Azhdarchidae
Die Azhdarchidae sind eine Gruppe der ausgestorbenen Flugsaurier (Pterosauria). Die weltweit verbreitete Gruppe ist seit der frühen Kreidezeit nachgewiesen, erreicht ihre größte Vielfalt jedoch in der späten Kreidezeit und starb erst am Ende dieser Epoche zusammen mit den Dinosauriern aus. Azhdarchiden schließen die größten flugfähigen Tiere der Erdgeschichte mit ein. Die größten Vertreter zeigten Flügelspannweiten von schätzungsweise 10 bis 11 Meter und erreichten mit Standhöhen von über fünf Meter die Höhe heutiger Giraffen. Azhdarchiden zählen zu den für den Laien am leichtesten zu erkennenden Flugsauriergruppen[2] – charakteristische Merkmale schließen die langen, unbezahnten Kiefer, den langen Hals, die proportional kurzen Flügel sowie die langen Hinterbeine mit ein.[3] Zur Lebensweise dieser Tiere wurde eine Vielzahl von Hypothesen postuliert; die jüngste Studie kommt zu dem Schluss, dass Azhdarchiden wie heutige Schreitvögel am Boden diversen Beutetieren nachstellten. Namensgebend für die zu den Kurzschwanzflugsauriern (Pterodactyloidea) zählenden Azhdarchiden ist die Gattung Azhdarcho, deren Bezeichnung auf Aždahā, den Namen für Drache aus dem Persischen zurückgeht.
| Azhdarchidae | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Plastische Lebendrekonstruktion von Hatzegopteryx | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Kreide (Berriasium bis Maastrichtium)[1] | ||||||||||||
| 145 bis 66 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Azhdarchidae | ||||||||||||
| Nessov, 1984 |
Merkmale
Azhdarchiden zählen zu den seltensten Wirbeltierfossilien,[4] obwohl es sich womöglich um häufige Elemente der kreidezeitlichen Faunen handelte.[5] Der sehr lückenhafte Fossilbericht dieser Gruppe könnte auf die Präferenz für festländische Habitate zurückzuführen sein, in welchen die fragilen Flugsaurierskelette ein geringeres Erhaltungspotential zeigen als in marinen Habitaten, die von vielen besser bekannten Flugsauriergruppen bevorzugt wurden.[6] Lediglich Quetzalcoatlus sp. und Zhejiangopterus sind durch jeweils mehrere fragmentarische Skelette bekannt, während alle übrigen Vertreter nur auf stark unvollständigen Resten basieren.[4] Die bekannten Fossilien lassen allerdings vermuten, dass sich der Skelettbau bei verschiedenen Vertretern der Gruppe nur geringfügig unterschied; größere Variationen finden sich lediglich im Bau des Schädels.[7]
Größe
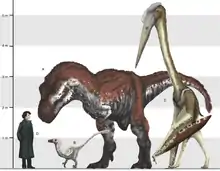
Die Körpergröße variiert bei Azhdarchiden stärker als bei jeder anderen Flugsaurier-Gruppe. Der kleinste bekannte Vertreter ist Montanazhdarcho mit einer Flügelspannweite von etwa 2,5 Metern. Eine Analyse der Wachstumsringe der Knochen zeigt, dass es sich bei dem einzigen bekannten Exemplar tatsächlich um ein ausgewachsenes Tier handelte, und nicht etwa um ein Jungtier. Vollständige Skelette sind von dem nur wenig größeren Zhejiangopterus (3,5 Meter Flügelspannweite) und dem mittelgroßen Quetzalcoatlus sp. (5 Meter Flügelspannweite) bekannt. Zu den größten Vertretern zählen Quetzalcoatlus nortrophi, Arambourgiania und Hatzegopteryx, welche vermutlich Flügelspannweiten von über 10 Meter erreichten; damit handelt es sich um die bei weitem größten Flugsaurier und die größten flugfähigen Tiere überhaupt.[8] Diese gigantischen Formen basieren lediglich auf sehr fragmentarischen Fossilien; so ist Arambourgiania durch einen unvollständigen Halswirbel, Quetzalcoatlus nortrophi durch einen Oberarmknochen und weiteren Fragmenten des Flügels, und Hatzegopteryx durch einen Oberarmknochen, einen Oberschenkelknochen sowie fragmentarischen Schädelknochen bekannt.[9] Frühe Schätzungen der Flügelspannweite von Quetzalcoatlus nortrophi reichen von 11 bis 21 Metern; erst mit zunehmendem Verständnis der Anatomie und Körperproportionen der Gruppe pendelten sich Schätzungen zwischen 10 und 13 Metern ein. Eine aktuelle Studie errechnet 10,5 Meter Flügelspannweite für Hatzegopteryx und Quetzalcoatlus nortrophi, weist jedoch darauf hin, dass derzeit keine verlässlichen Schätzungen für Arambourgiania möglich sind.[10] Ein Azhdarchide mit einer Flügelspannweite von 10,5 Metern hätte am Boden eine Schulterhöhe von 2,5 Metern und eine Gesamthöhe von über 5 Metern erreicht.[8]
Schätzungen des Körpergewichts dieser Tiere sind höchst umstritten.[10][11] Zahlreiche Knochen des Flugsaurier-Skeletts waren mit luftgefüllten Kammern durchzogen (pneumatisiert), was ursprünglich als Anpassung an eine extreme Leichtbauweise interpretiert wurde. Lange galten Flugsaurier somit als extrem leichtgewichtige Segler, die wegen der zur Gewichtsersparnis reduzierten Muskelmasse lediglich zu sporadischen Flügelschlägen in der Lage waren.[12] Für einen Quetzalcoatlus nortrophi mit einer Flügelspannweite von 10 Metern wurde von einer 1974er Studie ein Gewicht von nur 50 bis 60 Kilogramm vorgeschlagen.[10] Viele jüngere Studien gehen derweil davon aus, dass das Gewicht von Flugsauriern mit dem gleich großer moderner Vögel und Fledermäuse vergleichbar ist; demnach kann ein Azhdarchide mit einer Spannweite von 10 Meter auf bis zu 250 kg geschätzt werden.[12] Gewichtsschätzungen bestimmter Spezies hängen vor allem von der rekonstruierten Flügelspannweite ab; so wäre ein Tier mit einer Flügelspannweite von 13 Metern vermutlich fast doppelt so schwer gewesen wie ein Tier mit einer Flügelspannweite von 10 Metern.[10]
Paläontologen errechneten eine Flügelspannweite von 12 bis 13 Metern als Obergrenze für einen flugfähigen Azhdarchiden – bei noch größeren Spannweiten sei die Belastungsgrenze des Skeletts überschritten und ein effektiver Start nicht mehr möglich. Demnach haben Azhdarchiden ihren Gigantismus bis an die Grenzen des physikalisch Machbaren innerhalb ihres Bauplans gebracht. Der Gigantismus bot den Tieren vermutlich eine Reihe von Vorteilen, wie beispielsweise eine effizientere Fortbewegung, einen größeren Schutz gegenüber Räubern, die Überdauerung längerer Zeiten geringen Nahrungsangebots sowie eine effizientere Wärmeregulierung. Folglich wurden Azhdarchiden, ebenso wie viele andere Tiergruppen mit Gigantismus, möglicherweise schlicht deswegen so groß, weil ihr Skelettbauplan und ihre Ökologie dies zuließen.[7]
Schädel

Der Schädel war leicht gebaut, lang und in Seitenansicht dreieckig.[13] Vor den Augenhöhlen befand sich ein sehr großes Schädelfenster, das Nasoantorbitalfenster, welches unter anderem die Nasenöffnungen beherbergte. Während die Augenhöhlen auf den unteren Bereich des Schädels beschränkt waren, vereinnahmte das Nasoantorbitalfenster fast die gesamte Tiefe des Schädels. Die Kieferknochen trugen beim lebenden Tier einen Schnabel aus Horn, worauf kleine Öffnungen im Kiefer weisen.[14] Die Kiefermuskeln waren lediglich schwach ausgeprägt; der wichtigste Schließmuskel war wie bei heutigen Vögeln der vordere Flügelmuskel (Musculus pterygoideus anterior).[13] Das Hinterhaupt des Schädels war annähernd horizontal ausgerichtet – das Kopfgelenk befand sich also auf der Unterseite des Schädels, wodurch die Halswirbelsäule in einem Winkel von fast 90° relativ zur Längsachse des Schädels ansetzte.[14][13] Viele Flugsaurier zeigten auffällige Kopfkämme, die sowohl von den Kiefern als auch von der Ober- und Hinterseite des Schädels ausgehen konnten. Bei Azhdarchiden ist ein Kamm bisher lediglich bei Quetzalcoatlus sp. nachgewiesen, während der vollständige Schädel von Zhejiangopterus keinen Hinweis auf einen Kamm zeigt. Der Kamm von Quetzalcoatlus sp. ist buckelförmig und befand sich über der hinteren Hälfte des Nasoantorbitalfensters.[7]
Die bisher bekannten Schädelreste können möglicherweise einem „langschnäuzigen“ und einem „kurzschnäuzigen“ Bauplan zugeordnet werden. Zhejiangopterus, Quetzalcoatlus sp. und vermutlich Alanqa zeigten sehr lange und niedrige Schädel, die zehn Mal länger als breit werden konnten. Der Schädelbereich vor dem Nasoantorbitalfenster, das Rostrum, machte mehr als die Hälfte der Kieferlänge aus. Dagegen wiesen Bankonydraco und möglicherweise der noch unbeschriebene „Javelina-Azhdarchide“ sowie Azhdarcho vergleichsweise kurze Schädel auf; der Kiefer von Bankonydraco ist im Verhältnis zu seiner Breite lediglich halb so lang wie der von Quetzalcoatlus sp.[7] Über den Schädel der größten Vertreter der Gruppe ist wenig bekannt. Die Kiefer von Hatzegopteryx maßen fast einen halben Meter in der Breite – entspräche der Schädelbau dem des kurzschnäuzigen Bankonydraco, wären die Kiefer 2,5 Meter und der gesamte Schädel annähernd 3 Meter lang gewesen. Träfe diese Schätzung zu, hätte der Schädel von Hatzegopteryx zu den längsten aller bekannter landlebender Tiere gezählt.[15]
Restskelett
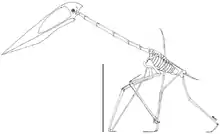
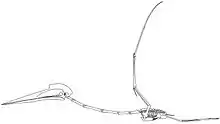
Der Großteil des sich dem Schädel anschließenden Skeletts war mit luftgefüllten Kammern durchzogen, einschließlich des gesamten Flügels. Diese Kammern wurden wie bei Vögeln durch Luftsäcke ausgefüllt, die mit der Lunge in Verbindung standen.[16][17]
Die charakteristischen röhrenförmigen Halswirbel sind die am häufigsten gefundenen und am besten untersuchten Azhdarchiden-Fossilien.[18] Durch Verlängerung des dritten bis achten Halswirbels war der Hals proportional länger als bei allen anderen Flugsauriern: der fünfte Halswirbel war dabei acht Mal so lang wie breit. Der unvollständige Halswirbel von Arambourgiania lässt vermuten, dass die größten Vertreter eine Halslänge von etwa drei Meter zeigten. In den meisten Halswirbeln fehlten Querfortsätze, während die Dornfortsätze nur als niedrige Kämme ausgeprägt oder vollständig reduziert waren.[18] Die Gelenkfortsätze waren groß und erlaubten nur geringe Bewegungen zwischen den einzelnen Wirbeln, was den Hals relativ inflexibel machte. Das schwach ausgeprägte Kopfgelenk lässt vermuten, dass auch die Beweglichkeit des Kopfes eingeschränkt war.[14]
Der Rumpf, obwohl robust gebaut, war verhältnismäßig klein: selbst bei den größten Vertretern maß er vermutlich lediglich 70 Zentimeter Länge.[16] Die vorderen Rückenwirbel waren miteinander zu einem Notarium verschmolzen, was möglicherweise ein Durchbiegen der Wirbelsäule beim Flügelschlag verhinderte.[19][16] Große Muskelansatzstellen in der Schulterregion lassen Gregory Paul (2002) vermuten, dass die Flugmuskeln der größten Vertreter so schwer waren wie ein ausgewachsener Mensch.[20] Sowohl Schulter- als auch Beckengürtel waren robust im Vergleich mit anderen Flugsauriern. Schulterblatt und Rabenbein entsprachen einander in ihrer Länge, und die hinter dem Hüftgelenk befindliche Erweiterung des Beckens, der Postacetabularprozess, war groß.[16][21]
Der Oberarmknochen war robust gebaut, der Knochenschaft erreichte bei den größten Vertretern eine Breite von acht Zentimetern. Wie bei allen Flugsauriern war der vierte Mittelhandknochen sehr groß, während die ersten drei Mittelhandknochen in ihrer Größe stark reduziert waren. Bei Azhdarchiden war der vierte Mittelhandknochen fast 2,5 Mal so lang wie der Oberarmknochen – damit war er proportional länger als bei allen anderen Flugsauriern und der längste Knochen des Flügels. Der vierte Mittelhandknochen fand seine Verlängerung im vierten Finger, der als stark vergrößerter Flugfinger ausgebildet war. Der Flugfinger war proportional kürzer als bei anderen Flugsauriern und machte weniger als 50 % der gesamten Flügellänge aus. Die vier Fingerglieder des Flugfingers wurden nach außen sukzessive kürzer. Die ersten drei Finger waren deutlich kleiner als der Flugfinger und dienten der vierfüßigen Fortbewegung auf dem Boden. Die Hintergliedmaßen waren im Verhältnis länger als bei anderen Flugsauriern, wobei der Oberschenkelknochen 80 % der Länge des Unterschenkels (Tibiotarsus) erreichte. Die Füße waren klein und schmal aber dennoch robuster als bei vielen anderen Flugsauriern.[16][21]
Entdeckungsgeschichte

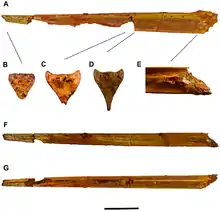
1959 beschrieb Camille Arambourg einen röhrenförmigen Knochen aus Jordanien als Titanopteryx („gigantischer Flügel“) – dieser Knochen war damit das erste Fossil eines Azhdarchiden, das einen Namen erhielt. Der Knochen ist unvollständig, zeigt aber dennoch eine Länge von 62 Zentimetern. Arambourg interpretierte den Knochen als Mittelhandknochen, der zu einem riesigen Flugsaurier mit einer Flügelspannweite von etwa sieben Metern gehört haben müsse. Erst in den 1970er Jahren, nachdem Fossilien des Azhdarchiden Quetzalcoatlus entdeckt wurden, wurde der Knochen als Halswirbel erkannt. Später wurde Titanopteryx in Arambourgiania umbenannt, da sich herausstellte, dass der Name Titanopteryx zum Zeitpunkt der Beschreibung bereits an eine Fliege vergeben war.[9]
Vollständigere Fossilien wurden schließlich zwischen 1972 und 1975 aus dem Big-Bend-Nationalpark (Javelina-Formation) in Texas geborgen. Die Funde umfassen einen gigantischen unvollständigen Flügel, der 1975 als Quetzalcoatlus northropi beschrieben wurde. Außerdem wurden mehrere unvollständige Skelette von kleineren Tieren geborgen, deren Flügelspannweite bei etwa fünf Metern lag; diese Exemplare wurden einer zweiten, noch unbenannten Quetzalcoatlus-Art (Quetzalcoatlus sp.) zugeschrieben. Obwohl es sich bei Quetzalcoatlus sp. um die bis heute besterhaltenen Azhdarchien-Fossilien handelt, steht eine genaue wissenschaftliche Beschreibung noch aus.[9] Heute gehört Quetzalcoatlus zu den in der Öffentlichkeit bekanntesten Flugsauriern.[22]
1984 stellte der russischen Paläontologe Lev Nessov schließlich die Unterfamilie Azhdarchinae auf. Namensgebende Gattung ist der zeitgleich beschriebene Azhdarcho (persisch für „Drache“), der auf fragmentarischen Knochen aus der frühen Oberkreide von Usbekistan basiert. Nur wenige Monate nach der Veröffentlichung des Namens Azhdarchidae erschien eine Arbeit von Kevin Padian, welche für dieselbe Gruppe den Namen Titanopterygiidae vorsah. Da Nessovs Veröffentlichung vor jener von Padian erschien, hat der Name Azhdarchinae nach den Internationalen Regeln für die Zoologische Nomenklatur Priorität. Zwei Jahre später benannte Padian die Azhdarchinae in Azhdarchidae um und erhob die Gruppe somit in den Rang einer Familie.[9][23]
Die wichtigste Entdeckung der 1990er Jahre ist Zhejiangopterus, der durch mehrere unvollständige und stark zerdrückte Skelette aus der späten Oberkreide von China bekannt ist. Ursprünglich wurde Zhejiangopterus 1994 als Vertreter der Flugsaurier-Gruppe Nyctosauridae beschrieben; erst 1997 wurde Zhejiangopterus als Vertreter der Azhdarchidae erkannt. Derzeit gilt die Gattung als die am besten untersuchte dieser Gruppe. Weitere in den 1990er Jahren beschriebene Gattungen schließen Bennettazhia mit ein, der lediglich auf einem Oberarmknochen aus der späten Unterkreide von Oregon basiert. Aus der späten Oberkreide von Montana stammt der kleinwüchsige Montanazhdarcho, der 1995 anhand eines fragmentarischen Skeletts beschrieben wurde. Ein bereits 1914 beschriebenes Fragment eines Halswirbels, das heute als Bogolubovia bekannt ist, wurde 1991 ebenfalls zu den Azhdarchidae gestellt. Das Fossil gilt heute jedoch als verschollen und die Gültigkeit der Gattung wird angezweifelt.[9]
2002 wurde mit Hatzegopteryx ein weiterer gigantischer Vertreter der Familie beschrieben. Ein sehr fragmentarisches Exemplar stammt aus der späten Oberkreide des rumänischen Hațeg-Beckens und schließt Schädelfragmente und einen fragmentarischen Oberarmknochen mit ein. Phosphatodraco, beschrieben im Jahr 2003, ist durch eine fragmentarische Halswirbelsäule aus phosphatreichen Ablagerungen des Ouled-Abdoun-Beckens in Marokko bekannt.[24][25] Etwas vollständigere Überreste aus der mittleren Oberkreide von Ungarn, die unter anderem einen vollständigen Kiefer mit einschließen, wurden 2005 Bakonydraco getauft. Weitere, jedoch nur sehr fragmentarisch überlieferte Vertreter wurden 2007 mit Aralazhdarcho aus Usbekistan und 2008 mit Volgadraco aus Russland beschrieben. Ein zweiter Azhdarchide aus Marokko, Alanqa, wurde 2010 beschrieben und basiert auf fragmentarischen Kiefern. 2011 wurde die Beschreibung von Navajodactylus aus der späten Oberkreide New Mexicos veröffentlicht. Die Gattung ist von umstrittener Gültigkeit, da sie lediglich auf dem Oberende des ersten Fingerglieds des Flugfingers basiert.[9] 2013 wurde mit Eurazhdarcho eine zweite Azhdarchiden-Gattung aus der späten Oberkreide von Rumänien publiziert.[26]
Systematik
Die genauen Verwandtschaftsbeziehungen mit anderen Flugsauriergruppen sind umstritten.[27] Zur Systematik der Flugsaurier existieren zwei konkurrierende Ansätze, die auf unterschiedlichen Datensätzen beruhen, teilweise ihre eigenen Bezeichnungen und Definitionen für übergeordnete Gruppen mitbringen und sich in wichtigen Punkten voneinander unterscheiden.[28] Moderne systematische Analysen sind meist von einem dieser beiden Ansätze abgeleitet.[28] Der erste Ansatz wurde von Alexander Kellner (2003) eingeführt und sieht die Azhdarchidae als Schwestergruppe der Tapejaridae (Tapejara + Tupuxuara); beide Familien werden als Azhdarchoidea zusammengefasst. Die Azhdarchoidea werden von Kellner den Dsungaripteroidea unterstellt, einer von zwei Gruppen innerhalb der Kurzschwanzflugsaurier (Pterodactyloidea).[29] Der zweite Ansatz von David Unwin (2003) zieht den Schluss, dass Tupuxuara näher mit den Azhdarchiden verwandt war als mit Tapejara. Mit Tupuxuara als Schwestergruppe bilden die Azhdarchiden demnach die Gruppe Neoazhdarchia, welche zusammen mit Tapejara bzw. der Tapejaridae die Gruppe Azhdarchoidea bildet. Laut Unwin gehören die Azhdarchoidea innerhalb der Kurzschwanzflugsaurier einer Lophocratia genannten Gruppe an.[30] Erst seit 2008 werden die Chaoyangopteridae als eigenständige Gruppe unterschieden, die womöglich ebenfalls nahe mit den Azhdarchiden verwandt gewesen sein könnten.[31] Die Verwandtschaftsbeziehungen innerhalb der Azhdarchidae können derzeit nicht aufgelöst werden.
Es folgen zwei Kladogramme: das linke illustriert die Verwandtschaftshypothese von Kellner (2003)[29]; das rechte jene von Unwin (2003)[30].
|
|
Stammesgeschichte und Verbreitung
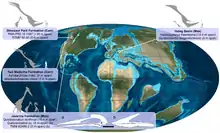
Über den Ursprung der Azhdarchiden ist wenig bekannt. Bis vor kurzem galten stark verlängerte Halswirbel aus dem Oberjura von Tansania und der Jura-Kreide-Grenze von England als die ältesten Nachweise von Azhdarchiden. Brian Andres und Ji Qiang (2008) zeigten jedoch, dass diese Halswirbel wahrscheinlich einer anderen Flugsauriergruppe zuzuordnen sind, der Ctenochasmatidae. In beiden, nur entfernt miteinander verwandten Gruppen hätten sich stark verlängerte Halswirbel unabhängig voneinander ausgebildet, was zeigen könnte, dass beide Gruppen ähnliche ökologische Nischen besetzten.[6][32] Derzeit gilt ein deutlich jüngerer Fund als möglicherweise ältestes Azhdarchiden-Fossil – ein 2010 beschriebener fragmentarischer Halswirbel aus dem Berriasium (Unterkreide, 145 bis 140 Mya) von Rumänien.[32] Sollte sich dieser Nachweis bestätigen, wäre die Gruppe über einen Zeitraum von etwa 80 Millionen Jahren nachgewiesen und hätte damit länger existiert als jede andere Flugsaurier-Gruppe.[1]
Azhdarchiden erreichten offenbar ihre Blütezeit während der Oberkreide – sämtliche benannten Vertreter stammen aus dieser Epoche.[6] Fossilien wurden vor allem in Nordamerika, Europa, Asien und Nordafrika entdeckt; vereinzelte Funde sind aus Argentinien, Japan und Australien bekannt.[4] Damit sind Azhdarchiden auf sämtlichen Kontinenten mit Ausnahme Antarktikas nachgewiesen. Verschiedene Gesteinsformationen enthalten Fossilien von mehr als einer Azhdarchiden-Spezies: So sind in der Javelina-Formation in Texas die Riesenform Quetzalcoatlus northropi sowie zwei mittelgroße Arten (Quetzalcoatlus sp. sowie ein noch unbenannter Vertreter) nachgewiesen. In der Two-Medicine-Formation von Montana tritt neben der kleinen Form Montanazhdarcho ein unbestimmter, deutlich größerer Vertreter auf. In der Dinosaurier-Park-Formation von Alberta ist vermutlich eine Riesenform sowie ein mittelgroßer Vertreter präsent, während im rumänischen Hațeg-Becken der sehr große Hatzegopteryx zusammen mit dem kleinen Vertreter Eurazhdarcho auftritt. Die jeweils gemeinsam auftretenden Spezies könnten zur Vermeidung von Konkurrenz unterschiedliche ökologische Nischen besetzt haben.[26]
Azhdarchiden starben mit dem Kreide-Tertiär-Massenaussterben vor 66 Millionen Jahren aus. Ein wichtiger Grund für das Aussterben liegt vermutlich in der Körpergröße dieser Tiere: Große Tiere sind generell stark spezialisiert, haben niedrige Populationsdichten und pflanzen sich nur langsam fort, weshalb sie sich an veränderte Umweltbedingungen weniger schnell anpassen können als kleine und unspezialisierte Tiere. Die meisten anderen Flugsaurier-Gruppen verschwinden bereits etwa 30 Millionen Jahre vor dem Massenaussterben aus dem Fossilbericht.[33] Umstritten bleibt allerdings, ob Flugsaurier tatsächlich lange vor dem Massenaussterben in ihrer Vielfalt stark dezimiert wurden, oder ob Fossilien schlicht deswegen unbekannt bleiben, weil entsprechende Fossillagerstätten fehlen (der sogenannte Lagerstätten-Effekt).[34] In diesem Zusammenhang sind die Funde von Flugsauriern einschließlich Vertretern der Azhdarchidae, der Nyctosauridae und der Pteranodontidae aus dem Ouled-Abdoun-Becken von Bedeutung.[24][25]
Paläobiologie
Flug

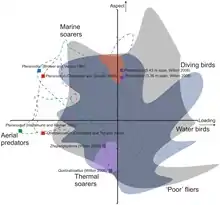
Kontrovers diskutiert wird die Frage, wie Flugsaurier und insbesondere große Vertreter wie Azhdarchiden ihren Flug starteten. Traditionell wird ein Start mit Hilfe der Hinterbeine angenommen, wie er bei heutigen Vögeln zu beobachten ist.[10] Chatterjee und Templin (2004) argumentierten, dass Quetzalcoatlus nortrophi nur durch seine sehr geringe Körpermasse von 70 Kilogramm zum Start befähigt sei; ein Start könne erfolgen, wenn sich das Tier auf zwei Beinen aufrichtet und gegen den Wind anläuft oder sich von einem Hang abstößt.[35] Jüngere Studien vermuten indes, dass Flugsaurier aus einer vierfüßigen Haltung heraus mit Hilfe der Vordergliedmaßen starteten, ähnlich wie heutige Fledermäuse. Auf diese Weise können die großen Flugmuskeln zum Emporhieven des Körpers genutzt werden; die Vordergliedmaßen großer Azhdarchiden seien stark genug, um ein Gewicht von annähernd 500 kg in die Luft zu befördern.[10]
Donald Henderson (2010) hypothetisiert, dass große Azhdarchiden wie Quetzalcoatlus nortrophi sekundär flugunfähig gewesen sein könnten. So seien die Flügel zu kurz für einen effektiven Flug, und die Hintergliedmaßen besser an eine Fortbewegung auf dem Boden angepasst als bei anderen Flugsauriern. Das Gewicht von Quetzalcoatlus nortrophi berechnete Henderson auf 544 Kilogramm, damit sei dieses Tier schlicht zu schwer zum Fliegen gewesen.[36] Mark Witton und Michael Habib (2010) halten dagegen, dass Henderson für die Gewichtsschätzung einen zu großen Rumpf annahm und somit ein Gewicht von 200 bis 250 kg realistischer sei. Auch große Azhdarchiden würden sämtliche Anpassungen an einen Flug zeigen, die sich auch bei kleineren Flugsauriern finden.[10]
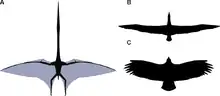
Witton und Darren Naish (2008) argumentieren, dass die kurzen und breiten Flügel an ein Fliegen über Land angepasst seien. Mit der durch die breiten Flügel bedingten geringen Flächenbelastung hätten Azhdarchiden möglicherweise thermische Aufwinde genutzt. Kurze und breite Flügel hätten die Tiere wendiger gemacht und ermöglichten außerdem einen schnellen Start und Aufstieg, was beispielsweise in vegetationsreichen Umgebungen vorteilhaft gewesen wäre. Ähnliche Längenverhältnisse der Flügel fänden sich bei heutigen landlebenden Vögeln wie beispielsweise dem Andenkondor, während marine Segler, wie beispielsweise der Wanderalbatros, generell eher lange und schlanke Flügel zeigen.[37]
Traditionell gelten große Flugsaurier als extrem leichtgewichtig, was einen Segelflug ermöglicht hätte, der deutlich langsamer war als der heutiger Vögel. Wird ein Gewicht angenommen, das mit dem heutiger Vögel und Fledermäuse vergleichbar ist, ändert sich dieses Bild drastisch: Witton und Habib (2010) berechnen, dass ein 200 Kilogramm schwerer Azhdarchide mit einer Flügelspannweite von 10 Metern nach dem Start für etwa zwei Minuten einen kraftvollen Flügelschlag aufrechterhalten konnte, bei dem das Tier Geschwindigkeiten von über 100 km/h erreichte, bevor das Tier in einem etwas langsameren Segelflug überging.[10][38] Ein solcher Azhdarchide sei in der Lage gewesen, über 16.000 Kilometer ohne Zwischenlandung zurückzulegen, mehr als jedes andere Tier.[39]
Fortbewegung auf dem Boden
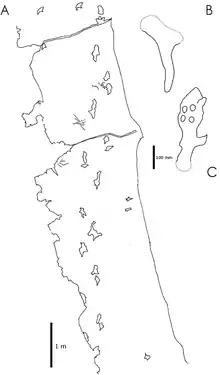
Fossile Fußspuren von gehenden und laufenden Flugsauriern zeigen, dass viele Spezies deutlich kompetenter in der Fortbewegung auf dem Boden waren als traditionell angenommen. In Korea entdeckte, Haenamichnus genannte Spuren werden einem sehr großen Vertreter der Azhdarchidae zugeschrieben: Die Hinterfußabdrücke messen 35 Zentimeter in der Länge, was auf ein Tier mit einer Schulterhöhe von fast drei Meter und einer Flügelspannweite von über 10 Meter weist. Diese Fußspuren sind die derzeit einzigen, die einer bestimmten Flugsaurier-Gruppe zugeschrieben werden können; keine andere bekannte Gruppe könnte Fußspuren dieser Größe hinterlassen haben. Eine der Fährtenfolgen misst sieben Meter Länge und ist damit die längste bekannte Fährtenfolge eines Flugsauriers. Diese Fährtenfolge zeigt, dass die langen Gliedmaßen der Azhdarchiden eine schnelle und effiziente Fortbewegung auf dem Boden ermöglichen: Die Fährtenfolge ist schmal, was senkrecht unter dem Körper stehende Gliedmaßen anzeigt, und die Hinterfußabdrücke überlagern regelmäßig die zuvor hinterlassenen Hand-Abdrücke. Die Zehen waren mit fleischigen Polstern versehen, wobei Krallenabdrücke fehlen.[40]
Ökologie

Douglas Lawson stellte 1975 in einem Bericht über den neu entdeckten Flugsaurier Quetzalcoatlus die erste Hypothese zur Ernährungsweise eines Azhdarchiden auf. Obwohl zu dieser Zeit Flugsaurier allgemein als Fischfresser angesehen wurden, argumentierte Lawon, dass es sich bei Quetzalcoatlus um einen Aasfresser gehandelt haben könnte, der seinen langen Hals zum Eindringen in Kadaver nutzte.[41] So träten die Fossilien dieser Gattung in kontinentalen Ablagerungen fernab der Küste auf und wären zusammen mit den Knochen der pflanzenfressenden sauropoden Dinosaurier vorgefunden worden.[42] Verschiedene spätere Autoren kritisierten diese Idee; so sei die Verbindung zu den Sauropoden-Fossilien zufälliger Natur und der Hals zu steif gewesen, um in Kadaver eindringen zu können.[41]
Seit Lawsons Aasfresser-Hypothese wurden zahlreiche weitere Hypothesen aufgestellt. Wann Langston (1981) spekulierte, dass Quetzalcoatlus mit seinem Schnabel im Schlick nach wirbellosen Tieren getastet haben könnte; hierauf würden Bauten von Gliederfüßern weisen, die zusammen mit den Knochen des Flugsauriers gefunden wurden. Gegen diese Hypothese wurde vorgebracht, dass die Verbindung mit den Spurenfossilien zufällig, der Hals zu steif und die Form des Schnabels ungeeignet sei; außerdem fänden sich keine Hinweise auf druckempfindliche Herbstsche Körperchen oder ähnliche Strukturen in den Kieferknochen, die bei vielen heutigen Vögeln der Erkennung von Beutetieren im Schlick dienen. Verschiedene andere Autoren schlugen vor, dass Azhdarchiden flache Gewässer durchwateten, um dort Beutetieren nachstellten. Die langen Beine sowie der lange Hals und Schädel unterstützen diese These; die proportional kleinen Füße eigneten sich jedoch vermutlich nicht für ein Waten auf weichem Sediment. Wieder andere Autoren zogen in Betracht, dass einige Vertreter der Gruppe Früchte gefressen[43] oder weniger gut fliegende Tiere in der Luft gejagt haben könnten.[41]
Der Großteil der Studien vertrat die Hypothese, dass Azhdarchiden Fischfresser gewesen sein könnten. So vermutete Lev Nessov (1984), dass Azhdarcho und andere Azhdarchiden im Flug oder beim Schwimmen mit ihren Schnäbeln das Wasser durchflügten, um bei Kontakt mit Beutetieren zuzuschnappen. Diese Ernährungsweise findet sich bei heutigen Scherenschnäbeln und ermöglicht diesen Vögeln, auch in trüben Gewässern Beutetiere zu fangen. Kritiker bemerken, dass Scherenschnäbel zahlreiche Anpassungen an diese sehr spezialisierte Ernährungsweise zeigen, wie beispielsweise ein verstärktes Kiefergelenk und sehr robuste Halswirbel, die bei Azhdarchiden nicht zu finden seien. Verschiedene Autoren schlugen derweil vor, dass Azhdarchiden im Flug ihre Schnäbel gezielt ins Wasser stießen, um Fische zu erbeuten, ähnlich wie heutige Möwen, Seeschwalben und Fregattvögel. Der lange Hals und Schädel hätte es diesen Flugsauriern ermöglicht, Fische zu erbeuten, ohne dass die Flügel die Wasseroberfläche berührten. Gegner der Hypothese argumentieren, dass die Halswirbelsäule beinahe im rechten Winkel zur Längsachse des Schädels stand; somit hätten die Tiere beim Zustoßen Kopf und Hals nicht in einer Linie anlegen können. Außerdem sei der Hals zu rigide gewesen, und die Schnauzenspitze nicht nach unten gebogen wie bei vielen heutigen Vögeln, die diese Ernährungsweise zeigen.[41]
Die jüngste Hypothese wurde von Witton und Naish (2008) postuliert. Laut diesen Forschern schritten Azhdarchiden trockenes Terrain ab, um opportunistisch kleine bis mittelgroße Wirbeltiere und große Wirbellose sowie sporadisch auch Aas, Eier oder Früchte aufzunehmen. Eine ähnliche Ernährungsweise findet sich bei einigen heutigen Schreitvögeln, beispielsweise bei den Eigentlichen Störchen. Wie die Forscher argumentieren, wurde die Mehrzahl der Azhdarchiden-Fossilien in Flusssedimenten entdeckt, die im Inland zur Ablagerung kamen. Vertreter der Gruppe scheinen somit kontinentale Lebensräume frequentiert zu haben, wie beispielsweise Wald- und Buschlandschaften und Flussniederungen.[26] Die proportional kleinen Füße, die mit fleischigen Polstern und lediglich kleinen Krallen ausgestattet waren, wären auf festem Untergrund am effizientesten gewesen. Die langen Beine seien zum Schreiten durch Vegetation ideal gewesen, der nach unten gerichtete Schnabel ermögliche ein einfaches Erreichen des Bodens, und der lange, steife Hals könne unter anderem genutzt worden sein, um von erhöhter Position aus nach Beute zu spähen oder um den Kopf an Beutetiere anzunähern, bevor diese von den Tritten des Flugsauriers aufgeschreckt werden.[44] Witton (2013) erwägt, dass einige Azhdarchiden in Gruppen auf Nahrungssuche gegangen sein könnten, da gelegentlich Fossilien mehrerer Individuen zusammen gefunden werden.[45] Die größten Azhdarchiden treten in der späten Oberkreide auf, als mittelgroße fleischfressende Dinosaurier auffällig selten waren. Witton (2013) spekuliert daher, dass Azhdarchiden zumindest teilweise die ökologische Nische mittelgroßer Fleischfresser in der späten Oberkreide besetzt haben könnten.[45]
Literatur
- Mark P. Witton: Pterosaurs. Natural History, Evolution, Anatomy. Princeton University Press, Princeton NJ 2013, ISBN 978-0-691-15061-1.
- Mark Witton: Titans of the skies: azhdarchid pterosaurs. In: Geology Today. Bd. 23, Nr. 1, 2007, ISSN 0266-6979, S 33–38, doi:10.1111/j.1365-2451.2007.00596.x.
- Mark P. Witton, Darren Naish: A Reappraisal of Azhdarchid Pterosaur Functional Morphology and Paleoecology. In: PLoS ONE. Bd. 3, Nr. 5, 2008, e2271, doi:10.1371/journal.pone.0002271.
Einzelnachweise
- Witton (2013): Pterosaurs. Natural History, Evolution, Anatomy. S. 249.
- Witton (2013): Pterosaurs. Natural History, Evolution, Anatomy. S. 244.
- Witton und Naish (2008): A reappraisal of Azhdarchid Pterosaur Functional Morphology and Paleoecology. S. 1.
- Attila Ősi, Éric Buffetaut, Edina Prondvai: New pterosaurian remains from the Late Cretaceous (Santonian) of Hungary (Iharkút, Csehbánya Formation). In: Cretaceous Research. Bd. 32, Nr. 4, 2011, ISSN 0195-6671, S. 456–463, doi:10.1016/j.cretres.2011.01.011.
- Witton (2013): Pterosaurs. Natural History, Evolution, Anatomy. S. 248–249.
- Brian Andres, Ji Qiang: A new pterosaur from the Liaoning Province of China, the phylogeny of the Pterodactyloidea, and convergence in their cervical vertebrae. In: Palaeontology. Bd. 51, Nr. 2, 2008, ISSN 0031-0239, S. 453–469, hier S. 453 und 462–463, doi:10.1111/j.1475-4983.2008.00761.x.
- Witton (2013): Pterosaurs. Natural History, Evolution, Anatomy. S. 251.
- Witton (2013): Pterosaurs. Natural History, Evolution, Anatomy. S. 249–251.
- Witton (2013): Pterosaurs. Natural History, Evolution, Anatomy. S. 244–248.
- Mark P. Witton, Michael B. Habib: On the Size and Flight Diversity of Giant Pterosaurs, the Use of Birds as Pterosaur Analogues and Comments on Pterosaur Flightlessness. In: PLoS ONE. Bd. 5, Nr. 11, 2010, e13982, doi:10.1371/journal.pone.0013982.
- Mark P. Witton: A new approach to determining pterosaur body mass and its implications for pterosaur flight. In: Zitteliana. Reihe B: Abhandlungen der Bayerischen Staatssammlung für Paläontologie und Geologie. Bd. 28, 2008, ISSN 1612-4138, S. 143–158, Digitalisat (PDF; 6,88 MB).
- Witton (2013): Pterosaurs. Natural History, Evolution, Anatomy. S. 58.
- Witton und Naish (2008): A reappraisal of Azhdarchid Pterosaur Functional Morphology and Paleoecology. S. 3–5.
- Mark Witton: Titans of the skies: azhdarchid pterosaurs. In: Geology Today. Bd. 23, Nr. 1, 2007, ISSN 0266-6979, S 33–38, doi:10.1111/j.1365-2451.2007.00596.x.
- Witton (2013): Pterosaurs. Natural History, Evolution, Anatomy. S. 252–253.
- Witton (2013): Pterosaurs. Natural History, Evolution, Anatomy. S. 254.
- Witton (2013): Pterosaurs. Natural History, Evolution, Anatomy. S. 37 und 58.
- Witton und Naish (2008): A reappraisal of Azhdarchid Pterosaur Functional Morphology and Paleoecology. S. 5.
- Peter Wellnhofer, Eric Buffetaut, Paul Gigase: A pterosaurian notarium from the Lower Cretaceous of Brazil. In: Paläontologische Zeitschrift. Bd. 57, Nr. 1/2, 1983, S. 147–157, doi:10.1007/BF03031757.
- Gregory S. Paul: Dinosaurs of the Air. The Evolution and Loss of Flight in Dinosaurs and Birds. Johns Hopkins University Press, Baltimore MD u. a. 2002, ISBN 0-8018-6763-0, S. 333.
- Witton (2013): Pterosaurs. Natural History, Evolution, Anatomy. S. 255.
- Alexander O. Averianov: The osteology of Azhdarcho lancicollis Nessov, 1984 (Pterosauria, Azhdarchidae) from the late Cretaceous of Uzbekistan. In: Proceedings of the Zoological Institute of the Russian Academy of Sciences. Bd. 314, Nr. 3, 2010, ISSN 0206-0477, S. 264–317, hier S. 265, Digitalisat (PDF; 4,63 MB).
- Brian Andres, Timothy S. Myers: Lone Star Pterosaurs. In: Transactions of the Royal Society of Edinburgh. Earth and Environmental Science. Bd. 103, Nr. 3/4, 2012, ISSN 1755-6910, S. 383–398. hier S. 391, doi:10.1017/S1755691013000303.
- Xabier Pereda Suberbiola Nathalie Bardet, Stéphane Jouve S, Mohamed Iarochène, Baâdi Bouya, Mbarek Amaghzaz: A new azhdarchid pterosaur from the Late Cretaceous phosphates of Morocco. In: Geological Society, London, Special Publications. Bd. 217, Nr. 3, 2003, S. 79–90.
- Nicholas R. Longrich, David M. Martill, Brian Andres: Late Maastrichtian pterosaurs from North Africa and mass extinction of Pterosauria at the Cretaceous-Paleogene boundary. In: PLoS Biology. Bd. 16, Nr. 3, 2018, S. e2001663, doi:10.1371/journal.pbio.2001663.
- Mátyás Vremir, Alexander W. A. Kellner, Darren Naish, Gareth J. Dyke: A New Azhdarchid Pterosaur from the Late Cretaceous of the Transylvanian Basin, Romania: Implications for Azhdarchid Diversity and Distribution. In: PLoS ONE. Bd. 8, Nr. 1, 2013, S. e54268, doi:10.1371/journal.pone.0054268.
- Witton (2013): Pterosaurs. Natural History, Evolution, Anatomy. S. 92.
- Witton (2013): Pterosaurs. Natural History, Evolution, Anatomy. S. 90.
- Alexander W. A. Kellner: Pterosaur phylogeny and comments on the evolutionary history of the group. In: The Geological Society, London. Special Publications. Bd. 217, Nr. 1, 2003, ISSN 0375-6440, S. 105–137, doi:10.1144/GSL.SP.2003.217.01.10.
- David M. Unwin: On the phylogeny and evolutionary history of pterosaurs. In: The Geological Society, London. Special Publications. Bd. 217, Nr. 1, 2003, ISSN 0375-6440, S. 139–190, doi:10.1144/GSL.SP.2003.217.01.11.
- Witton (2013): Pterosaurs. Natural History, Evolution, Anatomy. S. 228.
- Gareth J. Dyke, Michael J. Benton, Erika Posmosanu, Darren Naish: Early Cretaceous (Berriasian) birds and pterosaurs from the Cornet bauxite mine, Romania. In: Palaeontology. Bd. 54, Nr. 1, 2011, S. 79–95, hier S. 91–92, doi:10.1111/j.1475-4983.2010.00997.x.
- Witton (2013): Pterosaurs. Natural History, Evolution, Anatomy. S. 175.
- Witton (2013): Pterosaurs. Natural History, Evolution, Anatomy. S. 259.
- Sankar Chatterjee, R. Jack Templin: Posture, locomotion, and paleoecology of pterosaurs (= The Geological Society of America. Special Papers. Nr. 376). Geological Society of America, Boulder CO 2004, ISBN 0-8137-2376-0, S. 1–4.
- Donald M. Henderson: Pterosaur body mass estimates from three-dimensional mathematical slicing. In: Journal of Vertebrate Paleontology. Bd. 30, Nr. 3, 2010, ISSN 0272-4634, S. 768–785, doi:10.1080/02724631003758334.
- Witton und Naish (2008): A reappraisal of Azhdarchid Pterosaur Functional Morphology and Paleoecology. S. 8.
- Witton (2013): Pterosaurs. Natural History, Evolution, Anatomy. S. 256.
- Michael B. Habib, Mark P. Witton: Soaring efficiency and long distance travel in giant pterosaurs. In: Proceedings of the Third International Symposium on Pterosaurs (= Acta Geoscientica Sinica. Bd. 31, Supplement 1, 2010). Dizhi Chubanshe, Beijing 2010, S. 27–28.
- Witton und Naish (2008): A reappraisal of Azhdarchid Pterosaur Functional Morphology and Paleoecology. S. 8–9.
- Witton und Naish (2008): A reappraisal of Azhdarchid Pterosaur Functional Morphology and Paleoecology. S. 2 und 9–12.
- Douglas A. Lawson: Pterosaur from the Latest Cretaceous of West Texas: Discovery of the Largest Flying Creature. In: Science. Bd. 187, Nr. 4180, 1975, S. 947–948, doi:10.1126/science.187.4180.947, PMID 17745279.
- Attila Ősi, David B. Weishampel, Coralia M. Jianu: First evidence of azhdarchid pterosaurs from the Late Cretaceous of Hungary. In: Acta Palaeontologica Polonica. Bd. 50, Nr. 4, 2005, ISSN 0567-7920, S. 777–787, online.
- Witton und Naish (2008): A reappraisal of Azhdarchid Pterosaur Functional Morphology and Paleoecology. S. 12–14.
- Witton (2013): Pterosaurs. Natural History, Evolution, Anatomy. S. 258.