Michel Pacaud und Patrice Lebrun: Les phosphates du Maroc (2): des faunes d’invertébrés dominées par les bivalves et les gastéropodes. Fossiles 40, 2019, S. 17–37
- Henri Cappetta: Additions à la faune de sélaciens fossiles du Maroc. 1: Sur la présence des genres Heptranchias, Alopiaset Odontorhytisdans l’Yprésien des Ouled Abdoun. Geobios 14 (5), 1981, S. 563–575
- Abdelmajid Noubhani und Henri Cappetta: Les Orectolobiformes, Carcharhiniformes et Myliobatiformes (Elasmobranchii, Neoselachii) des bassins à phosphate du Maroc (Maastrichtien-Lutétien basal). Systématique, biostratigraphie, évolution et dynamique des faunes. Palaeo Ichthyologica 8, 1997, S. 1–327
- Jaques Herman: Contribution à la connaissance de la faune ichthyologique des phosphates du Maroc. Annales de la Société Géologique de Belgiqiie 95, 1972, S. 271–284
- Bruce J. MacFadden, Joann Labs-Hochstein, Irvy Quitmyer und Douglas S. Jones: Incremental growth and diagenesis of skeletal parts of the lamnoid shark Otodus obliquus from the early Eocene (Ypresian) of Morocco. Palaeogeography, Palaeoclimatology, Palaeoecology 206, 2004, S. 179–192
- Henri Cappetta, Sylvain Adnet, Driss Akkrim und Mohamed Amalik: New Squalicorax species (Neoselachii: Lamniformes) from the Lower Maastrichtian of Ganntour phosphate deposit, Morocco. Palaeovertebrata 38 (2), 2014, S. e3, doi: 10.18563/pv.38.2.e3
- Henri Cappetta: Révision de Cestracion duponti Winkler, 1874 (Selachii, Batomorphii) du Bruxellien de Woluwe-Saint-Lambert (Eocène moyen de Belgique). Mededelingen van de Werkgroep voor Tertiaire en Kwartaire Geologie 19 (4), 1982, S. 113–125
- Henri Cappetta: Découverte du genre Gymnura (Batomorphii, Myliobatiformes) dans le Thanétien des Ouled Abdoun, Maroc. Observations sur la denture de quelques espèces actuelles. Geobios 17 (5), 1984, S. 631–635
- Henri Cappetta: Un nouveau genre de sélacien (Batomorphii, Myliobatiformes) de l’Yprésien des Ouled Abdoun, Maroc. Geobios 19 (5), 1986, S. 635–640
- Henri Cappetta: Types dentaires adaptatifs chez les sélaciens actuels et postpaléozoïques. Palaeovertebrata 16 (2), 1986, S. 57–76
- Abdelmajid Noubhani und Henri Cappetta: Revision des Rhombodontidae (Neoselachi, Batomrphii) des bassins a phosphate du Maroc. Palaeovertebrala 23 (1–4), 1994, S. 1–49
- Henri Cappetta: Nouveaux Rhinobatoidei (Neoselachii, Rajiformes) à denture spécialisée du Maastrichtien du Maroc. Remarques sur l’évolution dentaire des Rajiformes et des Myliobatiformes. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 187 (1), 1992, S. 31–52
- Gérard R. Case und Jaques Herman: A dorsal fin spine of the chimeroid Edaphodon cf. bucklandi (Agassiz) from the Eocene of Morocco. Bulletin de la Société belge de Géologie, Paléontologie et Hydrologie 82 (3), 1973, S. 445–449
- Romain Vullo, Lionel Cavin, Bouziane Khallouf, Mbarek Amaghzaz, Nathalie Bardet, Nour-Eddine Jalil, Essaid Jourani, Fatima Khaldoune und Emmanuel Gheerbran: A unique Cretaceous–Paleogene lineage of piranha-jawed pycnodont fishes. Scientific Reports 7, 2017, S. 6802, doi:10.1038/s41598-017-06792-x
- Lionel Cavin, Nathalie Bardet, Henri Cappetta, Emmanuel Gheerbrant, Sidi Mohamed Iarochène und Jean Sudre: A new Palaeocene albulid (Teleostei: Elopomorpha) from the Ouled Abdoun phosphatic basin, Morocco. geological Magazine 137 (5), 2000, S. 583–591
- Bouziane Khalloufi, Khadija El Houssaini Darif, Essaid Jourani, Fatima Khaldoune und Nour-Eddine Jalil: A new Palaeocene Megalopidae (Teleostei, Elopomorpha) from the phosphate basins of Morocco. Historical Biology 31 (9), 2019, S. 1256–1265, doi:10.1080/08912963.2018.1443327
- Jean-Claude Rage und Georges Wouters: Découverte du plus ancien Palaeopheide (Reptilia, Serpentes) dans le Maestrichtien du Maroc. Geobios 12, 1979, S. 293–296
- Alexandra Houssaye, Jean-Claude Rage, Nathalie Bardet, Peggy Vincent, Mbarek Amaghzaz und Said Meslouh: New highlights about the enigmatic marine snake Palaeophis maghrebianus (Palaeophidae) from the Ypresian phosphates (Middle Eocene) of Morocco. Palaeontology 56 (3), 2013, S. 647–661, doi:10.1111/pala.12008
- Vivian de Buffrénil, Nathalie Bardet, Xabier Pereda Suberbiola und Baâdi Bouya: Specialization of bone structure in Pachyvaranus crassispondylus Arambourg, 1952, an aquatic squamate from the Late Cretaceous of the southern Tethyan margin. Lethaia 41, 2008, S. 59–69, doi:10.1111/j.1502-3931.2007.00042.x
- Alexandra Houssaye, Nathalie Bardet, Jean-Claude Rage, Xabier Pereda Suberbiola, Baâdi Bouya, Mbarek Amaghzaz und Moamed Amalik: A review of Pachyvaranus crassispondylus Arambourg, 1952, a pachyostotic marine squamate from the latest Cretaceous phosphates of Morocco and Syria. Geological Magazine 148 (2), 2011, S. 237–249, doi:10.1017/S0016756810000580
- Nathalie Bardet, Xabier Pereda Suberbiola, Mohamed Iarochène, Baâdi Bouya und Mbarek Amaghzaz: A new species of Halisaurus from the Late Cretaceous phosphates of Morocco, and the phylogenetical relationships of the Halisaurinae (Squamata: Mosasauridae). Zoological Journal of the Linnean Society 143, 2005, S. 447–472
- Michael J. Polcyn, Johan Lindgren, Nathalie Bardet, Dirk Cornelissen, Louis Verding und Anne S. Schulp: Description of new specimens of Halisaurus arambourgi Bardet & Pereda Suberbiola, 2005 and the relationships of Halisaurinae. Bulletin de la Societe Geologique de France 183 (2), 2012, S. 123–136
- Nicholas R.Longrich, Nathalie Bardet, Fatima Khaldoune, Oussama Khadiri Yazami und Nour-Eddine Jalil: Pluridens serpentis, a new mosasaurid (Mosasauridae: Halisaurinae) from the Maastrichtian of Morocco and implications for mosasaur diversity. Cretaceous Research, 2021, doi:10.1016/j.cretres.2021.104882
- Catherine R. C. Strong, Michael W. Caldwell, Takuya Konishi und Alessandro Palci: A new species of longirostrine plioplatecarpine mosasaur (Squamata: Mosasauridae) from the Late Cretaceous of Morocco, with a re-evaluation of the problematic taxon ‘Platecarpus’ ptychodon. Journal of Systematic Palaeontology, 2020, doi:10.1080/14772019.2020.1818322
- Nathalie Bardet, Xabier Pereda Suberbiola, Mohamed Iarochène, Fatima Bouyahyaoui, Baâdi Bouya und Mbarek Amaghzaz: Mosasaurus beaugei Arambourg, 1952 (Squamata, Mosasauridae) from the Late Cretaceous phosphates of Morocco. Geobios 37, 2004, S. 315–324, doi:10.1016/j.geobios.2003.02.006
- Trevor H. Rempert, Alexander P. M. Vinkeles Melchers, Ashley N. Rempert, Muhammad R. Haque und Andrew R. Armstrong: Occurrence of Mosasaurus hoffmannii Mantell, 1829 (Squamata, Mosasauridae) in the Maastrichtian Phosphates of Morocco. Journal of Paleontological Sciences 10, 2022, S. JPS.C.2022.0001 ()
- Aaron R. H. Leblanc, Michael W. Caldwell und Nathalie Bardet: A new mosasaurine from the Maastrichtian (Upper Cretaceous) phosphates of Morocco and its implications for mosasaurine systematics. Journal of Vertebrate Paleontology 32 (1), 2012, S. 82–104, doi:10.1080/02724634.2012.624145
- Nathalie Bardet, Xabier Pereda Suberbiola, Anne S. Schulp und Baâdi Bouya: New material of Carinodens (Squamata, Mosasauridae) from the Maastrichtian (Late Cretaceous) phosphates of Morocco. Bulletin of the Fort Hays State University, Special Issue 3, 2008, S. 29–36
- Anne S. Schulp, Nathalie Bardet und Baâdi Bouya: A new species of the durophagous mosasaur Carinodens (Squamata, Mosasauridae) and additional material of Carinodens belgicus from the Maastrichtian phosphates of Morocco. Netherlands Journal of Geosciences — Geologie en Mijnbouw 88 (3), 2009, S. 161–167
- Nicholas R. Longrich, Nathalie Bardet, Anne S. Schulp und Nour-Eddine Jalil: Xenodens calminechari gen. et sp. nov., a bizarre mosasaurid (Mosasauridae, Squamata) with shark-like cutting teeth from the upper Maastrichtian of Morocco, North Africa. Cretaceous Research, 2021, S. 104764, doi:10.1016/j.cretres.2021.104764
- Nathalie Bardet, Alexandra Houssaye, Peggy Vincent, Xabier Pereda Suberbiola, Mbarek Amaghzaz, Essaid Jourani und Saïd Meslouh: Mosasaurids (Squamata) from the Maastrichtian Phosphates of Morocco: Biodiversity, palaeobiogeography and palaeoecology based on tooth morphoguilds. Gondwana Research 27, 2015, S. 1068–1078, doi:10.1016/j.gr.2014.08.014
- Peggy Vincent, Nathalie Bardet, Xabier Pereda Suberbiola, Baâdi Bouya, Mbarek Amaghzaz und Saïd Meslouh: Zarafasaura oceanis, a new elasmosaurid (Reptilia: Sauropterygia) from the Maastrichtian Phosphates of Morocco and the palaeobiogeography of latest Cretaceous plesiosaurs. Gondwana Research 19, 2011, S. 1062–1073, doi:doi:10.1016/j.gr.2010.10.005
- Dean R. Lomax und William R. Wahl: A new specimen of the elasmosaurid plesiosaur Zarafasaura oceanis from the Upper Cretaceous (Maastrichtian) of Morocco. Paludicola 9 (2), 2013, S. 97–109
- Peggy Vincent, Nathalie Bardet, Alexandra Houssaye, Mbarek Amaghzaz und Saïd Meslouh: New plesiosaur specimens from the Maastrichtian Phosphates of Morocco and their implications for the ecology of the latest Cretaceous marine apex predators. Gondwana Research 24, 2013, S. 796–805, doi:10.1016/j.gr.2012.11.011
- Nathalie Bardet, J. Falconnet, V. Fischer, Alexandra Houssaye, Stéphane Jouve, Xabier Pereda Suberbiola, A. Pérez-García, J.-C. Rage und Peggy Vincent: Mesozoic marine reptile palaeobiogeography in response to drifting plates. Gondwana Research 26, 2014, S. 869–887, doi:10.1016/j.gr.2014.05.005
- Nathalie Bardet, Nour-Eddine Jalil, France de Lapparent de Broin, Damien Germain, Olivier Lambert und Mbarek Amaghzaz: A Giant Chelonioid Turtle from the Late Cretaceous of Morocco with a Suction Feeding Apparatus Unique among Tetrapods. PLoS ONE 8 (7), 2013, S. e63586, doi:10.1371/journal.pone.0063586
- Eugene S. Gaffney und Haiyan Tong: Phosphatochelys, a New Side-Necked Turtle (Pelomedusoides: Bothremydidae) from the Paleocene of Morocco. Bulletin of the American Museum of Natural History 279, 2003, S. 644–659
- Eugene S. Gaffney und Haiyan Tong: Redescription of the Skull of Ummulisani rutgersensis Gaffney, Tong, and Meylan, 2006, a Bothremydid Side-Necked Turtle from the Eocene of Morocco. American Museum Novitates 3615,2008, S. 1–20
- Eugene S. Gaffney, Haiyan Tong und Peter A. Meylan: Evolution of the side-necked turtles: the families Bothremydidae, Euraxemydidae, and Araripemydidae. Bulletin of the American Museum of Natural History, 300, 2006, S. 1–700
- France de Lappareat de Broin: African chelonians from the Jurassic to the present: Phase of development and preliminary catalogue of the fossil record. Palaeontologica africana 36, 2000, S. 43–82
- France de Lapparent de Broin, Nathalie Bardet, Mbarek Amaghzaz und Saïd Meslouh: A strange new chelonioid turtle from the Latest Cretaceous Phosphates of Morocco. Comptes Rendus Palevol 13, 2014, S. 87–95, doi:10.1016/j.crpv.2013.07.008
- Haiyan Tong und Ren Hirayama: First Cretaceous dermachelyid turtle from Africa. Revue de Paléobiologie 9 (spec.), 2004, S. 55–59
- Ren Hirayama und Haiyan Tong: Osteopygis (Testudines, Cheloniidae) from the Lower Tertiary of the Ouled Abdoun phosphate basin, Morocco. Palaeontology 46, 2003, S. 85–98
- James F. Parham: A reassesment of the referral of sea turtle skulls to the genus Osteopygis (Late Creatceous, New Jersey, USA). Journal of Vertebrate Paleontology 25 (1), 2005, S. 71–77
- Nour-Eddine Jalil, France de Lapparent de Broin, Nathalie Bardet, Renaud Vacant, Baâdi Bouya, Mbarek Amaghzaz und Saïd Meslouh: Euclastes acutirostris, a new species of littoral turtle (Cryptodira, Cheloniidae) from the Palaeocene phosphates of Morocco (Oulad Abdoun Basin, Danian-Thanetian). Coptes Rendus Palevol 8, 2009, S. 447–459, doi:10.1016/j.crpv.2009.03.002
- Haiyan Tong und Ren Hirayama: A new species of Argillochelys (Testudines: Cryptodira: Cheloniidae) from the Ouled Abdoun phosphate basin, Morocco. Bulletin de la Société Géologique de France 179 (6), 2008, S. 623–630
- Haiyan Tong und Ren Hirayama: A new species of Tasbacka (Testudines: Cryptodira: Cheloniidae) from the Paleocene of Ouled Abdoun phosphate basin, Morocco. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 5, 2002, S. 277–294
- Haiyan Tong, Ren Hirayama und Jerôme Tabouelle: Puppigerus camperi (Testudines: Cryptodira: Cheloniidae) from the Ypresian (Early Eocene) of Ouled Abdoun basin, Morocco. Bulletin de la Société Géologique de France 183 (6), 2012, S. 635–640
- Haiyan Tong und Peter Meylan: Morphology and Relationships of Brachyopsemys tingitana gen. et sp. nov. from the Early Paleocene of Morocco and Recognition of the New Eucryptodiran Turtle Family: Sandownidae. In: Donald B. Brinkman, Patricia A. Holroyd und James D. Gardner (Hrsg.): Morphology and Evolution of Turtles. Springer Science+Business, 2013, S. 187–212
- Stéphane Jouve, Mohamed Iarochène, Baâdi Bouya und Mbarek Amaghzaz: A new species of Dyrosaurus (Crocodylomorpha, Dyrosauridae) from the early Eocene of Morocco: phylogenetic implications. Zoological Journal of the Linnean Society 148, 2006, S. 603–656
- Stéphane Jouve, Baâdi Bouya und Mbarek Amaghzaz: A long-snouted dyrosaurid (Crocodyliformes, Mesoeucrocodylia) from the Paleocene of Morocco: Phylogenetic and palaeobiogeographic implications. Palaeontology 51(2), 2008, S. 281–294, doi:10.1111/j.1475-4983.2007.00747.x
- Stéphane Jouve, Baâdi Bouya und Mbarek Amaghzaz: A short-snouted dyrosaurid (Crocodyliformes, Mesoeucrocodylia) from the Palaeocene of Morocco. Palaeontology 48 (2), 2005, S. 359–369
- Stéphane Jouve, Mohamed Iarochène, Baâdi Bouya und Mbarek Amaghzaz: A new dyrosaurid crocodyliform from the Palaeocene ofMorocco and a phylogenetic analysis of Dyrosauridae. Acta Palaeontologica Polonica 50 (3), 2005, S. 581–594
- Stéphane Jouve und Nour-Eddine Jalil: Paleocene resurrection of acrocodylomorph taxon: Biotic crises, climatic and sea level fluctuations. GondwanaResearch, 2020, doi:10.1016/j.gr .2020.03.010
- Stéphane Jouve, Nathalie Bardet, Nour-Eddine Jalil, Baâdi Bouya und Mbarek Amaghzaz: The oldest African crocodylian: Phylogeny, paleobiogeography, and differential survivorship of marine reptils through the Cretaceous-Tertiary boundary. Journal of Vertebrate Paleontology 28 (2), 2008, S. 409–421
- Stéphane Hua und Stéphane Jouve: A primitive marine gavialoid from the Paleocene of Morocco. Journal of Vertebrate Paleontology 24 (2), 2004, S. 341–350
- Stéphane Jouve, Mohamed Iarochène, Baâdi Bouya und Mbarek Amaghzaz: New material of Argochampsa krebsi (Crocodylia: Gavialoidea) from the Lower Paleocene of the Oulad Abdoun Basin (Morocco): phylogenetic implications. Geobios 39, 2006, S. 817–832, doi:10.1016/j.geobios.2005.07.003
- Stéphane Jouve, Baâdi Bouya, Mbarek Amaghzaz und Saïd Meslouh: Maroccosuchus zennaroi(Crocodylia: Tomistominae) from the Eocene of Morocco: phylogenetic and palaeobiogeographical implications of thebasalmost tomistomine. Journal of Systematic Palaeontology 32 (5), 2015, S. 421–445, doi:10.1080/14772019.2014.913078
- Xabier Pereda Suberbiola, Nathalie Bardet, Stéphane Jouve S, Mohamed Iarochène, Baâdi Bouya und Mbarek Amaghzaz: A new azhdarchid pterosaur from the Late Cretaceous phosphates of Morocco. Geological Society, London, Special Publications 217 (3), 2003, S. 79–90
- Xabier Pereda Suberbiola, Nathalie Bardet, Mohamed Iarochène, Baâdi Bouya und Mbarek Amaghzaz: The first record of a sauropod dinosaur from the Late Cretaceous phosphates of Morocco. Journal of African Earth Sciences 40, 2004, S. 81–88, doi:10.1016/j.jafrearsci.2004.07.002
- Eric Buffetaut, François Escuillié und Burkhart Pohl: First theropod dinosaur from the Maastrichtian phosphates of Morocco. Kaupia 14, 2005, S. 3–8
- Nicholas R. Longrich, Xabier Pereda Suberbiola, R. Alexander Pyron und Nour-Eddine Jalil: The first duckbill dinosaur (Hadrosauridae: Lambeosaurinae) from Africa and the role of oceanic dispersal in dinosaur biogeography. Cretaceous Research 120, 2020, S. 104678, doi:10.1016/j.cretres.2020.104678
- Estelle Bourdon, Mbarek Amaghzaz und Baâdi Bouya: Pseudotoothed Birds (Aves, Odontopterygiformes) from the Early Tertiary of Morocco. American Museum Novitates 3704, 2010, S. 1–71
- Estelle Bourdon, Baâdi Bouya und Mohamed Iarochène: Earliest African neornithine bird: A new species of Prophaetontidae (Aves) from the Paleocene of Morocco. Journal of Vertebrate Paleontology 25 (1), 2005, S. 157–170
- Estelle Bourdon, Cécilie Mourer-Chauviré, Mbarek Amaghzaz und Baâdi Bouya: New specimens of Lithoptila abdounensis (Aves, Prophaetontidae) from the Lower Paleogene of Morocco. Journal of Vertebrate Paleontology 28 (3), 2008, S. 751–761
- Estelle Bourdon, Mbarek Amaghzaz und Baâdi Bouya: A new seabird (Aves, cf. Phaethontidae) from the Lower Eocene phosphates of Morocco. Geobios 41, 2008, S. 455–459, doi:10.1016/j.geobios.2007.11.002
- Emmanuel Gheerbrant, Jean Sudre, Mohamed Iarochène und Abdelkader Moumni: First ascertained African “Condylarth” mammals (primitive ungulates: cf. Bulbulodentata and cf. Phenacodonta) from the earliest Ypresian of the Ouled Abdoun Basin, Morocco. Journal of Vertebrate Paleontology 21 (1), 2001, S. 107–118
- Emmanuel Gheerbrant: Primitive Ungulates („Condylarthra“ and Stem Paenungulata). In: Lars Werdelin und William Joseph Sanders (Hrsg.): Cenozoic Mammals of Africa. University of California Press, Berkeley, Los Angeles, London, 2010, S. 563–571
- Emmanuel Gheerbrant, Mbarek Amaghzaz, Baadi Bouya, Florent Goussard und Charlène Letenneur: Ocepeia (Middle Paleocene of Morocco): The Oldest Skull of an Afrotherian Mammal. PLoS ONE 9 (1), 2014, S. e89739, doi:10.1371/journal.pone.0089739
- Emmanuel Gheerbrant, Andrea Filippo und Arnaud Schmitt: Convergence of Afrotherian and Laurasiatherian Ungulate-Like Mammals: First Morphological Evidence from the Paleocene of Morocco. PLoS ONE 11 (7), 2016, S. e0157556, doi:10.1371/journal.pone.0157556
- Emmanuel Gheerbrant: Paleocene emergence of elephant relatives and the rapid radiation of African ungulates. PNAS 106 (26), 2009, S. 10717–10721
- Emmanuel Gheerbrant, Baadi Bouya und Mbarek Amaghzaz: Dental and cranial anatomy of Eritherium azzouzorum from the Paleocene of Marocco, earliest known proboscidean material. Palaeontographica, Abteilung A 297 (5/6), 2012, S. 151–183.
- Emmanuel Gheerbrant, Jean Sudre und Henri Cappetta: A Palaeocene proboscidean from Morocco. Nature 383, 1996, S. 68–70
- Emmanuel Gheerbrant: The oldest known proboscidean and the role of Africa in the radiation of modern orders of placentals. Bulletin of the Geological Society of Denmark 44, 1998, S. 181–185
- Emmanuel Gheerbrant, Jean Sudre, Henri Cappetta und Gérard Bignot: Phosphatherium escuilliei du Thanétien du Bassin des Ouled Abdoun (Maroc), plus ancien proboscidien (Mammalia) d'Afrique. Geobios 30 (2), 1998, S. 247–269
- Emmanuel Gheerbrant, Jean Sudre, Pascal Tassy, Mbarek Amaghzaz, Baâdy Bouya und Mohamed Iarochène: Nouvelles données sur Phosphatherium escuilliei (Mammalia, Proboscidea) de l’Éocène inférieur du Maroc, apports à la phylogénie des Proboscidea et des ongulés lophodontes. Geodiversitas 27 (2), 2005, S. 239–333
- Emmanuel Gheerbrant, Jean Sudre, Henri Cappetta, Mohamed Iarochène, Mbarek Amaghzaz und Baâdi Bouya: A new large mammal from the Ypresian of Morocco: Evidence of surprising diversity of early proboscideans. Acta Palaeontologica Polonica 47 (3), 2002, S. 493–506.
- Emmanuel Gheerbrant, Mohamed Iarochène, Mbarek Amaghzaz und Baâdi Bouya: Early African hyaenodontid mammals and their bearing on the origin of the Creodonta. Geological Magazine 143 (4), 2006, S. 475–489, doi:10.1017/S0016756806002032
- Floréal Solé, Emmanuel Gheerbrant, Mbarek Amaghzaz und Baâdi Bouya: Further evidence of the African antiquity of hyaenodontid („Creodonta“, Mammalia) evolution. Zoological Journal of the Linnean Society 156, 2009, S. 827–846
- Henri Cappetta, Nathalie Bardet, Xabier Pereda Suberbiola, Sylvain Adnet, Driss Akkrim, Mohamed Amalik und Aziza Benabdallah: Marine vertebrate faunas from the Maastrichtian phosphates of Benguérir (Ganntour Basin, Morocco): Biostratigraphy, palaeobiogeography and palaeoecology. Palaeogeography, Palaeoclimatology, Palaeoecology 409, 2014, S. 217–238, doi:10.1016/j.palaeo.2014.04.020
- Abdelmajid Noubhani: The selachians’ faunas of the Moroccan phosphate deposits and the KT mass-extinctions. Historical Biology 22, (1), 2010, S. 71–77, doi:10.1080/08912961003707349
- Jörg Trappe: Microfacies zonation and spatial evolution of a carbonate ramp: marginal Moroccan phosphate sea during the Paleogene. Geologische Rundschau 81, 1992, S. 105–126
- André Charrière, Hamid Haddoumi, Pierre-Olivier Mojon, Jacky Ferrière, Daniel Cuche und Lamia Zili: Mise en évidence par charophytes et ostracodes de l’âge Paléocène des dépôts discordants sur les rides anticlinales de la région d’Imilchil (Haut Atlas, Maroc): conséquences paléogéographiques et structurales. Comptes Rendus Palevol 8, 2009, S. 9–19, doi:10.1016/j.crpv.2008.11.006
- Lászlό Kocsis, Emmanuel Gheerbrant, Mustapha Mouflih, Henri Cappetta, Alex Ulianov, Massimo Chiaradiaf und Nathalie Bardet: Gradual changes in upwelled seawater conditions (redox, pH) from the late Cretaceous through early Paleogene at the northwest coast of Africa: Negative Ce anomaly trend recorded in fossil bio-apatite. Chemical Geology 421, 2016, S. 44–54, doi:10.1016/j.chemgeo.2015.12.001
- C. Lécuyer, P. Grandjean, J. R. O'Neil, H. Cappetta und F. Martineau: Thermal excursions in the ocean at the Cretaceous-Tertiary boundary (northern Morocco): δ 16O record of phosphatic fish debris. Palaeogeography, Palaeoclimatology, Palaeoecologv 105, 1993, S. 235–243
- Vincent Balter, Christophe Lécuyer und Jean-Alix Barrat: Reconstructing seawater Sr/Ca during the last 70 My using fossil fish tooth enamel. Palaeogeography, Palaeoclimatology, Palaeoecology 310, 2011, S. 133–138
- Jeremy E. Martin, Peggy Vincent, Theo Tacail, Fatima Khaldoune, Essaid Jourani und Nathalie Bardet: Calcium Isotopic Evidence for Vulnerable Marine Ecosystem Structure Prior to the K/Pg Extinction. Current Biology 27, 2017, S. 1641–1644, doi:10.1016/j.cub.2017.04.043
- B. Mohr: Palynologischer Nachweis eines Mangrovenbiotops in der Südatlas-Randzone (Marokko) und seine paläoökologische Ausdeutung. Documenta naturae 33, 1986, S. 20–28
- Hans-Georg Herbig und H. J. Gregor: The mangrove-forming palm Nypa from the early Paleogene of southern Morocco. Paleoenvironment and paleoclimate. Géologie Mediteranéene 17 (2), 1990, S. 123–137
- Marie-Françoise Ollivier-Pierre: La microflore du Paléocène et de l’Éocène des séries phosphatées des Ganntour (Maroc). Sciences Géologiques. Bulletin 35 (3), 1982, S. 117–127
- László Kocsis, Alex Ulianov, Mustaphah Mouflih, Fatima Kaldoune und Emmanuel Gheerbrant: Geochemical investigation of the taphonomy, stratigraphy, and palaeoecology of the mammals from the Ouled Abdoun Basin (Paleocene-Eocene of Morocco), Palaeogeography, Palaeoclimatology, Palaeoecology 577, 2021, S. 110523, doi:10.1016/j.palaeo.2021.110523
- Robert Ambroggi und Camille Arambourg: Sur le Maestrichtien et l’Ėoocéne inférieur de la vallée du Sous (Sud Marocain). Comptes Rendus de l’Académie des Sciences de Paris 233, 1951, S. 170–171 ()
- Oliver W. M. Rauhut und Christa Werner: First record of a Maastrichtian sauropod dinosaur from Egypt. Palaeontologia africana 34, 1997, S. 63–67
- A. A. Tantawy, G. Keller, T. Adatte, W. Stinnesbeck, A. Kassab und P. Schulte: Maastrichtian to Paleocene depositional environment of the Dakhla Formation, Western Desert, Egypt: sedimentology, mineralogy, and integrated micro- and macrofossil biostratigraphies. Cretaceous Research 22,2001, S. 795–827
- M. H. El-Azabi und A. El-Araby: Depositional cycycles: an approach to the sequence stratigraphy of the Dakhla Formation, west Dakhla-Farafra stretch, Western Desert, Egypt. Journal of African Earth Sciences 30 (4), 2000, S. 971–996
- Hesham M. Sallam, Patrick M. O'Connor, Mahmoud Kora, Joseph J. W. Sertich, Erik R. Seiffert, Mahmoud Faris, Khaled Ouda, Iman El-Dawoudi, Sara Saber und Sanaa El-Sayed: Vertebrate paleontological exploration of the Upper Cretaceous succession in the Dakhla and Kharga Oases, Western Desert, Egypt. Journal of African Earth Sciences 117, 2016, S. 223–234, doi:10.1016/j.jafrearsci.2016.01.022
- Jo De Waele, Felice Di Gregorio, Nabil Gasmi, Maria Teresa Melis und Mohamed Talbi: Geomorphosites of Tozeur region (south-west Tunisia). Il Quaternario Italian Journal of Quaternary Sciences 18 (1), 2005, S. 221–230
- Stéphane Jouve: A new description of the skull of Dyrosaurus phosphaticus (Thomas, 1893) (Mesoeucrocodylia: Dyrosauridae) from the Lower Eocene of North Africa. Canadian Journal of Earth Science 42, 2005, S. 323–337, doi:10.1139/E05-008
- Nizar Ibrahim, Paul C. Sereno, David J. Varricchio, David M. Martill, Didier B. Dutheil, David M. Unwin, Lahssen Baidder, Hans C. E. Larsson, Samir Zouhri und Abdelhadi Kaoukaya: Geology and paleontology of the Upper Cretaceous Kem Kem Group of eastern Morocco. ZooKeys 928, 2020, S. 1–216, doi:10.3897/zookeys.928.47517
- Prince C. Owusu Agyemanga, Eric M. Roberts, Robert Bussert, David Evans und Johannes Müller: U-Pb detrital zircon constraints on the depositional age and provenance of the dinosaur-bearing Upper Cretaceous Wadi Milk formation of Sudan. Cretaceous Research 97, 2019, S. 52–72, doi:10.1016/j.cretres.2019.01.005
- Christa Werner: Der erste Nachweis von Gymnophionen (Amphibia) in der Kreide (Wadi-Milk-Formation, Sudan). Neues Jahrbuch für Geologie und Paläontologie Monatshefte 10, 1994, S. Jg. 1994 Heft 10 (1994), p. 633–640
- Oliver W. M. Rauhut und Christa Werner: First record of the family Dromaeosauridae (Dinosauria: Theropoda) in the Cretaceous of Gondwana (Wadi Milk Formation, northern Sudan). Paläontologische Zeitschrift 69 (3/4), 1995, S. 475–489
- Oliver W. M. Rauhut: A dinosaur fauna from the Late Cretaceous (Cenomanian) of Northern Sudan. Palaeontologia africana 35, 1999, S. 61–84
- Nicole Klein, Robert Bussert, David Evans, Khalaf Allah O. Salih, Ali A. M. Eisawi, Mutwakil Nafi und Johannes Müller: Turtle remains from the Wadi Milk Formation (Upper Cretaceous) of Northern Sudan. Palaeobiodiversity and Palaeoenvironments 96, 2016, S. 281–303
- Henry Cappetta: Sélaciens nouveaux (Chondrichthyes, Neoselachii) du Paléocène supérieur de la région d'Ouarzazate, Maroc. Paläontologische Zeitschrift 67 (1/2), 1993, S. 109–122
- Marc Augé und Jean-Claude Rage: Herpetofaunas from the Upper Paleocene and Lower Eocene of Morocco. Annales de Paléontologie 92 (1/2), 2006, S. 235–253
- Bernard Sigé, Jean-Jaques Jaeger, Jean Sudre und Monique Vianey-Liaud: Alliatlasius koulchii n. gen., n. sp., primate omomyidé du Paléocène supérieur du Maroc, et les origines des euprimates. Palaeantographica, Abteilung A 214, 1990, S. 31–56
- Emmanuel Gheerbrant: Les mammifères paléocènes du bassin d’Ouarzazate (Maroc). Teil 1: Introduction générale et Palaeoryctidae. Palaeontographica, Abteilung A 224, 1992, S. 67–132; Teil 2: Todralestidae (Proteutheria, Eutheria). Palaeontographica, Abteilung A 231, 1994, S. 133–188; Teil 3: Adapisoriculidae et autres mammifères (Carnivora, ?Creodonta, Condylarthra, ?Ungulata et incertae sedis). Palaeontographica, Abteilung A 237, 1995, S. 39–132
- Emmanuel Gheerbrant: Premières données sur les mammifères „insectivores“ de l’Yprésien du bassin d’Ouarzazate. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 187, 1993, S. 225–242
- Emmanuel Gheerbrant, Jean Sudre, Sevket Sen, Claude Abrial, Bernard Marandat, Bernard Sigé und Moniue Vianey-Liaud: Nouvelles données sur les mammifères du Thanétien et de l’Yprésien du Bassin d’Ouarzazate (Maroc) et leur contexte stratigraphique. Palaeovertebrata 27, 1998, S. 155–202
- Rodolphe Tabuce, Sylvain Adnet, Henri Cappetta, Abdelmajid Noubhani und Frédéric Quillevere: Aznag (bassin d’Ouarzazate, Maroc), nouvelle localité à sélaciens et mammifères de l’Eocène moyen (Lutétien) d’Afrique. Bulletin de la Societe géologique de France 176 (4), 2005, S. 381–400
- Mohammed Mahboubi, R. Ameur, Jean-Yves Crochet und Jean-Jacques Jaeger: El Kohol (Saharan Atlas, Algeria): A new Eocene mammal locality in northwestern Africa. Palaeontographica, Abteilung A 192, 1986, S. 15–49
- J. Sudre: Nouveaux mammifères éocènes du Sahara occidental. Palaeovertebrata 9, 1979, S. 83–115
- Mohammed Adaci, Rodolphe Tabuce, Fateh Mebrouk, Mustapha Bensalah, Pierre-Henri Fabre, Lionel Hautier, Jean-Jacques Jaeger, Vincent Lazzari, M’hammed Mahboubi, Laurent Marivaux, Olga Otero, Stéphane Peigné und Haiyan Tong: Nouveaux sites à vertébrés paléogènes dans la région des Gour Lazib (Sahara nord-occidental, Algérie). Comptes Rendus Palevol 6, 2007, S. 535–544
- Jean-Louis Hartenberger, Claude Martinez und Ahmed Ben Said: Découverte de mammifères d’âge Éocène inférieur en Tunisie centrale. Comptes rendus de l'Académie des sciences. Série 2 301, 1985, S. 649–652 ()
- Rodolphe Tabuce, Laurent Marivaux, Mohammed Adaci, Mustapha Bensalah, Jean-Louis Hartenberger, Mohammed Mahboubi, Fateh Mebrouk, Paul Tafforeau und Jean-Jacques Jaeger: Early Tertiary mammals from North Africa reinforce the molecular Afrotheria clade. Proceedings of the Royal Society B 274, 2007, S. 1159–1166
- Elwyn L. Simons: Eocene and Oligocene mammals of the Fayum, Egypt. In: First International Conference on the Geology of the Tethys, Cairo University, November, 2005, Volume II. Cairo 2005, S. 439–450
- Sylvain Adnet, Henri Cappetta und Rodolphe Tabuce: A Middle–Late Eocene vertebrate fauna (marine fish and mammals) from southwestern Morocco; preliminary report: age and palaeobiogeographical implications. Geological Magazine 147 (6), 2010, S. 860–870, doi:10.1017/S0016756810000348
- Samir Zouhri, Philip D. Gingerich, Najia Elboudali, Samira Sebti, Abdelmajid Noubhani, Meriem Rahali und Saïd Meslouh: New marine mammal faunas (Cetacea and Sirenia) and sea level change in the Samlat Formation, Upper Eocene, near Ad-Dakhla in southwestern Morocco. Comptes Rendus Palevol 13, 2014, S. 599–610, doi:10.1016/j.crpv.2014.04.002
- Samir Zouhri, Bouziane Khalloufi, Estelle Bourdoni, France de Lapparent de Broin, Jean-Claude Rage, Leila M'Haïdrat, Philip D. Gingerich und Najia Elboudali: Marine vertebrate fauna from the late Eocene Samlat Formation of Ad-Dakhla, southwestern Morocco. Geological Magazine 15 (7), 2018, S. 1596–1620, doi:10.1017/S0016756817000759
- Philippe Thomas: Description de quelques fossiles nouveaux ou critiques des terrains tertiaires et secondaires de la Tunisie recueillis en 1885 et 1886. Exploration Scientifique de la Tunisie. Paris: Imprimerie Nationale, 1893, S. 1–46 ()
- Abel Brives: Sur les terrains éocènes dans le Maroc occidental. Comptes rendus de l’Académie des Sciences 140, 1905, S. 395–397 ()
- Abel Brives: Sur le Sénonien et l’Eocène de la bordure nord de l’Atlas marocain. Comptes rendus de l’Académie des Sciences 146. 1908, S. 873–875 ()
- Abel Brives: Le Suessonien dans le Maroc central. Comptes Rendus de l’Académie des Sciences de Paris 168, 1917, S. 776–777 ()
- Léonce Joleaud: Sur l’âge des dépôts de phosphate de chaux du Sud marocain, algérien et tunisien. Comptes Rendus de l’Académie des Sciences de Paris 174, 1922, S. 178–181 ()
- Louis Gentil: Sur l’âge des phosphates marocains. Comptes rendus de l’Académie des Sciences de Paris 174, 1922, S. 42–44 ()
Weblinks
- Centre de recherche en paléontologie Paris: Current work and results from the phosphate Basins of Morocco; zuletzt abgerufen am 1. Juni 2010