Hyaenodonta
Die Hyaenodonta (selten auch Hyaenodontida) sind eine ausgestorbene Gruppe fleischfressender Säugetiere, die in Nordamerika, Europa, Asien und Afrika vorkamen. Sie erschienen mit Lahimia im mittleren Paläozän und starben in Nordamerika und Europa mit dem Ende des Oligozän, in Afrika und Asien während des Miozän aus.
| Hyaenodonta | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelett von Hyaenodon | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Mittleres Paläozän (Seelandium) bis Mittleres Miozän (Serravallium) | ||||||||||||
| 61,1 bis 11,1 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Hyaenodonta | ||||||||||||
| Van Valen, 1967 | ||||||||||||
Merkmale

Die Vertreter der Hyaenodonta hatten eine äußere Ähnlichkeit mit den heutigen Hunden und Hyänen. Sie besaßen einen plantigraden oder vollständig digitigraden (v. lat. digitus „Zehe“) Fuß (Zehengänger) und waren teilweise wie heutige Hunde und Hyänen an schnelles Laufen angepasst. Die Hauptachse des Fußes befand sich an der Mittelzehe. Das Wadenbein gelenkte mit dem Fersenbein, eine Artikulation des Sprungbeins mit dem Würfelbein fehlte. Die Zehenendglieder waren eingekerbt. Frühe Formen waren noch klein, Lesmesodon, dessen Fossilien in der Grube Messel gefunden wurden, wurde nur 20 cm lang, Arfia wurde schon so groß wie ein Fuchs. Die größten Formen der Hyaenodonta, Hyaenodon, Hyainailouros und Megistotherium hatten Schädel, die doppelt so groß waren wie die heutiger Löwen. Allgemein zeichnete sich der Schädel durch einen langschmalen Bau, eine enge Basis und ein hohes Hinterhauptsbein aus.[1] Ihr Kiefer beherbergten ein Brechscherengebiss, das von der gesamten Backenzahnreihe gebildet wurde, mit Betonung auf den zweiten Backenzahn (Molar) im Oberkiefer und den dritten Backenzahn im Unterkiefer. Bei den möglicherweise verwandten Oxyaenida hatten der erste Backenzahn im Oberkiefer und der zweite im Unterkiefer eine größere Bedeutung, bei den rezenten Raubtieren der vierte Prämolar im Oberkiefer und der erste Backenzahn im Unterkiefer.
Paläobiologie
Das Gehirn der meisten Hyaenodonten war langgestreckt. Das Volumen variierte von rund 3 ml bei kleinen Formen wie Proviverra bis hin zu 334 ml bei großen Vertretern wie Megistotherium. Bezogen auf das Körpergewicht ergibt sich so ein Enzephalisationsquotient von 0,42 bis 0,88. Auffallend ist, dass der Wert bei den Angehörigen der Linie der Hyainailouroidea im Verlauf ihrer Stammesgeschichte relativ gleich blieb mit Schwankungen von 0,4 bis 0,6. Es kam somit nicht zu einer signifikanten Gehirnvolumenzunahme. Allerdings erhöhte sich der Anteil der Großhirnrinde am Gehirnvolumen durch eine deutlichere Furchenbildung. Im Gegensatz dazu vergrößerte sich das Gehirn bei einigen Formen der Hyaenodontoidea auch absolut, da etwa bei Hyaenodon der Enzephalisationsquotient fast 0,9 erreichte. Die Ursachen für die unterschiedliche Entwicklung innerhalb der beiden Linien der Hyaenodonten ist bisher ungeklärt. Bei modernen Raubtieren hat dies variierende Hintergründe. Neben einer Ausweitung des Beutespektrums können hier auch eine verlängerte Lebenserwartung, stärkere Sozialbindungen, längere Tragzeiten oder Betreuungsphasen des Nachwuchses, aber auch größere Nachwuchsgruppen generell verursachend einwirken.[2]
Äußere Systematik
Die Hyaenodonta wurden ursprünglich (als Hyaenodontidae) zusammen mit den Katzen- bzw. Marderähnlichen Oxyaenodonta in die Ordnung der Creodonta, im deutschen auch Urraubtiere genannt, gestellt. Die Oxyaenodonta erschienen schon im mittleren Paläozän in der fossilen Überlieferung und starben früher als die Hyaenodonta wieder aus. Beide Gruppen teilen kaum Synapomorphien, so dass die Gültigkeit des Taxons Creodonta bezweifelt wird.[3][4] Zusammen mit den rezenten Raubtieren (Carnivora) und den Schuppentieren (Manidae) bilden die Hyaenodonta und die Oxyaenodonta das Taxon Ferae.
Innere Systematik
Allgemein
Innere Systematik der Hyaenodonta nach Solé & Mennecart 2019[5]
|
Die Gliederung der Hyaenodonta ist momentan nicht vollständig geklärt und in Diskussion. Generell lassen sich zwei größere taxonomische Einheiten unterscheiden: die Hyaenodontidae und die Hyainailouridae. Beide Gruppen können anhand skelettanatomischer Merkmale relativ gut voneinander getrennt werden. So besitzen die Hyaenodontidae einen Processus postorbitalis, der den Hyainailouridae in der Regel fehlt. Am Hinterhauptsbein reicht der markante Knochenwulst als Ansatzstelle der Nackenmuskulatur bei ersteren seitlich bis zum Processus mastoideus herab, bei letzteren wiederum nicht. Außerdem sind die Gelenkflächen dort zur Verbindung mit der Halswirbelsäule bei den Hyaenodontidae klein und bei den Hyainailouridae groß. Am Unterkiefer tritt das Foramen mentale bei den Hyaenodontidae meist gedoppelt auf und der Winkelfortsatz ist äußerst kräftig. Die Hyainailouridae verfügen häufig über mehr als zwei Foramina, außerdem zeigt sich ihr Winkelfortsatz eher schwach. Bedeutende Unterschiede finden sich auch an den Zähnen. Die typisch sectorialen Molaren weisen mehrere spitze Höcker auf, die drei größten umfassen im Oberkiefer den Para-, Meta- und das Protoconus, im Unterkiefer entsprechend das Para-, Meta- und Protoconid. Sowohl bei den Hyaenodontidae als auch bei den Hyainailouridae sind bei den oberen Mahlzähnen der Para- und der Metaconus zum Amphiconus verschmolzen. Allerdings überragt bei den Hyaenodontidae der Metaconus den Paraconus, bei den Hyainailouridae ist dies genau umgekehrt.[1][6]
Innerhalb der Hyainailouridae werden noch die Apterodontinae und die Hyainailourinae voneinander unterschieden. Beide Gruppen weichen in speziellen Zahnmerkmalen voneinander ab. Die Hyainailourinae bilden ein hypercarnivores Gebiss aus, das heißt, es kommt zu einem Rückgang in der Komplexität der Zahnstruktur. Dadurch verliert das Gebiss weitgehend seine durchlöchernde und brechende Funktion und entwickelt sich hin zu einem starken Schneidwerkzeug. Dies geschieht etwa im Unterkiefer durch die Reduktion des Metaconids (durchlöchernd) und des Talonids (brechend; hierbei handelt es sich um eine tieferliegende Zahnfläche der Unterkiefermolaren, in die bei Gebissschluss der Protoconus der Oberkiefermolare greift). Dadurch können zahlreiche Vertreter der Hyainailouridae als Aasfresser angesehen werden. Heutige hypercarnivore Raubtiere beziehen zu über 70 % ihrer Nahrung von Wirbeltieren. Die Apterodontinae verlieren an den Unterkiefermolaren zwar auch einzelne Höcker (Metaconid und Paraconid), allerdings bleibt das Talonid erhalten. Diese eher ungewöhnliche Zahngestaltung wird häufig mit einer Spezialisierung auf Fische oder auf Wirbellose interpretiert.[1][7]
Problematisch ist die genaue Stellung der Teratodontinae. Einige Forscher sehen sie als näher verwandt mit den Hyaenodontidae an und begründen dies mit dem ausgebildeten Processus postorbitalis oder dem kräftigen Winkelfortsatz.[1] Andere Wissenschaftler vereinen die Teratodontinae mit den Hyainailouridae in einer gemeinsamen höheren Gruppe, den Hyainailouroidea. Dies geschieht unter anderem aufgrund des weniger kräftigen Hinterhauptskamms oder der mehrfach ausgebildeten Foramina am Unterkiefer. Wie die Hyaenodontidae und die Hyainailouridae besitzen auch die Teratodontinae im Oberkiefer einen fusionierten Para- und Metaconus, wobei beide nur an der Basis vereint sind und die Spitzen frei stehen. In Übereinstimmung mit den Hyaenodontidae ist der Metaconus höher als der Paraconus.[6][8]
Abseits dieser größeren Gruppen werden davon abgetrennt die Limnocyoninae unterschieden. Bei diesen handelt es sich um relativ kleine Vertreter der Hyaenodonta, deren Hauptmerkmal der Verlust oder die Reduktion des oberen und unteren dritten Mahlzahns darstellt. Die oberen Molaren weisen einen fusionierten Para- und Metaconus auf, was aber innerhalb der einzelnen Gattungen graduell abweicht.[9] Die Proviverrinae wiederum vereinen eine eher urtümliche Stammgruppe der Hyaenodonta mit deutlich unterscheidbaren Höckern auf den Molaren.[4][10] Sie stehen manchmal aufgrund ihrer sehr ursprünglichen Merkmale in der eigenen höheren Gruppe der Proviverroidea,[3] eine Studie aus dem Jahr 2019 verweist sie dagegen in die Nahverwandtschaft der Hyaenodontidae.[5] Die Prionogalidae wurden ursprünglich aufgrund ihrer ungewöhnlichen Backenzahngestaltung als Höhere Säugetiere mit ungenauen Verwandtschaftsverhältnissen klassifiziert. Sie stellen zwergenhafte Beutegreifer dar, bei denen der jeweils hinterste Molar zurückgebildet ist, während die restlichen Backenzähne extrem hypercarnivore Eigenschaften aufweisen. Neuere phylogenetische Untersuchungen verweisen sie zu den Hyaenodonta, je nach Wichtung der Merkmale bilden sie entweder eine Klade mit den Hyaenodontidae oder mit den Hyainailouridae.[11][12]
Die systematische Zuweisung der säbelzähnigen Machaeroidinae ist unklar. Einerseits werden sie als näher verwandt mit den Limnocyoninae angesehen, andererseits auch den Oxyaenodonta zugeschlagen.[13] Eine im Jahr 2018 veröffentlichte Untersuchung eines nahezu vollständigen Skelettes aus Utah spricht aufgrund des Baus des Bewegungsapparates für eine Nahverwandtschaft mit den Oxyaenodonta.[14]
Überblick über die Familien und Gattungen der Hyaenodonta
Die hier dargestellte Gliederung basiert weitgehehend auf den Untersuchungen von Borths et al. 2016 und 2017.[6][8] Demnach setzen sich die Hyaenodonta folgendermaßen zusammen:[15][4][16][17][1][10][18][19][5][20][2]
- Ordnung Hyaenodonta Van Valen, 1967
- Familie incertae sedis
- Unterfamilie Arfiinae Solé, 2013[4] (auch Arfianinae)
- Arfia Van Valen, 1965
- Familie incertae sedis
- Unterfamilie Limnocyoninae Wortman, 1902
- Prolimnocyon Matthew & Granger, 1915
- Limnocyon Marsh, 1872
 Schädel von Limnocyon
Schädel von Limnocyon - Thinocyon Marsh, 1872
- Iridodon Morlo & Gunnell, 2003[9]
- Oxyaenodon Matthew, 1899
- Prolaena Xu et al., 1979
- Familie incertae sedis
- Sinopa-Klade
- Prototomus Cope, 1874
- Sinopa Leidy, 1871
 Skelett von Sinopa grangeri im National Museum of Natural History in Washington, DC
Skelett von Sinopa grangeri im National Museum of Natural History in Washington, DC - Acarictis Gingerich & Deutsch, 1989
- Proviverroides Bown, 1982
- Überfamilie Hyaenodontoidea Leidy, 1869
- Unterfamilie Proviverrinae Schlosser, 1886
- Allopterodon Ginsburg, 1978
- Parvagula Lange-Badré, 1987
- Lesmesodon Morlo & Habersetzer, 1999[22]
 Fossil von Lesmesodon edingeri im Museo Civico di Storia Naturale di Milano
Fossil von Lesmesodon edingeri im Museo Civico di Storia Naturale di Milano - Morlodon Solé, 2013[4]
- Proviverra Rütimeyer, 1862
- Minimovellentodon Solé, Falconnet & Yves, 2014[16]
- Alienetherium Lange-Badré, 1981
- Consobrinus Lange-Badré, 1979
- Deltatherium Cope, 1881
- Didelphodus Cope, 1882
- Paenoxyaenoides Lange-Badré, 1979
- Praecodens Lange-Badré, 1981
- Protoproviverra Lemoine, 1891
- Familie Prionogalidae Morales, Pickford & Salesa, 2008[23]
- Prionogale Schmidt-Kittler & Heizmann, 1991
- Namasector Morales, Pickford & Salesa, 2008[23]
- Familie Hyaenodontidae Leidy, 1869
- Eurotherium Polly & Lange-Badré, 1993[24]
- Cartierodon Solé & Mennecart, 2019[5]
- Prodissopsalis Matthes, 1950
- Preregidens Solé, Falconnet & Vidalenc, 2015[1]
- Leonhardtina Matthes, 1952
- Matthodon Lange-Badré & Haubold, 1990
- Oxyaenoides Matthes, 1967
- Thereutherium Filhol, 1876
- Boritia Solé, Falconnet & Yves, 2014[16]
- Cynohyaenodon-Klade
- Cynohyaenodon Filhol, 1873
- Quercytherium Filhol, 1880
- Paracynohyaenodon Martin, 1906
- Unterfamilie Hyaenodontinae Trouessart, 1885
- Propterodon Martin, 1906
- Hyaenodon Laizer & Parieu, 1838
 Schädel von Hyaenodon cayluxi im Muséum national d'Histoire naturelle in Paris
Schädel von Hyaenodon cayluxi im Muséum national d'Histoire naturelle in Paris - Isphanatherium Lavrov & Averianov, 1998[25]
- Megalopterodon Dashzeveg, 1964
- Neoparapterodon Lavrov, 1996
- Familie incertae sedis
- Lahimia Solé & Gheerbrant, 2009[26]
- Boualitomus Gheerbrant, Iarochene, Amghzaz & Bouya, 2006[27]
- Pyrocyon Gingerich & Deutsch, 1989
- Gazinocyon Polly, 1996
- Indohyaenodon Bajpai, Kapur & Thewissen, 2009[28]
- Koholia Crochet, 1988
- Tritemnodon Matthew, 1906
 Skelett von Tritemnodon
Skelett von Tritemnodon - Yarshea Egi, Holroyd, Tsubamoto, Shigehara, Takei, Tun, Aung & Soe, 2004[29]
- Galecyon-Klade
- Galecyon Gingerich & Deutsch, 1989
- Überfamilie Hyainailouroidea Borths, Holroyd & Seiffert, 2016[6]
- Familie incertae sedis
- Unterfamilie Teratodontinae Savage, 1965
- Furodon Solé, Lhuillier, Adaci, Bensalah, Mahboubi & Tabuce, 2014[17]
- Kyawdawia Egi, Holroyd, Tsubamoto, Soe, Takai & Ciochon, 2005[30]
- Paratritemnodon Ranga Rao, 1973
- Metasinopa Osborn, 1909
- Pakakali Borths & Stevens, 2017[8]
- Ekweeconfractus Flink, Cote, Rossie, Kibii & Werdelin, 2021[2]
- Glibzegdouia Crochet, Peigné & Mahboubi, 2001[31]
- Brychotherium Borths, Holroyd & Seiffert, 2016[6]
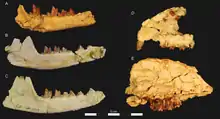 Unterkiefer- und Schädelreste von Brychotherium aus dem Fayyum
Unterkiefer- und Schädelreste von Brychotherium aus dem Fayyum - Masrasector Simons & Gingerich, 1974[32]
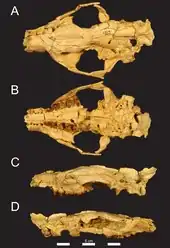 Schädel von Masrasector aus dem Fayyum
Schädel von Masrasector aus dem Fayyum - Teratodon Savage, 1965
- Anasinopa Savage, 1965
- Dissopsalis Pilgrim, 1910
- Familie Hyainailouridae Pilgrim, 1932
- Unterfamilie Apterodontinae Szalay, 1967
- Quasiapterodon Lavrov, 1999
- Apterodon Fischer, 1880
- Unterfamilie Hyainailourinae Pilgrim, 1932
- Orienspterodon Egi, Tsubamoto & Takai, 2007[33]
- Kerberos Solé, Amson, Borths, Vodalenc, Morlo & Bastl, 2015[13]
- Pterodon Blainville, 1839
- Metapterodon Stromer, 1922
- Hemipsalodon Cope, 1885
- Akhnatenavus Holroyd, 1999[34]
 Oberkiefer von Akhnatenavus
Oberkiefer von Akhnatenavus - Paroxyaena Martin, 1906
- Falcatodon Morales & Pickford, 2017[19]
- Isohyaenodon Savage, 1965
- Sectisodon Morales & Pickford, 2017[19]
- Exiguodon Morales & Pickford, 2017[19]
- Buhakia Morlo, Miller & El-Barkooky, 2007[35]
- Mlanyama Rasmussen & Gutierrez, 2009[36]
- Hyainailouros Biedermann, 1863
- Leakitherium Savage, 1965
- Megistotherium Savage, 1973
- Parapterodon Lange-Badré, 1979
- Parvavorodon Solé, Lhuillier, Adaci, Bensalah, Mahboubi & Tabuce, 2014[17]
- Sivapterodon Ginsburg, 1980
- Simbakubwa Borths & Stevens, 2019[37]

Als problematisch erwiesen sich mehrere weitere Unterfamilien, etwa die Koholiinae, 1988 von Jean-Yves Crochet eingeführt, welche Gattungen wie Koholia, Lahimia und Boualitomus zusammenführen.[26] Ähnliches gilt für die Sinopinae (oder Sinopaninae) von Floréal Solé aus dem Jahr 2013 um Sinopa, Prototomus, Tritemnodon und Pyrocyon[4] sowie für die Indohyaenodontinae von Solé und Kollegen 2014 mit Indohyaenodon, Paratritemnodon und Kyawdawia.[17] Hier zeigten phylogenetische Studien, dass die Zusammensetzung der Gruppen meist nicht in sich schlüssig ist.[6][8] Zudem bestehen innerhalb der Hyainailouridae zahlreichen Triben, so die Paroxyaenini, die Hyainailourini,[13] die Dissopsalini und die Metapterodontini.[19] Auch hier ergeben die phylogenetischen Analysen teils unterschiedliche Verwandtschaftsverhältnisse.[6][8]
Literatur
- Thomas S. Kemp: The Origin and Evolution of Mammals. Oxford University Press, Oxford u. a. 2005, ISBN 0-19-850760-7.
- Jordi Augusti, Mauricio Antón: Mammoths, Sabertooths, and Hominids. 65 Million Years of Mammalian Evolution in Europe. Columbia University Press, New York NY u. a. 2002, ISBN 0-231-11640-3.
- Alan Turner, Mauricio Antón: Evolving Eden. An Illustrated Guide to the Evolution of the African Large-Mammal Fauna. Columbia University Press, New York NY 2004, ISBN 0-231-11944-5.
Einzelnachweise
- Floréal Solé, Jocelyn Falconnet und Dominique Vidalenc: New fossil Hyaenodonta (Mammalia, Placentalia) from the Ypresian and Lutetian of France and the evolution of the Proviverrinae in Southern Europe. Palaeontology 58 (6), 2015, S. 1049–1072
- Therese Flink, Susanne Cote, James B. Rossie, Job M. Kibii und Lars Werdelin: The neurocranium of Ekweeconfractusamoruigen. et sp. nov. (Hyaenodonta,Mammalia) and the evolution of the brain in some hyaenodontan carnivores. Journal of Vertebrate Paleontology, 2021, S. e1927748, doi:10.1080/02724634.2021.1927748
- Michael Morlo, Gregg Gunnell und P. David Polly: What, if not nothing, is a creodont? Phylogeny and classification of Hyaenodontida and other former creodonts. Journal of Vertebrate Paleontology 29 (3 suppl), 2009, S. 152A
- Floréal Solé: New proviverrine genus from the Early Eocene of Europe and the first phylogeny of Late Paleocene-Middle Eocene hyaenodontidans (Mammalia). Journal of Systematic Paleontology 11, 2013, S. 375–398
- Floréal Solé und Bastien Mennecart: A large hyaenodont from the Lutetian of Switzerland expands the body mass range of the European mammalian predators during the Eocene. Acta Palaeontologica Polonica 64, 2019 doi:10.4202/app.00581.2018
- Matthew R. Borths, Patricia A. Holroyd und Erik R. Seiffert: Hyainailourine and teratodontine cranial material from the late Eocene of Egypt and the application of parsimony and Bayesian methods to the phylogeny and biogeography of Hyaenodonta (Placentalia, Mammalia). PeerJ 4, 2016, S. e2639 doi:10.7717/peerj.2639
- Floréal Solé und Sandrine Ladevèze: Evolution of the hypercarnivorous dentition in mammals (Metatheria, Eutheria) and its bearing on the development of tribosphenic molars. Evolution & Development 19 (2), S. 56–68
- Matthew R. Borths und Nancy J. Stevens: The first hyaenodont from the late Oligocene Nsungwe Formation of Tanzania: Paleoecological insightsinto the Paleogene-Neogene carnivore transition. PLoSONE 12 (10), 2017, S. e0185301 doi:10.1371/journal.pone.0185301
- Michael Morlo und Gregg F. Gunnell: Small Limnocyonines (Hyaenodontidae, Mammalia) From the Bridgerian Middle Eocene of Wyoming: Thinocyon, Prolimnocyon, And Iridodon, New Genus. Contributions from the Museum of Paleontology, University of Michigan 31 (2), 2003, S. 43–78
- Floréal Solé, Thierry Smith, Rodolphe Tabuce und Bernard Marandat: New Dental Elements of the Oldest Proviverrine Mammal from the Early Eocene of Southern France Support Possible African Origin of the Subfamily. Acta Palaeontologica Polonica 60 (3), 2015, S. 527–538
- Matthew R. Borths und Nancy J. Stevens: Taxonomic affinities of the enigmatic Prionogale breviceps, early Miocene, Kenya. Historical Biology, 2017 doi:10.1080/08912963.2017.1393075
- Jorge Morales und Martin Pickford: A reassessment of Prionogale and Namasector (Prionogalidae, Hyaenodonta, Mammalia) with descriptions of new fossils from Napak, Uganda and Koru, Kenya. Communications of the Geological Survey of Namibia 20, 2018, S. 114–139
- Floréal Solé, Eli Amson, Matthew Borths, Dominique Vidalenc, Michael Morlo und Katharina Bastl: A New Large Hyainailourine from the Bartonian of Europe and Its Bearings on the Evolution and Ecology of Massive Hyaenodonts (Mammalia). PLoS ONE 10 (9), 2015, S. e0135698 doi:10.1371/journal.pone.0135698
- Shawn P. Zack: A skeleton of a Uintan machaeroidine ‘creodont’ and the phylogeny of carnivorous eutherian mammals. Journal of Systematic Palaeontology, 2018 doi:10.1080/14772019.2018.1466374
- Leigh Van Valen: New Paleocene Insectivores and Insectivore classification. Bulletin of the American Museum of Natural History 135, 1967, S. 217–284
- Floréal Solé, Jocelyn Falconnet und Laurent Yves: New proviverrines (Hyaenodontida) from the early Eocene of Europe; phylogeny and ecological evolution of the Proviverrinae. Zoological Journal of the Linnean Society 171, 2014, S. 878–917
- Floréal Solé, Julie Lhuillier, Mohammed Adaci, Mustapha Bensalah, Mohammed Mahboubi und Rodolphe Tabuce: The hyaenodontidans from the Gour Lazib area (?Early Eocene, Algeria): implications concerning the systematics and the origin of the Hyainailourinae and Teratodontinae. Journal of Vertebrate Paleontology 12 (3), 2014, S. 303–322
- Rajendra S. Rana, Kishor Kumar, Shawn P. Zack, Floreal Solé, Kenneth D. Rose, Pieter Missiaen, Lachham Singh, Ashok Sahni und Thierry Smith: Craniodental and Postcranial Morphology of Indohyaenodon raoi from the Early Eocene of India, and Its Implications for Ecology, Phylogeny, and Biogeography of Hyaenodontid Mammals. Journal of Vertebrate Paleontology 35 (5), 2015, S. e965308 doi:10.1080/02724634.2015.965308
- Jorge Morales und Martin Pickford: New hyaenodonts (Ferae, Mammalia) from the Early Miocene of Napak (Uganda), Koru (Kenya) and Grillental (Namibia). Fossil Imprint 73 (3/4), 2017, S. 332–359
- Morgane Dubied, Floréal Solé und Bastein Mennecart: The cranium of Provierra typica (Mammalia, Hyaenodonta) and its impact on hyaenodont phylogeny and endocranial evolution. Palaeontology 2019, S. 1–19 doi:10.1111/pala.12437
- Emmanuel Gheerbrant: Les mammifères paléocènes du bassin d’Ouarzazate (Maroc). III. Adapisoriculidae et autres mammifères (Carnivora, ?Creodonta, Condylarthra, ?Ungulata et incertae sedis). Palaeontographica, Abteilung A 237, 1995, S. 39–132
- Michael Morlo und Jörg Habersetzer: The Hyaenodontidae (Crerodonta, Mammalia) from the lower Eocene (MP 11) of Messel (Germany) with special remarks on new x.ray methods. Courier Forschungsinstitut Senckenberg 216, 1999, S. 31–73
- Jorge Morales, Martin Pickford und M. J. Salesa: Creodonta and Carnivora from the Early Miocene of the Northern Sperrgebiet, Namibia. Memoirs of the Geological Survey of Namibia 20, 2008, S. 291–310
- Paul David Polly, Brigitte Lange-Badré: A new genus Eurotherium (Mammalia, Creodonta) in reference to taxonomic problems with some Eocene hyaenodontids of Eurasia. Comptes Rendus de l’Académie des Sciences Paris. Série 2 317, 1993, S. 991–996
- A. V. Lavrov und A. O. Averianov: The Oldest Asiatic Hyaenodontidae (Mammalia, Creodonta) from the Early Eocene of the Southern Fergana Basin (Andarak-2 Locality). Paleontological Journal 32 (2), 1998, S. 200–205
- Floréal Solé, Emmanuel Gheerbrant, Mbarek Amaghzaz und Baâdi Bouya: Further evidence of the African antiquity of hyaenodontid (‘Creodonta’, Mammalia) evolution. Zoological Journal of the Linnean Society 156 (4), 2009, S. 827–846
- Emmanuel Gheerbrant, Mohamed Iarochene, Mbarek Amaghzaz und Baâdi. Bouya: Early African hyaenodontid mammals and their bearing on the origin of the Creodonta. Geological Magazine 143 (4), 2006, S. 75–489
- Sunil Bajpai, Vivesh V. Kapur und Hans. Thewissen: Creodont and condylarth from the Cambay Shale (Early Eocene, 55-54MA), Vastan Lignite Mine, Gujarat, Western India. Journal of the Palaeontological Society of India 54 (1), 2009, S. 103–109
- Naoko Egi, Patricia A. Holroyd, Takehisa Tsubamoto, Nobuo Shigehara, Masanaru Takai, Soe Thura Tun, Aye Kung Aung und Aung Naing Soe: A new genus and species of hyaenodontid creodont from the Pondaung Formation (Eocene, Myanmar). Journal of Vertebrate Paleontology 24 (2), 2004, S. 502–506
- Naoko Egi, Patricia A. Holroyd, Takehisa Tsubamoto, Aung Naing Soe, Masanaru Takai und Russell L. Ciochon: Proviverrine hyaenodontids (Creodonta: Mammalia) from the Eocene of Myanmar and a phylogenetic analysis of the proviverrines from the para-Tethys area. Journal of Systematic Palaeontology 3 (4), 2005, S. 337–358
- Jean-Yves Crochet, Stéphane Peigné und Mohamed Mahboubi: Ancienneté des Carnivora (Mammalia) en Afrique. In: C. Denys, L. Granjon und A. Poulet (Hrsg.): African Small Mammals. Petits mammifères africains. Paris, 2001, S. 91–100
- Elwyn L. Simons und Philip D. Gingerich: New carnivorous mammals from the Oligocene of Egypt. Annals of the Geological Survey of Egypt 4, 1974, S. 157–166
- Naoko Egi, Takehisa Tsubamoto und Masanaru Takai: Systematic status of Asian "Pterodon" and early evolution of hyaenaelurine hyaenodontid creodonts. Journal of Paleontology 81 (4), 2007, S. 770–778
- Patricia A. Holroyd: New Pterodontinae (Creodonta: Hyaenodontidae) from the late Eocene-early Oligocene Jebel Qatrani Formation, Fayum Province, Egypt. PaleoBios 19 (2), 1999, S. 1–18
- Michael Morlo, Ellen R. Miller und Ahmed El-Barkkoky: Creodonta and Carnivora from Wadi Moghra, Egypt. Journal of Vertebrate Paleontology 27 (1), 2007, S. 145–159
- D. Tab Rasmussen und Mercedes Gutierrez: A Mammalian Fauna from the Late Oligocene of Northwestern Kenya. Palaeontographica Abteilung A 288 (1-3), 2009, S. 1–52
- Matthew R. Borths und Nancy J. Stevens: Simbakubwa kutokaafrika, gen. et sp. nov. (Hyainailourinae, Hyaenodonta, ‘Creodonta,’ Mammalia), A Gigantic Carnivore from the earliest Miocene of Kenya. Journal of Vertebrate Paleontology, 2019, S. e1570222 doi:10.1080/02724634.2019.1570222
Weblinks
- The Paleobiology Database: Hyaenodontidae
- Paleocene mammals of the world:Carnivores, creodonts and carnivorous ungulates: Mammals become predators