Gaviale
Die Gaviale (Gavialidae) sind eine Familie der Krokodile (Crocodylia). Als einziger rezenter Vertreter gilt traditionell der in Nepal und Nord-Indien lebende Gangesgavial (Gavialis gangeticus). Der in Malaysia und im Westen Indonesiens beheimatete Sundagavial (Tomistoma schlegelii) wird traditionell als Mitglied der Echten Krokodile (Crocodylidae) betrachtet. Die Ergebnisse molekulargenetischer Untersuchungen sprechen aber dafür, dass Gangesgavial und Sundagavial näher miteinander verwandt sind als mit allen anderen rezenten Krokodilen.[1][2][3][4][5] Daher wird der Sundagavial nunmehr auch taxonomisch zu den Gavialen gezählt.
| Gaviale | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Gangesgavial (Gavialis gangeticus) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Gavialidae | ||||||||||||
| Adams, 1854 |


Merkmale
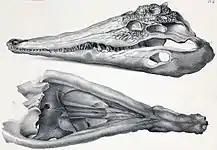
Gaviale leben semiaquatisch und ernähren sich überwiegend piscivor. Als Anpassung an die Fischjagd sind ihre Schädel longirostrin, das heißt, ihre Kiefer sind lang und schmal ausgebildet. Infolgedessen ist die Unterkiefersymphyse, das heißt, die Nahtstelle am vorderen Ende des Unterkiefers, an der die zahntragenden Knochen, die Dentalia, zusammentreffen, bei Gavialen extrem lang und erstreckt sich über etwa die Hälfte der Länge des Unterkiefers. Dadurch ist der Unterkiefer in Ansicht von unten (ventral) oder oben (dorsal) Y-förmig. Die weniger stark spezialisierten Kiefer der meisten Echten Krokodile und Alligatoren sind relativ breit und die Symphyse ist auf den vordersten Teil des Unterkiefers beschränkt, sodass der Unterkiefer in dorsaler bzw. ventraler Ansicht V- oder U-förmig ist. Diese Schnauzenform wird als mesorostrin bzw. brevirostrin bezeichnet.[6]
Darüber hinaus sind die Kiefer der Gaviale mit relativ vielen Zähnen bestückt. Beim Sundagavial sind es 76–84 und beim Gangesgavial 106–110 Zähne. Krokodile und Alligatoren haben meist deutlich weniger Zähne (je nach Art zwischen 64 und 82[7]). Die spitzkonischen Zähne der Gaviale haben eine eher einheitliche Größe. Diese echte Homodontie ist bei Gavialis besonders deutlich ausgeprägt. Bei den übrigen Krokodilen besitzen die Zähne zwar auch alle die gleiche Form, weisen aber teils deutliche Größenunterschiede auf. Diese Pseudoheterodontie wird zusätzlich akzentuiert durch die Wellung (Festonierung) der Kieferränder. Gaviale haben hingegen nur schwach festonierte Kieferränder.
Longirostrine Schädel sind innerhalb der Krokodile im weiteren Sinn (Crocodyliformes) jedoch kein Alleinstellungsmerkmal der Gaviale. Konvergent kommen sie unter anderem bei den ausgestorbenen Teleosauriden (Jura) vor.
Schädel, die hinsichtlich der Schnauzenmorphologie zwischen den mesorostrinen und den longirostrinen Formen stehen, das heißt, die eine zwar sehr schmale und nur schwach festonierte, aber keine deutlich verlängerte Schnauze haben und deren Unterkiefer noch eher V- als Y-förmig ist, werden als stenorostrin bezeichnet. Rezente Vertreter mit stenorostrinen Schädeln sind das Orinoco-Krokodil (Crocodylus intermedius), das Australien-Krokodil (Crocodylus johnstoni) und das Westafrikanische Panzerkrokodil (Mecistops cataphractus),[6] wobei letztgenanntes bisweilen auch als longirostrine Form eingestuft wird.[8]
Systematik und Fossilbericht
Die Gavialidae sind eine der drei rezenten Familien der Krokodile (Crocodylia). Traditionell gelten sie innerhalb der Krokodile als eher entfernt sowohl mit Echten Krokodilen (Crocodylidae) als auch mit Alligatoren und Kaimanen (Alligatoridae) verwandt. Auch in diesem Punkt widersprechen die Ergebnisse molekulargenetischer Untersuchungen der traditionellen Sicht. Stattdessen legen sie nahe, dass die Gaviale näher mit den Echten Krokodilen als mit den Alligatoren verwandt sind.[5] Die meisten aktuellen morphologischen Studien, die auch fossile Arten mit einbeziehen, kommen aber nach wie vor zu Ergebnissen, die die traditionelle Sicht stützen. Eine vergleichende Untersuchung der Ontogenese der rezenten Krokodile ergab, dass sich die Embryonalentwicklung des Gangesgavials grundsätzlich sowohl von jener der Echten Krokodile als auch von der des Sundagavials unterscheidet, wobei die Embryonalentwicklung der beiden letztgenannten Parallelen aufweist. Ein weiteres Problem ist, dass der Fossilbericht der Gavialoidea bereits mit longirostrinen Formen wie Eothoracosaurus (Oberkreide) einsetzt, während die Tomistominae erst im frühen Eozän (z. B. Kentisuchus) erscheinen, wobei sie zu diesem Zeitpunkt noch stenorostrin sind. Erst die geologisch jüngeren Vertreter dieser Unterfamilie sind longirostrin.[9]
Morphologischer Baum der rezenten Krokodile (nach Brochu, 1999, 2003):[10][11]
| Krokodile (Crocodylia) |
| ||||||||||||||||||
Molekularer Baum der rezenten Krokodile (nach Oaks, 2011):[5]
| Krokodile (Crocodylia) |
| ||||||||||||||||||
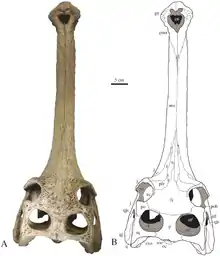

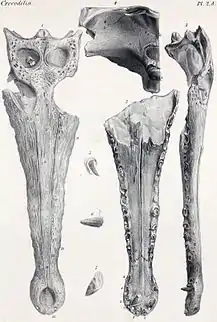





Bei Berücksichtigung fossiler Taxa werden die Gaviale in drei Unterfamilien gegliedert: Gavialinae, Gryposuchinae und Tomistominae. Die Gavialinae, zu denen auch der rezente Gangesgavial gehört, wurden fossil hauptsächlich in Asien gefunden. Eogavialis aus dem Oligozän, stammt hingegen aus Afrika. Jedoch ist unsicher ob es sich tatsächlich um den frühesten Vertreter der Gavialinae handelt, oder ob Eogavialis, noch außerhalb der Gavialinen, an der Basis der Gavialiden steht. Vertreter dieser Gattung wurden auch in marinen Ablagerungen gefunden. Im Miozän erschien die Gattung Gavialis. Funde von Gavialis-Arten auf Inseln Indonesiens und Ozeaniens lassen vermuten, dass diese, anders als der Gangesgavial, auch in Salzwasser lebten oder es zumindest nicht scheuten.
Die Gryposuchinae haben keine rezenten Vertreter. Sie wurden bislang nur in Südamerika und der Karibik gefunden. Sie erschienen erstmals im Oligozän und lebten bis zum frühen Pliozän. Ihre Fossilien kommen dabei auch in Ablagerungen ehemaliger Küstengebiete vor. Gryposuchus croizati war mit über 10 Metern Länge die größte Spezies.
Fossile Tomistominae wurden in Afrika, Europa, Asien und Nordamerika gefunden. Die ältesten Vertreter stammen aus dem frühen Eozän. Auch bei den Tomistominae wird davon ausgegangen, dass sie auch im Salzwasser lebten und sich durch das Überqueren von Meeren ausgebreitet haben. Rhamphosuchus aus Indien war mit geschätzten 10–12 Metern die größte Gattung.
Folgende fossile Arten gehören, unter Einschluss der Tomistominae, zu den Gavialidae (nach Vélez-Juarbe, Brochu & Santos, 2007;[12] Brochu & Storrs, 2012[13] und Martin & al., 2012[14]):
- Familie: Gavialidae Adams, 1854
- Unterfamilie: Gavialinae Nopcsa, 1923
- Unterfamilie: † Gryposuchinae Vélez-Juarbe, Brochu & Santos, 2007
- Gattung: † Aktiogavialis Vélez-Juarbe, Brochu & Santos, 2007
- † Aktiogavialis puertoricensis Vélez-Juarbe, Brochu & Santos, 2007
- Gattung: † Gryposuchus Gürich, 1912
- † Gryposuchus neogaeus (Burmeister, 1885)
- † Gryposuchus jessei Gürich, 1912
- † Gryposuchus colombianus (Langston, 1965)
- † Gryposuchus croizati Riff & Aguilera, 2008
- Gattung: † Ikanogavialis Sill, 1970
- † Ikanogavialis gameroi Sill, 1970
- † Ikanogavialis papuensis (De Vis, 1905), Zuordnung umstritten
- Gattung: † Siquisiquesuchus Brochu & Rincón, 2004
- † Siquisiquesuchus venezuelensis Brochu & Rincón, 2004
- Gattung: † Piscogavialis Kraus, 1996
- † Piscogavialis jugaliperforatus Kraus, 1996
- Gattung: † Hesperogavialis Bocquentin-Villanueva & Buffetaut, 1981
- † Hesperogavialis cruxenti Bocquentin-Villanueva & Buffetaut, 1981
- Gattung: † Aktiogavialis Vélez-Juarbe, Brochu & Santos, 2007
- Unterfamilie: Tomistominae Kälin, 1955
- Gattung: † Ferganosuchus Efimov, 1982
- † Ferganosuchus planus Efimov, 1982
- Gattung: † Kentisuchus Mook, 1955
- Gattung: † Dollosuchus Swinton, 1937
- † Dollosuchus dixoni (Owen, 1850), Status unklar
- Gattung: † Dollosuchoides Brochu, 2007
- † Dollosuchoides densmorei Brochu, 2007
- Gattung: † Megadontosuchus Mook, 1955
- † Megadontosuchus arduini (De Zigno, 1880)
- Gattung: † Gavialosuchus Toula & Kail, 1885
- † Gavialosuchus eggenburgensis Toula & Kail, 1885
- Gattung: † Toyotamaphimeia Aoki, 1983
- † Toyotamaphimeia machikanensis (Kobatake & al., 1965)
- Gattung: Tomistoma Müller, 1846
- Gattung: † Maroccosuchus Jonet & Wouters, 1977
- † Maroccosuchus zennaroi Jonet & Wouters, 1977
- Gattung: † Thecachampsa Cope, 1867
- Gattung: † Penghusuchus Shan & al., 2009
- † Penghusuchus pani Shan & al., 2009
- Gattung: † Paratomistoma Brochu & Gingerich, 2000
- † Paratomistoma courti Brochu & Gingerich, 2000
- Gattung: † Rhamphosuchus Falconer & Cautley, 1840
- † Rhamphosuchus crassidens Falconer & Cautley, 1840
- Gattung: † Ferganosuchus Efimov, 1982
Einzelnachweise
- John Harshman, Christopher J. Huddleston, Jonathan P. Bollback, Thomas J. Parsons, Michael J. Braun: True and false gharials: a nuclear gene phylogeny of crocodylia. Systematic Biology. Bd. 52, Nr. 3, 2003, S. 386–402, doi:10.1080/10635150390197028
- Axel Janke, Anette Gullberg, Sandrine Hughes, Ramesh K. Aggarwal, Ulfur Arnason: Mitogenomic Analyses Place the Gharial (Gavialis gangeticus) on the Crocodile Tree and Provide Pre-K/T Divergence Times for Most Crocodilians. Journal of Molecular Evolution. Bd. 61, Nr. 5, 2005, S. 620–626, doi:10.1007/s00239-004-0336-9 (freier Volltext: Researchgate)
- Ray E. Willis, L. Rex McAliley, Erika D. Neeley, Llewellyn D. Densmore III: Evidence for placing the false gharial (Tomistoma schlegelii) into the family Gavialidae: Inferences from nuclear gene sequences. Molecular Phylogenetics and Evolution. Bd. 43, Nr. 3, 2007, S. 787–794, doi:10.1016/j.ympev.2007.02.005
- Ray E. Willis: Transthyretin Gene (TTR) Intron One Elucidates Crocodylian Relationships. Molecular Phylogenetics and Evolution. Bd. 53, Nr. 3, 2009, S. 1049–1054, PMC 2787865 (freier Volltext)
- Jamie R. Oaks: A time-calibrated species tree of Crocodylia reveals a recent radiation of the true crocodiles. Evolution. Bd. 65, Nr. 11, 2011, S. 3285–3297, doi:10.1111/j.1558-5646.2011.01373.x
- Christopher W. Walmsley, Peter D. Smits, Michelle R. Quayle, Matthew R. McCurry, Heather S. Richards, Christopher C. Oldfield, Stephen Wroe, Phillip D. Clausen, Colin R. McHenry: Why the Long Face? The Mechanics of Mandibular Symphysis Proportions in Crocodiles. PLoS ONE. Bd. 8, Nr. 1, 2013, e53873, doi:10.1371/journal.pone.0053873.
- Richard Owen: Odontography; or, a treatise on the comparative anatomy of the teeth; their physiological relations, mode of development, and microscopic structure, in the vertebrate animals. Volume I: Text. Hippolyte Baillière, London 1840–1845, S. 285 ff. (BHL)
- L. Rex McAliley, Ray E. Willis, David A. Ray, P. Scott White, Christopher A. Brochu, Llewellyn D. Densmore III: Are crocodiles really monophyletic? – Evidence for subdivisions from sequence and morphological data. Molecular Phylogenetics and Evolution. Bd. 39, Nr. 1, 2006, S. 16–32, doi:10.1016/j.ympev.2006.01.012.
- P. Piras, P. Colangelo, D.C. Adams, A. Buscalioni, J. Cubo, T. Kotsakis, C. Meloro, P. Raia: The Gavialis-Tomistoma debate: the contribution of skull ontogenetic allometry and growth trajectories to the study of crocodylian relationships. In: Evolution & Development. Bd. 12, Nr. 6, 2010, S. 568–579. doi:10.1111/j.1525-142X.2010.00442.x.
- Christopher A. Brochu: Phylogenetics, Taxonomy, and Historical Biogeography of Alligatoroidea. Society of Vertebrate Paleontology Memoir. Bd. 6 (Journal of Vertebrate Paleontology, Bd. 19, Supplementum Nr. 2), 1999, S. 9–100, doi:10.1080/02724634.1999.10011201
- Christopher A. Brochu: Phylogenetic Approaches Toward Crocodylian History. Annual Review of Earth and Planetary Sciences. Bd. 31, 2003, S. 357–397, doi:10.1146/annurev.earth.31.100901.141308
- J. Vélez-Juarbe, C. A. Brochu, H. Santos: A gharial from the Oligocene of Puerto Rico: transoceanic dispersal in the history of a non-marine reptile. In: Proceedings of the Royal Society B. Bd. 274, Nr. 1615, 2007, S. 1245–1254. doi:10.1098/rspb.2006.0455. PMID 17341454. PMC 2176176 (freier Volltext).
- C. A. Brochu, G. W. Storrs: A giant crocodile from the Plio-Pleistocene of Kenya, the phylogenetic relationships of Neogene African crocodylines, and the antiquity of Crocodylus in Africa. In: Journal of Vertebrate Paleontology. Bd. 32, Nr. 3, 2012, S. 587. doi:10.1080/02724634.2012.652324.
- J. E. Martin, E. Buffetaut, W. Naksri, K. Lauprasert, J. Claude: Gavialis from the Pleistocene of Thailand and Its Relevance for Drainage Connections from India to Java. In: PLoS ONE. Bd. 7, Nr. 9, 2012, S. e44541. doi:10.1371/journal.pone.0044541.
- Stéphane Jouve: A new basal tomistomine (Crocodylia, Crocodyloidea) from Issel (Middle Eocene; France): palaeobiogeography of basal tomistomines and palaeogeographic consequences. Zoological Journal of the Linnean Society. Bd. 177, Nr. 1, 2016, S. 165–182, doi:10.1111/zoj.12357