Titanohyrax
Titanohyrax ist eine ausgestorbene Gattung aus der Ordnung der Schliefer. Innerhalb der Schliefer gehört sie zu einer frühen Entwicklungslinie. Die Tiere lebten vom Mittleren Eozän bis zum Unteren Oligozän, was den Zeitraum von vor 48 bis vor 28 Millionen Jahren umfasst. Das Fundmaterial besteht weitgehend aus fragmentierten Teilen des Gebisses und des Schädels und stammt von verschiedenen Fundstellen des nördlichen Afrikas. Herausragend ist dabei die Fundstelle des Fayyum in Ägypten. Diese barg das bisher umfangreichste und forschungsgeschichtlich älteste Material, es diente auch zur Aufstellung der Gattung im Jahr 1921. Titanohyrax schließt mehrere Arten ein. Bei diesen handelt es sich um mittelgroße bis sehr große Vertreter der Schliefer. Die größten Formen erreichten die Ausmaße kleiner Nashörner und repräsentieren die größten Schliefer ihrer Zeit. Es lassen sich aber einzelne Unterschiede in den Ausmaßen und in der Anatomie innerhalb der Arten feststellen, was möglicherweise in einem Geschlechtsdimorphismus begründet ist.
| Titanohyrax | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Mahlzähne von Titanohyrax | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Mittleres Eozän bis Unteres Oligozän | ||||||||||||
| 48 bis 28 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Titanohyrax | ||||||||||||
| Matsumoto, 1921 | ||||||||||||
Merkmale
Titanohyrax ist ein sehr großer Vertreter der Schliefer. Die größten Individuen wogen schätzungsweise rund eine Tonne, wodurch sie zu den größten bekannten Schlieferformen zählen. Die Größenvariation innerhalb der Gattung ist allerdings beträchtlich.[1] Das Fundmaterial, das der Gattung zugrunde liegt, besteht überwiegend aus fragmentierten Gebiss- und Schädelteilen. Aufgefundene postcraniale Reste wurden bisher noch nicht wissenschaftlich aufgearbeitet. Den diagnostizierbaren Schädelmerkmale an den vorhandenen Bruchstücken zufolge besaß Titanohyrax ein kurzes Rostrum, was mit anderen Titanohyraciden übereinstimmt, aber von den übrigen frühen Schliefern mit ihren eher langen Schnauzen abweicht. Die Symphyse verband wie bei allen Schliefern die beiden Unterkieferhälften fest miteinander. Sie war sehr kräftig und reichte nach hinten bis zum zweiten Prämolaren. Vorn war die Unterkante steil nach oben orientiert. Ein Foramen mentale öffnete sich unterhalb des zweiten Prämolaren, ein weiteres auf Höhe des letzten Prämolaren oder des ersten Molaren. Bei einigen Unterkiefern befand sich unterhalb des hinteren Molaren eine im Knochen eingebettete luftgefüllte Kammer. Der aufsteigende Ast erhob sich fast senkrecht über den horizontalen Knochenkörper. Das Merkmal stimmt mit zahlreichen anderen frühen Schliefern überein, ist aber abweichend von den heutigen Schliefern mit einem schräg nach hinten geneigten aufsteigenden Ast. Der Kronen- und der Gelenkfortsatz standen eng beieinander.[2][3]
Das Gebiss ist nicht vollständig bekannt, lediglich für die hintere Bezahnung liegt umfangreicheres Material vor. Am Unterkiefer waren aber wenigstens die beiden inneren Schneidezähne ausgebildet. Diese hatten eine nahezu gleiche Größe und waren von spatelartiger Gestalt, welche die typische Aufteilung in kleine, kammartig nebeneinanderliegende Spitzen anderer Schliefer vermissen lässt. Das Diastema zum hinteren Gebissabschnitt war in der oberen Zahnreihe sehr kurz, in der unteren kam möglicherweise keins vor. Die Prämolaren ähnelten weitgehend den Molaren und können damit als molarisiert angesprochen werden. Auf den oberen Molaren bestanden vier Haupthöcker, die den Zähnen einen bilophodonten Aufbau verliehen. Die beiden äußeren, lippenseitigen Höcker (Paraconus und Metaconus) verband eine W-förmig gestaltete Scherleiste, das Ectoloph; es gab den Mahlzähnen einen selenodonten Charakter. Auf der Zungenseite war der Protoconus abweichend von Rupestrohyrax aufgebläht, der Hypoconus wies seitliche Verschmälerungen auf und wirkte klein. Im Unterschied dazu besaß Megalohyrax zwei etwa gleichgroße zungenseitige Haupthöcker. Auch die unteren Molaren wurden von vier Haupthöckern dominiert (Paraconid, Metaconid, Hypoconid, Protoconid). Zusätzlich bestanden einzelne Nebenhöcker wie etwa das Metastylid, das beispielsweise bei anderen Vertretern der Titanohyracidae und bei Megalohyrax fehlt. Die Zahnkronen waren relativ hoch (hypsodont), das Merkmal variiert aber deutlich zwischen den einzelnen Arten von Titanohyrax.[4][5][3]
Fossilfunde
Titanohyrax trat erstmals im Übergang vom Unteren zum Mittleren Eozän vor etwa 48 Millionen Jahren in Erscheinung. Aus Chambi in Tunesien liegen ein Oberkieferfragment und einzelne isolierte obere und untere Backenzähne von T. tantulus vor, die noch auf ein eher kleines Tier verweisen. Die Zähne besaßen relativ niedrige Zahnkronen.[4] Als nahezu zeitgleich kann T. mongereaui aufgefasst werden. Bekannt ist die Art über einen schlecht erhaltenen mittleren oberen Mahlzahn aus Gour Lazib in Algerien, der aber schon deutlich größer war als entsprechende Funde aus Chambi.[6] Möglicherweise sind in Gour Lazib noch Reste weiterer Vertreter von Titanohyrax präsent. Gour Lazib bildet einen Fundstellenkomplex bestehend aus über einem Dutzend Einzelfundstellen. Die meisten Funde stammen aus der Glib-Zegdou-Formation, die sich weitgehend aus feinkörnigen Sedimentgesteinen zusammensetzt.[3] Nach diesem frühen Auftreten folgt eine zeitlich lange Lücke ohne Fossilnachweis der Gattung, die das gesamte Mittlere Eozän überspannt.[7][8]
Erst im Oberen Eozän und im darauffolgenden Unteren Oligozän ist Titanohyrax wieder fassbar. Das umfangreichste Fossilmaterial wurde bisher an der bedeutenden Lagerstätte des Fayyum im nördlichen Ägypten geborgen. Das Hauptfundgebiet bildet die Gebel-Qatrani-Formation mit ihren fein- bis gröberklastischen Sedimenten limnischen und fluviatilen Ursprungs. Die Reste von Titanohyrax lagerten in verschiedenen stratigraphischen Positionen, deren Alter nach paläomagnetischen Untersuchungen zwischen 33 und 28,5 Millionen Jahren beträgt. Sie lassen sich mehreren Arten zuordnen und geben somit Hinweise auf eine Radiation der Gattung in dieser Zeit, deren Ursprünge aber bislang unbekannt sind. Ein generelles Kennzeichen der Arten besteht in den im Vergleich zu den frühen Formen zumeist höheren Zahnkronen der Backenzähne.[8] Die ältesten Funde hier, repräsentiert durch verschiedene Zahnreste und zahlreiche postcraniale Skelettelemente, verweisen auf einen mittelgroßen und wohl eher grazilen Vertreter, eine formale Beschreibung blieb aber bisher aus. Sie kamen an der Fundstelle L-41 im unteren Abschnitt der Gebel-Qatrani-Formation zu Tage und gehören damit noch dem ausgehenden Obereozän an.[5] Der obere Abschnitt der Gesteinseinheit mit einer unteroligozänen Zeitstellung barg ebenfalls verschiedene Reste. Dazu zählt unter anderem T. angustidens aus den Fundstellen R, V und I. Diese kleinere bis mittelgroße Form ist durch mehrere Unter- und Oberkiefer belegt. Als besonderes Kennzeichen der Art waren die Backenzähne deutlich schmal ausgebildet.[2] T. andrewsi wiederum basiert auf relativ zahlreichem Material, zusammengesetzt aus Unterkiefern und einzelnen Oberkieferfragmenten.[3] Den größten Vertreter stellt T. ultimus dar. Er kann aber bisher nur einige wenige, stark abgekaute obere und untere Zähne und ein Oberkieferfragment vorweisen. Die Zahnkronen der Molaren sind etwas niedriger als die der anderen späteren Arten. Sowohl für T. andrewsi als auch für T. ultimus liegen nur wenige Informationen zu den exakten Fundlagen vor. Grund dafür ist das forschungsgeschichtlich frühe Auffinden der Reste, für die zumeist keine Lagedokumentation erfolgte.[9][10] Zwei Funde von T. ultimus fanden sich bei späteren Grabungen an der Fundstelle I im obersten Abschnitt der Gebel-Qatrani-Formation.[11][5][3]
Aus der späteren Radiationsphase von Titanohyrax wurden noch ein Ober- und ein Unterkiefer aus dem Guerran Member der Samlat-Formation im südwestlichen Marokko berichtet, welche dem Oberen Eozän angehört. Die Funde zeigen eine Mischung aus ursprünglichen und modernen Merkmalen und könnten auf den Beginn der Radiationsphase hinweisen. Sie geben darüber hinaus Aufschluss darüber, dass die Gattung während dieser Zeit nicht nur im Fayyum, sondern im gesamten Nordafrika anwesend war. Eine formale Artbeschreibung fand noch nicht statt.[8]
Paläobiologie
Die Vertreter der Gattung Titanohyrax zeigen eine starke Variabilität in der Körpergröße, die sich im Fundmaterial über die Größe der Zähne ausdrückt, als Basis für Gewichtsberechnungen wurde der zweite obere Mahlzahn genommen. Frühe Formen wie T. tantulus aus dem Mittleren Eozän von Chambi waren teilweise noch sehr klein, dessen zweiter Molar maß gerade einmal etwa 1,1 cm in der Länge.[4] Allerdings wies das etwa gleichalte T. mongereaui aus Gour Lazib mit einem 3,6 cm langen mittleren Mahlzahn schon eine deutlich größere Dimension auf.[6] Gewichtsschätzungen reichen bis zu 626 kg. Das unteroligozäne T. angustidens aus dem Fayyum in Ägypten wog dagegen nur rund 180 kg, was anhand des etwa 2,3 cm langen zweiten Molaren errechnet wurde.[2] Dagegen betrug die Länge des mittleren Mahlzahns von T. andrewsi aus der gleichen Region 3,3 cm, was in einem Körpergewicht von schätzungsweise 430 kg resultiert.[3] Alle diese Formen wurden von T. ultimus ebenfalls aus dem Fayyum übertroffen. Sein zweiter Molar wurde 4,1 cm lang,[9] die Tiere brachten wahrscheinlich ein Körpergewicht von 994 kg auf die Waage. Damit stellt Titanohyrax den größten bekannten Schliefer des Paläogens in Afrika dar. Erst in späterer Zeit erreichten einige Angehörige einer anderen Entwicklungslinie der Schliefer mit etwa 1000 bis 1300 kg wieder ähnliche Ausmaße, so etwa Sogdohyrax und Postschizotherium aus dem zentralen und östlichen Asien. Beide datieren in das Pliozän. Zum Vergleich dazu werden die heutigen Arten der Schliefer nur 1,5 bis 5,5 kg schwer bei einer entsprechenden Zahngröße des oberen zweiten Molaren von 0,7 bis 0,8 cm für sehr große Individuen.[12] Titanohyrax kann damit als Mitglied der Megafauna im nördlichen Afrika des Paläogens angesehen werden. Es teilte seinen Lebensraum mit anderen riesigen Säugetieren seiner Zeit wie etwa Arsinoitherium aus der Gruppe der Embrithopoda oder Barytherium und Palaeomastodon aus der Gruppe der Rüsseltiere.[1][5][3]
Neben den allgemeinen Größenunterschieden zwischen den verschiedenen Arten von Titanohyrax lassen sich aber auch Differenzen innerhalb der einzelnen Arten erkennen. Dies betrifft unter anderem T. andrewsi, von dem unterschiedlich massiv ausgebildete Unterkiefer vorliegen. Einige Unterkiefer weisen einen robusten horizontalen Knochenkörper mit einer Höhe unter dem zweiten Molaren von 8 cm auf, während er bei andern zwischen 3,4 und 4,2 cm variiert. Auch zeigen sich andere Abweichungen wie etwa ein stärker ausgebildeter Winkelfortsatz an den größeren Unterkiefern. Generell werden bei Schliefern größere und massivere Unterkiefer einer Art mit männlichen und kleinere mit weiblichen Tieren in Verbindung gebracht. Für T. andrewsi würde dies bedeuten, dass männliche Individuen rund 430 kg, weibliche dagegen rund 240 kg gewogen haben. Daraus ergibt sich ein Größenverhältnis der Geschlechter von 1,79:1, was wesentlich stärker als bei heutigen Schliefern ausfällt. Bei diesen treten zwar auch markante Geschlechtsunterschiede in verschiedenen Schädelmerkmalen auf, so in der Robustizität des Unterkiefers und der Ausprägung der oberen Schneidezähne, diese ergeben aber keine großen Abweichungen in der Gesamtkörpergröße. Das Verhältnis bei heutigen Schliefern beträgt daher 1,1:1 im Vergleich Männchen zu Weibchen. Der Unterschied bei T. andrewsi übertrifft auch den Geschlechtsdimorphismus zahlreicher Huftiere, befindet sich aber in der Variationsbreite der heutigen Elefanten.[3]
Weder T. tantulus noch T. mongereaui beziehungsweise T. ultimus können momentan Unterkiefer vorweisen. Von T. angustidens sind einige bekannt, die bisher kaum Größenvariationen zeigen. Dafür befindet sich im horizontalen Knochenkörper ein interner runder, mit Luft gefüllter Hohlraum unterhalb der beiden hinteren Molaren. Derartige Hohlräume treten auch bei anderen frühen Schliefern auf, beispielsweise bei Thyrohyrax oder bei Saghatherium und kommen bei diesen in manchen Arten exklusiv bei männlichen Tieren, in anderen in beiden Geschlechtern mit unterschiedlicher Ausprägung vor. Ihre Funktion ist unbekannt, teilweise werden sie aber mit der Lautgebung in Verbindung gebracht.[2][5] Heutige Schliefer verfügen nicht über ein derartiges Merkmal. Von T. angustidens liegen zu wenige Unterkiefer vor, um dieses als geschlechtsunterscheidendes Kriterium zu definieren. Zudem repräsentieren sie alle nicht vollständig ausgewachsene Individuen, so dass über die endgültige Form der Struktur keine Aussagen möglich sind. Es kann aber zusammengefasst werden, dass sich innerhalb der frühen Schliefer verschiedene und weitgehend unabhängige Formen des Geschlechtsunterschieds finden: einerseits die Ausprägung von internen luftgefüllten Kammern im Unterkiefer, andererseits durch Variationen in der Massivität des Unterkiefers. Als Erklärung für dieses Phänomen wird ein abweichendes Paarungsverhalten der einzelnen Arten vermutet.[13][14][3]
Systematik
Innere Systematik der frühen Schliefer nach Pickford 2015[15]
|
Titanohyrax ist eine Gattung aus der ausgestorbenen Familie der Titanohyracidae innerhalb der Ordnung der Schliefer (Hyracoidea). Die Schliefer bilden gemeinsam mit den Elefanten und Seekühen eine Verwandtschaftsgruppe, die als Paenungulata bezeichnet wird und innerhalb der Überordnung der Afrotheria steht. Die Titanohyracidae wiederum gehören zu den frühen Formen der Schliefer. Sie sind erstmals im Eozän im nördlichen Afrika nachweisbar. Gemeinsam mit den Namahyracidae, den Geniohyidae und den Saghatheriidae gehören sie zur ersten Radiationsphase der Schliefer, welche im Paläogen erfolgte und sich auf Afrika beschränkte. In dieser Zeit formten sich die Schliefer als eine sehr vielfältige Gruppe mit zahlreichen Vertretern heraus. Diese waren an die unterschiedlichsten Biotope angepasst. Die Titanohyracidae vereinen dabei mittelgroße bis sehr große Vertreter, einige Formen wie Titanohyrax erreichten die Ausmaße kleiner Nashörner und gehören damit zu den größten bekannten Schliefern überhaupt. Charakteristische Merkmale der Titanohyracidae stellen der gegenüber anderen frühen Schliefern gekürzte Schnauzenteil des Schädels, das typisch selenodonte Kauflächenmuster der Backenzähne und die stark spezialisierten Unterkieferschneidezähne dar. Im postcranialen Skelett waren die Titanohyracidae mitunter an eine schnelle Fortbewegung angepasst. Teilweise werden die Titanohyracidae zusammen mit den Geniohyidae und den Namahyracidae in die Unterordnung der Pseudhippomorpha verwiesen, welche die urtümlichen Schliefer zusammenfasst. Sie steht als Schwestergruppe den moderneren Procaviamorpha mit den Saghatheriidae, den Pliohyracidae und den Procaviidae gegenüber, letztere enthalten die heutigen Schliefer.[16][17][5]
Innerhalb der Gattung Titanohyrax werden mehrere Arten anerkannt:[5]
- T. andrewsi Matsumoto, 1921
- T. angustidens Rasmussen & Simons, 1988
- T. mongereaui Sudre, 1979
- T. tantulus Court & Hartenberger, 1992
- T. ultimus Matsumoto, 1921
Die einzelnen Arten zeigen eine drastische Variabilität in der Körpergröße. Weitere Unterschiede betreffen unter anderem die Kronenhöhe der Backenzähne, die bei frühen Gattungsvertretern generell niedriger ist als bei späteren. Zudem tritt an den oberen Molaren einiger früher Arten an der Scherleiste zwischen dem Proto- und dem Paraconus eine buckelartige Aufwölbung auf, die sogenannte „paraconulare Schwellung“. Neben den hier genannten Arten wurden mit T. palaeotherioides und T. schlosseri weitere eingeführt,[9][10] später aber zumeist mit T. andrewsi vereint. Eine bisher unbeschriebene kleinere Art wird für das Obere Eozän des Fayyum vermutet,[5] ebenso wie für die etwa gleichalte Samlat-Formation im südwestlichen Marokko.[8]
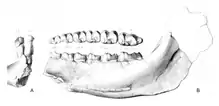
Die ersten Funde von Titanohyrax kamen bereits in der Wende vom 19. zum 20. Jahrhundert im Fossilgebiet des Fayyum zum Vorschein, wurden aber als solche nicht erkannt. Bereits 1904 hatte Charles William Andrews eine neue Art der Gattung Megalohyrax mit der Bezeichnung Megalohyrax minor eingeführt und ihr ein Oberkieferfragment zugewiesen. Einen teilweise erhaltenen Unterkiefer sah er als wahrscheinlichen Fossilrest der Art an.[18] In seinem umfassenden Katalogwerk A descriptive catalogue of the Tertiary Vertebrata of the Fayum, Egypt zur Fauna des Fayyum bildete Andrews die Funde ab.[19] Max Schlosser legte in den Jahren 1910 und 1911 eine Aufarbeitung der Fayyum-Fossilien mehrerer deutscher Expeditionen in das Fundgebiet vor. Hierin folgte er Andrews' Auffassung nur bedingt. Er teilte die Funde auf und behielt den Unterkiefer in der Gattung Megalohyrax, für den Oberkiefer mit einer nach seiner Auffassung einfacheren Zahngestaltung schuf er die neue Form Mixohyrax.[20][21] Eine Dekade später während der Bearbeitung der fossilen Schliefer aus amerikanischen und britischen Sammlungen revidierte Hikoshichiro Matsumoto die Gattung Megalohyrax. Er kam zu der Auffassung, das Schlossers Aufteilung des Fundmaterials zwar korrekt, seine daraus geschlussfolgerten Zuweisungen jedoch falsch waren. Demnach entsprach Schlossers Mixohyrax dem von Andrews etablierten Megalohyrax, Schlossers Megalohyrax repräsentierte eine eigenständige Gattung. Matsumoto kreierte für diese die Bezeichnung Titanohyrax und legte damit die wissenschaftliche Erstbeschreibung der Gattung vor. Der Name spielt auf die gewaltige Größe der Tiere an (nach den Titanen der griechischen Mythologie). Gleichzeitig erkannte er mit T. andrewsi, T. palaeotherioides, T. schlosseri und T. ultimus vier Arten an.[9][10] Im Jahr 1978 synonymisierte dann Grant E. Meyer T. palaeotherioides und T. schlosseri aufgrund nicht vorhandener morphologischer und metrischer Unterschiede mit T. andrewsi. Er verlegte dabei auch die Typusart von T. palaeotherioides auf T. andrewsi, da von ersterer neben nomenklatorischen Problemen auch das Ursprungsmaterial verloren gegangen war.[11] Andere Autoren sahen das problematischer. So erkannten D. Tab Rasmussen und Elwyn L. Simons 1988 T. palaeotherioides als durchaus unterschiedlich zu T. andrewsi an. Sie akzeptierten aber Meyers Argumentation zur Nomenklatur und führten die Art unter der neuen Bezeichnung T. angustidens wieder ein.[2] im gleichen Zeitraum wurden weitere Arten beschrieben.[6][4]
Literatur
- Hikoshichiro Matsumoto: Megalohyrax Andrews and Titanohyrax n. g. – A revision of the genera of Hyracoids from the Fayûm, Egypt. Proceedings of the Zoological Society of London, 1921, S. 839–850 ()
- Rodolphe Tabuce: A mandible of the hyracoid mammal Titanohyrax andrewsi in the collections of the Muséum National d'Histoire Naturelle, Paris (France) with a reassessment of the species. Palaeovertebrata 40 (1), 2016, S. e4 doi: 10.18563/pv.40.1.e4
Einzelnachweise
- Gary T. Schwartz, D. Tab Rasmussen und Richard J. Smith: Body-Size Diversity and Community Structure of Fossil Hyracoids. Journal of Mammalogy 76 (4), 1995, S. 1088–1099
- D. Tab Rasmussen und Elwyn L. Simons: New Oligocene Hyracoids from Egypt. Journal of Vertebrate Paleontology 8 (1), 1988, S. 67–83
- Rodolphe Tabuce: A mandible of the hyracoid mammal Titanohyrax andrewsi in the collections of the Muséum National d'Histoire Naturelle, Paris (France) with a reassessment of the species. Palaeovertebrata 40 (1), 2016, S. e4 doi: 10.18563/pv.40.1.e4
- Nicholas Court und Jean-Louis Hartenberger: A new species of the hyracoid mammal Titanohyrax from the Eocene of Tunisia. Palaeontology 35 (2), 1992, S. 309–317
- D. Tab Rasmussen und Mercedes Gutiérrez: Hyracoidea. In: Lars Werdelin und William Joseph Sanders (Hrsg.): Cenozoic Mammals of Africa. University of California Press, Berkeley, Los Angeles, London, 2010, S. 123–145
- J. Sudre: Nouveaux mammifères éocènes du Sahara occidental. Palaeovertebrata 9, 1979, S. 83–115
- Rodolphe Tabuce, Anne-Lise Charruault, Mohammed Adaci, Mustapha Bensalah, Mustapha Ben Haj Ali, El Mabrouk Essid, Laurent Marivaux, Monique Vianey Liaud und M’hammed Mahboubi: The early Eocene radiation of Hyracoidea (Mammalia, Afrotheria): New fieldwork evidence from northwestern Africa. In: Thomas Lehmann und Stephan F. K. Schaal (Hrsg.): The world at the time of Messel. Puzzles in Palaeobiology, Palaeoenvironment and the History of Early Primates. 22nd International Senckenberg Conference Frankfurt am Main, 15th - 19th November 2011, S. 161–162
- R. Tabuce, S. Adnet, M. Benammi, L. Marivaux, O. Saddiqi, L. Baidder und M. Benammi: A New Titanohyrax species from the Priabonian of Dakhla, Morocco bridges a gap in the fossil record of the ‘Giant’ Hyracoids (Mammalia). The First West African Craton and Margins International Workshop “WACMA1”, Dakhla, Morocco, 24 to 29th April 2017
- Hikoshichiro Matsumoto: Megalohyrax Andrews and Titanohyrax n. g. – A revision of the genera of Hyracoids from the Fayûm, Egypt. Proceedings of the Zoological Society of London, 1921, S. 839–850 ()
- Hikoshichiro Matsumoto: Contribution to the knowledge of the fossil Hyracoidea of the Fayum, Egypt, with description of several new species. Bulletin of the American Museum of Natural History 56, 1926, S. 253–350
- Grant E. Meyer: Hyracoidea. In: Vincent J. Maglio und H. B. S. Cooke (Hrsg.): Evolution of African Mammals. Harvard University Press, 1978, S. 284–314
- Martin Pickford: Fossil hyraxes (Hyracoidea: Mammalia) from the Late Miocene and Plio-Pleistocene of Africa, and the phylogeny of the Procaviidae. Palaeontologia africana 41, 2005, S. 141–161
- D. Tab Rasmussen und E. L. Simons: The oldest hyracoids (Mammalia: Pliohyracidae): new species of Saghatherium and Thyrohyrax from the Fayum. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 182, 1991, S. 187–209
- Donald D. de Blieux, Michael R. Baumrind, Elwyn L. Simons, Prithijit S. Chathrath, Grant E. Meyer und Yousry S. Attia: Sexual dimorphism of the internal mandibular chamber in Fayum Pliohyracidae (Mammalia). Journal of Vertebrate Paleontology 26 (1), 2006, S. 160–169
- Martin Pickford: New Titanohyracidae (Hyracoidea: Afrotheria) from the Late Eocene of Namibia. Communications of the Geological Survey of Namibia 16, 2015, S. 200–214
- Martin Pickford, Salvador Moyà Solà und Pierre Mein: A revised phylogeny of the Hyracoidea (Mammalia) based on new specimens of Pliohyracidae from Africa and Europe. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 205 (2), 1997, S. 265–288
- Martin Pickford: Revision of the Early Miocene Hyracoidea (Mammalia) of East Africa. Comptes Rendus Palevol 3, 2004, S. 675–690
- Charles W. Andrews: Further notes on the mammals of the Eocene of Egypt. Part III. Geological Magazine 5 (1), 1904, S. 211–215 ()
- Charles W. Andrews: A descriptive catalogue of the Tertiary Vertebrata of the Fayum, Egypt. London, 1906, S. 1–324 (S. 96–98 und Tafel 7)
- Max Schlosser: Über einige fossile Säugetiere aus dem Oligocän von Ägypten. Zoologischer Anzeiger 35, 1910, S. 500–508 ()
- Max Schlosser: Beiträge zur Kenntnis der Oligozänen Landsäugetiere aus dem Fayum, Ägypten. Beiträge zur Paläontologie und Geologie Österreich-Ungarns und des Orients 24, 1911, S. 51–167 ()