Muscheln
Die Muscheln (Bivalvia von dem lateinischen Wort bi-valvius „zwei-klappig“ abgeleitet) sind eine Klasse der Weichtiere (Mollusca). Merkmale der Klasse sind eine aus zwei kalkigen Klappen bestehende Schale und ein weitgehend reduzierter Kopf. Sie leben weltweit in Salzwasser (zu 80 Prozent), Brackwasser und Süßwasser und sind meist zwischen 0 und 100 m, selten bis 11.000 m Wassertiefe zu finden. Muscheln leben im Meeresgrund, sind an ihm festgewachsen oder liegen frei auf ihm. Die meisten Muschelarten ernähren sich von Plankton, das sie mit ihren Kiemen aus dem Wasser filtern. Die Lebenserwartung reicht von ca. 1 Jahr bis zu mehr als 500 Jahren.
| Muscheln | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Schalen verschiedener Meeresmuschelarten | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Bivalvia | ||||||||||||
| Linnaeus, 1758 |
Der Mensch nutzt Muscheln als Nahrungsmittel und Muschelschalen als Ausgangsmaterial für Schmuck z. B. als Perlenlieferant, Souvenir und früher auch als Muschelgeld. Darüber hinaus werden Muscheln auch in Teichen zur Reinigung verwendet.
Die Klasse enthält etwa 7500 bis 10.000 rezente und 20.000 fossile Arten, die in 106 Familien eingeteilt werden. Nach Huber (2010) ist heute von weltweit etwa 8000 marinen Arten in vier Unterklassen und 99 Familien mit 1100 Gattungen auszugehen. Die Familie der Venusmuscheln ist mit mehr als 680 Arten die größte marine Familie. Aus dem Süßwasser sind weitere etwa 1200 Arten in sieben zusätzlichen Familien bekannt. Die Fluss- und Teichmuscheln stellen hier mit etwa 700 Arten die artenreichste Familie.
Wissenschaftliche Bezeichnung
Der wissenschaftliche Name „Bivalvia“ (Carl von Linné, 1758) ist von der zweigeteilten Kalkschale abgeleitet, die ihren Körper je nach Art mehr oder weniger schützend umhüllt.
Die Bezeichnung „Lamellibranchia“ Henri Marie Ducrotay de Blainville 1824 wurde nach der Gestaltung der Kiemen (Blattkiemen) gewählt und ist in der wissenschaftlichen Literatur am häufigsten zu finden. Weitere vor allem historische Namen sind „Pelecypoda“ August Goldfuß 1820 nach der Gestaltung des Fußes oder „Acephale“ Georges Cuvier 1798 aufgrund des fehlenden Kopfes oder „Conchifera“ Jean-Baptiste de Lamarck 1818, wegen der Muschelschalen (Schalenträger).
Die Begriffe sind nicht ganz synonym, Probleme bilden die Abgrenzungen der Gruppen, so umfasste z. B. „Bivalvia“ für Linné auch die Armfüßer (Brachiopoden). Die deutsche Bezeichnung Muscheln wird nicht nur bei Weichtieren verwendet, sondern auch etwa für die zu den Krebsen gehörenden Entenmuscheln.
Körperbau
Weichkörper
Bei Muscheln ist der Kopf reduziert. Der Weichkörper der Muschel wird beidseitig von den Mantellappen bedeckt und umschließt die Kiemen und die meist getrenntgeschlechtlichen Gonaden. Das Kreislaufsystem mit einem Herzen ist offen. Typische Muskeln sind die zwei getrennten Schließmuskeln, die die beiden Schalenhälften zusammenziehen, die Fußrückziehmuskeln und die Mantelrückziehmuskeln. Der Fuß der Muscheln ist beweglich und mit Schleimdrüsen ausgestattet. Das Nervensystem besitzt zwei Nervenzellhäufungen: das Pedal- und das Viszeralganglion.
Rudimente des Kopfes
Aufgrund der ursprünglich im Sediment grabenden Lebensweise ist der Kopf mit Ausnahme der Mundregion zurückgebildet (deswegen nannte Frédéric Cuvier 1798 die Muscheln „Acephala“, die Kopflosen). Im Vergleich mit den anderen Weichtieren nur noch rudimentär sind die Tentaktel, die Kiefer, die Radula und die Zunge. Die Schlunddrüsen sind weitgehend reduziert. Neben der Mundöffnung sitzen noch paarige, flache Mundlappen. Auf diesen Mundlappen sitzen bei ursprünglichen Formen (wie den Nussmuscheln) noch bewimperte Taster, die Nahrungspartikel zur Mundöffnung transportieren.
Mantel

Der Weichkörper der Muschel wird beidseitig von den Mantellappen bedeckt und geschützt. Der von den beiden Mantellappen gebildete Raum wird als Mantelraum bezeichnet.
Der Mantelrand besteht aus drei Falten, die unterschiedliche Aufgaben erfüllen: Die äußerste Randfalte bildet Schale und Schalenhaut (Periostracum), die mittlere dient der Wahrnehmung sensorischer Aufgaben (z. B. Punktaugen) und die innere Falte reguliert den Wasserstrom in den Mantelraum zu den Kiemen.

Der Mantelrand ist bei den meisten Arten hell-dunkel-empfindlich. Bei schwimmenden Muscheln, wie Kamm- (Pecten) und Feilenmuscheln (Lima), die genauere Informationen über ihre Umgebung brauchen, ist der Mantelrand mit einfachen Ocellen (Punktaugen) besetzt. Bei der Gattung Arca können es über 200 Augen sein.
Ursprünglich verlaufen die Mantelränder getrennt und parallel zum Schalenrand (integripalliat). Abgeleitet sind bei den meisten Arten die aneinander liegenden Mantelränder mehr oder weniger miteinander verwachsen. Zum Teil bleiben nur drei Öffnungen bestehen: zwei Öffnungen am hinteren Ende der Muschel, durch die Atemwasser und Nahrung in den Mantelraum ein- und – durch die Kiemen gefiltert – wieder ausströmen, sowie einer Öffnung für den Fuß.
Der Mantelrand um die Atemöffnungen des Mantels sind bei grabenden oder bohrenden Muschelarten oft schlauchförmig verlängert, so dass die Muschel auch im Substrat mit Atemwasser und Nahrung versorgt ist. Die schlauchförmigen Mantelfortsätze bezeichnet man als Siphonen. Man unterscheidet einen zuführenden Sipho (Ingestionssipho) und einen ausführenden Sipho (Egestionssipho). Beide können zu einer einziehbaren Doppelröhre verwachsen sein, die im ausgestreckten Zustand länger sein kann als die Muschel selbst.
Damit dieser Sipho bei Gefahr in die Schale zurückgezogen werden kann, musste die Ansatzlinie der Mantelrandmuskeln nach innen ausweichen. Diese zurückgewichene Ansatzlinie ist in der Schale anhand einer mehr oder weniger ausgeprägten Mantelbucht der Mantellinie zu erkennen (sinupalliat).
Sandklaffmuscheln (Mya arenaria) beispielsweise leben eingegraben im Substrat des Wattenmeeres und versorgen sich durch den Sipho. Werden sie aus dem Substrat ausgespült, müssen sie sterben. Im Gegensatz dazu sitzt die Gemeine Miesmuschel (Mytilus edulis) angeheftet auf festen Untergrund und benötigt deshalb keinen Sipho. Sie stirbt daher, wenn sie von Substrat begraben wird.
Im Mantelrand der Riesenmuscheln (Tridacna) leben symbiotische Algen (Zooxanthellen), die von der Muschel geschützt werden. Im Gegenzug dafür kommt die Muschel in den Genuss der Photosyntheseprodukte der Algen.
Kiemen

Evolutionär haben sich aus einfachen Fiederkiemen zur Atmung die komplexeren Faden- und Blattkiemen zur Filtration entwickelt. Die im Mantelraum liegenden Kiemen der Muschel dienen dann nicht nur zur Atmung, sondern bei den meisten Arten außerdem zur Nahrungsaufnahme. Die Kiemen sind – wie die ganze Mantelhöhle – mit Wimpern besetzt, die einen Atemwasserstrom produzieren, der auch Nahrungspartikel einstrudeln kann. Die Kiemenbögen produzieren einen Schleim, in dem die Partikel hängenbleiben und auf Wimpernbändern zum Mund transportiert werden. Ungeeignete Partikel werden jedoch nicht verschluckt, sondern in Schleim zu größeren Scheinkotballen – Pseudofaeces – zusammengerollt und mit dem ausströmenden Wasser nach außen abgegeben.
Kiementypen:
- Die paarigen Fiederkiemen (Ctenidien) bestehen jeweils aus einem Schaft mit mehreren Kiemenblättchen.[1]
- Bei den Fadenkiemen (Filibranchien) hängen vor und hinter dem Fuß W-förmige Kiemenfäden in zwei Reihen in den Mantelraum. Die Kiemenfäden sind dorsal an der mittleren Spitze des W befestigt und die Schenkel des W untereinander durch kleine Wimpernstrukturen verbunden.
- Die netzartigen Schein-Blattkiemen (Pseudolamellibranchien) sind durch seitliche Verwachsung der Kiemenfäden, also Verbindungen zwischen aufeinanderfolgenden „Ws“ entstanden.
- Die echten Blattkiemen (Eulamellibranchien) sind durch echte, von Blutgefäßen durchzogene, Gewebebrücken zwischen den Kiemenfäden gekennzeichnet.
- Die Netzkiemen (Septibranchien) sind schmal, netzförmig und seitlich mit dem Mantel verwachsen. Diese Verwachsung ist muskulös. Bei einigen Formen sitzen die Kiemen in Lochreihen dieser Verwachsung die dann eine Scheidewand (Septum) ausbildet, selten sind die Kiemen ganz reduziert. Typischerweise sind die Netzkiemen-Arten Tiefseebewohner und saugen z. T. aktiv Nahrung ein.
Kreislaufsystem
Das Blutgefäßsystem der Muscheln ist, wie bei den meisten Weichtieren, offen. Das Herz hat zwei Vorkammern und eine Hauptkammer. Die Hauptkammer wird primär und sekundär vom Enddarm durchzogen.
Fuß
Der ursprünglich flach beilförmige Fuß der Muscheln kann, angepasst an Lebensweise und Fortbewegung, unterschiedliche Formen wie balkenförmig, zungenförmig oder wurmförmig annehmen. Schwimmende und festsitzende Muschelarten haben oft einen weitgehend zurückgebildeten Fuß. Der Fuß trägt die Byssusdrüsen, die vor allem bei Jungmuscheln Haftfäden (70 % Kollagen) produzieren. Bei einigen Arten produzieren auch die ausgewachsenen Tiere Byssusfäden, mit denen sich die Muscheln am Untergrund verankern (z. B. bei Miesmuscheln (Mytilidae), Archenmuscheln (Arcidae), Kammmuscheln (Pectinidae) und Steckmuscheln (Pinnidae)). Die Byssusverbindung kann später gelöst werden, indem die Muschel die Fäden sekretorisch abtrennt (Mytilus) oder ganz abstößt (Perlmuscheln (Pinctada)). Miesmuscheln nutzen ihre Byssusfäden auch zur Verteidigung, indem sie kleinere Schnecken, z. B. Reusenschnecken (Hinia) damit einspinnen.
Gehäuse
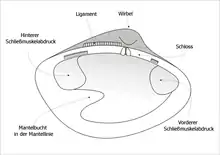
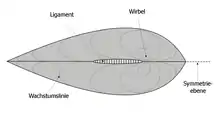

Das Gehäuse der Muscheln besteht aus einer rechten und einer linken Klappe und umgibt den Weichkörper der Muschel. Beide Klappen werden auf dem Rücken (also oben) mit einem Schloss und einem Schlossband (Ligament) zusammengehalten. Der Wirbel (Umbo) ist der älteste Teil des Muschelgehäuses. Farben und Formen sind sehr variabel. Das Gehäuse wird vom Mantel, einer Hautfalte der Muschel, gebildet und zwar in drei Schichten: der farbigen Schalenhaut (Periostracum), der Prismenschicht (Ostracum) und der inneren Kalkschicht (Hypostracum). Die beiden Hälften können durch zwei innere Schließmuskeln zusammengezogen werden.
Material
Die inneren Schalenschichten bestehen hauptsächlich aus Kalk, dessen Kristalle mittels einer organischen Substanz, dem Conchin, verkittet werden. Die Art der gebildeten Kalkstrukturen hat sich im Laufe der Evolution verändert: Ursprüngliches Merkmal ist die Bildung von perlmuttrigem Aragonit. Bei den heute dominierenden Heterodonta und bei den Taxodonta wird als abgeleitetes Merkmal ein kreuzlamellärer Aragonit gebildet. Parallel dazu entwickelten die Pteriida die Einlagerung von Calcit.
Calcit tritt auch abgeleitet in der mittleren Prismenschicht auf, und zwar bei den Anisomyaria und den ausgestorbenen Hippuritoida. Die Calcit- und Aragonitkristalle sind in der Prismenschicht in kleinen, eckigen Säulen (Prismen) angeordnet, die mehr oder weniger senkrecht nach außen zeigen.
Das äußere Periostracum besteht aus organischem Conchiolin und kann verschieden ausgeprägt sein. Am häufigsten wird eine dünne Haut ausgebildet, es kommen aber auch „Haare“ (z. B. Bärtige Archenmuschel) und lappige Vorhänge (z. B. Solemya togata) vor.
Ligament
Eigentlich ist die Schale nur im Bereich der beiden Kalkschichten (Ostracum und Hypostracum) in zwei Hälften geteilt. Die äußerste Schalenschicht, das Periostracum, überzieht die Schale auch hier, verstärkt sich zum Ligament und verbindet so rückenseitig die beiden Schalenhälften. Dieses Ligament zieht oft in den Scharnierbereich hinein und verdickt sich zum Resilinum (siehe Resilin und Abductin). Dieses Schlossband besteht aus nicht verkalkendem Conchin. Dieser Ligament-Resilinum-Komplex ist sehr elastisch und arbeitet antagonistisch zu den Schließmuskeln. Nach dem Tod des Tieres klaffen daher die beiden Schalen auseinander und werden durch mechanische Beanspruchung, wie etwa die Brandung, leicht getrennt, sodass man meist nur mehr einzelne Schalenhälften findet – im Gegensatz zur selteneren Dublette, die noch aus beiden Klappen besteht.
Schloss
Damit die beiden Hälften seitlich nicht verrutschen, tragen sie meist sogenannte Schlosszähne, deren Ausbildung auch Grundlage systematischer Zuordnungen ist. Diese Schlosszähne bestehen aus zahn- oder leistenförmigen Erhebungen am inneren Rückenrand der Klappen beziehungsweise entsprechenden Gruben auf der Gegenklappe, die ineinandergreifen. Auch ineinandergreifende Einkerbungen am unteren Schalenrand können ein seitliches Verrutschen verhindern. Diese Einkerbungen sind ebenfalls bestimmende Merkmale einer Muschelart.
Es werden bis zu neun verschiedene Schlosstypen unterschieden:
- taxodont (reihenzähnig): zahlreiche Zähne sind in einer (meist abnehmenden) Reihe am Rand der Muschel platziert (z. B. bei Archenmuscheln)
- heterodont (verschiedenzähnig): wenige große Zähne und max. vier Nebenzähne (z. B. bei Cerastoderma, Venusmuschel)
- desmodont (bandzähnig): Zwei Zähne zu einem verwachsen mit löffelförmigen Chondrophor, einem vorstehenden Träger des inneren Ligaments (z. B. bei Mya)
- pachyodont (dickzähnig): ein bis drei dicke Zapfen, die in tiefe Gruben der Gegenseite passen
- dysodont (schlecht bezahnt): ohne Zähne – nur kleine Erhebungen (z. B. bei Ostrea)
- schizodont (spaltzähnig): mittlerer Zahn der linken Klappe meist gespalten und rechts von zwei keilförmigen Zähnen umrahmt
- isodont (gleichzähnig): zwei bis vier Zähne symmetrisch auf beiden Klappen
- hemidapedont: schwach ausgebildete Hauptzähne, nur selten Nebenzähne
- anomalodesmatisch: ohne oder nur mit schwach ausgebildeten Zähnen, mit Chondrophor, einem vorstehenden Träger des inneren Ligaments (Schließknorpel oder Resilium)
Analog spricht man bei den Schalen der Articulata, die keine Muscheln sind, sondern zu den Armfüßern gehören, vom Schloss.
Muskelabdrücke
Die Ansatzstellen der Schließmuskeln (Schließmuskelabdrücke) und der Mantelrückziehmuskeln (die Mantellinie) spielen für Systematik und Bestimmung eine wichtige Rolle. Von den Schließmuskelabdrücken sind ursprünglich zwei gleich große vorhanden (isomyar). Im Lauf der Evolution entstanden Arten mit unterschiedlich großen Schließmuskelabdrücken (anisomyar oder heteromyar) und solche mit nur einem Abdruck (monomyar).
Die Mantellinie veränderte sich von einer einfachen gebogenen Linie, die die beiden Schließmuskelabdrücke verbindet, (integripalliat, vgl. das Foto einer Herzmuschel) hin zu einer eingebuchteten Form (sinupalliat, vgl. Zeichnung).
Formen

In Farbe, Form und Beschaffenheit sind die Schalen der einzelnen Arten sehr unterschiedlich. Weiße und stachlige existieren ebenso wie längliche, schwarze und glatte Schalen.
Die Schalenform der Muscheln ist an ihre jeweilige Lebensweise angepasst. Da Muscheln aus unterschiedlichen Gruppen oft ähnliche Lebensweisen besitzen, etwa im Sediment vergraben durch einen Sipho Wasser filtrieren, bilden sie trotz geringer Verwandtschaft oft ähnliche Schalenformen aus (Konvergenz). Der Zusammenhang zwischen Schalenform und Lebensweise ermöglicht auch die Rekonstruktion der Lebensweise fossiler Formen und eventuell auch deren Habitate.
Löcher
Am Strand angespülte Muschelschalen weisen manchmal kreisrunde, 1–3 mm große Löcher auf. Diese werden von Raubschnecken (meist Naticidae) erzeugt, welche die Muschel mit ihrer Raspelzunge aufbohren und dann die Weichteile verzehren.[2]
Lebensweise
Lebensräume

Muscheln sind an das Leben im Wasser gebunden. Sie kommen sowohl im Salzwasser, Brackwasser als auch Süßwasser vor, von der Arktis und Antarktis bis in die Tropen. Besonders im Wattenmeer findet man sehr große Mengen an Muscheln (beispielsweise Herzmuscheln und Miesmuscheln) im beziehungsweise auf dem Watt. Sie stellen eine wichtige Nahrung für Seevögel dar.
Die Art Enigmonia aenigmatica (HOLTEN, 1803) (eine Anomiidae) lebt in der Gezeitgischt auf Mangrovenblättern im Indopazifik und entspricht am ehesten einer Lebensweise an Land.
Muscheln leben als erwachsene Tiere überwiegend sessil (festsitzend), teils an festen Oberflächen, beispielsweise Felsen oder Steinen, teils im Sand oder Schlick. Sie gehören also zum Benthos. Sie sind von der Gezeitenzone bis in die Tiefsee verbreitet.
Fortbewegung und Verankerung
Folgende Fortbewegungsarten sind bei Muscheln beobachtet worden:
- Fortbewegung mit Hilfe des Fußes
- Die häufigste Fortbewegungsweise bei Muscheln ist die Fortbewegung mit Hilfe ihres Fußes. Dieser Körperteil kann zum Beispiel bei Herzmuscheln (Cardiidae) auf die dreifache Länge des Schalendurchmessers gestreckt werden. Dank ihres Fußes können sich Muscheln schnell eingraben, ruckweise kriechen oder sogar springen (Knotige Herzmuschel (Acanthocardia tuberculata) aus einem 20 cm hohen Aquarium). Für die Fortbewegung brauchen die Muscheln ein geeignetes Substrat, da sie im Gegensatz zu Schnecken nie einen echten Kriechfuß besitzen. Arten der Gattung Sphaerium sind in der Lage, sich auf Pflanzen spannerartig fortzubewegen, indem der Fuß ausgestreckt, die Spitze festgeklebt und der Körper nachgezogen wird.
- Durch Schalenklappern freies Schwimmen auf kurzen Strecken
- Arten der Gattung Lima schwimmen gut, einige der Familie Kammmuscheln (Pectinidae) ebenfalls. Sie können durch ruckartiges Zusammenklappen der Schalenhälften einen gerichteten Wasserstrom erzeugen und sich so nach dem Raketenprinzip ein Stückchen durch das Wasser bewegen. Kammmuscheln können angeblich Wasser auch gezielt durch die Öhrchen an den Seiten ausstoßen und so ihre Bewegungen, unterstützt durch ihre Linsenaugen, feiner koordinieren.
- Verankern mit Byssusfäden
- Einige Muscheln (unter anderem Miesmuscheln (Mytilus), Steckmuscheln (Pinnidae), Archenmuscheln (Arcidae) und Sattelmuschel (Anomia ephippium)) spinnen mit einem Sekret der Byssusdrüse an ihrem Fuß so genannte Byssusfäden, mit denen sie sich aneinander und an der Unterlage festkleben. Für Positionswechsel können zumindest Miesmuscheln die Byssusfäden mit einem Sekret wieder auflösen.
- Feste Verankerung mit der Schale am Untergrund
- Bekanntestes Beispiel sind die Austern (Ostreidae). Weitere festgewachsene Gattungen: Chama, Pseudochama, einige Anomia-Arten, Stachelaustern (Spondylus).
- Keine feste Verankerung, da von Substrat geschützt
- Bei Arten, die in Substrat wie Holz oder Stein bohren, kann die Bewegungsfähigkeit eingeschränkt und die Schale als Schutz reduziert sein. Beispiele: Schiffsbohrwürmer und Krause Bohrmuschel.
Ernährung
Die Muscheln waren ursprünglich Sedimentfresser, abgeleitet entstanden filtrierende Planktonfresser und Sonderformen wie Holzfresser, Jäger und Bakterienzüchter.
Die urtümlichen Muscheln aus der Unterklasse Protobranchia sammeln mit verlängerten Mundlappen vom umgebenden Substrat essbare Partikel, wie Protozoen, Eier, Larven und verdaulichen Detritus ein. Die Nahrung gelangt anschließend über eine Wimperrinne zur Mundöffnung.
Die meisten höher entwickelten Muscheln ernähren sich ausschließlich durch Filtration ihres Atemwassers. Im Mantelraum und auf den Kiemen gelegene Wimpern erzeugen einen gerichteten Wasserstrom, der durch eine Atemöffnung eintritt und durch die andere wieder austritt. Feste Teilchen im Atemwasser werden an den Kiemen aufgefangen und gelangen in einem Schleimpaket zur Mundöffnung. Die verwertbaren Partikel, meist Plankton, werden verdaut, der Rest in Kotschnüren ausgeschieden. Aufgrund ihrer Ernährung durch Filtration kommen Muscheln mit sehr großen Wassermengen in Kontakt, was sie besonders empfänglich für im Wasser enthaltene Schadstoffe macht und als Bioindikatoren prädestiniert.
Die Bohrmuscheln (Teredinidae) verwerten mit speziellen Enzymen das Holz, in dem sie bohren.
Einige, möglicherweise auch die meisten Arten der zu den Anomalodesmata gehörenden Septibranchia (darunter die Keulenmuscheln) sind Jäger, die mithilfe ihres muskulösen Septums in der Mantelhöhle aktiv kleine Krebstiere einsaugen.
Die Riesenmuscheln (Tridacna) und Arten der Gattung Solemya leben mit symbiotischen, Photosynthese betreibenden Algen im Mantelrand, und Tiefseearten an Black Smokern halten sich Sulfidbakterien in speziellen Strukturen ihrer Kiemen.
Fortpflanzung und Entwicklung
Die meisten Muschelarten sind getrennt geschlechtlich; es gibt sowohl männliche, als auch weibliche Tiere. Befruchtung und anschließende Larvenentwicklung finden äußerlich im Wasser statt. Der Ausstoß von Eiern und Samenzellen kann etwa bei den Riesenmuscheln (Tridacna) hormonell koordiniert sein. Nach einer Entwicklung über ein Larvenstadium vom Trochophora- oder Veliger-Typus entsteht aus der Larve nach einer Metamorphose die Jungmuschel, die sich während ihres Heranwachsens einen passenden Ort sucht, an dem sie das Erwachsenenleben verbringen kann. Bei Miesmuscheln (Mytilus), Austern (Ostrea) und anderen koloniebildenden Arten bleiben die Jungmuscheln meist in der Nähe der Kolonie und befestigen sich anschließend nicht nur am Untergrund, sondern auch an anderen Muscheln. So entstehen beispielsweise die Muschelbänke der Miesmuschel wie man sie aus dem Wattenmeer kennt.
Die im Süßwasser lebenden Muschelarten zeigen sehr unterschiedliche Fortpflanzungs- und Entwicklungsmethoden. Diese Muschelgruppen, ebenso wie die im Süßwasser und an Land lebenden Schneckengruppen, haben sich im Verlauf ihrer Entwicklung stark an die wechselhaften Lebensbedingungen, die das Süßwasser von den relativ konstanten Bedingungen im Meer unterscheiden, angepasst. Die Gruppe der Unionida, zu denen die in Mitteleuropa heimischen Großmuscheln (Maler-, Bach-, Teich- und Flussperlmuscheln) gehören, entwickelt sich über ein parasitisches Larvenstadium, die so genannten Glochidien, die sich zur weiteren Entwicklung erfolgreich an einem vorbeischwimmenden Fisch festheften müssen.
Im Gegensatz dazu sind die meisten im Süßwasser lebenden Kleinmuscheln (Erbsenmuscheln (Pisidium) und Kugelmuscheln (Sphaerium)) zwittrige Tiere, die lebende Larven gebären (Ovoviviparie). Die zu den Dreikantmuscheln gehörende Wandermuschel (Dreissena polymorpha) hingegen entwickelt sich, wie ihre meereslebenden Verwandten, über ein veligerähnliches planktontisches Larvenstadium.
Gefährdung
Durch Gewässerverschmutzung und Flussbegradigungen sind die großen Süßwassermuscheln (Unionida), zu denen auch die Flussperlmuschel (Margaritifera margaritifera) zählt, vielerorts stark in ihrem Bestand bedroht. Hinzu kommt die weitere Dezimierung durch die 1905 nach Eurasien eingeschleppte Bisamratte, die als Haupt-Fraßfeind dieser Muschelarten zählt.
Stammesgeschichte und Systematik
Evolution
Muscheln erscheinen fossil erstmals im Kambrium vor etwa 500 Millionen Jahren. Die frühen Formen waren noch einfach gebaute Weichtiere mit einer einklappigen Schale. Im mittleren Ordovizium erschienen erstmals Vertreter aller modernen Unterklassen.
Die rezenten Muscheln sind aus sedimentgrabenden Vorfahren entstanden. Diese hatten als Anpassung an das Leben im Meeresboden den Kopf bis auf die Mundöffnung und die Mundlappen reduziert. Jüngere Formen stellten auf eine Ernährung durch Filtration um. Evolutionär haben sich aus einfachen Fiederkiemen zur Atmung die komplexeren Faden- und Blattkiemen zur Filtration entwickelt.
Während der Kreidezeit bildeten schnell wachsende Muscheln riffähnliche Strukturen, die mit heutigen Korallenriffen vergleichbar sind. Diese Muscheln, deren Schalen Licht leitende Elemente enthielten, lebten teilweise in Symbiose mit Photosynthese betreibenden Einzellern.
In vielen Gesteinen zählen Muscheln zu den besonders häufigen Fossilien, da die harte Schale sich gut erhält. Sie dienen deswegen oft als Leitfossilien.
Die Evolutionsgeschwindigkeit der Muscheln ist sehr unterschiedlich. Während als Leitfossil dienende Arten im Durchschnitt 0,3 bis 1 Million Jahre existieren, sind einzelne Gattungen erheblich langlebiger. Die Gattung Gryphaea ist seit dem Unterjura (195 Mio. Jahre), die Gattung Spondylus seit dem Perm (285 Mio. Jahre) und Lima seit dem Jura nachgewiesen.
Das ursprüngliche, taxodonte Schloss der Muscheln besitzt eine große Zahl kleiner, gleichförmiger Zähnchen, während hoch abgeleitete, heterodonte Formen aus wenigen Hauptzähnen und bis zu vier leistenförmige Seitenzähnen bestehen oder gar keine Zähne besitzen.
Systematik der Großgruppen
Die Systematik der Muscheln war in der Vergangenheit größeren Veränderungen unterworfen. Auch heute kommen vor allem durch molekulargenetische Untersuchungen neue Daten hinzu, die selbst die Großsystematik noch geringfügig verändern könnten. Allerdings hat sich die Systematik und auch die Phylogenie der Großgruppen in den letzten 15 Jahren jedoch – mit wenigen Ausnahmen – weitgehend stabilisiert. Die Unterschiede in den verschiedenen Klassifikationen beruhen vor allem in der sehr subjektiven hierarchischen Stellung einiger weniger Gruppen (z. B. Ordnung Ostreida gegen Überfamilie Ostreoidea, Anomalodesmata Ordnung oder Überordnung).
Die Klassifikation der Muscheln nach Bieler & Mikkelsen (2006) (nur rezente Gruppen) und Amler et al. (2000) (auch fossile Gruppen) sowie nach dem Zoological Record stellt sich zurzeit so dar:
Klasse Bivalvia Linnaeus, 1758
- Infraklasse Protobranchi(at)a Pelseneer, 1889
- Ordnung Nuculida Dall, 1889
- Ordnung Solemyida Dall, 1889
- Ordnung Nuculanida Carter, Campbell & Campbell, 2000 (diese neue Ordnung hat noch keine allgemeine Akzeptanz gefunden (nach Zoological Record); alternativ können die in dieser Ordnung vereinigten Familien in die Ordnung Nuculida gestellt werden)
- Infraklasse Autolamellibranchi(at)a/Autobranchi(at)a Grobben, 1894
- Unterklasse Pteriomorphia Beurlen, 1944
- Ordnung † Praecardioida Newell, 1965
- Ordnung † Cyrtodontoida Scarlato & Starobogatov, 1971
- (Ordnung Ostreida Férussac, 1822) (diese Ordnung wird von Bieler & Mikkelsen (2006) nicht anerkannt, sondern zu einer Überfamilie innerhalb der Ordnung Pteriida „degradiert“. Demgegenüber verwendet die überwiegende Mehrzahl der Malakologen diese Großgruppe im Ordnungsrang; laut Zoological Record)
- Ordnung Arcida Stoliczka, 1870
- Ordnung Mytilida Férussac, 1822
- Ordnung Pteriida Newell, 1965
- Ordnung Limida Waller, 1978
- Ordnung Pectinida H. Adams & A. Adams, 1857
- Unterklasse Heteroconchia Hertwig, 1895
- Überordnung Palaeoheterodonta Newell, 1965
- Überordnung Heterodonta Neumayr, 1883
- Ordnung Carditida Dall, 1889
- Ordnung † Hippuritida Newell, 1965
- Ordnung Venerida H. Adams & A. Adams, 1856
- Ordnung Myida Stoliczka, 1870
- Ordnung Anomalodesmata Dall, 1889 (Diese Gruppe wird von den meisten Malakologen noch als Überordnung betrachtet; Harper et al. (2006) begründen die Rückstufung auf Ordnungsrang jedoch mit vielen neuen molekulargenetischen Daten).
- Unterklasse Pteriomorphia Beurlen, 1944
Für noch ältere Klassifikationen, die gelegentlich noch in populärwissenschaftlichen Muschelhandbüchern auftauchen, sei auf den Artikel Systematik der Muscheln verwiesen.
Stammbaum
Über die Phylogenie bzw. die Kladistik der heutigen Gruppen besteht insofern weitgehende Einigkeit unter den Malakologen, als die Protobranchi(at)a als die Schwestergruppe der restlichen Muscheln, den Autolamellibranchi(at)a (oder auch Autobranchi(at)a) angesehen werden. Einige halten die Protobranchia jedoch für eine paraphyletische Gruppierung. Innerhalb der Autolamellibranchia bilden Pteriomorphia und Heteroconchia wiederum Schwestergruppen. Die weitere Schwesterngruppengliederung innerhalb der Pteriomorphia ist noch unsicher. Innerhalb der Heteroconchia bilden Palaeoheterodonta und Heterodonta Schwestergruppen.
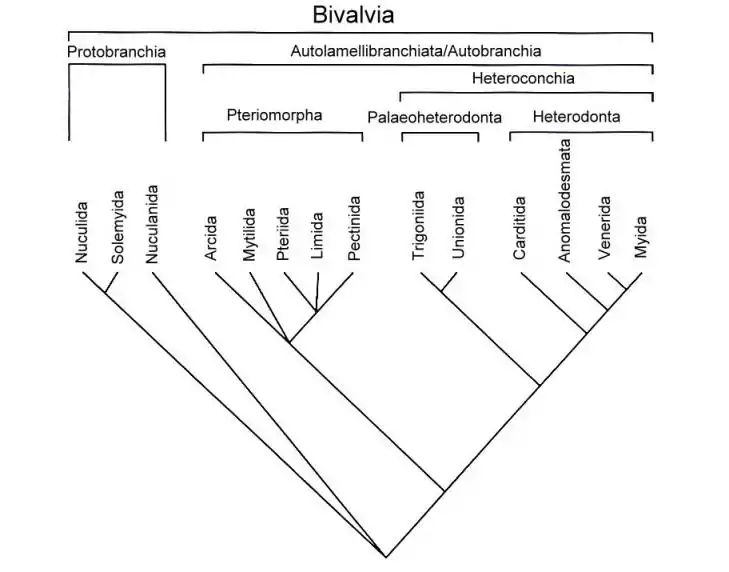
Zahl der Arten nach Großgruppen
Huber (2010) geht von weltweit 106 Familien mit insgesamt 1260 Gattungen und etwa 9200 rezenten Muschelarten aus. Diese verteilen sich wie folgt auf die jeweiligen Gruppen:
| Zahl der | Familien | Gattungen | Arten |
|---|---|---|---|
| Protobranchia | 10 | 49 | 700 |
| Nuculoidea | 1 | 8 | 170 |
| Sapretoidea | 1 | ca. 5 | 10 |
| Solemyoidea | 1 | 2 | 30 |
| Manzanelloidea | 1 | 2 | 20 |
| Nuculanoidea | 6 | 32 | 460 |
| Pteriomorpha | 25 | 240 (davon 2 Süßwasser) | 2000 (davon 11 Süßwasser) |
| Mytiloidea | 1 | 50 (1 Süßwasser) | 400 (5 Süßwasser) |
| Arcoidea | 7 | 60 (1 Süßwasser) | 570 (6 Süßwasser) |
| Pinnoidea | 1 | 3 (+) | 50 |
| Pterioidea | 5 | 9 | 80 |
| Ostreoidea | 2 | 23 | 80 |
| Dimyoidea | 1 | 3 | 15 |
| Anomioidea | 2 | 9 | 30 |
| Plicatuloidea | 1 | 1 | 20 |
| Pectinoidea | 4 | 68 | 500 |
| Limoidea | 1 | 8 | 250 |
| Palaeoheterodonta | 7 (davon 6 Süßwasser) | 171 (davon 170 Süßwasser) | 908 (davon 900 Süßwasser) |
| Trigonioidea | 1 | 1 | 8 |
| Unionoidea | (6 Süßwasser) | (170 Süßwasser) | (900 Süßwasser) |
| Heterodonta | 64 (davon 1 Süßwasser) | 800 (davon 16 Süßwasser) | 5600 (davon 270 Süßwasser) |
| Crassatelloidea | 5 | 65 | 420 |
| Thyasiroidea | 1 | ca. 12 | ca. 100 |
| Lucinoidea | 2 | ca. 85 | ca. 500 |
| Galeommatoidea | ca. 4 | ca. 100 | ca. 500 |
| Cyamioidea | 3 | 22 | 140 |
| Solenoidea | 2 | 17 (2 Süßwasser) | 130 (4 Süßwasser) |
| Hiatelloidea | 1 | 5 | 25 |
| Gastrochaenoidea | 1 | 7 | 30 |
| Chamoidea | 1 | 6 | 70 |
| Cardioidea | 2 | 38 | 260 |
| Tellinoidea | 5 | 110 (2 Süßwasser) | 900 (15 Süßwasser) |
| Glossoidea | 2 | 20 | 110 |
| Arcticoidea | 2 | 6 | 13 |
| Cyrenoidea | 1 | 6 (3 Süßwasser) | 60 (30 Süßwasser) |
| Sphaerioidea | (1 Süßwasser) | (5 Süßwasser) | (200 Süßwasser) |
| Veneroidea | 4 | 104 | 750 |
| Hemidonacoidea | 1 | 1 | 6 |
| Cyrenoidoidea | 1 | 1 | 6 |
| Ungulinoidea | 1 | 16 | 100 |
| Mactroidea | 4 | 46 | 220 |
| Dreissenoidea | 1 | 3 (2 Süßwasser) | 20 (12 Süßwasser) |
| Myoidea | 3 | 15 (1 Süßwasser) | 130 (1 Süßwasser) |
| Pholadoidea | 2 | 34 (1 Süßwasser) | 200 (3 Süßwasser) |
| Limoidea | 1 | 8 | 250 |
| (Anomalodesmata) | (14) | (71) | (770) |
| Pholadomyoidea | 2 | 3 | 20 |
| Clavagelloidea | 1 | 2 | 20 |
| Pandoroidea | 7 | 30 | 250 |
| Verticordioidea | 2 | 16 | 160 |
| Cuspidarioidea | 2 | 20 | 320 |
Muscheln und der Mensch
Muscheln als Nahrung


Menschen aus vielen historischen und prähistorischen Kulturen haben in ihren Küchenabfällen Hinweise darauf hinterlassen, dass Muscheln konsumiert wurden. Die ältesten Belege hierfür sind rund 165.000 Jahre alt und stammen von den so genannten Pinnacle-Point-Menschen aus Südafrika. In Dänemark werden solche urgeschichtlichen Muschelhaufen Køkkenmøddinger genannt.
Auch heute noch dienen viele Arten von Muscheln – in der westlichen Welt besonders Miesmuscheln, Austern, Venusmuscheln, Messermuscheln (wie Schwertförmige Scheidenmuschel) und Kammmuscheln – als Nahrung des Menschen. Allein in Europa werden jährlich rund 100.000 t Miesmuscheln verzehrt. Die Zucht von Muscheln auf Muschelbänken ist heute (besonders Austern und Miesmuscheln, die auch natürlich solche Bänke bilden) ein wichtiges Gewerbe an nahezu allen Küsten der Welt.
Die Angaben zum Nährwert unterscheiden sich zwischen Muschelarten nur geringfügig. Der Energiegehalt in kJ wurden nach den für Menschen gültigen Formeln berechnet. Wenn Muscheln als Nahrung für Tiere dienen, muss eventuell der Aufwand für die Verdauung der Muschelschale bedacht werden. Je nach Schalendicke kann daher die Energiebilanz negativ ausfallen.
| Energiegehalt und Nährstoffe pro 100 g in g (unfrittiert) (verzehrbarer Anteil für Frischware) | ||||
|---|---|---|---|---|
| Art | kJ | Protein | Fett | Kohlenhydrate |
| Jakobsmuschel | 336 | 11,9 | 1 | 5,5 |
| Klaffmuschel | 288 | 11,1 | 1,4 | 2,6 |
| Miesmuschel | 292 | 10,2 | 1,5 | 3,4 |
| Venusmuschel | 340 | 11,1 | 1,1 | 6,4 |
Perlen und Perlmutt
Neben der Nutzung als Nahrungsmittel ist die Verwendung von Muschelteilen, besonders Perlen und Perlmutt, als Schmuck von Bedeutung. Auch diese hat eine lange Tradition. Beispielsweise wurden im Neolithikum in Europa Muschelschalen über weite Strecken als Handelsware transportiert, wie Funde der Schalen der Stachelauster (Spondylus gaederopus) in der Bandkeramischen und Theiß-Kultur zeigen. Spondylus princeps, die an der Küste von Ecuador vorkommt, besaß eine große Bedeutung für die Menschen der präkolumbischen Zeit. Zu den Funden der asiatischen Mehrgarh-Kultur des frühesten Neolithikums gehört ebenfalls Schmuck aus Meeres-Muscheln.
Vertreter der Unionida und Pteriidae sind von jeher natürliche Quellen für Perlen und Perlmutt. Der hohe Wert von Perlen der Flussperlmuschel führte dazu, dass sie im Mittelalter in einigen Regionen der heutigen Bundesrepublik Deutschland unter Schutz gestellt und Wilderei massiv – etwa durch das Abhacken der Hand – bestraft wurde. Ab dem 5. Jahrhundert n. Chr. wurden in China so genannte „Buddha-Perlen“ in Süßwassermuscheln (wie z. B. Hyriopsis cumingii) gezüchtet, aber erst dem Japaner Kokichi Mikimoto gelang es in den frühen 1920er Jahren, die ersten vollrunden Zuchtperlen auf den Markt zu bringen und damit die Grundlage der heutigen Perlenzucht und des Handels mit Perlen zu legen.
Muschelwährungen
Viele Kulturen, etwa die Ureinwohner Nordamerikas, besaßen eine eigene Muschelwährung, da Muscheln wegen ihrer Seltenheit und ihres Schmuckwertes geschätzt wurden. So entstand der englische Begriff „shell out“ für „bezahlen“.
Schäden durch den Schiffsbohrwurm
Auch negative Auswirkungen von Muscheln auf den Menschen und seine Technik sind bekannt. Ein Beispiel für Bioerosion bietet der Schiffsbohrwurm (Teredo navalis), der sich vom Holz von Schiffen, Stegen, Buhnen oder hölzernen Deichtoren ernährt und durch seine langen Gänge das Holz brüchig macht. Seine Einschleppung aus Amerika führte zu einer Revolution des Schiffbaus, sein derzeitiges Vordringen in die Ostsee ist ein aktuelles Problem der Unterwasserarchäologie, da er dort historisch bedeutsame Holzschiffreste zerstört.
Symbole und Mythologie


Muscheln finden sich als Symbol in der Kunst und Mythologie, die wiederum die Benennung von Muscheln inspirierte. Zum Beispiel sind die Venusmuscheln (zu denen auch die Muschelgattung Venus gehört) benannt nach Venus, der antiken römischen Göttin der Liebe, des erotischen Verlangens und der Schönheit. Darstellungen zeigen oft die Geburt der Venus aus einer Muschel; allerdings wird dabei oft nicht eine Venusmuschel dargestellt, sondern eine Kammmuschel der Gattung Pecten (so auch im Bild rechts).
Seit dem Mittelalter dienen die stärker gewölbten rechten Schalenhälften der Großen Pilgermuschel Pecten maximus (bzw. Zinnabgüsse solcher Muscheln) den Jakobspilgern, die das Grab des heiligen Jakobus in Santiago de Compostela besuchen, als Erkennungszeichen. Die Mittelmeer-Pilgermuschel Pecten jacobaeus, die der Bezeichnung „Jakobsmuschel“ näher kommt, kann aufgrund ihres Verbreitungsgebietes nicht die von den Pilgern genutzte Art sein. Im Artikel Jakobsmuschel werden daher sowohl Pecten maximus als auch Pecten jacobaeus als Jakobsmuscheln bezeichnet.
Das Wappen von Guinea-Bissaus enthält eine stilisierte Kammmuschel als Symbol für die Lage des Landes an der Küste Afrikas. Der Ölkonzern Shell (englisch shell = Muschel) verwendet sie als Firmenlogo.
Das „Meeresrauschen“ in der Muschel
Es heißt, dass in Muscheln das Rauschen des Meeres zu hören sein soll. Man kann nämlich das Rauschen der Muschel mit einem Mikrofon aufnehmen, ohne dass sich ein Mensch in der Nähe befindet. Auf der Aufnahme wäre das Rauschen dann zu hören.
Tatsächlich handelt es sich hierbei um ein Resonanzphänomen: Wie in einem Blasinstrument befindet sich in der Schale eine Luftsäule, die eine bestimmte Eigenfrequenz besitzt. Das ist jener Ton, welcher entsteht, wenn man die Muschel zum Schwingen bringt. Die Luftsäule wird durch kaum merkliche Geräusche in der Umgebung, deren Frequenzen ungefähr der Eigenfrequenz der Muschel entsprechen, in Schwingung versetzt. Dadurch werden diese sonst nicht hörbaren Umgebungsgeräusche erheblich verstärkt. Das entstehende Tongemisch wird vom Menschen als Rauschen wahrgenommen.[3][4][5]
Siehe auch
- Muschelschale
- Muschelmuseum
- Køkkenmøddinger (prähistorische Muschelhaufen)
Literatur
- R. Tucker Abbott, S. Peter Dance: Compendium of Seashells. 1990, ISBN 0-915826-17-8
- Markus Huber: Compendium of Bivalves. A Full-Color Guide to 3'300 of the World's Marine Bivalves. A Status on Bivalvia after 250 Years of Research. 901 S. + CD. ConchBooks, Hackenheim 2010, ISBN 978-3-939767-28-2
- Rudolf Kilias: Marine Muscheln und Schnecken, Lexikon. 2. Auflage, Ulmer, Stuttgart 2000 (Erstausgabe 1997), ISBN 978-3-8001-3105-1
- Gert Lindner: Muscheln und Schnecken der Weltmeere, Aussehen, Vorkommen, Systematik [Bestimmungsbuch], 5., überarbeitete und erweiterte Auflage. BLV, München / Wien / Zürich 1999, ISBN 3-405-15438-3
- Gert Lindner: Muscheln und Schnecken sammeln und bestimmen: Die häufigsten und schönsten Arten. Für die Strände Europas [Bestimmungsbuch], In: Der zuverlässige Naturführer. 3. Auflage, BLV, München 2008, ISBN 978-3-8354-0374-1
- Arno Hermann Müller: Lehrbuch der Paläozoologie. Band. II, Teil 1, Protozoa – Mollusca 1, Fischer, Jena 1993, ISBN 3-334-60409-8
- Alfred Kaestner (Begründer): Lehrbuch der Speziellen Zoologie. Band I, Teil 3 Mollusca etc., Fischer, Stuttgart 1982, ISBN 3-437-20260-X
- Rüdiger Bieler, Paula M. Mikkelsen: Bivalvia – a look at the Branches. Zoological Journal of the Linnean Society, 148: 223–235, London 2006.
- Michael Amler, Rudolf Fischer, Nicole Rogalla: Muscheln. Haeckel-Bücherei, Band 5. Enke Verlag, Stuttgart 2000, ISBN 3-13-118391-8
- Elisabeth M. Harper, Hermann Dreyer, Gerhard Steiner: Reconstructing the Anomalodesmata (Mollusca: Bivalvia): morphology and molecules. Zoological Journal of the Linnean Society, 148(3): 395–420, London 2006
- D. C. Campbell: Paleontological and molecular evidence on Bivalve phylogeny. In: Evolutionary Paleobiology and Paleoecology of the Bivalvia 2002
- J. A. Schneider: Bivalve systematics in the 20th century. Journal of Paleontology, 75: 1119–1127 2001.
- Gonzalo Giribet, Daniel L. Distel: Bivalve phylogeny and molecular data. In: Lydeard, Charles; Lindberg, David R. [Eds]. Molecular systematics and phylogeography of mollusks. pp.45–90. [Smithsonian Series in Comparative Evolutionary Biology.]. Smithsonian Books, Washington and London 2003.
- Robert Prezant: Classification. In: P.L. Beesley, G. J. B. Ross & A. Wells (Hrsg.): Mollusca. The Southern Synthesis, Part A Fauna of Australia, vol.5, pp. 224–226, Collingwood, CSIRO Publishing, 1998
- J. G. Carter, D. C. Campbell, M. R. Campbell: Cladistic perspectives of early bivalve evolution. In: Geological Society Special Publications, pp. 47–79, Geological Society, London 2000.
- Norman D. Newell: Classification of Bivalvia. In: R.C. Moore (Hrsg.): Treatise on Invertebrate Paleontology, Part N, Mollusca 6, pp. N205-N218, University of Kansas Press, Lawrence, Kansas 1969.
- Literarische Darstellung
- Birgit Vanderbeke: Das Muschelessen [Erzählung]. Fischer-Taschenbuch 13783, Frankfurt am Main 1997 (Erstausgabe: Rotbuch, Berlin 1990, ISBN 3-88022-757-8), ISBN 978-3-596-13783-1 (Ingeborg Bachmann-Preis 1990).
Weblinks
- Muscheln auf weichtiere.at
- http://www.bivalvia.net
- www.wissenschaft.de: Warum das Verschwinden der Haie die Muscheln bedroht – Forscher zeigen den Zusammenhang der Abnahme von Hai- und Muschelbeständen auf
Einzelnachweise
- Pia Andrea Egger: Anatomisch-histologische Untersuchung der Kiemenretraktoren bei ausgewählten Arten der Caudofoveata (Mollusca). (PDF) Diss. Universität Wien, 2008.
- Wolfgang von Buddenbrook: Vergleichende Physiologie. Band III: Ernährung, Wasserhaushalt und Mineralhaushalt der Tiere. Springer Basel 1956, S. 262
- 121 – Akustische Hohlraumschwingungen. (Nicht mehr online verfügbar.) Friedrich-Schiller-Universität Jena, Physikalisch-Astronomische Fakultät, 13. August 2010, archiviert vom Original am 13. Mai 2012; abgerufen am 19. Oktober 2011 (Versuch mit einer leeren Flasche).
- Vladimir Rydl: Wissensfrage. Meeresrauschen oder Ohrensausen – was hört man, wenn man sich das Gehäuse einer Meeresschnecke ans Ohr hält? In: Planet Wissen. 1. Juni 2009, abgerufen am 19. Oktober 2011.
- Was rauscht in der Muschel? (Nicht mehr online verfügbar.) Schweizer Fernsehen, 19. April 2007, ehemals im Original; abgerufen am 19. Oktober 2011. (Seite nicht mehr abrufbar, Suche in Webarchiven)