Flugsaurier
Die Flugsaurier (Pterosauria) sind ausgestorbene Reptilien, die etwa gleichzeitig mit den Nichtvogeldinosauriern lebten und wie die Dinosaurier zur Gruppe der Ornithodira innerhalb der Archosauria gehören. Sie waren durch ihre großen, tragflächenartigen Flughäute als erste Wirbeltiere in der Lage, aktiv zu fliegen. Die Vögel und Fledertiere entwickelten ihr Flugvermögen später und jeweils eigenständig.
| Flugsaurier | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Flugsaurier (Pterosauria) | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Karnium (Obertrias) bis Maastrichtium (Oberkreide) | ||||||||||||
| 235 bis 66 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Pterosauria | ||||||||||||
| Kaup, 1834 |
Die frühesten Funde von Flugsauriern reichen zurück bis in die untere Obertrias (Karnium) vor etwa 228 Millionen Jahren. Sie waren auf der Erde präsent bis zum großen Massenaussterben an der Wende von der Kreidezeit zum Paläogen (Kreide-Paläogen-Grenze) vor etwa 66 Millionen Jahren.
Bei den Flugsauriern werden zwei Typen unterschieden: Die „Langschwanzflugsaurier (Rhamphorhynchoidea)“, die sich durch einen langen Schwanz und relativ kurze Mittelhandknochen auszeichnen, waren die frühesten Formen und verschwanden im oberen Jura. Die Kurzschwanzflugsaurier (Pterodactyloidea), die keinen oder einen nur sehr kurzen Schwanz und lange Mittelhandknochen besaßen, erschienen im Mittleren Jura und starben zum Ende der Kreidezeit aus. Diese klassische Einteilung wird mittlerweile in der Taxonomie nicht mehr verwendet, da die Kurzschwanzflugsaurier von den Langschwanzflugsauriern abstammen. Die Langschwanzflugsaurier sind daher im Sinne der phylogenetischen Systematik eine paraphyletische Gruppe.
Lebensweise
Fortbewegung


Eine wichtige Frage zur Lebensweise stellt sich bezüglich der Flugfähigkeit der Flugsaurier. Dabei wird allgemein davon ausgegangen, dass gerade die kleinen Arten aktiv sehr gut fliegen konnten. Bei den großen Flugsauriern der Kreidezeit handelte es sich demgegenüber wahrscheinlich primär um Segelflieger, die wie die Greifvögel auf den Aufwind angewiesen waren, oder dynamische Gleitflieger, die wie die Albatrosse unterschiedlich schnelle horizontale Winde im Lee von Wellentälern nutzten, um mit möglichst wenig Energieaufwand weite Strecken hinter sich zu bringen. Für einen dynamischen Gleitflug spricht die maritime Lebensweise, da es über dem Meer keine Aufwinde gibt, und die hohe Flügelstreckung vieler Arten. Doch auch diese Tiere mussten zumindest aus dem Wasser heraus in der Lage sein, aktiv zu starten und Höhe zu gewinnen.
Gerade aufgrund des hohen Energieaufwandes, der zum Fliegen notwendig ist, geht man heute oft davon aus, dass die Tiere Warmblüter und zu einem großen Teil behaart waren. Dafür spricht auch der Fund des behaarten Sordes pilosus („Behaarter Teufel“) und haarähnliche Abdrücke bei anderen Flugsaurierfossilien aus dem Solnhofener Plattenkalk und dem Posidonienschiefer von Holzmaden. Dem widersprechen die britischen Paläontologen David M. Martill und David Unwin in einer im September 2020 veröffentlichten Studie. Ihrer Ansicht nach waren die Tiere weder behaart noch befiedert, sondern bei den als Haare oder Protofedern interpretierten Strukturen handelt es sich um Fasern aus den Flügelmembranen.[1]
Da die Hinterbeine bei den Flugsauriern nur sehr klein ausgebildet waren, stellt sich die Frage, wie die Tiere sich an Land fortbewegen konnten. Zur Beantwortung dieser Frage werden zwei alternative Theorien diskutiert. Nach Ansicht von Kevin Padian und Jeremy Raynor nutzten die Tiere ihre Beine wie die heutigen Vögel und gingen zweibeinig (biped). Die Autoren führen dies darauf zurück, dass die Beine ähnlich gebaut sind wie die der heutigen Vögel und entsprechend wohl auch ähnlich eingesetzt wurden. David Unwin und Peter Wellnhofer gehen stattdessen davon aus, dass die Flugsaurier vierbeinig (quadruped) gingen und ihre Arme dabei einsetzten (wie die heutigen Vampirfledermäuse), wobei der Flügelfinger nach hinten eingeschlagen wurde.[2] Sie nehmen dabei an, dass die Flughäute auch die Hinterbeine mit umfassten. Die letztere Theorie ist die aktuell bevorzugte. Untersuchungen der Beckenstruktur von Anhanguera und Campylognathoides haben gezeigt, dass die Flugsaurier die Hinterbeine nicht senkrecht unter den Körper stellen konnten[3] und besonders die großen Flugsaurier sich nur sehr schwerfällig allein auf den Hinterbeinen fortbewegt haben können.
Möglich ist zudem, dass unterschiedliche Arten hier unterschiedliche Lösungen nutzten. So ist es leicht, sich eine der kleineren langschwänzigen Formen auf den Hinterbeinen vorzustellen, jedoch eher schwer bei den großen und größten Formen der ausgehenden Kreidezeit.
Ernährung, Lebensraum und Fortpflanzung

Die Flugsaurier waren wahrscheinlich durchweg Jäger und Fleischfresser. Dabei jagten die frühen Langschwanzflugsaurier vor allem nach Wirbellosen, später entwickelte sich eine größere Diversität in der Ernährung, und auch Fischfresser und Jäger von Landwirbeltieren traten auf. Dimorphodon ernährte sich wahrscheinlich von Wirbeltieren, Austriadactylus war auf hartschalige Wirbellose spezialisiert, Coloborhynchus und Rhamphorhynchus waren Fischfresser und Pterodactylus ernährte sich von verschiedenen Wirbellosen.[4] Ein sehr ungewöhnlicher Flugsaurier namens Pterodaustro aus Südamerika hatte sehr lange und extrem dünne Zähne, die eine Reuse darstellten. Er filterte hiermit Meerwasser und ernährte sich von Plankton wie die heutigen Flamingos.
Obwohl die meisten Fossilien von Flugsauriern bislang von Lebensräumen an größeren Gewässern stammen, ist es heute unbestritten, dass viele Arten auch im Inland, etwa in Wäldern, Bergen oder sogar Wüsten lebten. Der fehlende Fossilbefund ist auf die sehr schlechten Fossilisierungsbedingungen dieser Lebensräume und die sehr dünnen Knochen der Tiere zurückzuführen.
Flugsaurier waren wie alle Archosaurier eierlegende Tiere. Ein im Jahre 2004 publizierter Fund eines fossilen Embryos aus der frühen Kreidezeit vor etwa 121 Millionen Jahren, der aus der Liaoning-Provinz Nordostchinas stammt, belegt dies eindrücklich. Das gefundene Ei war etwas kleiner als ein Hühnerei und enthielt ein gut erhaltenes Skelett, daneben waren auch Eindrücke von Haut- und Flügelgewebe zu erkennen. Die bei dem Tier, das vermutlich kurz vor dem Schlüpfen stand, bereits vorhandene Flügelspanne von 27 Zentimetern wird von den Entdeckern als Hinweis darauf gewertet, dass die Jungtiere wahrscheinlich Frühentwickler waren, die bereits kurz nach dem Schlüpfen fliegen und sich unabhängig von ihren Eltern ernähren konnten.[5][6]
Dass Flugsaurier in Kolonien zusammenlebten, basiert dagegen in erster Linie auf theoretischen Annahmen sowie auf einem Fund in der Atacama-Wüste in Chile, wo auf sehr engem Raum sehr viele Überreste junger Flugsaurier gefunden wurden.
Anatomie
Anatomie der Flügel
Das auffälligste Merkmal der Flugsaurier sind die zu großen Schwingen umgestalteten Vorderbeine, die den Tieren sowohl den Gleitflug als auch den aktiven Flug ermöglicht haben. Dabei spannte sich die Flughaut zwischen dem Körper der Tiere und dem Arm mit dem stark verlängerten vierten Finger der Tiere, der dem Ringfinger beim Menschen entspricht, auf. Die anderen drei Finger bildeten Krallen außerhalb der Flughaut, der fünfte Finger fehlte.
Die Flügelknochen der Flugsaurier waren in der Regel hohl und sehr dünnwandig. Sie enthielten viele luftgefüllte Bereiche, damit das Gewicht der Knochen nicht zu hoch wurde. Durch Knochenbälkchen wurden vor allem die Enden der größeren Knochen verstärkt. In der Regel werden aus diesem Grunde nur Fragmente der Flügelknochen gefunden, intakte Knochen sind äußerst selten. Die Flügelknochen der beiden Flugsaurierformen unterscheiden sich dabei deutlich.
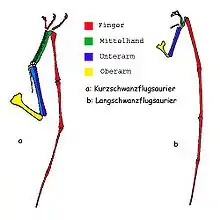
Der Flügel der Langschwanzflugsaurier besteht aus einem recht kurzen Oberarmknochen (Humerus) und einem relativ kurzen Mittelhandknochen. Die vier Fingerknochen sind dagegen recht lang. Die Krallen sitzen ebenfalls an sehr kurzen Mittelhandknochen. Der Unterarm besteht wie bei allen Landwirbeltieren aus zwei Knochen, wobei die Elle (Ulna) etwas länger ist als die Speiche (Radius). Im Querschnitt sind beinahe alle Knochen im Schaftbereich oval, während sie zu den Enden herzförmig werden.
Im Gegensatz dazu sind bei den Kurzschwanzflugsauriern sowohl der Oberarm als auch die Elle massiver und länger, die Speiche ist ebenso lang wie die Elle, nur weniger dick. Besonders die Mittelhandknochen sind sehr viel länger als die der früheren Flugsaurierformen, während entsprechend die Finger in Relation zu den anderen Knochen kürzer werden. Auch Kurzschwanzflugsaurier hatten meist vier Fingerknochen. Die Krallen sind sehr gut ausgebildet und enden ebenfalls an verlängerten Mittelhandknochen. Die Knochenwand ist dünner als die der Langschwanzflugsaurier und verdickt sich zum Ende der Knochen. Der Querschnitt ist hohl und dreieckig und wird zum Ende hin oval. Besonders auffällig sind diese Merkmale bei den sehr großen Formen.
Die Flügel selbst bestanden aus einer Hautstruktur, die durch eng beieinander liegende Fasern, so genannte Aktinofibrillen verstärkt wurde und so zum einen für eine Stabilität des Flügels und zum anderen für eine Rissfestigkeit sorgten. Diese Strukturen konnten an den gut erhaltenen Flughäuten einiger Flugsaurier aus der Santana-Formation untersucht werden.
Wirbelsäule
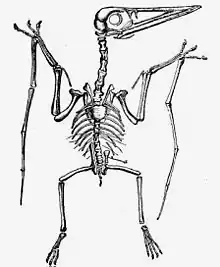


Die Wirbelsäule der Flugsaurier unterscheidet sich von den früheren Reptilien deutlich und ist der fliegenden Lebensweise und den damit verbundenen Ansprüchen an das Achsenskelett angepasst. In einigen Bereichen ähnelte sie sehr stark der Wirbelsäule der Vögel (Konvergenz) mit einer ausgeprägten Schulterregion und einer massiven Region im Bereich des Beckens. Der Rücken war nur sehr kurz und erlaubte entsprechend nur wenige Bewegungen, den längsten Bereich stellte die Halswirbelsäule dar, entsprechend sind die meisten Einzelfunde von Wirbeln Halswirbel.
Die Halswirbelsäule ist im Vergleich mit anderen Wirbeltieren sehr lang. Der Hals besteht dabei aus acht Wirbeln bei den frühen Flugsauriern, während einige der größeren Spätformen die Anzahl auf sechs reduziert haben. Die beiden ersten Wirbel dieses Abschnitts, der Atlas und der Axis, waren meistens zu einem einzigen Wirbelknochen verschmolzen. Die Knochenwand der Wirbel war dünn, und der Innenraum bestand aus einem schwammigen Knochengebälk mit großen Lufträumen. Halsrippen existierten im Normalfall nicht.
Die Brustwirbelsäule bestand aus bis zu zwölf Wirbeln, wobei die meisten Arten weniger besaßen. Die ersten Wirbel waren meist zu einer massiven Schulterstruktur verwachsen, an der die Schulterblätter (Scapulae) ansetzten. Dies bildete bei einigen Arten einen langen Knochen, der über sechs bis acht Wirbel reichte.
Die Rückenwirbelsäule bestand aus etwa sechs Wirbelkörpern und war sehr kurz, aber kompakt. Wie alle anderen Wirbelknochen waren sie auch mit einem Gebälk gefüllt, sie wiesen jedoch außerdem noch seitlich ein großes Loch auf, welches den Knochen vollständig durchdrang. Bei den meisten Arten trugen diese Wirbel kurze Bauchrippen.
Die Sakral- oder Lendenwirbelsäule war normalerweise zu einem massiven Knochen verschmolzen, mit dem die Beckenknochen in einer engen Verbindung standen. Bei den frühen Langschwanzflugsauriern war diese Struktur noch relativ locker, bei den Kurzschwanzflugsauriern handelte es sich jedoch um einen einzelnen Beckenknochen. In diesem Bereich befanden sich in der Regel sechs bis acht Wirbelknochen.
Die Schwanzwirbelsäule war bei den Langschwanzflugsauriern sehr lang und konnte aus mehr als 35 Schwanzwirbeln bestehen. Bei einigen dieser Flugsaurierfossilien findet sich am Ende des Schwanzes ein Segelabdruck, dieses diente wahrscheinlich der Navigation und Stabilisierung im Flug. Bei den Kurzschwanzflugsauriern wurde die Anzahl der Schwanzwirbel auf eine geringe Zahl reduziert.
Schädel und Zähne
Der Schädel der Flugsaurier ähnelte wie andere Teile des Skeletts dem der Vögel, vor allem durch die verschiedenen Einsparungen und Reduzierungen, die den Schädel leichter machten. Der wesentliche Unterschied betrifft jedoch die Bezahnung, die besonders bei den frühen Flugsauriern sehr gut ausgebildet war. Die massivsten Strukturen und damit diejenigen, die am häufigsten unter Fossilienfunden auftreten, waren die Kieferspitzen sowie der Kieferansatz am Quadratum.
Ein wesentliches Merkmal der Flugsaurier ist ein Ring aus Knochenplatten, der um das Auge lag und dessen biologischer Sinn bislang nicht eindeutig geklärt werden konnte. Wahrscheinlich diente er als Druckausgleichselement. Des Weiteren besaßen die Tiere wie alle anderen Archosaurier ein drittes Schädelfenster vor den Augen (Präfrontales Fenster), während sie das Schläfenfenster reduziert hatten. Viele der Kurzschwanzflugsaurier wiesen hohe und lange Schädelkämme auf, die je nach Interpretation der Schallverstärkung und/oder der Flugstabilisierung gedient haben.

Die Kiefer trugen keine Zähne.
Auch die Zähne sind sehr oft fossil erhalten, allerdings gab es besonders in der späten Kreidezeit eine Reihe von Arten, die keine oder nur sehr kleine Zähne aufwiesen. Die frühesten Flugsaurier hatten mehrspitzige Zähne (in der Regel mit drei Spitzen: Tricuspide Zähne), die etwa bei Rhamphorhynchus von langen und spitzen Zähnen abgelöst wurden. Diese Zähne standen in den langen und dünnen Kiefern über den Mundrand hinaus. Einige der Langschwanzflugsaurier besaßen allerdings auch gar keine Zähne.
Während des oberen Jura nahm die Anzahl der Zähne bei den Flugsauriern ab. Einige Arten behielten noch Zähne am vorderen Ende der Kiefer, andere verloren alle Zähne, wodurch ein schnabelähnlicher Kiefer entstand. Die großen Flugsaurier der späten Kreide hatten durchweg keine Zähne.
Entdeckungsgeschichte
Wie die Dinosaurier gehörten auch die Flugsaurier zu den Tiergruppen, die bei ihrer ersten Entdeckung eine enorme Faszination auslösten. Fossilien der Flugsaurier gehören zu den ersten gefundenen Fossilien von Landwirbeltieren überhaupt und wurden entsprechend bereits zu Beginn der Paläontologiegeschichte im frühen 19. Jahrhundert untersucht. Die erste Beschreibung eines Flugsauriers stammt aus dem Jahr 1784 von Cosimo Alessandro Collini. Besonders gut erhaltene Fossilfunde aus Südengland und Deutschland machten die Flugsaurier sowohl innerhalb der wissenschaftlichen Kreise als auch beim Volk populär. Unter den ersten Funden befanden sich Dimorphodon macronyx, der von Mary Anning in der „Liasformation“ bei Lyme Regis entdeckt wurde,[7] sowie Rhamphorhynchus und Pterodactylus aus dem späten Jura, welche im Plattenkalk von Solnhofen gefunden wurden. Die Funde führten dazu, dass sie etwa als most wondrous creatures of the prehistoric world beschrieben wurden. Der deutsche Naturforscher Georg August Goldfuß vertrat 1831 die Ansicht, dass diese Tiere wohl eher aussähen, als seien sie der Fantasie denn der Natur entsprungen.
Im Laufe des 19. und des 20. Jahrhunderts stieg die Anzahl der Fossilfunde, und vor allem die Artenvielfalt der Gruppe wurde mehr und mehr offensichtlich. Besonders die Plattenkalke in Solnhofen und Eichstätt stellten sich als sehr reiche Lagerstätten fossiler Flugsaurier heraus. So wurde hier der sehr gut erhaltene Rhamphorhynchus longicephalus 1839 als erster Flugsaurier mit einem langen und vollständigen Schwanz entdeckt. In den gleichen Schichten entdeckte man daneben auch kurzschwänzige Tiere, die offensichtlich gleichzeitig gelebt haben müssen. In England war vor allem die Wealden-Formation reich mit Fossilien der Tiere, welche von Gideon Mantell erst als Vögel beschrieben wurden (1822, 1827 und 1837) und deren Zugehörigkeit zu den Flugsauriern Owen 1846 erkannte. Seeley ordnete sie der Gattung Ornithocheirus zu. 1870 entdeckte Othniel Charles Marsh in Kansas Überreste des riesigen Pteranodon und damit des ersten Flugsaurierskelettes in Nordamerika. 1895 führte Felix Plieninger die Unterteilung der Flugsaurier in Langschwanzflugsaurier und Kurzschwanzflugsaurier ein, die erst in den letzten beiden Jahrzehnten durch phylogenetische Untersuchungen revidiert wurde.
Auch im Verlauf des 20. Jahrhunderts nahmen die Fossilfunde und der bekannte Formenreichtum zu. Zu den klassischen Fundstätten kamen weitere hinzu, die eine weltweite Verbreitung der Tiere dokumentieren. Heute liegen etwa besonders ergiebige Fundstellen in der Santana-Formation in Brasilien, in Paki im Senegal und in Nordchina.
Flugsaurier der Trias
Die ältesten Flugsaurier stammen aus der Obertrias und haben entsprechend vor etwa 228 Millionen Jahren gelebt. Von den meisten Vertretern aus dieser Zeit sind allerdings nur Knochenfragmente gefunden worden, vollständige Schädel oder Skelette sind äußerst selten. Die frühen Arten lassen sich vor allem an ihren Zähnen erkennen, die mit mehreren Spitzen ausgestattet sind. Die Zähne späterer Arten haben nur eine Spitze und sind langoval. Des Weiteren sind die Knochen der frühen Flugsaurier massiver und haben eine dickere Außenschicht, während das Innere weniger Freiräume besitzt.
|
Flugsaurier aus der Trias:
|
|---|
Alle Flugsaurier der Trias und des frühen Jura hatten lange Schwänze, zudem zeichnen sich die meisten frühen Flugsaurier durch sehr lange Zähne aus, wie etwa Rhamphorhynchus aus dem Solnhofener Plattenkalk.
Das wahrscheinlich am besten erhaltene Skelett eines frühen Flugsauriers stellt Eudimorphodon ranzi dar. Das Exemplar stammt aus dem Kalkstein von Cene in der Nähe der norditalienischen Stadt Bergamo. Im Bereich des Magens wurden sogar noch Reste der letzten Mahlzeit in Form von Fischschuppen gefunden. Offensichtlich war diese Art ein Fischfresser.
Die Eudimorphodontidae spielen zu Beginn der Evolution der Flugsaurier eine wesentliche Rolle, aus ihnen entwickelten sich bereits in der Trias die ersten Vertreter der Dimorphodontidae und der Rhamphorhynchidae. Diese beiden Taxa sind auch in der Jurazeit noch präsent, während die Eudimorphodontidae zum Ende der Trias verschwunden sind.
Die meisten und die vollständigsten Funde der Trias stammen aus Italien: aus dem Zorzino-Kalkstein bei Cene in der Lombardei und aus dem Preone-Tal in den Alpen.
Flugsaurier des Jura
In der Jurazeit kam es bei den Flugsauriern zu großen evolutionären Veränderungen. Sie breiteten sich weltweit aus und besiedelten viele verschiedene Lebensräume. Die Langschwanzflugsaurier dieser Zeit hatten meist lange Zähne, die sich zur Spitze hin stark verschmälern. Bei einigen Arten waren die Zähne gekerbt, die meisten hatten jedoch glatte Zähne.
|
Flugsaurier aus dem Jura
|
|---|
Die Langschwanzflugsaurier verschwanden etwa zur Mitte des Jura. Sie wurden ersetzt durch die zu Beginn des Jura aus einer Teilgruppe von ihnen entstandenen Kurzschwanzflugsaurier oder Pterodactyloidea. Diese hatten den Schwanz weitgehend reduziert und zeigen in ihrer Evolution eine Verkürzung der Zähne und eine Verlängerung der Mittelhandknochen am flughauttragenden Finger.
Im Laufe des Jura entwickelten sich die Flugsaurier zu einer sehr erfolgreichen Wirbeltiergruppe. In der frühen und mittleren Jurazeit entstanden ausschließlich aus den bereits in der Trias bekannten Dimorphodontidae und Rhamphorhynchidae mit den Scaphognathinae, Campylognathoides und Rhamphocephalidae weitere Taxa der Langschwanzflugsaurier.

Erst im späteren Jura spielten die ungeschwänzten Kurzschwanzflugsaurier, vor allem repräsentiert durch die Pterodactylidae, eine große Rolle. Zu dieser Zeit kam es zur größten Artenvielfalt der Flugsaurier insgesamt, sie besiedelten die verschiedensten Lebensräume in allen Teilen der Welt.
Eine sehr reichhaltige Lagerstätte für Flugsaurier des Jura bildet der Plattenkalk bei Eichstätt und Solnhofen, in dem auch der Urvogel Archaeopteryx gefunden wurde. Die Bedingungen zur Fossilisation in diesem Material sind ausgesprochen gut, so dass sogar einige Flughäute und Weichgewebeabdrücke erhalten sind. Unter den Funden in diesen Plattenkalken sind etwa die Skelette von Rhamphorhynchus und Pterodactylus.
Ebenfalls als exzellente Lagerstätte des Jura werden die Qaratai- oder Karatau-Berge in Kasachstan angesehen. Hier wurde zum Beispiel der offensichtlich behaarte Sordes pilosus mit konservierten Flügelmembranen gefunden.
Flugsaurier der Kreidezeit
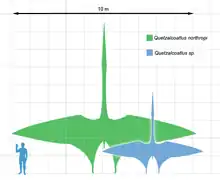
In der Kreidezeit entwickelten sich die größten Formen der Flugsaurier, gleichzeitig entstanden auch Riesenformen der Dinosaurier und anderer Tiergruppen (Plesiosaurier, Mosasaurier, Ichthyosaurier). Wie bereits dargestellt, handelt es sich bei diesen Flugsauriern ausschließlich um Kurzschwanzflugsaurier.
|
Flugsaurier aus der Kreide
|
|---|
Die großen Flugsaurier waren dabei überall in der Welt anzutreffen, allerdings gab es auch weiterhin kleinere Formen. Diese kleineren Flugsaurier standen wahrscheinlich in direkter Konkurrenz mit den sich neu entwickelnden Vögeln, die sich zu dieser Zeit rasch ausbreiteten.
Bis in die 1970er Jahre galt Pteranodon aus Nordamerika als größter Flugsaurier, seine Flügelspannweite erreichte etwa acht Meter. Er wurde durch den Fund des Quetzalcoatlus (benannt nach dem Aztekengott Quetzalcoatl, der u. a. in Gestalt einer gefiederten Schlange dargestellt wurde) übertroffen, der nach Hochrechnung seiner Schädelmaße und einem Vergleich mit anderen Flugsauriern eine Spannweite von bis zu 12 Metern erreicht haben könnte. Ähnliche Ausmaße erreichte Hatzegopteryx aus der europäischen Oberkreide, dessen im rumänischen Hațeg-Becken entdeckte Skelettfragmente erst Ende der 1990er Jahre als Fossilien eines Flugsauriers identifiziert wurden.
Eine sehr wichtige Lagerstätte für Flugsaurierfossilien der oberen Kreide ist das Araripe-Becken in Brasilien und hier vor allem die als Crato-Formation und Santana-Formation bekannten Schichten. Aus diesem Gebiet stammen sehr unterschiedliche Flugsaurier, darunter etwa Anhanguera, Criorhynchus, Ornithocheirus, Tapejara, Thalassodromeus, Ludodactylus und Tupuxuara.
Eine weitere bedeutende Fundstätte für Flugsaurier der Kreidezeit ist Xinjiang in China, wo auch wichtige Dinosaurier- (darunter einige gefiederte Dinosaurier) und Säugetierfunde (etwa Hadrocodium wui) gemacht wurden. Unter den Flugsauriern stammen etwa Dsungaripterus wei und Noripterus complicidens aus dieser Region, die besonders durch ihre kräftigen Kiefer auffallen. Ähnlich wie in Solnhofen und in der Crato-Formation, zeichnen sich die Funde aus der Jehol-Gruppe in Liaoning durch „Weichteilerhaltung“ aus. Die chinesischen Arten ähneln sowohl Funden aus Solnhofen als auch aus Brasilien. Die bisherigen Funde aus China lassen eine ähnliche, wenn nicht noch größere Diversität als dort vermuten.
Als reiche Flugsaurierregion gelten zudem der Niobrara-Kalk und andere Gesteine in Texas und Kansas, in denen einige der größten der bislang bekannten Flugsaurier entdeckt wurden, etwa Pteranodon ingens und Quetzalcoatlus. Die Knochen dieser Tiere sind oft sehr zerbrechlich und wurden weit verstreut gefunden.
Eine erst in jüngster Zeit erschlossene Lagerstätte im Hinblick auf Flugsaurier-Fossilien ist das Ouled-Abdoun-Becken im nördlichen Marokko (rund 70 km von Casablanca entfernt). Das Gelände ist für seine reichhaltigen und industriell verwertbaren Phosphatablagerungen bekannt, die während der Übergangszeit von der Kreide zum Paläogen entstanden. In diesen Schichten wurden von einzelnen Knochen bis hin zu Teilskeletten über 200 Pterosaurierfragmente gefunden, die zu 3 Familien (Pteranodontidae, Nyctosauridae und Azhdarchidae) mit insgesamt 7 Arten gehören. Sämtliche Funde stammen aus dem späten Maastrichtium mit einer zeitlichen Nähe zur Kreide-Paläogen-Grenze von höchstens einer Million Jahre. Dies deutet darauf hin, dass die Artenvielfalt der Pterosauria zumindest in dieser Region bis zum Ende der Kreide im Wesentlichen gewahrt blieb.[8]
Evolution der Flugsaurier
Die Flugsaurier stammen von frühen Archosauriern ab, die auch die Vorfahren der Krokodile und der Dinosaurier waren. Der Fossilbefund der Abstammungslinie der Flugsaurier fehlt, entsprechend konnte diese Abstammung nur durch einen Merkmalsvergleich der unterschiedlichen Tiergruppen nachgewiesen werden. Als Hauptindiz kann der Schädelbau der Flugsaurier angesehen werden. Dieser besitzt wie der anderer Archosaurier ein drittes Schädelfenster vor dem Auge (Präorbitales Fenster) und das Scheitelfenster (Foramen parietale) ist reduziert. Als weitere Anhaltspunkte zur Zuordnung besitzen die Flugsaurier typische Archosaurierzähne, und die fünfte Zehe ist reduziert. Innerhalb der Archosaurier stellen die Flugsaurier wahrscheinlich die Schwestergruppe der Dinosaurier dar, welche beide gemeinsam in das Taxon Ornithodira aufgrund des gleichen Aufbaus der Fußgelenke gestellt werden. Das Schwestergruppenverhältnis ist allerdings nicht so gut begründet und schlüssige gemeinsame Merkmale fehlen. Der aktuell akzeptierte Stammbaum entspricht also der folgenden Grafik:[9]
| Diapsiden |
| ||||||||||||||||||||||||

Als nächster Verwandter der Flugsaurier und zugleich der Dinosaurier wird ein Fossil aus dem schottischen Lossiemouth diskutiert. Dieses Tier, benannt als Scleromochlus taylori, teilt mit beiden Gruppen einige anatomische Merkmale. Aufschlussreich könnte es vor allem für die Evolution des Fliegens bei den Flugsauriern sein, da angenommen wird, dass Scleromochlus taylori ein guter Springer war, der vielleicht sogar eine Vorstufe der Flughäute der Flugsaurier besaß. Bereits 1914 schrieb Huene über Scleromochlus: „ … it was a parachuting animal, which had skin duplications on the forelimbs, perhaps also in other places.“ Er nahm an, dass das Tier in den Bäumen lebte und diese als Absprungpunkt für seinen Gleitflug nutzte (arboreale Hypothese).
Präzisiert wird die Theorie der Evolution der Flugsaurier aus Scleromochlus-ähnlichen Vorläufern durch Michael Benton 1999, der weitere Optionen der Lebensweise von Scleromochlus taylori darstellt und vor allem eine sehr detaillierte Merkmalsanalyse des Tieres bietet. Im Gegensatz zu Huene favorisiert er die von Woodward 1907 aufgestellte Theorie, dass Scleromochlus zwar ein Springer war, jedoch nicht Bäume als Absprungpunkt nutzte, sondern ein Bodenläufer mit gut ausgeprägtem Sprungvermögen war (saltatorische Hypothese).

Demnach soll sich aus dieser Art des Springers der erste fliegende Flugsaurier entwickelt haben.[10] Bislang ist nicht geklärt, welche der beiden Theorien zutreffend ist, spannend ist jedoch, dass für die Evolution des Fluges bei den Vögeln die gleichen Theorien aufgestellt wurden.
Ende 2020 wurden die Lagerpetidae, kleine, auf zwei Beinen laufenden Reptilien aus der Trias, als Vorfahren der Flugsaurier ermittelt. Beide Gruppen besitzen ein gleich geformtes Innenohr und verfügten wahrscheinlich über einen ähnlichen, sehr gut entwickelten Gleichgewichtssinn.[11]
Systematik


Die klassische Zweiteilung der Flugsaurier wurde durch phylogenetische Untersuchungen der letzten 25 Jahre aufgelöst. Demnach stellen die Kurzschwanzflugsaurier tatsächlich eine natürliche systematische Einheit dar, sind jedoch eingebettet in die verschiedenen Gruppen, die bislang zu den Langschwanzflugsauriern zusammengefasst wurden. Es wurden eine Reihe von Familien und anderer Untertaxa beschrieben, die jedoch meist schlecht definiert sind. Im Folgenden wird ein Kladogramm einer möglichen inneren Systematik aus einem Standardwerk von David M. Unwin wiedergegeben:[12]
| Flugsaurier |
| |||||||||||||||||||||||||||||||||||||||||||||||||||
Eine vollständige Systematik mit allen Gattungen findet sich unter Systematik der Flugsaurier.
Literatur
- Éric Buffetaut: Flugsaurier – Warmblütige Pioniere der Lüfte. In: Spektrum der Wissenschaft. Nr. 11, 2004, ISSN 0170-2971, S. 24–31.
- Eric Buffetaut, Jean-Michel Mazin (Hrsg.): Evolution and Palaeobiology of Pterosaurs (= The Geological Society. Special Publication. Band 217). The Geological Society, London 2003, ISBN 1-86239-143-2.
- Michael Fastnacht: The first dsungaripterid pterosaur from the Kimmeridgian of Germany and the biomechanics of pterosaur long bones. In: Acta Palaeontologica Polonica. Band 50, Nr. 2, 2005, ISSN 0567-7920, S. 273–288, online.
- Hartmut Haubold: Literaturbericht: Pterosauria 2004–2008 (= Zentralblatt für Geologie und Paläontologie. Teil 2: Paläontologie. Jg. 2009, Heft 1/2, ISSN 0044-4189). Schweizerbart, Stuttgart 2009.
- David W. E. Hone, Eric Buffetaut (Hrsg.): Flugsaurier. Pterosaur papers in honour of Peter Wellnhofer (= Zitteliana. Reihe B: Abhandlungen der Bayerischen Staatssammlung für Paläontologie und Geologie. Band 28, ISSN 1612-4138). Bayerische Staatssammlung für Paläontologie und Geologie, München 2008.
- David M. Unwin: The Pterosaurs. From Deep Time. PI Press, New York NY 2006, ISBN 0-13-146308-X.
- Peter Wellnhofer: Pterosauria (= Handbuch der Paläoherpetologie. Band 19). Pfeil u. a., München u. a. 1978, ISBN 3-437-30269-8.
- Peter Wellnhofer: Die große Enzyklopädie der Flugsaurier. Illustrierte Naturgeschichte der fliegenden Saurier. 100 Arten. Mosaik-Verlag, München 1993, ISBN 3-576-10174-8.
- Peter Wellnhofer: Illustrated Encyclopedia of Pterosaurs. Crescent Books, New York NY 1991, ISBN 0-517-03701-7.
Weblinks
- Pterosaur.net (englisch)
- The Pterosaur Database (englisch)
- Pterosaur Site (deutsch)
- Pterosauria Translation and Pronunciation Guide (Memento vom 13. Oktober 2011 im Internet Archive)
- The Pterosaur Species List (Memento vom 30. September 2011 im Internet Archive) (englisch, PDF, 789 kB)
Fußnoten
- David M. Unwin, David M. Martill. No protofeathers on pterosaurs. Nature Ecology & Evolution, 2020; DOI: 10.1038/s41559-020-01308-9
- Animiertes Drahtgittermodell von Anhanguera santanae (Gehen)
- Peter Wellnhofer: Illustrated Encyclopedia of Pterosaurs. 1991, S. 76–77 und 126.
- Jordan Bestwick, David M. Unwin, Richard J. Butler und Mark A. Purnell. 2020. Dietary Diversity and Evolution of the earliest Flying Vertebrates revealed by Dental Microwear Texture Analysis. Nature Communications. 11, 5293. DOI: 10.1038/s41467-020-19022-2
- Xiaolin Wang, Zhonghe Zhou: Palaeontology: Pterosaur embryo from the Early Cretaceous. In: Nature. Band 429, Nr. 6992, 2004, S. 621, doi:10.1038/429621a.
- David Michael Unwin, D. Charles Deeming. Prenatal development in pterosaurs and its implications for their postnatal locomotory ability. Proceedings of the Royal Society B: Biological Sciences, 2019; 286 (1904): 20190409 DOI: 10.1098/rspb.2019.0409
- seinerzeit noch unter dem Namen Pterodactylus macronyx beschrieben, siehe William Buckland: On the discovery of a new species of pterodactyle in the Lias at Lyme Regis. In: Transactions of the Geological Society of London. 2. Serie, Band 3, Teil 1, 1829, S. 217–222 (BHL)
- Nicholas R. Longrich, David M. Martill, Brian Andres: Late Maastrichtian pterosaurs from North Africa and mass extinction of Pterosauria at the Cretaceous-Paleogene boundary. In: PLOS Biology. März 2018. doi:10.1371/journal.pbio.2001663.
- Klassifikation der Reptilien in: Michael J. Benton: Paläontologie der Wirbeltiere. Übersetzung der 3. englischen Auflage durch Hans-Ulrich Pfretzschner. Pfeil, München 2007, ISBN 978-3-89937-072-0, S. 419 f.
- Michael J. Benton: Scleromochlus taylori and the origin of the pterosaurs. In: Philosophical Transactions of the Royal Society London. Series B: Biological Sciences. Band 354, Nr. 1388, 1999, ISSN 0080-4622, S. 1423–1446, doi:10.1098/rstb.1999.0489, Digitalisat (PDF; 576,33 kB).
- Martín D. Ezcurra, Sterling J. Nesbitt, Mario Bronzati, Fabio Marco Dalla Vecchia, Federico L. Agnolin, Roger B. J. Benson, Federico Brissón Egli, Sergio F. Cabreira, Serjoscha W. Evers, Adriel R. Gentil, Randall B. Irmis, Agustín G. Martinelli, Fernando E. Novas, Lúcio Roberto da Silva, Nathan D. Smith, Michelle R. Stocker, Alan H. Turner, Max C. Langer. Enigmatic dinosaur precursors bridge the gap to the origin of Pterosauria. Nature, 2020; DOI: 10.1038/s41586-020-3011-4
- David M. Unwin: The Pterosaurs. From Deep Time. 2006, S. 228.
- Junchang Lü, David M. Unwin, Xingsheng Jin, Yongqing Liu, Qiang Ji: Evidence for modular evolution in a long-tailed pterosaur with a pterodactyloid skull. In: Proceedings of the Royal Society. Series B: Biological Sciences. Band 277, Nr. 1680, 2010, ISSN 0950-1193, S. 383–389, doi:10.1098/rspb.2009.1603.
{kind=link}