Knochenfische
Knochenfische (Osteichthyes, von altgriechisch ὀστέον ostéon „Knochen“ und ἰχθύς ichthýs „Fisch“) oder Knochenfische im weiteren Sinne sind nach traditionellem Verständnis diejenigen Fische, deren Skelett im Gegensatz zu dem der Knorpelfische (Chondrichthyes) vollständig oder teilweise verknöchert ist. Von den Osteichthyes sind die Knochenfische im engeren Sinne, die Echten Knochenfische (Teleostei), als untergeordnetes Taxon zu unterscheiden.
| Knochenfische | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Veraltete systematische Gruppe Das hier behandelte Taxon ist nicht Teil der in der deutschsprachigen Wikipedia dargestellten Systematik. Näheres hierzu findet sich im Artikeltext. | ||||||||||||||

Dunkler Riesenzackenbarsch (Epinephelus lanceolatus) | ||||||||||||||
| Systematik | ||||||||||||||
| ||||||||||||||
| Wissenschaftlicher Name | ||||||||||||||
| Osteichthyes | ||||||||||||||
| Huxley, 1880 | ||||||||||||||
Die Knochenfische gliedern sich in zwei Großgruppen: die Strahlenflosser (Actinopterygii) und die Fleischflosser (Sarcopterygii). Aus der Sicht der modernen Systematik (Kladistik) gehören aber auch die Landwirbeltiere (Tetrapoda) zu den Fleischflossern und damit zu den Knochenfischen.[1] In der traditionellen Systematik hingegen zählt man die Landwirbeltiere nicht zu den Knochenfischen. Die Knochenfische im klassischen Sinne sind demnach keine natürliche Verwandtschaftsgruppe (kein monophyletisches Taxon). Das klassische Taxon „Osteichthyes“ wird daher in der zoologischen und paläontologischen Systematik immer weniger verwendet, und der Taxonname dient nur mehr als informeller Sammelbegriff für Fische mit knöchernem Skelett oder mit knorpeligem aber ursprünglich knöchernem Skelett (z. B. Störe und Mondfische).
Allgemeines
Mit Ausnahme der Neunaugen gehören alle in Binnengewässern Mitteleuropas lebenden Fischarten zu den Knochenfischen und – bis auf die Störartigen – zu den Echten Knochenfischen.
Die Knochenfische sind die artenreichste Gruppe der Wirbeltiere (Vertebrata). Auch heute noch werden sowohl im Meer als auch in Süßgewässern kontinuierlich neue Arten entdeckt. Die Fleischflosser, in deren fossiler Verwandtschaft sich der gemeinsame Vorfahr aller Landwirbeltiere findet, sind unter den Fischen nur noch mit acht rezenten (heute lebenden) Arten vertreten. Die Strahlenflosser stellen mit wenigstens 29.000 rezenten Arten mehr als 96 % der gesamten rezenten Fischfauna.
Strahlenflosser und Fleischflosser treten seit dem obersten Silur im Fossilbericht in Erscheinung. Im anschließenden Devon durchliefen sie eine rasche Aufspaltung in zahlreiche Arten (adaptive Radiation).[2]
Merkmale
.svg.png.webp)
1 Kiemendeckel
2 Seitenlinie
3 Rückenflosse (Dorsalis)
4 Fettflosse
5 Schwanzstiel
6 Schwanzflosse (Caudalis)
7 Afterflosse (Analis)
8 Leuchtorgane
9 Bauchflossen (Ventrales)
10 Brustflossen (Pectorales)
Die meisten Knochenfische haben einen spindelförmigen, der Bewegung im Wasser wenig Widerstand leistenden Körper. Paarige und unpaarige Flossen dienen dem Antrieb und der Stabilisierung.
Anhand der Flossenanatomie lassen sich Strahlenflosser und Fleischflosser unterscheiden: Die Flossen der Strahlenflosser werden von einer Reihe Radien gebildet, schmalen Strahlen aus Knochensubstanz oder Knorpel. Bei den Fleischflossern werden die Flossen hingegen von einem einzelnen, von Muskeln bewegten basalen Knochen gestützt. Aus den paarigen Brust- und Bauchflossen dieser Gruppe sind die Extremitäten der Landwirbeltiere hervorgegangen.[2]
Es kann je nach Fischfamilie eine, zwei oder drei Rückenflossen geben. Manche, wie die Alt- und Neuwelt-Messerfische, haben sie auch reduziert. Zwischen der Rückenflosse und der Schwanzflosse haben einige Taxa, wie viele Ostariophysi, noch eine strahlenlose, kleine Fettflosse. Die Flossen und die Anzahl und Art der Flossenstrahlen sind wichtige Merkmale in der Taxonomie.
Die Länge ausgewachsener Knochenfische schwankt je nach Art zwischen einem knappen Zentimeter (Paedocypris progenetica) und 8 Metern (Regalecus glesne). Der schwerste Knochenfisch der Welt ist der bis zu drei Meter lange, vier Meter hohe und bis zu 2,3 Tonnen schwere Mondfisch (Mola mola).
Vielfalt der Körperformen

Einige Knochenfische weichen in ihrer Gestalt sehr vom üblichen Bauplan der Fische ab. Bekanntestes Beispiel sind die Plattfische (Pleuronectiformes), die sich im Verlauf ihrer Ontogenese auf eine Körperseite legen, die von da ab die Unterseite bildet. Das Auge dieser Seite wandert auf die andere Körperseite, die von da an die Oberseite ist; die Brustflosse der Blindseite kann verkümmern. Die Plattfische verlieren dadurch ihre Symmetrie.
Andere Fischfamilien, wie die Anglerfische (Antennariidae), die Steinfische (Synanceiidae) und die Fetzenfische (Solegnathinae), lösen mit einer Vielzahl von Körperauswüchsen ihre Konturen auf, um entweder als Räuber oder als potentielle Beute getarnt zu sein.
Die pelagischen Mondfische sind höher als lang, haben keine Bauch- und keine Schwanzflosse und werden auch als „schwimmender Kopf“ bezeichnet.
Haut
Die Haut der Fische besteht aus zwei Schichten: der bindegewebshaltigen Lederhaut, in der sich Schuppen und Farbzellen befinden, und der mit Schleimdrüsen versehenen Oberhaut. Der Schleim hat eine schützende Wirkung auf die Haut und die darin liegenden Schuppen, da er antibakteriell wirkt. Die Schuppen verringern den Strömungswiderstand, da sie die Grenzgeschwindigkeit für die Entstehung von Turbulenzen heraufsetzen (am besten die Ctenoid-Schuppen).
Anatomie und Physiologie
Skelett
Das Skelett besteht aus Knochen, bei primitiven (aber auch abgeleiteten) Arten teilweise noch aus Knorpel. Am Schädel befindet sich das Kiemenskelett aus sieben Kiemenbögen, von denen der vordere zum Unterkiefer umgebildet ist. In der Rumpfregion tragen die Wirbel Rippen, über die ganze Länge Dornfortsätze auf der Rückenseite, im Schwanzbereich auch auf der Bauchseite. In den Muskelscheiden von Knochenfischen befinden sich häufig Gräten aus verknöchertem Bindegewebe. Zur Steuerung und Fortbewegung dienen die Flossen, die bis auf die Fettflosse (bei Lachsartigen, Salmlerartigen, Welsartigen) mit knöchernen Flossenstrahlen (Radien, aus Schuppenreihen entstanden) verstärkt sind. Brust- und Bauchflossen sind paarig und gelenken am Schultergürtel beziehungsweise am Becken. Die unpaarigen Rücken-, After- und Schwanzflosse(n) sind über Flossenträger (Radialia) mit der Wirbelsäule verbunden. Die erwähnte Fettflosse hat kein Skelett, kann aber hornartig verstärkt sein.
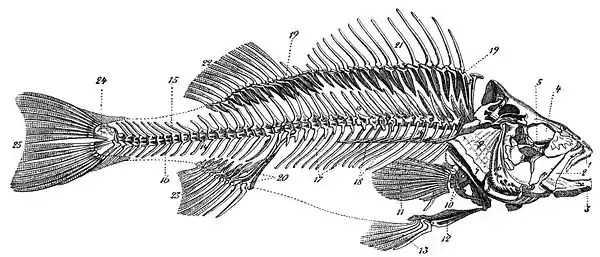
Die nebenstehende Abbildung zeigt das Skelett eines Flussbarsches (Perca fluviatilis). Die Namen der wichtigsten Knochen lauten in der weitgehend international gültigen Nomenklatur:
1 Praemaxillare (bezahnt), 2 Maxillare, 3 Unterkiefer, bestehend aus Dentale (bezahnt), Articulare und Angulare, 4 Augenhöhle (Orbita), 5 (sechs) Suborbitalia (das vordere, größte heißt Lacrimale), 6 Praeoperculare, 7 Interoperculare, 8 Suboperculare, 9 Operculare, 10 Schultergürtel mit Scapulare und Coracoideum, 11 Pectoralis (13 Radien auf vier Radialia), 12 Beckenknochen (Pelvis), 13 Pelvicalis (sechs Radien; brustständig), 14 (40) Wirbel (Vertebrae), 15 Neurapophysen, 16 Haemapophysen, 17 Rippen (Costae), 18 Gräten (Epipleuralia), 19, 20 Flossenträger (Pterygiophoren), 21, 22 zwei Dorsales, 23 Analis, 24 Hypuralia, 25 Caudalis (mit 17 Radien).
Muskeln
Die Muskulatur der Knochenfische wird in Rumpf- und Flossenmuskulatur unterteilt. Die Rumpfmuskulatur besteht aus Einzelsegmenten, deren Anzahl meist der der Wirbel entspricht. Der Mondfisch hat nur 17 Wirbel, der Schnepfenaal (Nemichthys scolopaceus) über 600. Am Schädel setzt naturgemäß eine große Zahl von Muskeln für Nahrungserwerb und Atmung an. Sie sind segmental in Myomere gegliedert. Die Scheidewände zwischen den Myomeren heißen Myosepten. Die Myomere verlaufen W-förmig wenn man sie von der Seite betrachtet. Bei manchen Arten (vor allem Pflanzenfresser) entwickelt sich ein muskelstarker Kaumagen.
Kreislauf und Atmung
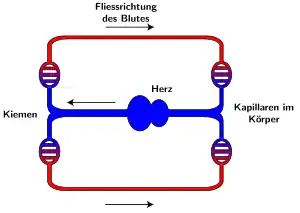
Knochenfische haben einen geschlossenen Blutkreislauf mit einem nahe an den Kiemen gelegenen, einfachen Herzen, das aus einem Vorhof und einer Herzkammer besteht. Es pumpt venöses Blut direkt in die Kiemen, die aus zahlreichen, mit Kapillaren durchzogenen Blättchen, feinen Hautausstülpungen, bestehen. Die Kiemen nehmen über ihre große Oberfläche gelösten Sauerstoff aus dem Wasser auf und geben ihn an das Blut weiter. Die sehr hohe Effektivität der Sauerstoffaufnahme über die Kiemen (bis zu 70 % des verfügbaren Sauerstoffs) kann auf das Prinzip des Gegenstrom-Austausches in den Kiemen und die sehr hohe Affinität von einigen Knochenfischhämoglobinen zu Sauerstoff zurückgeführt werden.
Die Kiemen sind in der Kiemenhöhle durch den knöchernen Kiemendeckel (Operculum) geschützt. Durch die Absenkung des Mundbodens und das gleichzeitige Abheben des Kiemendeckels strömt Atemwasser durch das Maul ein, während dies am Kiemendeckel durch eine weiche Membran verhindert wird. Beim Auspressen schließen sich im vorderen Maulbereich zwei Klappen (Valven) und das Wasser wird durch die Kiemendeckelspalte nach hinten ausgepresst; zu beachten ist, dass diese beiden Vorgänge fast synchron ablaufen, so dass ständig Wasser durch die Kiemenspalten fließen kann.
Bei einigen Knochenfischen (beispielsweise Aalen) ist die Kiemenhöhle nur durch einen kleinen Spalt nach außen geöffnet, wodurch die Kiemen auch auf dem Trockenen eine gewisse Zeit feucht und damit funktionsfähig bleiben. Einige mehr oder weniger amphibisch lebende Fischarten verfügen über weitere Atmungsorgane: Schlammspringer können Luftsauerstoff durch Hautatmung aufnehmen. Verbreiteter ist die Darmatmung, beispielsweise beim Schlammpeitzger, dabei wird Luft geschluckt, der Gasaustausch findet an den Wänden des stark durchbluteten Vorderdarms statt. Einige Fische (Labyrinthfische) atmen Luft über Schädelkammern, indem sie Ausstülpungen im Kiemenbereich zur Aufnahme von Sauerstoff nutzen. Fischen, deren Schwimmblase noch mit dem Darm verbunden ist, wie den Knochenhechten, dient auch die Schwimmblase zur Luftatmung. Bei Flösselhechten und Lungenfischen haben sich diese Darmausstülpungen bereits zu Lungen entwickelt.
Das Blut dient auch dem Transport der in den inkretorischen Drüsen gebildeten Hormone. Sowohl die Drüsen als auch die Hormone gleichen schon prinzipiell denen des Menschen.
Darm

1 Leber, 2 Magen, 3 Darm, 4 Herz, 5 Schwimmblase, 6 Niere, 7 Hoden, 8 Harnleiter, 9 Samenleiter, 10 Harnblase, 11 Kiemen
Der Darm lässt sich unterteilen in: Mundhöhle (Zähne äußerst vielgestaltig, selten fehlend, z. B. bei Cypriniformes), Kiemendarm (Pharynx mit Spalten zu den Kiemen, meist mit Reusenbildung; am Ende die Schlundknochen mit Zähnen, die sehr selten fehlen), Speiseröhre (Oesophagus), Magen (vielgestaltig, fehlt z. B. bei den Cypriniformes), Mitteldarm (sehr verschieden lang, je nach Ernährung; am Anfang können Pförtner-Blindschläuche von ihm ausgehen, 0 bis 1000 an Zahl, Funktion unklar; Leber und Bauchspeicheldrüse oft noch nicht geschieden), Enddarm (bei Pflanzenfressern mitunter mit Blinddarm; eine Spiralfalte darin wie bei den Knorpelfischen findet sich noch bei allen Nicht-Teleostei unter den Osteichthyes, wird aber mehr und mehr rückgebildet).
Schwimmblase

Bei den meisten Knochenfischarten hat die Schwimmblase die Funktion, das spezifische Gewicht der Fische (die Dichte) zu regulieren, so dass sie ohne Kraftanstrengung im Wasser schweben können oder nur sehr langsam zu Boden sinken. Hervorgegangen ist sie aus einer Ausstülpung an der Oberseite des Vorderdarms; sie kann, wie bei den Karpfen, noch mit ihm verbunden sein (physostom) oder, wie bei den Barschen, vom Darm getrennte Kammern bilden (physoklist).
Zur Regulierung des spezifischen Gewichts werden Gase (Sauerstoff, Kohlendioxid, Stickstoff) aus dem Blut über die so genannte Gasdrüse oder den Verbindungsgang zum Darm, den Ductus pneumaticus, in die Schwimmblase abgegeben. Zur Volumenverminderung der Schwimmblase gelangt das Gas entweder über den Ductus pneumaticus in den Vorderdarm (und wird ausgespien) oder über einen stark durchbluteten Bereich in der Schwimmblasenwand, das Oval, wieder zurück ins Blut.
Knorpelfischen, aber auch bodenlebenden oder besonders gut schwimmenden Knochenfischen fehlt die Schwimmblase – sie sinken zu Boden, wenn sie sich nicht fortbewegen. Durch Knochenreduktion, Fetteinlagerung u. ä. kann aber zum Beispiel der Mondfisch (Mola) auch ohne Schwimmblase an der Meeresoberfläche treiben.
Nervensystem und Sinnesorgane
Das Nervensystem ist einfach, das Gehirn klein, eine ausgeprägte Großhirnrinde fehlt. Der Geruchssinn der Fische ist meist sehr ausgeprägt, Nasen- und Mundhöhle sind voneinander getrennt. Die Gleichgewichts- und Gehörorgane bestehen aus geschlossenen, flüssigkeitsgefüllten Blasen. Bei einigen Arten (Karpfen, Salmler, Welse) sind sie durch bewegliche Knochen mit der Schwimmblase verbunden und werden als Weberscher Apparat bezeichnet.[3] Im Aufbau ähneln sie dem Innenohr der Säugetiere, eine Hörschnecke ist jedoch nicht ausgebildet. Die meisten Fischarten können Signale unterhalb von 1 kHz wahrnehmen, einige haben ihr Hörvermögen durch spezielle Strukturen verbessert und den Hörbereich bis 5 kHz oder mehr erweitert.
Auge
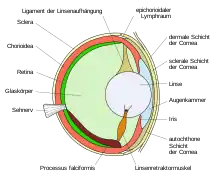
Das Auge der Knochenfische ist in seinem Aufbau und seiner Funktion ein weitgehend typisches Wirbeltierauge und somit grundsätzlich ähnlich dem Auge des Menschen. Ein bedeutender Unterschied zum Auge der Landwirbeltiere besteht darin, dass die Öffnung in der Regenbogenhaut (Iris), die Pupille, zumeist starr ist, sodass die Menge des in das Auge einfallenden Lichtes nicht reguliert werden kann. Der größte Unterschied zum Auge der Landwirbeltiere besteht jedoch in der Form der Linse. Diese ist kugelförmig und das einzige Teilorgan des Auges, das für die Brechung der Lichtstrahlen verantwortlich ist, wohingegen bei Landwirbeltieren der Hauptteil der Lichtbrechung (ca. 85 %[4]) bereits beim Passieren der Hornhaut infolge des deutlichen Dichteunterschiedes zwischen Luft und Hornhaut erfolgt. Eine Konsequenz der Kugelform wäre jedoch eine starke sphärische Aberration und damit die Erzeugung eines unscharfen Bildes auf der Netzhaut (Retina). Um diesen Fehler zu verringern, weist der Linsenkörper inhomogene optische Eigenschaften auf: der Brechungsindex nimmt von der Mitte zum Rand der Linse hin ab. Eine solche Linse wird als Gleitindexlinse oder Matthiessenlinse bezeichnet. Außerdem ist die Linse des Fischauges nicht flexibel und besitzt folglich eine feste Brennweite. Die Akkommodation, das heißt die Umstellung von Nah- auf Fernsicht, erfolgt durch Kontraktion des sogenannten Linsenretraktormuskels (Musculus retractor lentis), wodurch die Linse entlang der optischen Achse näher an die Netzhaut herangeführt wird. Absatz bis hierher, wo nicht anderweitig zitiert, nach [5][6] und [7] Das Auge der Knochenfische ist folglich auf Nahsicht voreingestellt.
Für das Auge der Knorpelfische galt vormals, dass dort die Akkommodation genau umgekehrt laufe: das Auge sei auf Fernsicht voreingestellt und die Kontraktion eines Protraktormuskels sorge für die Umstellung auf Nahsicht. Dies konnte jedoch durch verschiedene Untersuchungen an mehreren Haiarten nicht bestätigt werden, sodass es nunmehr als unklar gilt, wie und ob überhaupt die Augen der Knorpelfische akkommodieren.[8][9] Viele Fische sind farbtüchtig und nehmen auch ultraviolettes Licht wahr.
Tastsinn
Besonders bei bodenlebenden Fischen ist der Tastsinn gut entwickelt. Vor allem an Lippen und Barteln befinden sich in der Oberhaut Sinneszellen, die Berührungs-, aber natürlich auch Geschmacksreize übertragen. Als „sechsten Sinn“ verfügen Fische über Organe zur Wahrnehmung von Wasserströmungen, die Seitenlinienorgane, die sich seitlich etwa in der Mitte über einen Großteil der Körperlänge erstrecken. Sie bestehen aus einer Reihe von Hauteinstülpungen, in denen sich Sinneszellen mit Sinneshaaren befinden, die durch Strömungsveränderungen erregt werden. Wenn diese „Seitenlinie“ fehlt (z. B. beim Hering), gibt es doch auch am Kopf solche Organe bei (fast) allen Knochenfischen.
Fortbewegung
Zur Fortbewegung dient – bei seitlich schlängelnder Körperbewegung – vor allem die Schwanzflosse. Lippfische und Doktorfische bewegen sich jedoch überwiegend mittels der Brustflossen fort und nutzen die Schwanzflosse lediglich als Steuerruder (labriform). Bei den Kugelfischverwandten dienen die Rücken- und die Afterflosse, die sich genau gegenüberliegen, der Fortbewegung. Den Neu- und Altwelt-Messerfischen dient allein die wellenartige Bewegung der Afterflosse dem Vorwärtskommen. Sie können durch Umkehrung der Wellenbewegung ebenso geschickt rückwärts schwimmen. Für die Einstellung der Körperorientierung dient neben dem Gleichgewichtssinn auch der Lichtrückenreflex.
Fortpflanzungsbiologie

Die Nieren sind ein paariges (oder unpaarig verschmolzenes) langgestrecktes Organ unterhalb der Wirbelsäule. Bei allen anderen „Fischen“ dienen die Harnleiter (oder davon abgegliederte Röhren) auch der Ausleitung der Keimzellen – aber die Echten Knochenfische haben dazu eigene Ei- und Samenleiter entwickelt. Ihre Fortpflanzungsorgane befinden sich seitlich und oberhalb des Darms.
Die Befruchtung findet bei den meisten Arten ohne Kopulation statt, stattdessen werden der weibliche Rogen (die Eier) und die männliche Milch (der Samen) etwa gleichzeitig ins Wasser abgesetzt. (Der zur Fortpflanzung fähige weibliche Fisch heißt „Rogner“, der fortpflanzungsfähige männliche Fisch wird „Milchner“ genannt.) Dabei schwankt die Anzahl der Eier extrem: Störe legen mehrere Millionen, die brutpflegenden Stichlinge nicht mehr als hundert. Bei brutpflegenden Arten sind es häufig die Männchen, die die Pflege übernehmen.
Es gibt bei den brutpflegenden Arten verschiedene Formen: Offen-, Höhlen- und Maulbrüter.
- Bei Offenbrütern werden die Eier vom Weibchen auf Pflanzen oder anderen festen Materialien abgelegt und anschließend vom Männchen besamt.
- Bei Höhlenbrütern werden die Eier an der Decke von Höhlen abgelegt. Nach dem Schlüpfen werden die Jungen noch eine Zeit lang auf Blättern oder in Gruben abgelegt (bis der Dottersack verbraucht ist) und bewacht. Danach werden die Larven von den Eltern geführt, wobei sich die Eltern entweder abwechseln oder ein Elternteil, meistens das Männchen, die Reviergrenzen bewacht. Geht ein Großteil der Brut verloren, so kann es vorkommen, dass die Eltern die wenigen überlebenden Larven fressen und schnell wieder mit einer neuen Brut beginnen.
- Bei Maulbrütern nehmen die Weibchen die Eier nach der Ablage ins Maul, das Männchen schwimmt über die Eier und gibt dabei seinen Samen ab. Die Jungen der Maulbrüter sind, wenn sie das Maul verlassen, schon weit entwickelt, sie kehren nach dem Schlüpfen aber noch eine Zeit lang bei Gefahr ins Maul zurück.
Die Larven der Knochenfische entwickeln sich durch Metamorphose zu Adulttieren. Bei der Metamorphose werden die larvalen Organe resorbiert oder abgestoßen und die vorhandenen Anlagen der Adultorgane zur Funktionsfähigkeit entwickelt.
Beispiele für Besonderheiten bei der Fortpflanzung:
- Kuckucks-Fiederbartwelse haben sich darauf spezialisiert, ihre Eier zu den Eiern des Wirtsfisches abzulegen, die dann vom Weibchen mit ins Maul aufgenommen und dort „ausgebrütet“ werden. Die sich schneller entwickelnden Welslarven fressen dann die Eier oder Larven des Wirtsfisches.
- Diskusfische sondern nach dem Schlüpfen ihrer Brut ein Hautsekret ab, das von den Larven abgeweidet wird und als Erstnahrung dient.
- Bei Labyrinthfischen bilden die Männchen häufig an der Wasseroberfläche zwischen Schwimmpflanzen ein Schaumnest und treiben dann das Weibchen zur Eiablage unter das Schaumnest. Dabei umschlingt das Männchen mit seinem Körper das Weibchen und dreht es auf den Rücken, so dass die Eier nach oben schwimmen. Nach dem Besamen wird das Weibchen vertrieben und die Brut vom Männchen allein behütet, bis die Larven schlüpfen, danach kümmert sich auch das Männchen nicht weiter um die Brut.
- Es gibt auch (selten) Knochenfische, die über Kopulationsorgane verfügen, wie beispielsweise die Lebendgebärenden Zahnkarpfen (Poeciliinae) oder die Falschen Dornwelse (Auchenipteridae).
- Einige Knochenfische, besonders marine Barschverwandte, haben kein genetisch bestimmtes Geschlecht, es prägt sich erst durch den Kontakt mit Partnern oder durch Umweltbedingungen aus und kann im Laufe des Lebens gewechselt werden.
Lebensräume
_yellowtail_barracuda.JPG.webp)


Ozeane
Die Weltmeere haben einen Anteil von etwa 70 % an der Oberfläche der Erde und sind damit der größte Lebensraum. Die Knochenfische, die die obere Schicht des offenen Ozean bis in eine Tiefe von 200 Meter als Lebensraum gewählt haben, machen aber nur etwas mehr als ein Prozent aller Arten aus, zum größten Teil Angehörige der Makrelen und Thunfische (Scombridae) und der Fliegenden Fische (Exocoetidae). Weitere fünf Prozent der Knochenfische leben pelagisch unterhalb von 200 Metern. Es sind zum großen Teil Heringsartige (Clupeiformes) und Laternenfische (Myctophidae).
Der größte Teil der marinen Knochenfische lebt in der Nähe der Küsten. Die kalten Küsten bewohnen hauptsächlich Dorsche (Gadidae), Aalmuttern (Zoarcidae), Schleimfischartige (Blennioidei), Panzerwangen (Scorpaeniformes, keine Verwandtschaftsgruppe) und Antarktisfische (Notothenioidei). Etwas mehr als fünf Prozent der Knochenfischarten bevorzugen diesen Lebensraum, der zugleich ein wichtiges Fischereigebiet ist, weil die kalten, küstennahen Meere sehr nährstoffreich sind und die Fische in großen Schwärmen vorkommen. Die warmen Küsten der Kontinente und tropischen Inseln sind wegen der vielfältigen ökologischen Nischen in Korallenriffen und Mangroven mit einem Anteil von mehr als 40 % an der gesamten Knochenfischfauna der artenreichste marine Lebensraum. Die hier hauptsächlich vorkommenden Arten gehören zum großen Teil zu den Barschverwandten (Percomorphaceae), im Einzelnen z. B. zu den Barschartigen (Perciformes), den Schleimfischartigen (Blennioidei) und den Grundelartigen (Gobiiformes). Außerdem gibt es sehr viele Aalartige (Anguilliformes) an warmen Küsten, z. B. die Muränen.
Besondere Lebensräume bilden die Kontinentalabhänge und die bodennahe Zone der Tiefsee, auf und über denen etwas mehr als sechs Prozent aller Knochenfische leben. Grenadiere (Macrouridae), Eingeweidefischartige (Ophidiiformes), Aalmuttern (Zoarcidae), viele Aalartige (Anguilliformes) und Panzerwangen (Scorpaeniformes) haben hier ihren Lebensraum.
Binnengewässer
Obwohl der Anteil der Binnengewässer (Süßgewässer und auch Salzseen) am Wasservorkommen der Erde nur 2,6 bis 3 % beträgt, beherbergen sie insgesamt etwa 40 % aller Knochenfischarten. Die geografische Isolierung und Unterschiede in vielen Bedingungen, wie Untergrund, Temperatur, Fließgeschwindigkeit, Jahreszeiten, Sauerstoffgehalt, Härte und pH-Wert, führten zu einer großen Vielfalt von Anpassungen und Arten. Die meisten im Süßwasser lebenden Knochenfische mit ca. 6000 Arten gehören zu den Karpfenartigen (Cypriniformes), den Welsartigen (Siluriformes) und den Salmlerartigen (Characiformes), die zusammen mit den artenärmeren Ordnungen der Neuwelt-Messerfische (Gymnotiformes) und der Sandfischartigen (Gonorynchiformes; marin!) das Taxon der Ostariophysi bilden. Aus dem Verbreitungsgebiet dieser Ordnungen kann man schließen, dass die Ostariophysi entstanden, als alle Kontinente außer Australien, wo sie nicht vorkommen, noch zusammenhingen. Die Ostariophysi haben sich früh von einem gemeinsamen Vorfahren mit den Heringsartigen (Clupeiformes) abgespalten und auf das Leben in Süßgewässern spezialisiert. Sie sind primäre Süßwasserfische. Die bedeutendsten Süßwasserfische nach den Ostariophysi sind die Buntbarsche (Cichlidae), die die Süßwasserfauna der ostafrikanischen Seen und Mittelamerikas dominieren, daneben aber auch in Westafrika, Südamerika, Madagaskar und Südindien vorkommen.
Phylogenie, Systematik und Fossilbericht
Äußere Systematik: Ursprung der Knochenfische

Wann genau der erste Knochenfisch lebte, ist unbekannt. Man muss eine nicht fossil dokumentierte Geschichte dieser Gruppe ab dem frühen Silur annehmen. Das älteste bekannte relativ vollständige Fossil eines Knochenfisches stammt aus Ablagerungen des Ober-Silur (Ludlowium) im heutigen China.[10] Dieser etwa 26 Zentimeter lange, schlanke, stromlinienförmige Vertreter, der den wissenschaftlichen Namen Guiyu oneiros trägt, wird den Muskelflossern (Sarcopterygii) zugerechnet, besitzt aber noch Merkmale, die sich auch bei anderen sehr ursprünglichen Wirbeltiergruppen (Knorpelfische, Acanthodier, Placodermen) finden und erst im weiteren Verlauf der Evolution der Knochenfische verloren gingen. Andreolepis hedei aus dem nördlichen Europa stammt aus geringfügig älteren Schichten als Guiyu und gilt damit als der älteste bekannte Knochenfisch überhaupt. Er scheint noch ursprünglicher als Guiyu zu sein und repräsentiert offenbar einen Entwicklungsstand vor der Trennung der Linien der Muskelflosser und Strahlenflosser.[10][11] Allerdings sind seine Überreste deutlich bruchstückhafter und weniger vollständig als die von Guiyu. Sowohl Guiyu als auch Andreolepis sowie einige andere bruchstückhaft erhaltene ober-silurische Fossilfunde von Knochenfischen stammen aus Meeresablagerungen, d. h., nach aktuellem Kenntnisstand sind die Knochenfische im Meer entstanden.
Da die Knochenfische Ähnlichkeiten zu den ausgestorbenen Acanthodiern zeigen, nimmt man an, dass diese die Schwestergruppe der Knochenfische sind. Mit ihnen haben sie den Kiemenaufbau aus vier Paar Kiemen mit Kiemenspalte und Kiemendeckel gemeinsam. Acanthodier und Knochenfische haben wiederum gemeinsame Vorfahren mit den Knorpelfischen. Die ältesten und ursprünglichsten Kiefermäuler sind die Placodermen, die sich einen gemeinsamen Vorfahren mit dem Taxon aus Knorpelfischen, Acanthodiern und Knochenfischen teilen. Es gilt als sicher, dass die Knochenfische nicht unmittelbar mit den Knorpelfischen verwandt sind. Nach bisherigen Erkenntnissen sieht der wahrscheinlichste Stammbaum der Knochenfische (Osteichthyes) so aus:[1]
| Kiefermäuler (Gnathostomata) |
| ||||||||||||||||||||||||
1) Das Taxon, welches diesen Zweig des Stammbaums einnimmt, wird, um Verwechslungen mit dem paraphyletischen Taxon Osteichthyes der klassischen Systematik zu vermeiden, auch Euteleostomi oder Osteognathostomata genannt (siehe Knochenfische und Landwirbeltiere).
Innere Systematik: Evolutionsgeschichte der Knochenfische


.jpg.webp)
Bereits im frühen Devon finden sich Knochenfisch-Fossilien auch im Süßwasser. So stammen die ältesten als Quastenflosser (Coelacanthimorpha) identifizierten Fossilien aus Süßwassersedimenten des Pragiums in China (Euporosteus yunnanensis) und Australien (Eoactinistia foreyi). Einer der frühesten Vertreter jener Entwicklungslinie, die zu den modernen Lungenfischen und Tetrapoden führt, Youngolepis, wurde wiederum in China gefunden, in Süßwassersedimenten des Lochkoviums. Der älteste zweifelsfreie Vertreter der Strahlenflosser Cheirolepis trailli stammt aus mittel-devonischen (Emsium) Schichten des Old-Red-Sandsteins in Schottland. Zu dieser Zeit unterscheiden sich die Vertreter der beiden Hauptlinien der Knochenfische, Muskelflosser und Strahlenflosser, bereits deutlich in ihren Körpermerkmalen und haben zudem viele ursprüngliche Merkmale verloren, welche auf die gemeinsame Vorfahrenschaft mit anderen basalen Wirbeltiergruppen zurückgingen. Sowohl die frühen Strahlenflosser als auch die frühen Muskelflosser sind aktive Räuber und die Muskelflosser sind während des Mittel-Devon die dominierenden Fische der Süßgewässer. Aus ihnen gehen im Laufe des Ober-Devon die ersten Landwirbeltiere (Tetrapoda) hervor, belegt durch Übergangsformen, wie Eusthenopteron, Panderichthys und Tiktaalik. Im Zuge des Aussterbe-Ereignisses am Ende des Perm wird die Artenzahl der Muskelflosser stark reduziert. Während die Tetrapoden im Mesozoikum eine erneute Blüte erleben, bleiben ihre fischartigen Muskelflosser-Verwandten vergleichsweise artenarm und verschwinden mit dem Ende der Kreide ganz aus der Fossilüberlieferung. Erst im 19. Jahrhundert wurden die sechs heute lebenden Arten der Lungenfische und 1938 der Komoren-Quastenflosser (Latimeria chalumnae) als „Lebende Fossilien“ entdeckt.
Die Flösselhechte (Polypteriformes) sind das ursprünglichste Taxon der heute lebenden Strahlenflosser. Die ältesten Fossilien stammen zwar aus der frühen Ober-Kreide, aber die Geschichte dieser Gruppe muss mindestens bis ins Karbon zurückreichen, wo die Vertreter ihrer nur fossil bekannten Schwestergruppe, die Guildayichthyiden lebten.[12] Sowohl die Guildayichthyiden als auch die Flösselhechte, die zusammen als Cladistia bezeichnet werden, waren aber offenbar nie sonderlich artenreich.
Im Gegensatz dazu durchliefen die Knorpelganoiden (Chondrostei)[An 1] eine rasche Radiation im Karbon. Eine nur durch Fossilien bekannte Gruppe dieser ersten großen Evolutionswelle sind die Palaeonisciformes. Zu ihnen gehört u. a. der häufigste Fisch im Kupferschiefer der Zechsteinserie, einer permischen Gesteinsabfolge in Mitteleuropa, Palaeoniscus freieslebeni. Vertreter der Familie Redfieldiidae sind typisch für triassisch-jurassische Süßwasserablagerungen der Newark-Supergruppe, einer Gesteinsabfolge am Ostrand Nordamerikas, die mit der Entstehung des Atlantiks in Zusammenhang steht. Die Störartigen (Acipenseriformes) sind die noch heute lebenden Repräsentanten der Knorpelganoiden. Sie sind fossil erst ab dem unteren Jura bekannt, allerdings wurde vor kurzem der Fund eines frühen Störartigen aus dem Perm von China vermeldet (Eochondrosteus sinensis).[13]
Schon ab dem Ende der Trias erleben die Knorpelganoiden einen Niedergang, möglicherweise u. a. ausgelöst durch zunehmende Konkurrenz von Vertretern der basalen Neuflosser (Neopterygii), die während der Trias rasch evolvierten. Die nord- und mittelamerikanischen Knochenhechte (Lepisosteidae) und der nordamerikanische Kahlhecht oder Schlammfisch (Amia calva), die beide zusammen mit ihren ausgestorbenen Verwandten als Knochenganoiden (Holostei)[An 2] bezeichnet werden, sind Überlebende dieser zweiten Evolutionswelle der Strahlenflosser, die erst in der späten Kreide beendet war. Zwei eng mit den heute lebenden Holosteer-Arten verwandte Vertreter aus dem Eozän, Cyclurus kehreri und Atractosteus strausi, gehören zu den häufigsten Wirbeltierfossilien in der berühmten Grube Messel.
Das folgende Kladogramm zeigt die Verwandtschaft der basalen Taxa der rezenten Knochenfische. In Klammern ist die Anzahl der heute lebenden Arten angegeben.

| Knochenfische (Osteichthyes) |
| ||||||||||||||||||||||||||||||||||||


Bereits in der späten Trias vor etwa 220 Millionen Jahren erscheinen erstmals Vertreter der Echten Knochenfische (Teleostei) in der Fossilüberlieferung. Die Teleosteer zeichnen sich u. a. dadurch aus, dass sie keine stark mineralisierten Schuppen mehr haben, was sich im Fossilbericht daran zeigt, dass von diesen Vertretern nurmehr das Innenskelett fossil überliefert wird. Zu den basalen Teleosteern gehört auch Leedsichthys problematicus aus dem Jura, der größte Knochenfisch der Erdgeschichte, der möglicherweise sogar eine Länge von bis zu 25 Metern erreicht hat. Diese basalen Linien starben allerdings alle noch vor dem Ende der Kreide aus. Dies könnte wiederum damit zusammenhängen, dass die Teleosteer im Ober-Jura und vor allem im Laufe der Kreidezeit einen weiteren Evolutionsschub erfuhren, im Zuge dessen fast alle modernen Großgruppen entstanden. Diese moderneren Vertreter dürften den frühen Teleosteern zunehmend Konkurrenz gemacht haben. In der oberen Kreide und, als Folge des Massenaussterbens an der Kreide-Tertiär-Grenze, vor allem im unteren Tertiär[14] vollzog sich die letzte große Phase rascher evolutionärer Aufspaltung bei den Teleosteern, die im Wesentlichen die Barschverwandten (Percomorphaceae) und die Ostariophysi (Karpfen, Welse und Verwandte) betraf. Diese beiden Gruppen sind heute die artenreichsten (Barschverwandte: ca. 14.000 Arten[15], Ostariophysi: ca. 6000 Arten, darunter zwei Drittel aller Süßwasserfischarten).
Nur 57 % aller heute lebenden Familien der Echten Knochenfische sind im Fossilbericht vertreten. Fast 70 Familien sind ausgestorben.
Der Erfolg der Echten Knochenfische beruht zu einem großen Teil auf der Anpassung an das Leben im freien Wasser. Es gibt im Laufe der Knochenfischevolution eine Tendenz zur Verminderung der Anzahl der Wirbel, wodurch der Fischkörper steifer und strömungsgünstiger wird. Eine weitere Ursache dürfte die im Vergleich zu basaleren Knochenfischen verbesserte Schädelkinetik, einhergehend mit der Entwicklung eines hochentwickelten Kieferapparates sein. Letztgenannter erlaubt Räubern eine effektivere Jagd durch Saugschnappen, bildet aber auch die Basis für die Entwicklung sehr spezieller Ernährungsformen, wie z. B. das Abweiden des Aufwuchses von Felsen und in Korallenriffen. Die Teleosteer stellen heute mit über 40 Ordnungen und über 400 Familien den größten Teil der Fischarten.
Einige Taxa der Echten Knochenfische, wie die Maulstachler (Stomiiformes), die Eidechsenfischverwandten (Aulopiformes) und die Laternenfischverwandten (Myctophiformes) passten sich einem Leben in der Tiefsee an.
Kladogramm zur inneren Systematik der Echten Knochenfische (Teleostei) nach Betancur-R. et al. (2016):[16]
| Neoteleostei |
| ||||||||||||||||||||||||||||||||||||||||||||||||
Die Evolution und die phylogenetische Verwandtschaft unter den Arten, Gattungen und Familien der Barschverwandten (Percomorphaceae) sind noch weitgehend ungeklärt. Die Fülle von fast 15.000 Arten erschwert die genaue Erforschung.
Eine Übersicht aller Ordnungen und Familien ist unter Systematik der Knochenfische zu finden.
Knochenfische und Landwirbeltiere

Aus Sicht der kladistischen Systematik gehören auch die Landwirbeltiere (Tetrapoda) zu den Knochenfischen i. w. S. Ohne sie sind die Knochenfische ein so genanntes paraphyletisches Taxon, das heißt, dass sie nicht alle Nachkommen des gemeinsamen Vorfahren der Knochenfische umfassen – eben weil die Landwirbeltiere mit zu diesen Nachkommen gehören. So sind beispielsweise die Lungenfische oder auch die Quastenflosser enger mit den Landwirbeltieren verwandt als mit anderen Fischgruppen. Deswegen lehnt die kladistische Systematik ein Taxon Knochenfische ohne Landwirbeltiere ab. Um Verwechslungen mit den paraphyletischen Osteichthyes auszuschließen, führten einige Kladisten die neuen Namen Osteognathostomata (Knochenkiefermäuler), Neognathostomata (Neukiefermäuler) und Euteleostomi für jenes Taxon ein, das auch die Landwirbeltiere einschließt.
Anmerkungen
- Der Gebrauch des Begriffes „Chondrostei“ ist in der Fachliteratur nicht einheitlich. Zum einen steht er, wie hier im Artikel, für eine größere Gruppe ursprünglicher Strahlenflosser, zum anderen wird er nur für die Störartigen und ihre nächsten fossilen Verwandten genutzt.
- Die Holosteer wurden in jüngerer Vergangenheit als paraphyletisch betrachtet. In neueren Verwandtschaftsanalysen bilden sie jedoch wieder ein Monophylum.
Literatur
- Robert L. Carroll: Paläontologie und Evolution der Wirbeltiere. Georg Thieme Verlag, Stuttgart 1993, ISBN 3-13-774401-6.
- Rudie H. Kuiter, Helmut Debelius: Atlas der Meeresfische. Kosmos-Verlag, 2006, ISBN 3-440-09562-2.
- Kurt Fiedler: Lehrbuch der Speziellen Zoologie, Band II, Teil 2: Fische. Gustav Fischer Verlag, Jena 1991, ISBN 3-334-00338-8.
- Karl Albert Frickhinger: Fossilien Atlas Fische. Mergus-Verlag, Melle 1999, ISBN 3-88244-018-X.
- Hans-Eckard Gruner, Horst Füller, Kurt Günther: Urania Tierreich, Fische, Lurche, Kriechtiere. (7 Bände), Urania-Verlag, 1991, ISBN 3-332-00376-3.
- Juraj Holcik: The Freshwater Fishes of Europe. Band 1/II, AULA-Verlag, Wiesbaden 1989, ISBN 3-89104-431-3.
- Oskar Kuhn: Die vorzeitlichen Fischartigen und Fische. A. Ziemsen Verlag, Wittenberg 1967.
- Joseph S. Nelson: Fishes of the World. John Wiley & Sons, 2006, ISBN 0-471-25031-7.
- Günther Sterba: Süsswasserfische der Welt. 2. Auflage. Urania, Leipzig/Jena/Berlin 1990, ISBN 3-332-00109-4.
- Volker Storch, Ulrich Welsch: Kükenthal, Zoologisches Praktikum, Kapithel Osteichthyes; Spektrum Akademischer Verlag; 26. Auflage; ISBN 978-3-8274-1998-9.
Weblinks
Einzelnachweise
- Hans-Peter Schultze: Gnathostomata, Kiefermäuler. In Wilfried Westheide, Reinhard Rieger (Hrsg.): Spezielle Zoologie. Teil 2: Wirbel- oder Schädeltiere. 2. Auflage. Spektrum Akademischer Verlag, Heidelberg 2010, ISBN 978-3-8274-2039-8, S. 211–215.
- Michael J. Benton: Paläontologie der Wirbeltiere. Übersetzung der 3. englischsprachigen Auflage (Übersetzer: Hans-Ulrich Pfretzschner). Verlag Dr. Friedrich Pfeil. München 2007. 472 Seiten. ISBN 978-3-89937-072-0.
- WDR.de: Sascha Ott: Geschwätzig wie ein Fisch – Unter Wasser spielt die akustische Kommunikation eine erstaunlich große Rolle. S. 7, (PDF 110kB), Leonardo – Wissenschaft und mehr. 29. Januar 2010.
- David J. Randall, Roger Eckert, Warren Burggren, Kathleen French: Tierphysiologie. 4. Auflage. Thieme, 2002, ISBN 3-13-664004-7, S. 282.
- Staatsinstitut für Schulqualität und Bildungsforschung: Grundlagen der Biophysik. BRIGG Pädagogik Verlag, München 2009, ISBN 978-3-87101-640-0, S. 13–14.
- Jan-Peter Hildebrandt, Horst Beckmann, Uwe Homberg: Penzlin – Lehrbuch der Tierphysiologie. 8. Auflage. Springer Spektrum, Berlin/Heidelberg 2015, ISBN 978-3-642-55369-1, S. 673.
- Andreas Sebastian Reimann: Anatomisch-makroskopische Untersuchungen von Fischaugen. Eine interaktive Anleitung zur Herstellung und Fotografie von ophthalmologischen Präparaten als Grundlage zur Fischophthalmologie. Dissertation. Tierärztliche Fakultät der Ludwig-Maximilians-Universität München, 2015, urn:nbn:de:bvb:19-180417, S. 3–18.
- Quentin Bone, Richard H. Moore: Biology of Fishes. 3. Auflage. Taylor & Francis, 2008, ISBN 978-0-415-37562-7, S. 313 f.
- Volker Storch, Ulrich Welsch: Kükenthal – Zoologisches Praktikum. 26. Auflage. Spektrum Akademischer Verlag, Heidelberg 2009, ISBN 978-3-8274-1998-9, S. 353 f.
- Zhu Min, Zhao Wenjin, Jia Liantao, Lu Jing, Qiao Tuo, Qu Qingming: The oldest articulated osteichthyan reveals mosaic gnathostome characters. In: Nature. Band 458, 2009, S. 469–474, doi:10.1038/nature07855.
- Hector Botella, Henning Blom, Markus Dorka, Per Erik Ahlberg, Philippe Janvier: Jaws and teeth of the earliest bony fishes. In: Nature. Band 448, 2007, S. 583–586, doi:10.1038/nature05989.
- Richard Lund: The new Actinopterygian order Guildayichthyiformes from the Lower Carboniferous of Montana (USA) In: Geodiversitas. Bd. 22, Nr. 2, 2000, S. 171–206, sciencepress.mnhn.fr.
- Lu Liwu, Li Daqing, Yang Liangfeng: Notes on the discovery of Permian Acipenseriformes in China. In: Chinese Science Bulletin. Band 50, Nr. 12, 2005, S. 1279–1280, doi:10.1007/BF03183706.
- Elizabeth C. Sibert, Richard D. Norris. New Age of Fishes initiated by the Cretaceous−Paleogene mass extinction. In: Proceedings of the National Academy of Sciences. Bd. 112, Nr. 28, 2015, S. 8537–8542, doi:10.1073/pnas.1504985112.
- G. David Johnson, E. O. Wiley: Percomorpha. Tree of Life Web Project (Version vom 9. Januar 2007).
- R. Betancur-R., E. Wiley, N. Bailly, A. Acero, M. Miya, G. Lecointre, G. Ortí: Phylogenetic Classification of Bony Fishes – Version 4 (2016)