Fayyum (Fossillagerstätte)

Das Fayyum ist eine Region und eine wichtige Fossillagerstätte im nördlichen Ägypten. Die Region umfasst das landwirtschaftlich intensiv genutzte Fayyum-Becken und daran anschließende Gebiete, die bedeutenden Fundbereiche sind zumeist nördlich und westlich des Qarun-Sees aufgeschlossen. Den südwestlichen Abschluss bildet das Wadi al-Hitan, bekannt durch zahlreiche Walfossilien und seit dem Jahr 2005 UNESCO-Weltnaturerbe. Die Ablagerungen des Fayyums gehören verschiedenen geologischen Formationen an. Sie setzen sich vorwiegend aus Kalk-, Schluff- und Sandsteinen zusammen. Die unteren Abschnitte bestehen aus Meeresablagerungen, die oberen, festländischen Sedimente sind in einer küstennahen Landschaft entstanden. Der Bildungszeitraum reicht vom Mittleren über das Obere Eozän bis zum Unteren Oligozän, was einem Alter von vor rund 41 bis 28 Millionen Jahren entspricht. Der gesamte Sedimentkomplex wird von Basalt überlagert, der auf vulkanische Aktivitäten vor etwa 24 Millionen Jahren zurückgeht.
Die sehr reichhaltigen fossilen Zeugnisse des Fayyum umfassen Pflanzen, Wirbellose und Wirbeltiere, darüber hinaus sind auch zahlreiche Spurenfossilien überliefert. Unter den Wirbeltieren finden sich Fische, Reptilien, Vögel und Säugetiere. Einige Formen stellen wichtige evolutionäre Bindeglieder in der Entwicklung einzelner Linien dar, insbesondere bei den Schliefern, den Rüsseltieren, den Seekühen und den Walen. Unter den fossilen Primaten befinden sich unter anderem frühe Formen der Altweltaffen. Darüber hinaus kommen auch Beuteltiere, Tenrekartige, Paarhufer, Fledertiere und Nagetiere vor. Einige Gruppen wie die riesigen, sich pflanzenfressend ernährenden Embrithopoda oder die räuberisch lebenden Hyaenodonta beziehungsweise die wohl insektenfressenden Ptolemaiida haben keine heute lebenden Nachfahren. Aufgrund dieser Vielfalt stellt das Fayyum die bedeutendste Fossilfundstelle des afrikanischen Paläogens dar. Anhand der Landwirbeltiere lässt sich eine von Wasserläufen durchzogene, deltaartige Landschaft in Nähe der Meeresküste unter tropischen Klimabedingungen rekonstruieren.
Die ersten Fossilfunde im Fayyum reichen in die Mitte des 19. Jahrhunderts zurück. Eine große Untersuchungsphase fand am Übergang vom 19. zum 20. Jahrhundert statt, als zeitweise zugleich englische, amerikanische, deutsche und französische Forscher vor Ort tätig waren. Während dieser Zeit wurden zahlreiche Fossilien geborgen; dies führte zu einigen wichtigen Entdeckungen, die zur überregionalen Bekanntheit des Fayyum-Gebietes beitrugen. Zusätzlich erfolgten auch erste Untersuchungen zur Geologie. Die moderne Forschung begann in den 1960er Jahren und hält bis heute an. Sie ist international organisiert. Analysen des Fundmaterials führen fast im Jahresschritt zur Beschreibung neuer Fossilformen aus dem Fayyum.
Geographische Lage
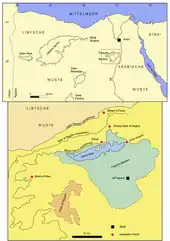
Die Fayyum-Region erstreckt sich über eine Fläche von rund 12.000 km² und befindet sich rund 70 km südwestlich von Kairo westlich des Nils inmitten der Libyschen Wüste. Das Zentrum bildet das Fayyum-Becken, ein etwa 1500 km² großes Depressionsgebiet. Es weist eine im Umriss dreieckige Form auf mit nach Süden zeigender Spitze. Zentralort ist al-Fayyūm, der zudem auch die Hauptstadt des Gouvernements al-Fayyum bildet. Im Norden des Beckens liegt der etwa 230 km² große Qarun-See, ein abflussloser See, der eine Länge von 40 km von Nordost nach Südwest aufweist bei einer durchschnittlichen Tiefe von rund 5 m und einer maximalen von 7,5 m. Als einzelne Landmasse ragt die Insel Geziret el-Qorn aus dem Wasser. Der Qarun-See (deutsch für „See der Hörner“, nach den beiden spitzen Halbinseln, die vom Norden in den See hineinreichen) ist der größte See des nördlichen Afrikas, er stellt aber nur den Rest eines noch größeren Sees dar. Dieser bestand in prähistorischer und historischer Vergangenheit und trägt die Bezeichnung Moeris-See. Die Depression senkt sich von Süd nach Nord ab. Befindet sich der Zentralort al-Fayyūm noch bei etwa 24 m über dem Meeresspiegel, so liegt die Wasserfläche des Qarun-Sees bei etwa 45 m unter dem Meeresspiegel.[1][2][3][4][5] Das gesamte Fayyum-Becken wird landwirtschaftlich genutzt, zwei Wassersammler (El Bats im Osten und El Wadi im Westen) drainieren das überschüssige Wasser in den Qarun-See.[6]
Die Umgebung des Fayyum-Beckens besteht weitgehend aus wüstenartigem Gelände. Im Osten trennt ein 5 bis 12 km breiter Rücken die Depression vom Niltal ab, seine Oberkante liegt bei etwa 30 bis 90 m über dem Meeresspiegel.[7] Diese Randberge zeigen einen etwa 1,5 km breiten Durchbruch, durch den auch der Bahr Yusuf verläuft. Der Kanal verbindet auf einer Länge von 270 km den Nil mit der Fayyum-Senke. Er endet bei al-Fayyūm und verästelt sich hier in zahlreiche kleine Arme. Im Süden folgt der kleine Kessel von el-Gharaq, der eine Tiefe von maximal 4 m unter der Meeresoberfläche besitzt. Ihm schließt sich im Südwesten und Westen des Fayyum-Beckens das Wadi el-Rayyan an. Hierbei handelt es sich um eine rund 700 km² große Senke mit dem tiefsten Punkt bei etwa 42 m unter dem Meeresspiegel. Das Wadi wird von eigenen Quellen gespeist und ist heute teilweise vom el-Fayyum-See ausgefüllt, einem zweigeteilten künstlichen See von 51 beziehungsweise 62 km² Größe. Bedeutend ist jedoch die Nordgrenze des Fayyum-Beckens, wo sich mehrere Steilstufen erheben. Sie verlaufen von Südwest nach Nordost über eine Länge von rund 70 km und sind durch plateauartige Ebenen voneinander getrennt. Das Gelände steigt so vom Qarun-See mit seiner niedrigsten Lage auf durchschnittlich 350 m über den Meeresspiegel an. Die wichtigsten Steilstufen sind von Süd nach Nord das Birket Qarun Escarpment, das Qasr el-Sagha Escarpment und das Gebel Qatrani Escarpment. Dazwischen gibt es noch kleinere Stufen wie etwa das el-Ekhwat Escarpment und das Talata Escarpment.[1][2][3][4][5]
Das Südwestende der Steilstufen markiert das Wadi el-Hitan („Tal der Wale“, auch Zeuglodon valley), rund 50 km von Qarun-See entfernt. Das Gebiet ist seit dem Jahr 2005 als UNESCO-Weltnaturerbe ausgezeichnet. Es handelt sich um ein breites, flaches, rund 10 km langes Tal, das kein tatsächliches Wadi im geomorphologischen Sinn darstellt. Es wird im Nordwesten vom Birket Qarun Escarpment begrenzt. Im Südwesten dagegen bildet eine Reihe von flachgipfligen Hügeln den Abschluss (die sogenannten „Grenzhügel“). Durch diese beiden geographischen Grenzen engt sich das Tal von etwa 4 km Breite im Nordosten auf rund 0,5 km im Südwesten ein. Etwa 5 km weiter nordöstlich in Richtung des Fayyum-Beckens erstreckt sich eine weitere Gruppe von Hügeln, unter ihnen erhebt sich der Garet Gehannam als wichtige Landmarke etwa 120 m über die Umgebung.[3][5]
Geologie
Allgemein und Untergrund
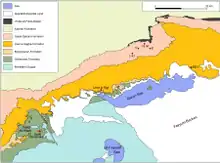
Die geologische Situation der Fayyum-Region wird seit dem Ende des 19. Jahrhunderts intensiv erforscht. Bedeutende Arbeiten dazu wurden 1905 von Hugh John Llewellyn Beadnell,[1] 1988 von Thomas M. Bown und Mary J. Kraus[2] sowie 1992 von Philip D. Gingerich[3] veröffentlicht. Das Fayyum-Becken stellt eine tektonisch entstandene Struktur dar, die in den triassischen Untergrund eingetieft ist. Gesteinsschichten aus dieser Zeit wie etwa der Bahariya-Formation treten aber nur in den äußersten Randlagen der Region an die Oberfläche, etwa im Nordosten. Die Hauptstörungen verlaufen von Südwest nach Nordost und gehören dem Syrian arc fold belt an, der in einem S-Bogen von der Levante über die Sinai-Halbinsel nach Nordafrika zieht. Sie bedingen auch die Entstehung der großen Steilstufen nördlich des Qarun-Sees. Untergeordnet kommen noch von Nordwest nach Südost sowie von Ost nach West gerichtete Störungszonen vor. Sie rahmen das Fayyum-Becken auf allen Seiten ein. Dadurch kann angenommen werden, dass das Becken durch tektonische Zugkräfte entstanden ist. Diese formten die Depression im Übergang vom Oligozän zum Miozän heraus, möglicherweise in Verbindung mit lokalem Vulkanismus. In der Region selbst sind vor allem Ablagerungen des Eozän und des Oligozän ausgebildet, deren Gesamtmächtigkeit bis zu 800 m beträgt.[8][9]
Mokattam-Gruppe

Das Liegende der Fayyum-Region wird allgemein der Mokattam-Gruppe (auch Wadi-el-Rayyan-Serie) zugewiesen, benannt nach dem Gebel Mokattam östlich von Kairo. Sie ist marinen Ursprungs und besteht aus vier Gesteinseinheiten, der Muweilih-Formation, der Midawara-Formation, der Sath-el-Hadid-Formation und der Gharaq-Formation. Alle vier Formationen setzen sich aus Kalksteinen zusammen, die sich aber in ihrer Struktur und in dem Gehalt von Nummuliten und Glaukoniten unterscheiden. Der obere Abschluss der Mokattam-Gruppe ist durch einen rapiden Rückgang an Numuliten und einen Anstieg an Glaukoniten definiert. Die Gharaq-Formation ist die am weitesten aufgeschlossene Einheit der Mokattam-Gruppe in der Region und findet sich zumeist im südwestlichen Fayyum-Gebiet vom Wadi el-Rayyan bis zum südlichen Teil des Wadi el-Hitan und bis zum Garet Gehannam.[10][5]
Maadi-Gruppe
Die Gehannam-Formation (auch Ravine beds) ist das Basisglied der Maadi-Gruppe. Sie besteht hauptsächlich aus marinen Tonen, Schluffen und sandigen Schluffen, in die mergelige und tonige Kalksteine eingearbeitet sind. Ihre Gesamtmächtigkeit liegt bei etwa 46 m. Die Basis besteht aus glaukonitischen Ton-/Schluffsteinen, die fein laminiert sind und nach oben in kalkreiche Sandsteine übergehen. Darauf folgen Schluff-, Kalk- und Mergelsteine. An einigen Aufschlüssen stellt wiederum ein Kalkstein den oberen Abschluss dar. Teilweise sind in den oberen Abschnitten der Gehannam-Formation einzelne Gipslagen eingebettet. Besonders gut ist die Gehannam-Formation im westlichen Fayyum-Bereich beim Wadi el-Hitan und am Garet Gehannam aufgeschlossen. Sie formt außerdem den Untergrund des größten Teils des kultivierten Landes im Becken.[3][10][5][11]

Alle anderen Formationen sind weitgehend in den Steilhängen nördlich des Qarun-Sees verbreitet. Die Birket-Qarun-Formation bildet den unteren, unmittelbar nördlich an den Qarun-See anschließenden Steilhang aus, das Birket Qarun Escarpment. Sie stellt sich als bis zu 85 m mächtige Gesteinseinheit dar, deren Hauptkomponenten sich aus feinen bis sehr feinen Sanden und Schluffen zusammensetzen, die mitunter stark kalkhaltig sind. Basal besteht ein Sandstein reich an Seeigeln, der als Schizaster-lybica-Sandstein bezeichnet wird. In einzelnen Bereichen sind Gipsausfällungen ausgebildet. Allgemein gehen die Ablagerungen auf ein küstennahes Meer zurück. In den Aufschlüssen nördlich des Qarun-Sees zeigen sie sich als einheitlicher Sedimentkomplex, dieser fächert nach Westen zum Wadi el-Hitan hin auf und verzahnt sich in seinen unteren und mittleren Abschnitten mit der Gehannam-Formation. Hier kann die Birket-Qarun-Formation in vier Einzelkörper untergliedert werden, die sich in der Regel eindeutig von der Gehannam-Formation abtrennen lassen. Als auffällige Erscheinung tritt eine Serie von schmalen, teils nur 30 cm mächtigen, blockartigen Sandsteinlagen auf, deren oberste als Camp White Layer bezeichnet wird. Letztere ist sehr markant, da sie neben zahlreichen Walknochen auffällige senkrechte Strukturen enthält, die wahlweise als Wurzeln von Mangroven oder Grabspuren der Spurengattung Thalassinoides gedeutet werden (das Camp White Layer wird in einzelnen älteren Arbeiten als Grenzschicht zwischen der Gehannam- und der Birket-Qarun-Formation aufgefasst,[3] jüngere Untersuchungen sehen es aber innerhalb der oberen Gesteinseinheit).[10][5][11]
Die Garet-el-Naqb-Formation wurde als mögliche Gesteinsformation erst im Jahr 2014 definiert, als besondere Bildung fiel sie schon vorher auf.[3][12] Es handelt sich um einen dunkelgrauen Ton, dessen Mächtigkeit von Süd nach Nord ansteigt. Seine größte Dicke erreicht er nördlich des Wadi el-Hitan mit rund 25 m. Die Gesteinseinheit kommt weitgehend nur im westlichen Fayyum-Gebiet vor, nördlich des Qarun-Sees ist sie nicht dokumentiert. In den Aufschlüssen des westlichen Fayyum teilt die Garet-el-Naqb-Formation als markant dunkler Horizont die Birket-Qarun-Formation oder liegt der Gehannam-Formation direkt auf.[5][11]

Die Qasr-el-Sagha-Formation (auch Carolia beds) schließt die Maadi-Gruppe ab. Sie ist am Qasr el-Sagha Escarpment aufgeschlossen und wird bis zu 200 m mächtig. Die Gesteinseinheit besteht aus Sandsteinen, sandigen Ton-/Schluffsteinen und kalkhaltigen geschichteten Gesteinen mit teilweise dazwischen geschalteten Lagen von Kalksteinen. Es lassen sich vier Untereinheiten auseinanderhalten. Zuunterst liegt das Umm Rigl Member mit einer 30 bis 65 m mächtigen Sequenz aus sandigen, teils bioklastischen, harten Kalksteinen, die sich mit gips- und kalkhaltigen, feingeschichteten Ton- und Schluffsteinen abwechseln. Darauf folgt das Harab Member, das sich als 30 bis 40 m dickes Paket aus braunen, strukturlosen Tonen darstellt. Das Temple Member wiederum ähnelt in seiner Struktur dem Umm Rigl Member. Das hangende Dir Abu Lifa Member wird bis zu 77 m mächtig und charakterisiert sich durch eine Wechsellage aus kreuzgeschichteten Sandsteinen und feinkörnigen, gipshaltigen Ton-/Schluff- und Sandsteinen. Die Ablagerungen gehen auf ein flaches, küstennahes Meer zurück. Die Basis der Formation wurde in der Vergangenheit nicht einheitlich beurteilt. Einige Autoren legten sie mit dem ersten Auftreten der Muschel Carolia fest, andere mit einem tonhaltigen Kalk, der einige Meter höher liegt.[3][12] Der obere Übergang zur nächsten Gesteinseinheit ist diskontinuierlich.[2][3][5][11]
Gebel-Qatrani-Formation
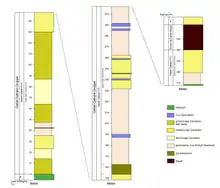

Der Qasr-el-Sagha-Formation liegt diskordant die Gebel-Qatrani-Formation (auch fluvio-marine series) auf. Sie erreicht eine Mächtigkeit von 340 m und bildet mit dem Gebel Qatrani Escarpment den höchstgelegenen Steilhang. Nach Westen zu dünnt die Formation deutlich aus und wird dann teilweise weniger als 100 m mächtig. Sie hebt sich von der Qasr-el-Sagha-Formation durch einen Aufbau aus verschiedensten feinen bis gröberen Sandsteinen sowie Ton-/Schluffsteinen ab. Prinzipiell sind drei Zonen unterscheidbar: eine untere Zone aus groben, lockeren Sandsteinen (die lower fossil wood zone, Mächtigkeit 153 m), eine mittlere aus einem bankigen, mittel-bis grobkörnigen und teils extrem gipshaltigen Sandstein als Markerhorizont (Mächtigkeit 7 bis 10 m) und eine obere, bestehend aus einem Wechsel aus Sandstein, sandigem Ton-/Schluffstein mit eingearbeiteten Linsen aus groben Sandstein und Kalkstein (die upper fossil wood zone, Mächtigkeit 190 m). Die Bezeichnungen lower und upper fossil wood zone gehen auf die hier gefundenen fossilisierten Baumstämme zurück. Sowohl die untere als auch die obere Serie lassen sich in zahlreiche Untereinheiten auflösen, besonders hervorzuheben sind mehrere Sequenzen farbig gebänderter Sedimentfolgen, die sich über die gesamte Gebel-Qatrani-Formation verteilen und als variegated sequences bezeichnet werden. Insbesondere in der upper fossil wood zone werden sie bis zu 70 m mächtig. Die Bänderungen gehen auf fossile Bodenbildungsprozesse und die damit verbundene Verlagerung und Anreicherung von löslichen Eisenverbindungen zurück. Darüber hinaus gibt es weitere Hinweise auf Paläoböden wie Tonanlagerungen. Die Böden lassen verschiedene Entwicklungsstadien von initialen (Inceptisole) bis zu entwickelten Bodenbildungen (Spodosole und Ultisole) erkennen. Insgesamt verweisen die sekundären Bodenüberprägungen auf ein eindeutig terrestrisches Ablagerungsmilieu der Gebel-Qatrani-Formation hin. Sie kann daher als kontinental-fluviatile Sedimentfolge angesehen werden, die durch das Wirken zahlreiche Flüsse und Wasserläufe entstanden ist.[13][2]
Widan-el-Faras-Basalt
Der obere Abschluss der eozän-oligozänen Serie wird durch den Widan-el-Faras-Basalt markiert, von der liegenden Gebel-Qatrani-Formation ist er durch eine Schichtlücke getrennt. Er besteht aus dichten, extrem feinkörnigen und eisenhaltigen Basalten. Einzelne eingeschaltete Sandhorizonte zeigen, dass die Basalte mehrere Ströme formen, deren Dicke und Ausprägung aber stark variabel sind. Die größte Dicke wird mit 25 m erreicht, stellenweise kann der Widan-el-Faras-Basalt auch nur 2 m mächtig werden.[2]
Deckschichten
Auf der paläogenen Sedimentfolge lagern die neogenen Deckschichten. Sie werden im Fayyum-Gebiet der Kashab-Formation zugeordnet, die sich wiederum von den Widan-el-Faras-Basalten durch einen Hiatus abhebt. In Gebieten, in denen der Basalt nicht ausgebildet ist, bedeckt sie direkt die Gebel-Qatrani-Formation. Hauptsächlich besteht die Kashab-Formation aus Schwemmsedimenten, die unteren 10 bis 12 m werden durch ein geröllhaltiges Konglomerat angezeigt. Ihre Gesamtmächtigkeit beträgt über 100 m. Die Kashab-Formation bildet das nördliche Hinterland der Fayyum-Region.[2]
Entwicklung der Fayyum-Senke im Pleistozän und Holozän
Während der wechselnden Kalt- und Warmzeitphasen des Pleistozän formte sich die Gestalt des heutigen Fayyum-Beckens heraus. Die extremen Trockenperioden während der Kaltzeiten führten mehrfach zur Ausräumung des Kessels durch Deflation bis auf den Grundwasserspiegel. In den feuchteren Warmzeiten füllte sich das Becken mit Wasser und bildete ein Sumpfgebiet. Diese Sumpfgebiete waren noch bis in historische Zeit nachweisbar und führten auch zur Namensgebung der Fayyum-Region (vom ägyptischen phiom für „Sumpf“). Die jüngste Bildung des Holozän wird als Moeris-See bezeichnet. Seine natürliche Entwicklung begann bereits im Unteren Holozän und verlief in mehreren Phasen, die sich vor allem anhand der Verbreitung von Kieselalgen, aber auch alter Uferlinien nachweisen lassen. Sie führten zu schwankenden Seespiegellagen, was teilweise mit der Entstehung und dem Abbruch der natürlichen Verbindung zum Nil zusammenhängt. Die älteste stellt der „Paläomoeris“ dar, er bestand von 8.830 bis 8.220 Jahre BP. Während dieser Zeit erreichte der Seespiegel einen Höchststand von 17 m über dem Meeresspiegel. Ihr folgte die „Prämoeris“- (zwischen 8.200 und 7.500 Jahre BP) und der „Protomoeris“-Phase (7.500 bis 7.140 Jahre BP), während der der Seespiegel zeitweise zwischen 28 und 8 m über dem Meeresniveau schwankte. Die letzte Phase umfasst die „Moeris“-Phase, sie begann etwa um 6.095 BP und hielt bis etwa 5.500 Jahre BP an. Hier lag der Seeoberfläche bei bis zu 20 m über dem Meeresspiegel und formte einen See mit einer Fläche von 2100 km² und einem Volumen von 53 km³.[14] In der Folgezeit fiel er aber wieder durch den Verlust der Verbindung zum Nil um mehrere Meter ab, und es entstand ein sumpfiges Gelände.[7][6][15][14]
In der altägyptischen Zeit wurde unter anderem durch wasserwirtschaftliche Baumaßnahmen Neuland zugänglich und urbar gemacht. Dies geschah beispielsweise im Mittleren Reich durch den Bau des Bahr Yusuf entlang des alten Nilzugangs und durch ein Drainagesystem im Fayyum-Becken. Auf diese Weise konnten ungefähr 450 km² Land genutzt werden. Später kamen die Errichtung eines 8 km langen Absperrdamms bei el-Lahun und die Schaffung weiterer Frischhaltebecken im Südosten des Fayyum-Gebietes hinzu. Bis zur ptolemäischen Zeit wurde so die nutzbare Fläche auf rund 1300 km² ausgedehnt, was einem Großteil des heute kultivierten Landes entspricht. Der Moeris-See war zu dieser Zeit auf rund 415 km² geschrumpft und nahm den Bereich des heutigen Qarun-Sees ein.[16] Weitere Rückgänge prägten die römische Epoche, während der die Wasseroberfläche von 7 m unter dem Meeresspiegel auf 17 m absank. In der ersten Hälfte des 19. Jahrhunderts lag sie bereits bei rund 40 m, während der Tiefpunkt mit 46 m unter dem Meeresspiegel in den 1930er Jahren erreicht wurde. Der Qarun-See entwickelte sich dadurch auch von einem einstigen Süßwasserreservoir zu einem stark salzhaltigen Gewässer, was einerseits durch den gegenüber der Verdunstung geringeren Wasserzufluss bedingt, andererseits durch die Einleitung von chemisch belasteten Abwässern der Landwirtschaft noch verstärkt wird. Abseits davon stellt der See ein wichtiges Refugium für Zug- und Wasservögel dar. Er dient als Brutgebiet des Hirten- und des Seeregenpfeifers sowie der Rotflügel- und der Zwergseeschwalbe, während der Hauben- und der Schwarzhalstaucher hier überwintern.[4]
Fossilfundstellen


Die Fossilien stammen aus mehreren geologischen Formationen. Pflanzenfossilien treten in größerer Anzahl nur in der Gebel-Qatrani-Formation auf. Dagegen können Wirbellose in nahezu allen Gesteinseinheiten angetroffen werden, während Wirbeltiere wiederum stärker beschränkt sind. Marine Formen verteilen sich auf die unteren Einheiten und kommen häufig in der Muweilih-, der Gharaq-, der Gehannam- und der Birket-Qarun-Formation vor, sie sind aber auch aus der Qasr-el-Sagha-Formation überliefert. Mitunter erhielten sich vor allem die größeren Meeressäuger in vollständigen Skeletten.[11] Die Qasr-el-Sagha-Formation und die folgende Gebel-Qatrani-Formation stellen die Hauptfundgebiete der terrestrischen Lebewesen dar, das Material ist überwiegend fragmentiert und disartikuliert, so dass von gewissen Umlagerungen auszugehen ist. Im gesamten Fayyum-Gebiet bestehen unzählige Fundstellen und Aufschlüssen. Sie liegen in den verschiedenen stratigraphischen Einheiten und konzentrieren sich in der Regel nördlich und westlich des Qarun-Sees. Allein für die terrestrischen Säugetiere sind über 100 Fundstellen belegt. Diese verteilen sich auf einige vereinzelte in der Birket-Qarun-Formation und etwas weniger als ein Dutzend in der Qasr-el-Sagha-Formation. Der weitaus größere Teil von über 90 % gehört der Gebel-Qatrani-Formation an.[17][18][19] Von großer Bedeutung ist die Fundstelle BQ-2 aus dem Umm Rigl Member der Qasr-el-Sagha-Formation (einzelne, vor allem paläontologische Studien verweisen das Umm Rigl Member zur Birket-Qarun-Formation,[19] was auf ungenügende Aufschlussbedingungen an der Fundstelle selbst zurückgeführt wird[5]) nahe der östlichsten Spitze des Qarun-Sees. BQ-2 wurde erst im Jahr 2000 entdeckt, von dort stammt die bisher am stärksten diversifizierte Wirbeltierfauna des gesamten Fayyum-Gebiets.[19] Die meisten anderen Fundplätze der Qasr-el-Sagha-Formation sind, mit Ausnahme einer unstratifizierten Fundstelle aus dem Temple Member, dem Dir Abu Lifa Member zuzuweisen. Generell treten aber landlebende Säugetiere hier seltener auf.[17][18]
Von den Fundstellen der Gebel-Qatrani-Formation wiederum sind acht (A, B, E, G, I, M, V und L-41) von besonderem Interesse, da diese rund 90 % des gesamten Fundmaterials terrestrischer Wirbeltiere enthalten.[17] Die bedeutendste Fundstelle hier findet sich mit L-41. Sie wurde 1983 vom Geologen Thomas M. Bown entdeckt. Bown war an den von Elwyn L. Simons seit den 1960er Jahren hier getätigten Forschungsarbeiten beteiligt, er wurde durch zwei Unterkiefer von Schliefern auf diese Fundlokalität aufmerksam. Eine genauere Beschreibung der Fundstelle legte einige Jahre später unter anderem D. Tab Rasmussen vor. Mit einer Lage von 47 m oberhalb der Basis der Gesteinseinheit markiert L-41 die bisher älteste Fundstelle der Formation, zudem gilt sie als die ertragsreichste.[20][21] In der geologischen Folge etwas höher, aber immer noch im Bereich der unteren Zone, sind die Fundstellen A, B und E lokalisiert. Sie wurden bereits zu Beginn des 20. Jahrhunderts geöffnet und bilden die forschungsgeschichtlich ältesten stratifizierten Fossillager. Ebenfalls eine gewisse Bedeutung weist die Fundstelle G auf, die etwa 16 m oberhalb der Sandsteinbank liegt, welche die untere von der oberen Zone trennt. Die hauptsächlichen Untersuchungen fanden in den 1960er Jahren statt. Noch einmal 66 m höher finden sich die beiden Fundstellen I und M, die demnach zu den jüngsten innerhalb der Gebel-Qatrani-Formation gehören.[22][23]
Generell sind die Fossilreste diagenetisch überprägt und permineralisiert, so dass das organische Material durch Anhydrit und Gips ersetzt wurde. Allerdings scheinen die Knochen und Zähne aus dem Dir Abu Lifa Member der Qasr-el-Sagha-Formation einen weiteren diagenetischen Prozess durchlaufen und zu haben. Vielfach wurden hier die Sulfate durch Karbonate ersetzt. Dadurch erhielten sich die Oberflächenstrukturen der Knochen und Zähne weniger gut als die Funde aus der Gebel-Qatrani-Formation.[18]
Funde
Flora

Makroreste von Pflanzen liegen in Form von Hölzern, Blättern, Früchten und Samen vor. Vor allem aus den älteren Gesteinseinheiten sind gelegentlich Blattreste aus der Gruppe der Cymodoceaceae belegt, so unter anderem von den „Seegräsern“ Thalassodendron und Cymodocea.[10] Ansonsten beschränken sich Pflanzenreste weitgehend auf die Gebel-Qatrani-Formation. Bei den Hölzern treten verschiedenste Größen auf, angefangen von kleinen Zweigen bis zu 44 m langen und über 2 m dicken Baumstämmen. An einigen Stämmen ist noch die Rinde vorhanden. Teilweise liegen diese Baumstämme als fossilisierte Wälder zusammen, die unter anderem in den unteren Abschnitten der Gebel-Qatrani-Formation aus bis zu 200 Einzelstücken bestehen. Die Vegetation umfasst rund zwei Dutzend verschiedene Familien, unter den Hölzern beziehungsweise den Früchten und Samen konnten jeweils acht und unter den Blättern 13 identifiziert werden. Typisch sind unter anderem die an stehende oder langsam fließende Gewässer gebundenen Schwimmfarne wie Salvinia, zudem auch Mangrovenfarne wie der im Brackwasser lebende Acrostichum. Unter den Einkeimblättrigen heben sich die Palmenartigen hervor, von denen unter anderem die ausgestorbene Gattung Palmoxylon beschrieben wurde.[24] Weitere Einkeimblättrige finden sich mit den Efeututen aus der Gruppe der Aronstabgewächse, die lianenartig an Bäumen klettern. Deren fragile Früchte können teilweise noch im intakten Zustand geborgen werden. Am häufigsten sind jedoch Zweikeimblättrige nachgewiesen, darunter etwa Lotosgewächse, Malvengewächse, Johannisbrotgewächse oder Sapotengewächse. Die gesamte Flora zeigt Beziehungen zum indo-malaysischen Raum.[25][13][2][26]
Wirbellose
Wirbellose Tiere finden sich in allen Gesteinsschichten, in der Regel handelt es sich dabei um Reste der Meeresfauna. Einen Großteil davon nehmen die Foraminiferen ein, gehäusetragende Einzeller, die im Fayyum sowohl als planktonische wie auch als benthische Formen auftreten. Planktonische Foraminiferen werden durch Truncorotaloides, Turborotalia oder Globigerinatheka repräsentiert, benthische durch Operculina Discocyclina oder Nummulites. Einige Formen wie Frondicularia oder Pullenia verweisen auf recht große Meerestiefen. In einzelnen Gesteinsabschnitten sind die Foraminiferen mit bis zu über 30 Arten recht zahlreich.[27][28][5] Andere Wirbellose gehören zu den Seeigeln wie beispielsweise Schizaster, ein Vertreter der Herzseeigel, oder seltener Lanzenseeigel. Daneben sind auch Weichtiere belegt. Unter den Muscheln haben Austern, Mondmuscheln und Sattelmuscheln hohe Bedeutung, bei letzteren bildet Carolia einen wichtigen stratigraphischen Anzeiger. An Schnecken lassen sich wiederum Mondschnecken und Turmschnecken vorweisen. Darüber hinaus kommen Nesseltiere vor. Gelegentlich sind Krebse überliefert, wie einzelne Gehäuseteile von Lobocarcinus aus der Gruppe der Taschenkrebse in der mittleren Birket-Qarun-Formation zeigen. Die Panzer haben Breiten von 9 bis 14 cm, wobei in der Panzerform markante Geschlechtsunterschiede vorliegen.[29][5]
Fische und Amphibien
Fische zeigen ebenfalls eine weite stratigraphische Verbreitung im Fayyum-Gebiet. Der überwiegende Teil des Fundmaterials besteht aus Zähnen und vereinzelten isolierten Knochen, gelegentlich treten auch artikulierte Skelettreste auf. Meeresbewohnende Fische kommen in allen geologischen Abschnitten der Fayyum-Region, sie sind aber vermehrt in der Mokattam-Gruppe und in den unteren Bereichen der Maadi-Gruppe zu beobachten. Die Fauna wird hauptsächlich durch Haie und Rochen geprägt. Große Haie finden sich mit dem Tigerhai, der beispielsweise in der Midawara-Formation sehr häufig ist. Andere Formen sind Scharfnasenhaie, Hammerhaie und Makrelenhaie, bei letzteren unter anderem ein bis zu 5 m langer Vertreter der Gattung Otodus.[30] Die umfangreichsten Funde können aber den Zitronenhaien zugewiesen werden, die mit einer größeren und kleineren Form vorkommen und in der Maadi-Gruppe ihren Verbreitungsschwerpunkt haben. Daneben wurde noch eine Vielzahl an mittelgroßen und kleineren Haien belegt, etwa Ammenhaie, Fuchshaie, Sandhaie, Sechskiemerhaie und Glatthaie. Auffallend ist die Gattung Moerigaleus, die typisch für Wieselhaie über ein Gebiss mit variantenreich gestalteten Zähnen verfügte, von denen bisher über 60 bekannt sind.[31] Mit dem Fossilhai, der ebenfalls zu dieser Gruppe zählt, tritt des Weiteren eine Form auf, dessen heutige Angehörige im Schelfgebiet zumeist Kopffüßer jagen. Zu nennen sind auch einige Requiemhaie wie Carcharhinus und Misrichthys, die eine gewisse Süßwasserkomponente bilden und in vergleichsweise größerer Anzahl in den oberen Abschnitten der Maadi-Gruppe auftreten. Ein vermehrter Fossilnachweis in diesen geologischen Schichten ist auch für das verwandte Abdounia zu verzeichnen. Große Rochen gehören zumeist der Gruppe der Stechrochenartigen an, so unter anderem Kuhnasenrochen, die in der Midawara- und in der Qasr-el-Sagha-Formation recht häufig auftreten, dazwischen aber nahezu verschwinden. Andere Formen wie Leidybatis oder Lophobatis sind demgegenüber generell selten. Weiter nachgewiesen wurden Propristis und Anoxypristis, die die Sägerochen repräsentieren. Dagegen gehören Coupatezia und Hypolophodon in die Verwandtschaft der Stechrochen, sie verweisen in den oberen Abschnitten der Maadi-Gruppe wieder auf einen stärkeren Süßwassereinfluss. Ein sehr rares Faunenelement findet sich mit den Zitterrochen.[32][33][34]

Generell treten marine Elemente in den oberen Abschnitten der Maadi-Gruppe ab der Qasr-el-Sagha-Formation in den Hintergrund und es kommen vermehrt Süßwasser- oder Brackwasserfische vor. Hier dominieren die Knochenfische mit variantenreichen Formen. Noch aus der Gehannam-Formation liegen aber Reste von Pycnodus aus der ausgestorbenen Gruppe der Pycnodontiformes vor, der sich offensichtlich von hartschaligen Muscheln ernährte.[30] Aus den jüngeren Ablagerungsserien konnten dann beispielsweise die Flösselhechte mit Polypterus belegt werden. Hervorzuheben sind auch der Großnilhecht und Welsartige wie Chrysichthys und Auchenoglanis. Ein nahezu vollständiges, gut 23 cm langes Individuum liegt von Chasmoclupea aus der Gruppe der Heringe vor.[35] Kreuzwelse wiederum geben Hinweise, dass es noch einen marinen Einfluss gab, was unter anderem durch die Gattung Qarmoutus mit einem weitgehend vollständigen Skelett dokumentiert wurde.[36] Dafür spricht auch der Nachweis einzelner Schädelteile von Xiphiorhynchus, einem Verwandten des Schwertfischs, das längste Stück misst etwa 33 cm.[37] Die Gattung Lates ist wiederum mit mehreren Schädeln überliefert, sie stellen den ältesten Beleg dieses Vertreters der Riesenbarsche in Afrika dar.[38] Die gleiche Aussage trifft auf Parachanna zu, einen Vertreter der Schlangenkopffische, von dem aus dem Fayyum ebenfalls einzelne Schädelteile überliefert sind.[39] Daneben kommen Buntbarsche, Afrikanische Salmler und Lungenfische vor.[40][41][33][42]
Amphibien sind bisher kaum untersucht. Aus der Gebel-Qatrani-Formation wurden Reste von Fröschen berichtet.[41]
Reptilien und Vögel
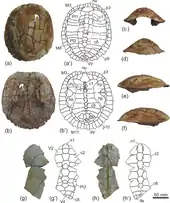
Die Reptilien sind wiederum sehr divers und mit allen heute noch bestehenden Ordnungen vertreten. In der Regel liegen die Reste in der Qasr el-Sgha- und in der Gebel-Qatrani-Formation, nur selten in älteren Gesteinsschichten. Das umfangreiche Material der Schildkröten setzt sich überwiegend aus den Panzerresten zusammen, jedoch liegen auch Schädel und Elemente des Körperskeletts vor. Es kommen Vertreter aus den beiden heutigen Ordnungen vor. Zur Gruppe der Halsberger-Schildkröten kann Gigantochersina verwiesen werden, eine Landschildkröte von der Größenordnung der Galapagos-Schildkröten. Von dieser Gattung sind mehrere vollständige Panzer von bis zu 88 cm Länge erhalten. Anfänglich wurden die Funde den Paläarktischen Landschildkröten zugesprochen.[43] Bei den Halswender-Schildkröten sind unter anderem die Gattungen Cordichelys, Dacquemys, Albertwoodemys, Shetwemys und Stereogenys hervorzuheben. Alle gehören zur Familie der Podocnemididae, welche vornehmlich Süßwasserformen enthält. Es handelt sich meist um kleine bis mittelgroße Formen, als große Form hat Stereogenys eine Panzerlänge von bis zu 46 cm.[44][45][46][47]
Von den Schuppenkriechtieren liegen rund 100 Wirbelfragmente vor. Zu ihnen gehören unter anderem die ältesten Vertreter der Warane. Ebenso tritt noch eine etwas urtümlichere, bisher unbestimmte Form auf. Die Funde lassen annehmen, dass die gesamte Gruppe in Afrika ihren Ursprung hat.[48][49] Auch die Schlangen sind überwiegend über Wirbel belegt, das überwiegende Material wird der Qasr el-Sagha-Formation zugewiesen. Unter ihnen beeindruckt vor allem Gigantophis, eine Riesenschlange, deren Länge mit durchschnittlich 6,9 m angegeben wird, große Exemplare mögen 9 bis 11 m Länge erreicht haben. Wahrscheinlich lebten die Tiere unterirdisch. Bisher ist kein Schädelmaterial bekannt, so dass über die Ausmaße und Dehnungsfähigkeit des Mauls und somit über die Beutegröße nichts ausgesagt werden kann; andere Vertreter der Madtsoiidae besaßen aber nicht die Anpassung an eine extreme Mauldehnung wie bei heutigen Riesenschlangen. Pterosphernus war dagegen an ein Leben im Wasser angepasst, worauf die seitlich stark gepressten Wirbel schließen lassen. Auch sie erreichte große Ausmaße. Neben diesen beiden häufigsten Vertretern kommen noch einige kleinere, bisher unbestimmte Boas vor, ebenso wie Erdboas. Mit Renenutet tritt auch eine Form der Nattern- und Vipernartigen auf, bisher sind aber nur drei Wirbel überliefert.[50][51]
Innerhalb der Krokodile lassen sich im Fayyum-Gebiet zwei Formengruppen unterscheiden. Die eine besteht aus langschnäuzigen Vertretern, die stammesgeschichtlich in der Nähe der heutigen Gaviale stehen. Als älteste Form ist Paratomistoma noch aus der Gehannam-Formation des Wadi el-Hitan anhand eines Schädels beschrieben worden. Ihre Fundlage in marinen Sedimenten lässt auf eine Anpassung an ein Meeresleben schließen. Alle anderen Funde sind stratigraphisch jünger. Von Bedeutung ist hier Eogavialis, dessen Fundmaterial ursprünglich dem rezenten Sunda-Gavial zugesprochen wurde.[52] Allerdings erwies sich Eogavialis als sehr basal stehend in der Entwicklung der Gaviale, möglicherweise gehört er in eine Linie noch vor der Aufspaltung in die heutigen südostasiatischen und die ausgestorbenen südamerikanischen Formen.[53] Die zweite Gruppe wird durch Tiere mit breiten Schnauzen repräsentiert, die wohl eine engere Beziehung zu den Echten Krokodilen zeigen. Zuerst der heutigen Gattung Crocodylus zugeordnet, handelt es sich hier aber sehr wahrscheinlich um stammesgeschichtlich ursprünglichere Angehörige.[54][55] Neben diesen Formen liefert ein Unterkieferfragment eines bisher unbenannten Mitglieds der Sebecosuchia einen der seltenen Nachweise dieser Gruppe aus Afrika. Gemeinsam mit einigen Funden aus Südamerika ist das Stück aus dem Fayyum auch einer der jüngsten Belege eines Vertreters aus dieser urtümlichen und entfernteren Krokodilsverwandtschaft.[56]
Fossilien von Vögeln sind eher spärlich mit einzelnen Bein- und Fußknochen, manchmal auch mit Schädelfragmenten überliefert. Den frühesten Nachweis der Vögel in der Region bildet ein Tibiotarsus aus der Birket-Qarun-Formation des Wadi el-Hitan. Für ihn wurde die Gattung Eopelecanus etabliert, er ist gleichzeitig einer der ältesten Belege für die Pelikane.[57] Die übrigen Funde entstammen fast ausnahmslos der Gebel-Qatrani-Formation und verteilen sich auf rund ein halbes Dutzend Ordnungen. Nur wenige Objekte lassen sich genau zuweisen. Hierzu gehören die Gattungen Nupharanassa und Janipes, die den Blatthühnchen innerhalb der Regenpfeiferartigen nahestehen. Die Schreitvögel dagegen repräsentiert Xenerodiops, der etwas kleiner als heutige Störche war und über einen Schädel und einen Oberarmknochen dokumentiert ist. Ein weiterer Schädel wird Palaeoephippiorhynchus zugesprochen, ebenfalls ein Verwandter der Störche. Einige andere Funde stehen wiederum mit der heutigen Gattung Nycticorax und damit mit den Reihern innerhalb der Ruderfüßer in Verbindung. Zur gleichen Verwandtschaftsgruppe gehören auch die Schuhschnabel, welche ein eher seltenes Fossilelement bilden. Sie sind mit Goliathia im Fayyum vertreten. Darüber hinaus waren Greifvögel (unter anderem mit den Habichtartigen und den Fischadlern) sowie Kranichvögel (mit den Rallen) anwesend, ebenso wie Turakos und Flamingos, hier ist das Material aber zu fragmentiert für genauere Aussagen. Neben den bekannten Vogelgruppen lassen sich einige überlieferte Formen nicht exakt einordnen. Zu nennen ist hier Eremopezus, ein wohl flugunfähiger Vogel mit den Ausmaßen heutiger Nandus. Dieser mit langen und schlanken Beinen ausgestattete Vogel, allein der Tarsometatarsus misst gut 34 cm, ähnelte keiner der bekannten Gruppen großer Laufvögel. Eine ursprünglich als Stromeria benannte ähnliche Form aus dem Fayyum-Gebiet galt anfänglich sogar als Basalform der madagassischen Elefantenvögel. Heute wird Stromeria als synonym zu Eremopezus aufgefasst. Wahrscheinlich handelt es sich um eine eigenständige Entwicklung innerhalb der Vogelwelt Afrikas.[58][59] Ein weiterer wohl flugunfähiger Vogel, angezeigt durch einen stark fragmentierten Beinknochen, könnte den ausgestorbenen Ameghinornithidae angehören; diese wurden bisher nur aus Europa beschrieben.[60]
Säugetiere
Die Säugetiere sind sehr umfangreich und mit mehr als einem Dutzend, zum Teil heute ausgestorbenen Ordnungen belegt. Sie kommen in allen Bereichen der Maadi-Gruppe vor. Zu unterscheiden ist zwischen meeresbewohnenden und landlebenden Formen. Erstere finden sich überwiegend in den unteren (Gehannam- und der Birket-Qarun-Formation), letztere in den oberen (Qasr-el-Sagha- und Gebel-Qatrani-Formation) Abschnitten. Die ursprünglichste Gruppe bilden die Beuteltiere, von denen wenigsten zwei Vertreter vorliegen. Dazu gehört Peratherium, das das erste in Afrika überhaupt nachgewiesene Beuteltier repräsentiert und in die nähere Verwandtschaftsgruppe der Beutelratten gehört.[61][62][63] In einer ähnlichen Beziehung steht wohl Ghamidtherium.[64] Beide Formen sind lediglich über mehrere Unterkiefer und einzelne Oberkieferreste bekannt. Die Zuweisung von Ghamidtherium zu den Beuteltieren wird aber teilweise in Frage gestellt, andere Autoren gruppieren die Reste eher mit den Insektenfressern.[65] Mit den Ptolemaiida wurde eine ausgestorbene Ordnung kleiner, wohl insektenfressend lebender Säugetiere eingeführt, deren systematische Stellung zunächst unklar war. Kennzeichnend sind die überdimensionierten mittleren Backenzähne von den hinteren Prämolaren bis zum vordersten Molar, während die anderen Backenzähne relativ klein bleiben. Wenigstens drei Gattungen wurden aus dem Fayyum beschrieben. Die bekannteste ist die Typusform Ptolemaia, deren Erstnachweise in Form von Gebissresten bereits vom Beginn des 20. Jahrhunderts stammen.[66] Später kamen auch Schädelfunde zu Tage. Weitere Formen aus dem Fayyum sind Qarunavus und Cleopatrodon.[67][68] Bemerkenswert bei den Funden der Ptolemaiida im Fayyum ist, dass sie fast alle in einem Umkreis von rund 1 km zum Vorschein kamen, so dass die Tiere möglicherweise ökologisch eng begrenzt waren. Heute werden die Ptolemaiidae mit den Afrotheria assoziiert.[69]
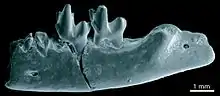
Die eigentlichen Afrotheria bilden eine der Hauptfundgruppen in der Fossilgemeinschaft von Fayyum. Von den kleinen, insektenfresseartigen Formen sind zumeist Gebissfragmente überliefert. Das gilt für Eochrysochloris als urtümlichem Angehörigen der Goldmulle ebenso wie für einige weitere, eher entfernte Verwandte der Tenrekartigen. Zu nennen wären hier etwa Dilambdogale, Widanelfarasia oder Qatranilestes. Sowohl die heutigen Goldmulle als auch die Tenrekartigen zeichnen sich durch drei markante Höckern und durch eine V-förmige Scherleiste auf jedem Molaren aus (zalambdodontes Zahnmuster), die drei letztgenannten fossilen Gattungen besitzen auf den vorderen Mahlzähnen aber noch eine urtümlichere W-förmige Scherleiste (dilambdodontes Zahnmuster).[70][71][72] Herodotius wiederum kann als ein sehr früher Vertreter der Rüsselspringer angesehen werden. Die Gattung basiert auf mehreren Schädel- und Unterkieferfragmenten. Ein weiterer Unterkiefer und ein Zahn sind von Metoldobotes bekannt, einer deutlich moderneren und größeren Form.[73][74]
Herausragend ist der Fossilbericht der Schliefer im Fayyum, allein in der Gebel-Qatrani-Formation stellen sie fast ein Drittel aller Säugetierfunde.[75] Die Schliefer gehören zu den wichtigsten biostratigraphischen Elementen des paläogenen Afrikas. Von Dimaitherium sind Schädel- und Körperelemente erhalten. Es repräsentiert mit seinem noch langgestreckten Schädel einen archaischen Schliefer, der Bau seines Fußes verweist auf eine mögliche kletternde Fortbewegung mit rapiden aber nicht ausdauernden Bewegungen.[76] Andere frühe Formen sind Geniohyus, Bunohyrax oder Pachyhyrax, häufig unterscheiden sie sich nur durch die Modifikationen der Mahlzähne von einem buckeligen (bunodonten) Kauflächenmuster hin zu einem mit einer mondsichelförmigen Scherleiste (selenodont).[77] Deutlich moderner wirken Saghatherium und Thyrohyrax, was durch den gekürzten Schädel angezeigt wird. Ersteres stellte möglicherweise einen schnellen Läufer dar. Bei beiden Vertretern sind an der Innenseite der Unterkiefer kammerartige Hohlräume ausgebildet, die nur bei männlichen Tieren auftreten, deren Funktion aber unklar ist.[78][21] Bei Antilohyrax lässt die Hinterfußkonfiguration ein sich wohl springend fortbewegendes Tier vermuten.[79] Die meisten der hier genannten Schliefer waren relativ klein und übertrafen die heutigen Arten nur geringfügig. Dem gegenüber erreichte Megalohyrax die Größe eines Esels, während Titanohyrax schätzungsweise über 800 kg wog und somit die Ausmaße eines kleinen Nashorns besaß. Von beiden sind vorwiegend Schädel- und Gebissreste erhalten.[80][77][69]

Eine der bekanntesten und wohl auch charakteristischsten Fossilformen der gesamten Fayyum-Region stellt Arsinoitherium dar, ein gewaltiges Tier mit bis zu 3,4 m Körperlänge. Nahezu das gesamte Skelett des Tieres ist von mehreren Dutzend Individuen bekannt. Dessen Hauptmerkmale finden sich in den zwei knöchernen Hornpaaren auf dem Schädel, von denen das größere vordere aus dem Nasenbein gebildet wurde. Ebenso auffallend ist das Gebiss mit 44 hochkronigen Zähnen, mit denen die wohl weiche Pflanzenkost zerquetscht wurde. Äußerlich ähnelte Arsinoitherium den Nashörnern, mit denen es dem Skelettbau nach nicht näher verwandt ist, es wird in die eigene, ausgestorbene Gruppe der Embrithopoda verwiesen.[81][15] Außerordentlich hohe Bedeutung erlangte die Fayyum-Region durch die Fossilien von Rüsseltieren. Sie gehören alle einer sehr frühen Entwicklungsphase an, deren gemeinsames Kennzeichen sich im vertikalen Zahnwechsel findet. Moeritherium bildete innerhalb dieser wohl einen Seitenzweig. Es zeigte sich äußerlich schweineartig und verfügte noch nicht über einen Rüssel. Dagegen wirkte das deutlich größere Barytherium schon eher wie ein klassisches Rüsseltier, es besaß aber insgesamt acht kleine Stoßzähne. Beide Formen waren ausgesprochene Sumpfbewohner. Zwei weitere nachgewiesene Gattungen sind Palaeomastodon und Phiomia. Beide erscheinen durch ihre vorderen Molaren mit drei quergestellten Leisten gegenüber zwei bei den zuvor genannten Vertretern etwas moderner, außerdem sind die Stoßzähne länger. Es wird teilweise angenommen, das ersteres der Entwicklungslinie der Mammutiden näher steht, letzteres den Gomphotherien und den Elefanten.[81][18][15] Die Seekühe kommen stratigraphisch weit gefasst vor. Protosiren tritt bereits in der Gehannam-Formation mit einigen gut erhaltenen Skeletten auf. Sein sehr früher Abzweig innerhalb der Seekühe ist unter anderem noch an den deutlich ausgebildeten Vorder- und Hinterextremitäten und den kräftigen Dornfortsätzen der Brustwirbel erkennbar, die wohl ein semi-aquatisches Leben erlaubten.[82] Die anderen Gattungen, Eosiren und Eotheroides, sind sowohl stammesgeschichtlich als auch stratigraphisch jünger und stehen den heutigen Dugongs näher, sie besaßen ein wesentlich reduziertes Gliedmaßenskelett und stärker geschwollene Knochen. Das bis zu 2,5 m lange Eosiren lebte möglicherweise nicht rein meeresbewohnend, sondern kam auch flussaufwärts vor.[83] Die verschiedenen Seekühe des Fayyum ernährten sich von Seegräsern, wahrscheinlich besetzten sie unterschiedliche ökologische Nischen.[10]



Neben den Afrotheria mit den Seekühen entstanden auch innerhalb der Laurasiatheria mit den Walen meeresbewohnende Säugetiere. Sie sind im Fayyum sehr reichhaltig mit teils vollständigen Skeletten nachgewiesen. Verschiedentlich treten Reste bereits in der Mokattam-Gruppe auf, sehr häufig werden Wale in der Gehannam- und Birket-Qarun-Formation gefunden, in der Qasr-el-Sagha-Formation geht ihr Anteil aber wieder zurück.[11] Allein aus dem Wadi el-Hitan sind gut 500 Skelette bekannt, die die Größe und individuelle Entwicklung der Tiere zu studieren erlauben. Weitere Funde kamen in den Aufschlüssen nördlich und südlich des Qarun-Sees zu Tage. Die bisher aus dem Fayyum-Gebiet beschriebenen Formen gehören einer ursprünglichen Walgruppe an, die als Archaeoceti bezeichnet wird. Diese unterscheiden sich von modernen Walen durch einen vollständigen Gebissschluss, Zähne mit zahlreichen kleinen Höckerchen, weniger spezialisierte Flipper und Hinterbeine, an denen noch die Füße und Zehen ausgebildet waren. Der Nachweis der komplett entwickelten Hinterextremitäten konnte anhand der Fayyum-Fossilien erstmals erbracht werden.[84] In Übereinstimmung mit den heutigen Walen war die Lendenwirbelsäule bereits extrem verlängert. Die ältesten Funde entstammen der Midawara-Formation im Wadi el-Rayyan. Dies zeigt sich unter anderem an den Hintergliedmaßen von Rayanistes, einem Mitglied der Remingtonocetidae. Diese sehr urtümlichen Wale sind bisher weitgehend vom Indischen Subkontinent belegt.[85] Die Gesteinseinheit barg zusätzlich ein Teilskelett von Phiomicetus, das wiederum den Protocetidae zuzuweisen ist, einer ebenfalls sehr ursprünglichen Verwandtschaftsgruppe der Wale.[86] Zu ihr zählt gleichfalls Aegicetus, der mit bisher zwei Individuen aus der Gehannam-Formation des Wadi el-Hitan vorliegt. Die nur knapp 900 kg schweren Tiere hatten noch relativ gleichgroße Hände und Füße. Demzufolge bewegten sie sich weniger undulierend im Wasser fort, sondern nutzten noch stärker ihre Gliedmaße für den Vortrieb. Hinzu kommen die schwächer abgeplatteten Schwanzwirbel, so dass möglicherweise noch keine Fluke ausgebildet war. Ebenso besaß Aegicetus ein noch voll entwickeltes Brustbein, abweichend von späteren Walen.[87] Die anderen Wale aus dem Fayyum-Gebiet gehören zu den Basilosauridae, die bekanntesten und häufigsten Vertreter hier sind Basilosaurus, ein 18 m langer Riese, und das bis zu 5 m lange Dorudon. Anhand von Magenresten von Basilosaurus aus dem Wadi el-Hitan konnte aufgezeigt werden, dass der riesige bezahnte Wal Jagd auf seinen kleineren Verwandten Dorudon machte und wohl bevorzugt dessen Nachwuchs nachstellte. Als weiteres Beutetier ließ sich der ausgestorbene Knochenfisch Pycnodus nachweisen. Damit nahm Basilosaurus wohl die Position eines Spitzenprädators in den Meeren des ausgehenden Eozäns ein.[30] Dorudon hingegen ernährte sich wohl überwiegend von Fischen und nutzte die damals seichten Meeresgewässer zur Geburt des Nachwuchses, wie einzelne Funde von Jungtieren annehmen lassen. Es besaß ein Gewicht von rund 2,2 t, ein kleines, nur 980 g schweres Gehirn und war gemäß dem Bau des Schädels noch nicht zur Echoortung befähigt.[88] Etwa die gleichen Ausmaße wie Dorudon erreichte Ancalecetus, es zeigte aber ein abweichend konfiguriertes Vorderbein.[89] Saghacetus ist mit 3 m Körperlänge recht klein und kommt vorwiegend in der Qasr-el-Sagha-Formation vor.[3][90] Eher selten treten Stromerius und Masracetus in Erscheinung, von letzterem fanden sich unter anderem einzelne Zähne und Teile des Körperskeletts in der Gehannam-Formation nördlich des Qarun-Sees.[91][92] Die nächsten Verwandten der Wale finden sich in den Paarhufern, von diesen sind im Fayyum lediglich Vertreter der Anthracotheriidae belegt, die in der Entwicklungslinie zu den Flusspferden stehen. Die äußerlich schweineähnlichen Tiere waren aufgrund ihres Körperbaus wohl ebenfalls an ein semi-aquatisches Leben angepasst. Während von Qatraniodon und Nabotherium jeweils nur wenige Funde vorliegen, setzt sich das Fossilmaterial von Bothriogenys aus mehr als 2000 Zahn- und Knochenlelementen von Tieren aller Altersstufen zusammen.[93][94][95]
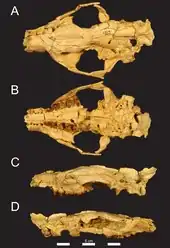
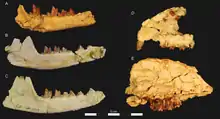
Die Hyaenodonta ersetzten im Paläogen die Raubtiere in Afrika und waren dort die ausschließlichen terrestrischen Beutegreifer. Sie werden teilweise in die umfassendere, aber in sich nicht geschlossene Gruppe der „Creodonta“ gestellt. Im Allgemeinen sind die afro-arabischen Hyaenodonta wenig untersucht, frühere Arbeiten verglichen sie zumeist mit Formen wie Pterodon oder Apterodon, die schon von anderen Kontinenten bekannt waren.[96][18] Das Fundmaterial des Fayyum besteht weitgehend aus Schädeln und Unterkiefern, postcraniale Körperteile sind wenig vorhanden. Unter anderem wurde Masrasector aus dem Fayyum beschrieben, der die Nische der kleinen, bodenlebenden Räuber einnahm, sein Körpergewicht betrug rund 1 kg.[67][97] Demgegenüber wurde Brychotherium sechsmal so schwer, beide übertraf Akhnatenavus, der etwa 20 kg und mehr wog. Letzteren charakterisierte eine extrem entwickelte Brechschere.[98] Ein weiterer nachgewiesener Vertreter ist Metapterodon.[99] Zwei eingekerbte Endphalangen des Vorder- und Hinterfußes verweisen auf Schuppentiere, eine genauere Bestimmung fand noch nicht statt. Sie liegen mit 2,7 beziehungsweise 1,1 cm Länge im Größenbereich der heutigen Arten.[100] Fledertiere bilden ein eher rares Element unter den Fayyum-Fossilien. Ihr Beleg basiert auf etwas mehr als drei Dutzend Einzelfunden, die vorwiegend Gebissreste und isolierte Zähne einschließen. Die Gattungen Philisis, Witwatia und möglicherweise auch Vampyravus gehören einem ausgestorbenen Zweig der Ordnung an. Vampyravus ist dabei im Gegensatz zu den anderen Fledertieren nur über einen 4,9 cm langen Oberarmknochen bekannt. Dieser bereits Anfang des 20. Jahrhunderts vorgestellte Knochen[73] war der erste Hinweis auf ausgestorbene Fledertiere im Fayyum (und in Afrika). Darüber hinaus übertrifft Vampyravus mit einem Körpergewicht um 120 g die meisten anderen fossil überlieferten Fledertiere. Lediglich Aegyptonycteris besaß eine ähnliche Größe, hierbei handelt es sich um ein großes, räuberisch lebendes Tier.[101] Andere Fledertiere können mit heute bestehenden Linien in Verbindung gebracht werden. So steht Dhofarella den Glattnasen-Freischwänzen nahe, Qarunycteris gehört wiederum zu den Mausschwanzfledermäusen und Saharaderma zu den Großblattnasen. Letztere sowie zusätzlich noch Khonsunycteris, eine Glattnase, können mit etwa 30 g Körpergewicht als die kleinsten Fledermäuse aus dem Fayyum angesehen werden. Bemerkenswert ist auch Phasmatonycteris, da die nächsten Verwandten der Gattung heute mit den Madagassischen Haftscheibenfledermäusen nur auf Madagaskar zu finden sind.[102][103][104]
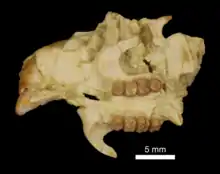
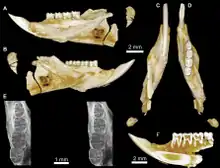
Die Überordnung der Euarchontoglires ist ungemein vielgestaltig vertreten und nimmt allein in der Gebel-Qatrani-Formation mehr als 50 % der Funde ein. Der größere Anteil fällt davon auf die Nagetiere.[75] Hier wiederum dominieren die Stachelschweinverwandten, deren Kennzeichen der hystricognathe Unterkiefer ist. Die Nagetiergruppe vereint ein umfangreiches Schädel- und Gebissmaterial. Phiomys wurde bereits Anfang des 20. Jahrhunderts an einem Unterkiefer definiert[66] und ist vielfach belegt.[22] Näher verwandt sind unter anderem Protophiomys, Acritophiomys und Talahphiomys, aber auch Qatranimys, Waslamys oder Gaudeamus. Innerhalb der Stachelschweinverwandten bilden alle genannten Formen zumeist ausgestorbene Linien.[105][106][107][108][109] Allerdings könnten Birkamys und Mubhammys in die Entwicklungslinie der Rohrratten gehören, ebenso wie Monamys.[110][111] Eine weitere besondere Gruppe findet sich in den Dornschwanzhörnchenverwandten, die bisher nur in den unteren Abschnitten der Qasr-el-Sagha-Formation auftreten. Sie können nur wenig Schädel- und Zahnmaterial vorweisen, nachgewiesene Gattungen sind Shazurus und Kabirmys. Dabei ist ersterer relativ klein, ähnelt im Zahnbau aber schon den Echten Dornschwanzhörnchen. Letzterer stellt die größte bekannte Form der Dornschwanzhörnchenverwandten des Paläogens dar.[112][113]
-2.JPG.webp)
Eine hohe Diversität erreichen auch die Primaten, die neben den Afrotherien eine der angestammten Säugetiergruppen des afrikanischen Kontinents bilden. Loriartige Formen treten unter anderem mit Karanisia und Saharagalago auf,[114] ebenso mit dem zwergenhaften Wadilemur, von dem neben Gebissresten auch Teile des Körperskeletts bekannt sind.[115][116] Die beiden letztgenannten könnten zur Stammgruppe der Galagos gehören, erster hingegen zu den Loris. Plesiopithecus, definiert anhand eines Unterkiefers und später ergänzt durch zusätzliche Funde wie einen Teilschädel,[117][118] reiht sich wiederum in die kontinentalen Vorläuferformen des Fingertiers von Madagaskar ein.[119][120] Eine weitere eher ursprüngliche und ausgestorbene Linie der Primaten wurde mit den Adapiformes etabliert. Ihre Stellung innerhalb der Feuchtnasen- oder innerhalb der Trockennasenprimaten ist teils umstritten. Aus dem Fayyum wurde Afradapis beschrieben, dessen Mahlzähnen mit hohen Höckern und langen Scherkanten eine blattfressende Ernährung anzeigen.[121] Masradapis, der rund 900 g wog, dürfte wiederum aufgrund seiner größeren hinteren Molaren und seines kräftigeren Unterkiefers einen höheren Anteil an Samen und Früchten verzehrt haben. Etwa gleich groß war der verwandte Aframonius, von dem nicht nur Teile des Gebisses, sondern auch des Schädels überliefert sind. Die geringe Größe seiner Augenhöhle spricht für einen tagaktiven Primaten.[122] Andere Formen repräsentieren die Affen und somit die höheren Primaten. Der älteste Vertreter ist Biretia, er kommt bereits im unteren Abschnitt der Qasr-el-Sagha-Formation vor. Anhand der bisher wenigen Zähne und Schädelreste kann ein Tier von etwas mehr als 200 g Gewicht rekonstruiert werden, das abweichend von den meisten heutigen Affen eine nachtaktive Lebensweise pflegte.[123] Im Gegensatz dazu sind alle anderen Affen bisher nur in der Gebel-Qatrani-Formation aufgefunden worden. Es lassen sich verschiedene Entwicklungslinien differenzieren. So gehören etwa die Gattungen Apidium, Parapithecus, Proteopithecus oder Qatrania zu ursprünglichen Seitenlinien der Affen, wobei vor allem Apidium eine sehr große Häufigkeit erreicht.[124][20][125] Die Tiere ernährten sich vorwiegend von Früchten und bewegten sich ihrem Körperskelett nach offensichtlich springend durch die Bäume, auch wenn einige Untersuchungen ihre Agilität als weniger groß einschätzen.[126][127] Andere Formen reihen sich wahrscheinlich in die Stammgruppe der Altweltaffen ein, so unter anderem Catopithecus und Oligopithecus sowie Propliopithecus und Aegyptopithecus.[20][128] Erstere beiden werden in die Verwandtschaftsgemeinschaft der Oligopithecidae eingegliedert, letztere beiden in die der Propliopithecidae. Dabei kommen die Oligopithecidae vor allem in stratigraphisch älteren, die Propliopithecidae in jüngeren Abschnitten der Gebel-Qatrani-Formation vor und schließen einander so weitgehend aus. Ein einzelner Unterkiefer eines wohl verzwergten, aber noch nicht genau bestimmten Oligopitheciden wurde jedoch in einer deutlich höheren Lage (Fundstelle M) aufgefunden und stellt möglicherweise einen der letzten Nachweise dieser Primatengruppe dar. Der Fund trat hier vergesellschaft mit verschiedenen Vertretern der Propliopitheciden auf.[129] Besonders hervorzuheben ist allerdings Aegyptopithecus, der einige gute Schädelfunde vorweisen kann. Dadurch ließ sich ein beträchtlicher Sexualdimorphismus herausarbeiten, der zu den frühesten Belegen bei höheren Primaten gehört. Das Gehirnvolumen der Affenform betrug etwa 20 cm³.[130][131] Zwei weitere Primaten, Nosmips und Afrotarsius sind in ihrer phylogenetischen Stellung unklar. Von beiden liegen nur einige wenige Zähne und Gebissreste vor, die Mischmerkmale von ursprünglicheren und höheren Primaten aufweisen.[132][133][134]
Spurenfossilien

Die Fayyum-Region zeichnet sich durch ausgezeichnet erhaltene Spurenfossilien aus, die in allen stratigraphischen Einheiten auftreten können, in größerer Reichhaltigkeit jedoch in den Bodenbildungen der kontinentalen Gebel-Qatrani-Formation überliefert sind. Es sind mehr als zwei Dutzend verschiedene Formen bekannt. Unter ihnen dominieren vor allem die Lebensspuren von Wirbellosen. Es handelt sich meist um Tunnel und Röhren oder kugelförmige Bildungen, die als Fraß- beziehungsweise Grabgänge von Insekten, Würmern, Weichtieren oder Krebsen gedeutet werden. Unter anderem treten aber auch komplexe Gebilde von bis zu 65 cm Durchmesser auf, die häufig aus verschiedensten Gängen, Kammern und Galerien bestehen. Sie stellen fossilisierte Nester von Termiten dar, die der Spurengattung Termitichnus zugewiesen werden. Andere wiederum beinhalten Gänge in mehreren Ebenen, die von Masrichnus stammen und eventuell auf Bienen zurückgehen. Daneben treten auch deutlich größere Strukturen auf, die teils spiralförmige oder U-förmige Gänge von bis zu 1,5 m Länge und 20 cm Durchmesser darstellen. Ihre Entstehung dürfte auf Wirbeltiere zurückgehen, wobei für die mitunter komplexen Tunnelsysteme grabende Säugetiere als Verursacher in Frage kommen. Eine dritte große Gruppe von Spurenfossilien können als Rhizolithe (Wurzelgänge von Pflanzen) angesprochen werden. Sie treten in allen Größen auf und spiegeln die damals vielfältige Vegetation wider. So sind unter anderem Spuren der Brettwurzeln von Bäumen dokumentiert, die wohl auf Mangroven schließen lassen. Einige dieser Wurzelstrukturen haben einen Durchmesser von 3,2 m, so dass entsprechend große Bäume angenommen werden können.[135][13][136][12]
Altersstellung

Untersuchungen zur Altersstellung der Aufschlüsse im Fayyum und deren Fossilien basieren auf drei unterschiedlichen Ansätzen. Bereits im ausgehenden 19. Jahrhundert wurde für die Fossilien des Fayyums aus biostratigraphischen Erwägungen, etwa in der Zusammensetzung der Haifauna, eine Einstufung in das Obere Eozän und das Untere Oligozän vermutet.[137] Ähnliche Ergebnisse lieferte auch die Molluskengemeinschaft,[138][139] so dass in der Wende vom 19. zum 20. Jahrhundert an einer „frühtertiären“ (Paläogen) Alterseinstufung kaum zu zweifeln war. Die Biostratigraphie mit ihrer relativchronologischen Betrachtung wurde in der Folgezeit noch verfeinert und eine Faunensukzession von den unteren zu den oberen Abschnitten herausgearbeitet. Vor allem für die marinen Ablagerungen erwiesen sich Wirbellose wie etwa Mollusken und Foraminiferen als gute Leitfossilien. Für letztere konnte für die unteren Abschnitte der Mokattam-Gruppe eine Alterseinstufung in das Mittlere Eozän, genauer in das Lutetium, vorgenommen werden. Die oberen Ablagerungssequenzen der Gruppe beginnend mit der Gharaq-Formation gehören dem Bartonium vor 41 bis 38 Millionen Jahren an. Der Übergang zum Priabonium und damit zum Oberen Eozän vor 38 bis 34 Millionen Jahren findet sich in der oberen Gehannam-Formation und wird durch den Wechsel von Truncorotaloides zu Globigerinatheka und das Auftreten von Turborotalia angezeigt. Es gibt hier allerdings eine gewisse Streubreite, so dass keine exakte stratigraphische Position angegeben werden kann.[27][5][140] Eine Alterseinschätzung für die obere terrestrische Abfolge wurde mehrfach mit Hilfe der Säugetiere vorgenommen. Hier ergeben sich aber gewisse Schwierigkeiten, da die Säugetiere in weiten Teilen stark endemisch erscheinen. Sie setzen sich aus originär afrikanischen Formen (Afrotheria) und primär eurasischen Vertretern (Laurasiatheria und Euarchontoglires) zusammen. Als problematisch erwies sich vor allem die Stellung der Gebel-Qatrani-Formation, die je nach Untersuchung entweder vollständig dem Oberen Eozän[141] oder dem Unteren Oligozän[142] angehörte beziehungsweise in den Übergang der beiden Zeitstufen datierte.[17] Anhand einzelner Fossilgruppen konnte dennoch eine relative Altersstellung abgeleitet werden. So wirken die Primaten des Fayyum vielfach ursprünglicher als die stammesgeschichtlich jüngeren Formen des Unteren Miozäns im östlichen Afrika.[143] Wegen der Besonderheit der terrestrischen Säugetierfauna in der Fayyum-Region wurde im Jahr 1991 von John A. Van Couvering und Judith A. Harris das „Phiomian“ als Bezeichnung für eine regionalstratigraphische Stufe vorgeschlagen.[141]
Die im Verlauf des 20. Jahrhunderts entwickelten neuen Datierungsmethoden erlaubten unter anderem absolute Datierungsansätze. Bereits in den 1960er Jahren wurden erste Daten für den auf der eozänen und oligozänen Sedimentabfolge auflagernden Widan-el-Faras-Basalt veröffentlicht, die auf der Kalium-Argon-Methode basierten. Sie ergaben damals ein Alter von 24,7 bis 27 Millionen Jahren, was dem Oberen Oligozän entspricht. Die Werte konnten somit als Mindestalter für die liegenden Sedimentschichten angesehen werden.[22] Weitere radiometrische Analysen aus den 1980er Jahren wiesen dem untersten Abschnitt des Widan-el-Faras-Basalt mit 31 Millionen Jahren ein etwas höheres Alter zu.[143] Das Ergebnis erwies sich im Folgenden aber als ungenau. In den 1990er Jahren durchgeführte Messungen der gleichen Bereiche verbunden mit der Überprüfung der zuvor gewonnenen Altersansätze bestätigten die oberoligozäne Stellung für den Basalt, die neuen Werte betrugen rund 23,6 Millionen Jahre.[144]
Direkte Datierungen der marinen sowie fluviatil-lakustrinen fossilführenden Ablagerungsgesteine des Fayyums sind bisher nicht möglich. Allerdings waren die radiometrischen Untersuchungen der 1990er Jahre mit Messungen zum Paläomagnetismus verbunden. Die analysierten Sequenzen betrafen den oberen Abschnitt der Qasr-el-Sagha-Formation (Dir Abu Lifa Memeber) und die gesamte Gebel-Qatrani-Formation. Innerhalb der Ablagerungsfolge ließ sich ein mehrfacher Wechsel der Polarität des Erdmagnetfeldes nachweisen, den die Autoren mit den magnetostratigraphischen Abschnitten Chron 16 bis Chron 12 in Verbindung brachten. Die Abschnitte entsprachen einer (damaligen) Alterseinstufung von rund 37 bis 33 Millionen Jahren. Als Ergebnis der Arbeit konnte demnach angenommen werden, dass die Fayyum-Ablagerungsserie beinahe vollständig dem Eozän angehörte, nur der oberste Abschnitt der Gebel-Qatrani-Formation fiel in den Übergang zum Unteren Oligozän.[144] Spätere Analysen erfassten eine weitaus umfangreichere Sedimentabfolge beginnend in den basalen Bereichen der Qasr-el-Sagha-Formation und führten auch einen Vergleich mit anderen Fundstellen im näheren und weiteren Umfeld aus. Sie korrigierten und präzisierten die Ergebnisse der ersten Messungen. Aus Sicht der Magnetostratigraphie reicht die Ablagerungssequenz von Chron 17 (Qasr-el-Sagha-Formation) bis Chron 10 (Gebel-Qatrani-Formation), die entsprechenden Altersdaten betragen 38 bis 28,5 Millionen Jahre. Der Übergang vom Eozän zum Oligozän findet sich im Chron 13, welches den unteren Teil der Gebel-Qatrani-Formation einnimmt. Demnach ist der weitaus größere Teil der Gesteinseinheit oligozänen Ursprungs, lediglich die untersten 48 m entstanden bereits im Oberen Eozän. Die bedeutende Fundstelle L-41 befindet sich genau in diesem Übergangsbereich, ihr Alter datiert auf rund 34 Millionen Jahre. Die stratigraphisch jüngeren Fundstellen I und M sind in das Chron 11 eingebettet und somit etwa 29,5 Millionen Jahre alt. Deutlich älter fällt die Einstufung der Fundstelle BQ-2 aus, die einige der frühesten terrestrischen Fossilfunde enthält und sich im unteren Abschnitt der Qasr-el-Sagha-Formation befindet. Sie gehört in das Chron 17 mit einem absoluten Alterswert von etwa 37 Millionen Jahren.[123][145][19]
Landschaftsrekonstruktion
Heute liegt die Fayyum-Region inmitten der Sahara und ist durch Wüstenklima mit einer durchschnittlichen Jahrestemperatur von rund 22 °C bei absoluten Maximalwerten um 49 °C und Minimalwerten um −1 °C charakterisiert. Der jährliche Niederschlag beträgt nur 10 mm und ist weitgehend auf das Winterhalbjahr beschränkt, die Verdunstungsrate kann demgegenüber mehr als das 200fache erreichen.[146]
Im Paläogen stellte Afrika gemeinsam mit der Arabischen Halbinsel eine kontinentale Insel dar. Diese afroarabische Landmasse war durch den Tethys-Ozeans vom eurasischen Kontinent getrennt, eine Landbrücke bildete sich erst im Übergang zum Miozän vor rund 24 Millionen Jahren mit der allmählichen Schließung der Tethys. Die weitgehend aus Kalksteinen bestehenden Formationen der Mokattam-Gruppe sind marinen und flachmarinen Ursprungs und lagerten sich demzufolge im Mittleren Eozän innerhalb der Tethys ab. Gleiches gilt für Teile der Maadi-Gruppe, jedoch macht sich hier bereits der Einfluss der Küstennähe am südlichen Rand der Tethys bemerkbar. Die Birket-Qarun-Formation und die unteren Abschnitte der Qasr-el-Sagha-Formation (Umm Rigl Member bis Temple Member) zeigen den weiteren Rückzug des Meerwassers nach Norden an, wodurch sich im Verlauf des Oberen Eozäns eine bucht-, später lagunenartige Landschaft herausformte. Die auf den flachmarinen Sedimenten geschalteten gröberklastischen Ablagerungen des Dir Abu Lifa Members entstanden dagegen unter den Bedingungen eines Flussdeltas oder eines Ästuars, was zunehmend terrestrische Bedingungen verdeutlicht.[147][8][12][5]

Die Gebel-Qatrani-Formation kappt die unterlagernden Schichten. Sie geht auf ein küstennahes Sumpf- oder Überschwemmungsgebiet zurück, das im Übergang vom Eozän zum Oligozän in Form einer Mangroven- oder Terra-Firma-Landschaft bestand.[136] Das Gebiet wurde von mehreren Flüssen durchzogen, die relativ breit und kräftig genug waren, größere Gesteinskomponenten zu bewegen. An den Flussufern wuchsen Wälder, die auch in das Hinterland hineinreichten. Die relative Meeresnähe bewirkte einen gewissen Einfluss, so dass in einzelnen Bereichen Brackwasserbedingungen vorherrschten. Diese Landschaft trug eine reichhaltige und vielfältige Fauna, die sich aus landlebenden und wasserbewohnenden Tieren zusammensetzte, letztere bestand aus Süß-, Salz- und Brackwasserfomen. Den weiteren paläontologischen und geologischen Daten zufolge herrschte während der Bildung der Gebel-Qatrani-Formation im Unteren Oligozän ein tropisches bis subtropisches Klima vor. Anhand der Ausbildung der Paläoböden lässt sich ein Wechsel von feuchten und trockenen Perioden ablesen, was möglicherweise auf den Einfluss eines Monsuns zurückgeht.[13][2][147]
Vergleich mit regional und überregional bedeutenden Fundstellen
Die Fayyum-Region stellt eine der bedeutendsten spät-paläogenen Fossillagerstätten in Afrika und dem Rest der Welt dar. Sie gehört zudem zu den wenigen Fundstellen auf dem afrikanischen Kontinent, die den Zeitraum vom Oberen Eozän und Unteren Oligozän abdecken. Demgegenüber sind ältere Fundstellen im nördlichen Afrika vergleichsweise häufiger. Eine der frühesten terrestrischen Fossilgemeinschaften des Känozoikums wurde im Ouled-Abdoun-Becken in Marokko entdeckt und datiert in das Paläozän und das Untere Eozän. Ältere fundführende Schichten reichen hier noch bis in das Maastrichtium zurück, so dass ein Fundzeitraum von vor 72 Millionen Jahren bis vor etwa 48 Millionen Jahren erfasst wird.[148][149] Das Becken ist unter anderem durch seinen Phosphatreichtum bekannt. Der obere Teil der Abfolge enthält sehr urtümliche Formen an Säugetieren, etwa Ocepeia und Abdounodus, zwei den heutigen Afrotheria nahestehende Formen. Mit Eritherium und Phosphatherium treten auch zwei der frühesten bekannten Rüsseltiere in Erscheinung, des Weiteren wurde mit Stylolophus ein ursprünglicher Angehöriger der Embrithopoda entdeckt.[150] Als vergleichbar mit dem obersten Abschnitt des Ouled-Abdoun-Becken kann El-Kohol im nördlichen Algerien angesehen werden. Hier wurden unter anderem Beuteltiere, Hyaenodonten, Schliefer und Rüsseltiere aufgefunden. Die Säugetierfauna steht der von Fayyum schon deutlich näher, erscheint mit dem Schliefer Seggeurius und dem Rüsseltier Numidotherium aber immer noch vergleichsweise archaisch.[151] Eine stärkere Vermittlerrolle nimmt der unter- bis mitteleozäne Fundstellenkomplex Gour Lazib, westliches Algerien, ein, da hier mit Megalohyrax und Titanohyrax bereits Formen auftreten, die auch aus Fayyum bekannt sind. Als Besonderheiten sind der sehr frühe Primat Azibius und Helioseus, ein Säugetier mit nicht genau bekannter Verwandtschaft, belegt. Das Material ist zumeist sehr kleinstückig, was die Aussagemöglichkeiten einschränkt.[152][153] Auch Chambi in Tunesien zeigt schon Verbindungen zu Fayyum, neben Schliefern und Primaten wurden hier zusätzlich einige der ältesten Rüsselspringer dokumentiert.[154][155] Von Bedeutung ist auch die weitgehend nur aus Zähnen bestehende Fundkollektion das Ouarzazate-Becken in Marokko. Die zahlreichen Fundstellen des Beckens streuen zeitlich vom ausgehenden Paläozän bis zum Mittleren Eozän. Ältere Lokalitäten wie Adrar Mgorn und N’Tagourt erbrachten urtümliche insektenfresserartige Säugetiere wie Afrodon oder Todralestes, die eventuell einer afrikanisch-endemischen Fauna entsprechen. Hinzu kommen mit Altiatlasius ein Primat, der wohl an der Schwelle zur Entwicklung der Affen steht, und mit Tinerhodon ein Vertreter der Hyaenodonta.[156][157][158] Die Sammlung der jüngsten Fundstelle, Aznag, setzt sich aus spitzmausartige Insektenfresser, Fledertieren, Rüsselspringern und urtümlichen Huftieren zusammen. Es handelt sich bei Aznag um die bisher einzige eindeutig mitteleozäne Fundstelle mit Landsäugetieren des nördlichen Afrika.[159][160]
Aus dem Oberen Eozän kann die Fundstelle Nementcha (auch Bir el-Ater) im nördlichen Algerien genannt werden, die somit zeitgleich zur Qasr el-Sagha-Formation ist. Angezeigt wird das etwa durch Moeritherium und Bunohyrax, zudem konnten die ersten Anthracotherien als eurasische Immigranten in Afrika nachgewiesen werden. Des Weiteren kommen unter den Kleinsäugern Nagetiere der Familie der Phiomyidae und Rüsselspringer vor.[161][162] Ein vergleichbares Alter haben die Funde aus Ad-Dakhla in der Westsahara. Die im Guerrani Member der Samlat-Formation lagernden Fossilien setzen sich neben zahlreichen Fischresten aus Schildkröten, Krokodilen, vereinzelten Vögeln sowie meer- und landbewohnenden Säugetieren zusammen. Unter den Meeressäugern stechen Reste der Wale Saghacetus, Basilosaurus und Dorudon beziehungsweise der Seekuh Eosiren hervor. Als landbewohnendes Säugetier wurde unter anderem der Zahn eines Rüsseltiers entdeckt, der eventuell mit Numidotherium übereinstimmt.[163][164][165]
Die Fundstellen Dor el-Talha und Jebel al-Hasawnah, beide Libyen, bilden dagegen ein zeitliches Äquivalent zur Gebel-Qatrani-Formation im Fayyum. Alle drei Fundgebiete vereint eine vergleichbare Zusammensetzung der Faunengemeinschaft. So sind unter den Großsäugern von Dor el-Talha etwa Barytherium, Moeritherium und Palaeomastodon anzutreffen, genauso wie Arsinoitherium, unter den Reptilien stechen beispielsweise gavialartige Formen hervor. Anzumerken sind auch frühe Primaten wie Karanisia und frühe Affen wie Biretia oder Talahpithecus sowie frühe Rüsselspringer, beispielsweise Eotmantsoius.[166][167][168] In Jebel al-Hasawnah ließen sich unter anderem Saghatherium und Titanohyrax belegen. Die Fundstelle ist auch aus taphonomischen Gründen herausragend, da sie als bisher einzige Fossillagerstätte des paläogenen Afrikas Skelette landlebender Säugetiere im anatomischen Zusammenhang hervorbrachte.[169] In einem ähnlichen zeitlichen Kontext steht der Fundstellenkomplex von Zallah im Sirte-Becken des zentralen Libyens. Hier wurden seit den 1960er Jahren Fossilien geborgen, sie beschränken sich aber weitgehend auf Zähne. Sehr zahlreich ist die Nagetierfauna mit Resten verschiedenster Stachelschweinverwandter wie Gaudeamus, Metaphiomys oder Neophiomys sowie Dornschwanzhörnchenverwandter wie Kabirmys. Daneben sind zudem mit Thyrohyrax auch Schliefer und mit Bothriogenys Vertreter der Anthracotheriidae nachgewiesen.[170][171][172] Africtis wiederum stellt die bisher früheste bekannte afrikanische Form aus dem Verwandtschaftsumfeld der Raubtiere dar, welches seinen Ursprung in Eurasien hat.[173] Die erst im Jahr 2007 entdeckte Fossillagerstätte von Minqar Tibaghbagh im Südwesten der Qattara-Senke entspricht mit ihren gebänderten Sandsteinen zeitlich ebenfalls der Gebel-Qatrani-Formation. Die oligozänen Ablagerungen enthalten nicht nur Reste von Haien, Schildkröten und Krokodilen, sondern auch von landlebenden Säugetieren, beschrieben wurden bisher Phiomia, Antilohyrax und Bothriogenys. Ein älterer Fundhorizont aus dem Oberen Eozän barg zudem Fossilien von Walen und Seekühen.[174][175] Von der Arabischen Halbinsel können noch die Fundstellen Thaytiniti und Taqah im Oman genannt werden, die eine etwa zeitgleiche Fauna wie die oberen Abschnitte von Fayyum beinhalten.[145][160]
Aus dem Oberen Oligozän sind vergleichsweise wenige Fundstellen in Afrika belegt. Eine der bedeutendsten ist Chilga in Äthiopien, deren Alter etwa 27 Millionen Jahre beträgt. Die Großsäuger umfassen sowohl Rüsseltiere als auch Schliefer und Embrithopoda. Sie zeigen eine Mischung aus sowohl älteren Formen wie sie auch im Fayyum vorkommen, etwa Arsinoitherium, Megalohyrax oder Palaeomastodon, aber auch moderneren Elementen. Beispielsweise treten bei den Rüsseltieren Chilgatherium und Gomphotherium hinzu, die beide zu stammesgeschichtlich jüngeren Linien wie die Deinotherien und die Gomphotherien gehören.[176][177] Entlang des Großen Afrikanischen Grabenbruchs finden sich mehrere oberoligozäne, fossilführende Bereiche, die sich vom nordwestlichen Kenia bis in das südwestliche Tansania erstrecken. Erwähnt werden sollen hier beispielhaft nur die Eragaleit Beds westlich des Turkanasee. Auch hier tritt eine Faunenmischung aus älteren und jüngeren Elementen auf. Neben einigen schon seit dem Fayyum bekannten Formen (Arsinoitherium, Thyrohyrax) sind hier Losodokodon als ein basaler Angehöriger der Mammutiden-Linie und Kamoyapithecus, ein Nahverwandter von Proconsul und damit der miozänen Menschenartigen nachgewiesen, auch kommt mit Mioprionodon erstmals ein eindeutiger Angehöriger der aus Eurasien eingewanderten Raubtiere vor.[178][160]
Forschungsgeschichte
Die Anfänge bis ins 19. Jahrhundert
Es ist anzunehmen, dass die Fayyum-Senke als fossilführender Fundplatz bereits in altägyptischer Zeit bekannt war. Aus den oberen Bereichen der Qasr-el-Sagha-Formation sind Steinbrüche belegt, in denen bereits im Alten Reich der zahlreich vorkommende Gips abgebaut und zu Gefäßen verarbeitet wurde. Die Steinbrüche befinden sich in unmittelbarer Nähe zu fossilisierten Baumstammlagen.[179] Der Widan-el-Faras-Basalt diente in gleicher Zeit als Rohstofflager für die Auskleidung von Totentempeln.[180] Die Region galt den Menschen jener Zeit zudem als Ursprung des Lebens, hier soll auch die Göttin Isis die Knochen ihres verstorbenen Mannes Osiris vergraben haben.[15]
Die ersten Fossilfunde der modernen Zeit gehen auf Arthur Bedford Orlebar (1810–1866) zurück, der im Jahr 1845 einige versinterte Baumstümpfe von bis zu 20 m Länge in Sandsteinlagen fand.[181] Weitere Funde sind dem deutschen Geologen Georg Schweinfurth (1836–1925) zu verdanken, der ein erfahrener Afrikaforscher war. Er entdeckte 1879 auf der Insel Geziret el-Qorn im Qarun-See neben zahlreichen Mollusken auch Zähne und Knochen von Haien sowie Knochen von Walen. Allgemein werden die Funde heute der Birket-Qarun-Formation zugewiesen. Die Wirbeltierreste wurden 1883 von Wilhelm B. Dames aufgearbeitet, der unter anderem zwei Arten von Zeuglodon (heute zumeist Basilosaurus) erkannte.[137] Im gleichen Jahr nahm sich der Schweizer Geologe Karl Mayer-Eymar der Weichtiere an.[182] Schweinfurth kehrte Mitte der 1880er Jahre nach Fayyum zurück und untersuchte die Gebiete nördlich des Qarun-Sees. Er entdeckte dabei nicht nur die Tempelruinen von Qasr el-Sagha (auch „Schweinfurths Tempel“ genannt), sondern weitere Wirbeltierreste. Auch diese wurden wiederum von Dames wissenschaftlich ausgewertet, unter anderem beschrieb er die neue Walart Zeuglodon osiris (Saghacetus).[183][184] Über seine Reise veröffentlichte Schweinfurth einen eigenen Bericht, der 1886 erschien.[185] In den 1890er Jahren hielt sich dann auch Mayer-Eymar mehrfach für kurze Zeit im Fayyum auf.[3][15]
Vom 19. zum 20. Jahrhundert – eine große Forschungsperiode
Der Übergang vom 19. zum 20. Jahrhundert fällt in eine intensive Forschungsphase im Fayyum, bei der unterschiedlichste Gruppen tätig wurden. Den Auftakt bildete der britische Paläontologe Hugh John Llewellyn Beadnell (1874–1944). Er kartographierte ab 1898 im Auftrag des Geologischen Dienstes von Ägypten die nördliche und östliche Grenze des Beckens. Ziel war es, ein Bewässerungssystem für die landwirtschaftliche Nutzung des Beckens zu schaffen. Beadnell entdeckte bei seinen Arbeiten zahlreiche Fossilien von Fischen und Krokodilen, aber auch von Walen und Seekühen. Diese sandte er an das Natural History Museum in London, wo sie das Interesse von Charles William Andrews (1866–1924) weckten.[186] Andrews schloss sich ab 1901 den Untersuchungen Beadnells vor Ort an. Zusammen erkundeten sie den nördlichen Teil des Fayyum-Beckens, was bis 1904 andauerte. Ihre Untersuchungen betrafen sowohl die stratigraphisch älteren sowie tiefer gelegenen (Birket-Qarun- und Qasr-el-Sagha-Formation), als auch die jüngeren und höheren Aufschlüsse (Gebel-Qatrani-Formation, hauptsächlich die untere Zone) nördlich des Qarun-Sees. Zu ihren bedeutendsten Entdeckungen aus ersteren gehören die Riesenschlange Gigantophis, des Weiteren auch Barytherium und Moeritherium, zwei Rüsseltiere, sowie die Seekuh Eosiren. In letzteren kamen wiederum Arsinoitherium und Phiomia zum Vorschein, ebenso wie ursprüngliche Schliefer, etwa Saghatherium. Im Jahr 1902 entdeckte Beadnell auch das Wadi el-Hitan. Sowohl Andrews als auch Beadnell publizierten die ersten Ergebnisse ihrer Arbeiten in kurzen Aufsätzen, in der Regel verbunden mit wissenschaftlichen Erstbeschreibungen neuer Arten und Gattungen. Diese erschienen häufig im Geological magazine,[52][187][188][189][54] aber auch anderweitig.[190][191][44] Darüber hinaus gab Beadnell einen kurzen Überblick über seine geologischen Arbeiten,[192] dem 1905 eine ausführliche Monographie folgte. Hier benannte er erstmals das Wadi el-Hitan mit Zeuglodon valley.[1] Ein Jahr später wiederum veröffentlichte Andrews einen umfassenden Katalog zu den Wirbeltierfunden unter dem Titel A descriptive catalogue of the Tertiary Vertebrata of the Fayum, Egypt, der noch heute zu den Standardwerken zu den Fayyum-Fossilien gehört.[81] Seine letzte Reise in das Fayyum-Becken tätigte er im Frühjahr des gleichen Jahres.[193][3][15][90]

Im gleichen Zeitraum fanden auch Forschungstätigkeiten deutscher Wissenschaftler statt. Bereits im Jahr 1900 hatte Max Blanckenhorn (1861–1947) nach einem kurzen Aufenthalt im Fayyum seine geologischen Aufnahmen vorgestellt.[138] Er besuchte zwei Jahre darauf zusammen mit Ernst Stromer (1871–1952) die Fossilfundstellen des Beckens. Das von ihnen gesammelte Material wurde relativ zeitig veröffentlicht.[194][195][196] Ihnen folgte der österreichische Fossiliensammler Richard Markgraf (1869–1916), der seine Heimat aus gesundheitlichen Gründen Richtung Ägypten verlassen hatte. Markgraf traf dort 1897 den deutschen Forscher Eberhard Fraas (1862–1915). Fraas selbst plante weitere Aufenthalte im östlichen und südlichen Afrika, konnte dies aber vorerst nicht verwirklichen. Stattdessen kehrte er nach Stuttgart zurück und unterrichtete Markgraf fernschriftlich in der Fossiliensuche. Im Gegenzug dazu sandte ihm Markgraf seine Fossilien zu, die er zuerst am Gebel Mokattam östlich von Kairo sammelte. Im Jahr 1903 traf Markgraf auf Stromer und schloss sich dessen dreimonatiger Expedition ins Fayyum-Becken an. Beide erforschten die nördlichen Teile der Depression, welche auch im Fokus der britischen Wissenschaftler standen. Danach wurde Markgraf wieder für Fraas tätig. Dieser wiederum organisierte im Jahr 1906 eine Reise nach Ägypten, um sich dort mit Markgraf zusammenzutun. Die Expedition fand schließlich nach einer Reihe von Schwierigkeiten ab dem 11. März für 10 Tage statt. Dabei standen die Qasr-el-Sagha-Formation und die Gebel-Qatrani-Formation im Zentrum ihrer Aufmerksamkeit. Sie entdeckten unter anderem Funde von Arsinoitherium und Basilosaurus, aber auch von Krokodilen.[197] Eine umfangreiche Publikation über die Tätigkeiten der deutschen Forscher im Fayyum-Becken gab Max Schlosser im Jahr 1911 heraus. Darin beschrieb er mit Propliopithecus und Parapithecus auch die ersten eindeutigen Primaten aus dem Fundgebiet.[73] Stromer kehrte 1914 auf seiner dritten Ägyptenexpedition noch einmal kurz ins Fayyum zurück, das Hauptaugenmerk der Reise lag aber auf der Oase Bahariyya. Zu Stromers Forschungen erschienen in den folgenden zwei Dekaden zahlreiche Fachaufsätze in den Abhandlungen der Bayerischen Akademie der Wissenschaften. Markgraf blieb noch bis zu seinem Tod im Jahr 1916 in Ägypten und sammelte regelmäßig im Fayyum-Becken, seine Funde bot er verschiedenen wissenschaftlichen Instituten an.[3][15]

Parallel zu Markgrafs Tätigkeiten vor Ort und nach dem Erscheinen der Publikationen von Beadnell und Andrews organisierte das American Museum of Natural History unter Federführung von Henry Fairfield Osborn (1857–1935) eine Expedition in das Fayyum-Becken. Es handelt sich hierbei um die erste Auslandsexpedition des Museums, der noch einige weitere, sehr erfolgreiche unter anderem in die Mongolei folgen sollten. Unterstützt wurde die Expedition vom damaligen US-Präsidenten Theodore Roosevelt. Die Amerikaner trafen dort Anfang Februar 1907 ein und blieben bis Ende Mai des gleichen Jahres. Als Assistenten Osborns fungierten Walter W. Granger (1872–1942) und George Olsen. Osborn selbst blieb nur für den Anfang der Expedition in Ägypten, er unternahm aber einen kurzen Abstecher ins Wadi el-Hitan, das er gemäß Beadnell Zeuglodon valley nannte. Die verbliebenen Expeditionsteilnehmer arbeiteten in verschiedenen Aufschlüssen und trafen während der Zeit mehrfach Markgraf. Dieser wurde von den Amerikanern auch zur Fossiliensuche eingestellt, arbeitete aber unabhängig. Ein Großteil der Forschungen betraf die Gebel-Qatrani-Formation und setzte die Arbeiten in den bereits von Andrews und Beadnell erschlossenen Fundstellen in der unteren Zone fort. Die Forscher entdeckten aber zusätzlich auch erstmals Funde in der oberen Zone.[66] Zu den aufgefundenen Fossilien gehören unter anderem Reste von Arsinoitherium, mehreren Rüsseltierformen, Hyaenodonten, Nagetieren und frühen Schliefern. Das gesamte Fundmaterial, etwa 550 Einzelobjekte, wurde nach Amerika verschifft. Im Nachklang der Expedition brachte Osborn mehrere Artikel über die Fossilfunde heraus,[66][96] ebenso wie über die Expedition selbst.[198][199] Der vollständige Bericht, der das von Granger geführte Tagebuch beinhaltet, wurde erst 2002 veröffentlicht.[22][15]
Unabhängig von den vorhergehenden Untersuchungen führte das Muséum national d’histoire naturelle von Paris unter organisatorischer Leitung von Marcellin Boule und Jean Albert Gaudry im März/April 1904 eine zweiwöchige Expedition ins Fayyum durch, die finanziell von Edmond de Rothschild unterstützt wurde. Beteiligt war unter anderem auch René Fourtau, ein französischer Ingenieur, der seit 1888 in Ägypten lebte und sich für Geologie interessierte. Fourtau analysierte unter anderem für den Geologischen Dienst Ägyptens fossile Wirbellose, vorwiegend Seeigel. Die Arbeiten fanden in der Gebel-Qatrani-Formation statt. Als Ergebnis brachten die Franzosen rund 80 Wirbeltierfossilien nach Paris, darunter auch Reste von Titanohyrax, einem riesigen Schliefer. Im Gegensatz zu den anderen Forschungsunternehmungen wurden die des Muséum national d’histoire naturelle kaum aufgearbeitet, so dass die Expedition des Jahres 1904 fast in Vergessenheit geriet.[200][201]
Intermezzo
In den folgenden, nahezu fünf Dekaden nach den überwiegend britischen, deutschen und amerikanischen Forschungen vor Ort fanden kaum Aktivitäten im Fayyum statt. Eine Ausnahme bildet eine pan-afrikanische Expedition der University of California im Jahr 1947 unter Wendell Phillips. Mehrere Wissenschaftler gruben in bereits zuvor geöffneten Aufschlüssen. Aufgrund der begrenzten Zeit ist das Fundmaterial nicht sehr umfangreich, setzt sich aber aus Rüsseltieren, Hyaenodonten, Anthracotherien und Embrithopoda zusammen. Es befindet sich heute in Berkeley. Drei Jahre später wurde zudem noch ein Schädel eines urtümlichen Wales entdeckt.[22][69][90]
Moderne Forschung
In den 1950er Jahren beschrieb Elwyn L. Simons (1930–2016) den Schädelknochen eines Primaten aus dem Fayyum, der in den Beständen des American Museum of Natural History lagerte.[202] Daraufhin leitete er eine Initiative zu weiteren Feldforschungen im Fayyum ein, die unter anderem der Frage nach dem Ursprung der nächsten Menschenverwandten nachgehen sollten. In Kooperation der Yale University und später der Duke University mit dem Geologischen Dienst von Ägypten und dem Geologischen Museum von Kairo begannen die Arbeiten vor Ort im Jahr 1961 und endeten vorerst 1967. Im Mittelpunkt standen neben den frühen Primaten hauptsächlich Kleinvertebraten, die zuvor weniger stark beachtet worden waren. Die jährlichen Feldforschungen führten unter anderem zur Beschreibung von Oligopithecus und Aegyptopithecus, zwei Primatenformen, beziehungsweise Phiocricetomys, einem Nagetier. Daneben gab es aber auch Untersuchungen zur Stratigraphie und zur Altersstellung. Ein erklärtes Ziel war es beispielsweise, die exakte Position der älteren Aufschlüsse zu dokumentieren. Simons gab 1968 einen Überblick über die geologisch-paläontologischen Arbeiten heraus.[22][69]
Im Jahr 1977 startete eine zweite Phase der Untersuchungen im Fayyum, wieder unter der Gesamtleitung von Simons. Sie dauerte bis 2005 und damit über einen Zeitraum von 28 Jahren. Geprägt war die zweite Phase von einer zunehmend interdisziplinären Arbeitsweise. Beteiligt waren zahlreiche international bedeutende Wissenschaftler, die jeweils ihre eigenen Forschungsschwerpunkte setzten. Dazu gehörten unter anderem Philip D. Gingerich (Meeressäuger), David Tab Rasmussen (Vögel, Schliefer) und Erik R. Seiffert (Primaten). Zusätzlich standen verstärkt andere Arbeitsfelder im Fokus, etwa die Paläoumwelt, die Paläomagnetik oder Bodenkunde. Darüber hinaus wurden auch neue Fossiliengruppen wie etwa die Spurenfossilien mit eingeschlossen. Ein weiterer Fokus betraf die Ausbildung des wissenschaftlichen Nachwuchses. Durch Fortschritte in der Grabungsmethodik konnte die Fundausbeute deutlich erhöht werden. Allein Ende der 1970er Jahre wurden dadurch mehr als 1400 Säugetierreste geborgen.[23][69] Die Arbeiten vor Ort gingen danach weiter, die Aufarbeitung des Fundmaterials hält bis heute an, wodurch beinahe jährlich neue Arten und Gattungen aus dem Fayyum-Becken beschrieben werden.[90]
Fundaufbewahrung
Bis auf den heutigen Tag sind Zehntausende von Fossilfunden im Fayyum gemacht worden, die vorwiegend im Ägyptischen Geologischen Museum in Kairo, im Peabody Museum of Natural History der Yale University, im Duke Lemur Center der Duke University, im American Museum of Natural History in New York, im Natural History Museum in London, im Muséum national d’histoire naturelle in Paris, im Staatlichen Museum für Naturkunde in Stuttgart und in der Bayerischen Staatssammlung für Paläontologie und Geologie in München aufbewahrt werden.
Literatur
- Charles W. Andrews: A descriptive catalogue of the Tertiary Vertebrata of the Fayum, Egypt. London 1906, S. 1–324
- Vincent L. Morgan und Spencer G. Lucas: Notes From Diary–Fayum Trip, 1907 (based on the expedition diary and photographs of Walter Granger). Bulletin of the New Mexico Museum of Natural History and Science 22, 2002, S. 1–148
- Elwyn L. Simons: Eocene and Oligocene mammals of the Fayum, Egypt. In: First International Conference on the Geology of the Tethys, Cairo University, November, 2005, Volume II. Cairo 2005, S. 439–450
Einzelnachweise
- Hugh John Llewellyn Beadnell: The Topography and Geology of the Fayum Province of Egypt. Cairo, 1905, S. 1–101
- Thomas M. Bown und Mary J. Kraus: Geology and Paleoenvironment of the Oligocene Jebel Qatrani Formation and Adjacent Rocks, Fayum Depression, Egypt. U. S. Geological Survey Professional Paper 1452, 1988, S. 1–60
- Philip D. Gingerich: Marine mammals (Cetacea and Sirenia) from the Eocene of Gebel Mokattam and Fayum, Egypt: Stratigraphy, age, and paleoenvironments. Papers on Paleontology 30, 1992, S. 1–84
- Gamal M. El-Shabrawy und Henri J. Dumont: The Fayum Depression and Its Lakes. In: Henri J. Dumont (Hrsg.): The Nile. Origin, Environments, Limnology and Human Use. Monographiae Biologicae, Springer, 2009, S. 95–124
- Chris King, Charlie Underwood und Etienne Steurbaut: Eocene stratigraphy of the Wadi Al-Hitan World Heritage Site and adjacent areas (Fayum, Egypt). Stratigraphy 11 (3–4), 2014, S. 185–234
- Ulrich Jux und Thomas Steubner: Der Karun-See im ägyptischen Fayum. Seine Sedimente im Spiegel der Geschichte. Naturwissenschaften 77, 1990, S. 262–270
- Rushdi Said, Claude C. Albritton, Fred Wendorf, Romuald Schild und Michał Kobusiewicz: Remarks on the Holocene geology and archaeology of Northern Fayum desert. Archaeologia Polona 13, 1972, S. 7–22
- El-Sayed A. A.Youssef, M.A. Abdel Fattah und A. Refaat: Sequence stratigraphy of the Late Eocene/Oligocene alluvial/paralic succession in the Fayum area, Western Desert, Egypt. Bulletin of the Tethys Geological Society 1, 2006, S. 59–70
- Timothy M Kusky, Talaat M Ramadan, Mahmoud M Hassaan und Safwat Gabr: Structural and Tectonic Evolution of El-Faiyum Depression, North Western Desert, Egypt Based on Analysis of Landsat ETM+, and SRTM Data. Journal of Earth Science 22 (1), 2011, S. 75–100
- Iyad S. Zalmout und Philip D. Gingerich: Late Eocene Sea Cows (Mammalia, Sirenia) from Wadi Al Hitan in the Western Desert of Fayum, Egypt. Papers on Paleontology 37, 2012, S. 1–158
- M. Gameil, M. Al Anbaawy, M. Abdel Fattah und G. Abu El-Kheir: Lithofacies and biofacies characteristics and whales skeletons distribution in the Eocene rock units of Fayoum Area, Egypt. Journal of African Earth Sciences 116, 2016, S. 42–55
- Zaki A. Abdel-Fattah, Murray K. Gingras, Michael W. Caldwell und S. George Pemberton: Sedimentary environments and depositional characteristics of the Middle to Upper Eocene whale-bearing succession in the Fayum Depression, Egypt. Sedimentology 57, 2010, S. 446–476
- Thomas M. Bown, Mary J. Kraus, Scott L. Wing, John G. Fleagle, Bruce H. Tiffney, Elwyn L. Simons und Carl F. Vondra: The Fayum Primate Forest Revisited. Journal of Human Evolution 11, 1982, S. 603–632
- Abdelfattah A. Zalat: Holocene diatom assemblages and their palaeoenvironmental interpretations in Fayoum depression, Western Desert, Egypt. Quaternary International 369, 2015, S. 86–98
- Vincent L. Morgan und Spencer G. Lucas: Notes From Diary–Fayum Trip, 1907 (based on the expedition diary and photographs of Walter Granger). Bulletin of the New Mexico Museum of Natural History and Science 22, 2002, S. 1–148 (online)
- Gunther Garbrecht: Historical water storage for irrigation in the Fayum depression (Egypt). Irrigation and Drainage Systems 10, 1996, S. 47–76
- David Tab Rasmussen, Thomas M. Bown und Elwyn L. Simons: The Eocene-Oligocene transition in continental Africa. In: Donald R. Prothero und William A. Berggren (Hrsg.): Eocene-Oligocene Climatic and Biotic Evolution. Princeton University Press, Princeton, 1992, S. 548–566
- Patricia A. Holroyd, Elwyn L. Simons, Thomas M. Bown, Paul D. Polly und Mary J. Kraus: New records of terrestrial mammals from the Upper Eocene Qasr el Sagha Formation, Fayum depression, Egypt. Palaeovertebrata 25 (2–4), 1996, S. 175–192
- Erik R. Seiffert, Thomas M. Bown, William C. Clyde und Elwyn Simons: Geology, Paleoenvironment, and Age of Birket Qarun Locality 2 (BQ-2), Fayum Depression, Egypt. In: J. G. Fleagle und C. C. Gilbert (Hrsg.): Elwyn Simons: A Search for Origins. Springer, 2008, S. 71–86
- Elwyn L. Simons: Description of two genera and species of Late Eocene Anthropoidea from Egypt. PNAS 86, 1989, S. 9956–9960
- David Tab Rasmussen und E. L. Simons: The oldest hyracoids (Mammalia: Pliohyracidae): new species of Saghatherium and Thyrohyrax from the Fayum. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 182, 1991, S. 187–209
- Elwyn L. Simons und Albert E. Wood: Early Cenozoic Mammalian Faunas Fayum Province, Egypt. Bulletin of the Peabody Museum of Natural History 28, 1968, S. 1–105
- Elwyn L. Simons: Egyptian Oligocene Primates: A Review. Yearbook of Physical Anthropology 38, 1995, S. 199–238
- Samar Nour-El-Deen, Romain Thomas und Wagieh El-Saadawi: First record of fossil Trachycarpeae in Africa: three new species of Palmoxylon from the Oligocene (Rupelian) Gebel Qatrani Formation, Fayum, Egypt. Journal of Systematic Palaeontology 16 (9), 2018, S. 741–766, doi:10.1080/14772019.2017.1343258
- Richard Kräusel: Ergebnisse der Forschungsreisen Prof. E. Stromers in den Wüsten Ägyptens. IV. Die fossilen Floren Ägyptens. Abhandlungen der Bayerischen Akademie der Wissenschaften Mathematische-Naturwissenschaften Abteilungen, Neue Folge, 47, 1939, S. 1–140 ()
- Wagieh E. El-Saadawi: On the fossil flora of Jebel Qatrani area, Fayum, Egypt. Taeckholmia 26, 2006, S. 131–140
- Amin Strougo, Mahmoud Faris, Mona A. Y. Haggag, Radwan, A. Abdul-Nasr und Philip D. Gingerich: Planktonic foraminifera and calcareous nannofossil biostratigraphy through the middle to late Eocene transition at Wadi Hitan, Fayum Province, Egypt. Contributions from the Museum of Paleontology 32, 2013, S. 111–138
- Akmal Mohamed Marzouk, Ahmed Moustafa El Shishtawy und Atef Masoud Kasem: Calcareous nannofossil and planktonic foraminifera biostratigraphy through the Middle to Late Eocene transition of Fayum area, Western Desert, Egypt. Journal of African Earth Sciences 100, 2014, S. 303–323
- Jessica L. Anderson und Rodney M. Feldmann: Lobocarcinus lumacopius (Decapoda: Cancridae), a new species of cancrid crab from the Eocene of Fayum, Egypt. Journal of Paleontology 69 (5), 1995, S. 922–932
- Manja Voss, Mohammed Sameh M. Antar, Iyad S. Zalmout and Philip D. Gingerich: Stomach contents of the archaeocete Basilosaurus isis: Apex predator in oceans of the late Eocene. PLoS ONE 14 (1), 2019, S. e0209021, doi:10.1371/journal. pone.0209021
- Charlie J. Underwood und David J. Ward: New Hemigaleid Shark from the Late Eocene of Wadi Al-Hitan, Egypt. Journal of Vertebrate Paleontology 31 (3), 2011, S. 707–711
- Gerard R. Case und Henri Cappetta: The Eocene selachian fauna from the Fayum Depression in Egypt. Palaeontographica Abteilung A 212, 1990, S. 1–30
- Alison M. Murray, Todd D. Cook, Yousry S. Attia, Prithijit Chatrath und Elwyn L. Simons: A Freshwater Ichthyofauna from the Late Eocene Birket Qarun Formation, Fayum, Egypt. Journal of Vertebrate Paleontology 30 (3), 2010, S. 665–680
- Charlie J. Underwood, David J. Ward, Christopher King, Sameh M. Antar, Iyad S. Zalmout und Philip D. Gingerich: Shark and ray faunas in the Middle and Late Eocene of the Fayum Area, Egypt. Proceedings of the Geologists' Association 122, 2011, S. 47–66
- Alison M. Murray, Elwyn L. Simons und Yousry S. Attia: A new clupeid fish (Clupeomorpha) from the Oligocene of Fayum, Egypt, with notes on some other fossil clupeomorphs. Journal of Vertebrate Paleontology 25 (2), 2005, S. 300–308
- Sanaa E. El-Sayed, Mahmoud A. Kora, Hesham M. Sallam, Kerin M. Claeson, Erik R. Seiffert und Mohammed S. Antar: A new genus and species of marine catfishes (Siluriformes; Ariidae) from the upper Eocene Birket Qarun Formation, Wadi El-Hitan, Egypt. PLoS ONE 12 (3), 2017, S. e0172409, doi:10.1371/journal.pone.0172409
- Harry L. Fierstine und Philip D. Gingerich: A second and more complete rostrum of Xiphiorhynchus aegyptiacus Weiler, 1929 (Perciformes: Xiphioidei, Xiphiidae, Xiphiorhynchinae), from the Birket Qarun Formation, late Eocene, Egypt. Journal of Vertebrate Paleontology 29 (2), 2009, S. 589–593
- Alison M. Murray und Yousry S. Attia: A new species of Lates (Teleostei: Perciformes) from the Lower Oligocene of Egypt. Journal of Vertebrate Paleontology 24 (2), 2004, S. 299–308
- Alison M. Murray: A new channid fish (Teleostei: Channiformes) from the Eocene and Oligocene of Egypt. Journal of Paleontology 80 (6), 2006, S. 1172–1178
- Alison M. Murray: Lower pharyngeal jaw of a cichlid fish (Actinopterygii; Labroidei) from an early Oligocene site in the Fayum, Egypt. Journal of Vertebrate Paleontology 22 (2), 2002, S. 453–455
- Alison M. Murray: Late Eocene and Early Oligocene teleost and associated ichthyofauna of the Jebel Qatrani Formation, Fayum, Egypt. Palaeontology 47 (3), 2004, S. 711–724
- Alison M. Murray, Thodoris Argyriou und Todd D. Cook: Palaeobiogeographic relationships and palaeoenvironmental implications of an earliest Oligocene Tethyan ichthyofauna from Egypt. Canadian Journal of Earth Sciences 51 (10), 2014, S. 909–918
- Patricia A. Holroyd und James Ford Parham: The antiquity of African tortoises. Journal of Vertebrate Paleontology 23 (3), 2003, S. 688–690
- Charles W. Andrews: On some Pleurodiran Chelonians from the Eocene of the Fayum, Egypt. Annals and magazine of natural history 7 (11), 1903, S. 115–121 ()
- Eugene S. Gaffney, Donald D. DeBlieux, Elwyn L. Simons, Marcelo R. Sánchez-Villagra und Peter A. Meylan: Redescription of the Skull of Dacquemys Williams, 1954, a Podocnemidid Side-Necked Turtle from the Late Eocene of Egypt. American Museum Novitates 3372, 2002, S. 1–16
- Eugene S. Gaffney, Peter A. Meylan, Roger C. Wood, Elwyn Simons und Diogenes De Almeida Campos: Evolution of the Side-Necked Turtles: The Family Podocnemididae. Bulletin of the American Museum of Natural History 350, 2011, S. 1–237
- Adán Pérez-García: New shell information and new generic attributions for the Egyptian podocnemidid turtles „Podocnemis“ fajumensis (Oligocene) and „Podocnemis“ aegyptiaca (Miocene). Fossil Record 24, 2021, S. 247–262, doi:10.5194/fr-24-247-2021
- Krister T. Smith, Bhart-Anjan S. Bhullar und Patricia A. Holroyd: Earliest African Record of the Varanus Stem-Clade (Squamata: Varanidae) from the Early Oligocene of Egypt. Journal of Vertebrate Paleontology 28 (3), 2008, S. 909–913
- Robert B. Holmes, Alison M. Murray, Yousry S. Attia, Elwyn L. Simons und Prithijit Chatrath: Oldest known Varanus (Squamta: Varanidae) from the Upper Eocene and Lower Oligocene of Egypt: Suport for an African origin of the Genus. Palaeontology 53 (5), 2010, S. 1099–1110
- Jacob A. McCartney und Erik R. Seiffert: A late Eocene snake fauna from the Fayum depression, Egypt. Journal of Vertebrate Paleontology 36 (1), 2016, S. e1029580, doi:10.1080/02724634.2015.1029580
- Jonathan P. Rio und Philip D. Mannion: The osteology of the giant snake Gigantophis garstini from the upper Eocene of North Africa and its bearing on the phylogenetic relationships and biogeography of Madtsoiidae. Journal of Vertebrate Paleontology 37 (4), 2017, S. e1347179, doi:10.1080/02724634.2017.1347179
- Charles W. Andrews: Preliminary Note on some Recently Discovered Extinct Vertebrates from Egypt. Geological Magazine, Part I. 4 (8), 1901, S. 400–409 (), Part II. 4 (8), 1901, S. 436–444 (), Part III. 4 (9), 1902, S. 291–295 ()
- Eric Buffetaut: Systématique, origine et évolution des gavialidae Sud-Américains. Geobios 15 (suppl. 1), 1982, S. 127–140
- Charles W. Andrews: Notes on some new Crocodilia from the Eocene of Egypt. Geological Magazine 5 (2), 1905, S. 481–484 ()
- Christopher Brochu und Philip D. Gingerich: New tomistomine crocodylians from the Middle Eocene (Bartonian) of Wasdi Hitan, Fayum Province, Egypt. Contributions from the Museum of Paleontology, The University of Michigan 30, 2000, S. 251–268
- Candice M. Stefanic, Jennifer H. Nestler, Erik R. Seiffert und Alan H. Turner: New crocodylomorph material from the Fayum Depression, Egypt, including the first Occurrence of a sebecosuchian in African late Eocene deposits. Journal of Vertebrate Paleontology 39 (6), 2019, S. e1729781, doi:10.1080/02724634.2019.1729781
- Joseph J. El Adli, Jeffrey A. Wilson Mantilla, Mohammed Sameh M. Antar und Philip D. Gingerich: The earliest recorded fossil pelican, recovered from the late Eocene of Wadi Al-Hitan, Egypt. Journal of Vertebrate Paleontology, 2021, S. e1903910, doi:10.1080/02724634.2021.1903910
- David Tab Rasmussen, Storrs L. Olson und Elwyn L. Simons: Fossil Birds from the Oligocène Jebel Qatrani Formation, Fayum Province, Egypt. Smithsonian Contribution to Paleobiology 62, 1987, S. 1–20
- David Tab Rasmussen, Elwyn . L. Simons, F. Hertel und A. Judd: Hindlimb of a giant terrestrial bird from the Upper Eocene, Fayum, Egypt. Palaeontology 44 (2), 2001, S. 325–337
- Thomas A. Stidham und N. Adam Smith: An ameghinornithid-like bird (Aves, Cariamae, ?Ameghinornithidae) from the early Oligocene of Egypt. Palaeontologia Electronica 18 (1), 2015, S. 5A ()
- Elwyn L. Simons und Thomas M. Bown: A New Species of Peratherium (Didelphidae; Polyprotodonta): The First African Marsupial. Journal of Mammalogy 65 (4), 1984, S. 539–548
- Thomas M. Bown und Elwyn L. Simons: First record of marsupials (Metatheria: Polyprotodonta) from the Oligocene in Africa. Nature 308, 1984, S. 447–449
- Jerry J. Hooker, Marcelo R. Sánchez-Villagra, Francisco J. Goin, Elwyn L. Simons, Yousry Attia und Erik R. Seiffert: The origin of Afro-Arabian „didelphimorph“ marsupials. Palaeontology 51 (3), 2008, S. 635–648
- Marcelo R. Sánchez-Villagra, Erik R. Seiffert, Thomas Martin, Elwyn L. Simons, Gregg F. Gunnell und Yousry Attia: Enigmatic new mammals from the late Eocene of Egypt. Paläontologische Zeitschrift 81 (4), 2007, S. 406–415
- Vicente D. Crespo und Francisco J. Goin: Taxonomy and affinities of African Cenozoic metatherians. Spanish Journal of Palaeontology 36 (2), 2021, S. 1–16, doi:10.7203/sjp.36.2.20974
- Henry Fairfield Osborn: New fossil mammals from the Fayum Oligocene, Egypt. Bulletin of the American Museum of Natural History 24, 1908, S. 265–272
- Elwyn L. Simons und Philip D. Gingerich: New carnivorous mammals from the Oligocene of Egypt. Annals of the Geological Survey of Egypt 4, 1974, S. 157–166
- Elwyn L. Simons und Thomas M. Bown: Ptolemaiida, a new order of Mammalia – with description of the first cranium of Ptolemaia grangeri. PNAS 92, 1995, S. 3269–3273
- Elwyn L. Simons: Eocene and Oligocene mammals of the Fayum, Egypt. In: First International Conference on the Geology of the Tethys, Cairo University, November, 2005, Volume II. Cairo, 2005, S. 439–450
- Erik R. Seiffert und Elwyn L. Simons: Widanelfarasia, a diminutive placental from the late Eocene of Egypt. PNAS 97 (6), 2000, S. 2646–2651
- Erik R. Seiffert, Elwyn L. Simons, Timothy M. Ryan, Thomas M. Bown und Yousry Attia: New remains of Eocene and Oligocene Afrosoricida (Afrotheria) from Egypt, with implications for the origin(s) of afrosoricid zalambdodonty. Journal of Vertebrate Paleontology 27 (4), 2007, S. 963–972
- Erik R. Seiffert: The oldest and youngest records of afrosoricid placentals from the Fayum Depression of northern Egypt. Acta Palaeontologica Polonica 55 (4), 2010, S. 599–616
- Max Schlosser: Beiträge zur Kenntnis der Oligozänen Landsäugetiere aus dem Fayum, Ägypten. Beiträge zur Paläontologie und Geologie Österreich-Ungarns und des Orients 24, 1911, S. 51–167 ()
- Elwyn L. Simons, Patricia A. Holroyd und Thomas M. Bown: Early tertiary elephant-shrews from Egypt and the origin of the Macroscelidea. PNAS 88, 1991, S. 9734–9737
- Mario Gagnon: Ecological diversity and community ecology in the Fayum sequence (Egypt). Journal of Human Evolution 32, 1997, S. 133–160
- Eugenie Barrow, Erik R. Seiffert und Elwyn L. Simons: A primitive hyracoid (Mammalia, Paenungulata) from the early Priabonian (Late Eocene) of Egypt. Journal of Systematic Palaeontology 8 (2), 2010, S. 213–244
- David Tab Rasmussen und Elwyn L. Simons: New Oligocene Hyracoids from Egypt. Journal of Vertebrate Paleontology 8 (1), 1988, S. 67–83
- Grant E. Meyer: A new Oligocene hyrax from the Jebel Qatrani formation, Fayum, Egypt. Postilla Peabody Museum Yale University 163, 1973, S. 1–11
- David Tab Rasmussen und Elwyn L. Simons: Ecomorphological diversity among Paleogene hyracoids (Mammalia): a new cursorial browser from the Fayum, Egypt. Journal of Vertebrate Paleontology 20 (1), 2000, S. 167–176
- Hikoshichiro Matsumoto: Contribution to the knowledge of the fossil Hyracoidea of the Fayum, Egypt, with description of several new species. Bulletin of the American Museum of Natural History 56, 1926, S. 253–350
- Charles W. Andrews: A descriptive catalogue of the Tertiary Vertebrata of the Fayum, Egypt. London, 1906, S. 1–324
- Daryl P. Domning und Philip D. Gringerich: Protosiren smithae, new species (Mammalia, Sirenia) from the Late Middle Eocene of Wadi Hitan, Egypt. Contributions from the Museum of Paleontology University of Michigan 29 (3), 1994, S. 69–87
- Daryl P. Domning, Philip D. Gringerich, Elwyn L. Simons und Friderun A. Ankel-Simons: A new Early Oligocene dugongid (Mammalia, Sirenia) from Fayum Province, Egypt. Contributions from the Museum of Paleontology University of Michigan 29 (4), 1994, S. 89–108
- Philip D. Gringerich, B. Holly Smith und Elwyn L. Simons: Hind limbs of Eocene Basilosaurus: evidence of feets in whales. Science 249, 1990, S. 154–157
- Ryan M. Bebej, Iyad S. Zalmout, Ahmed A. Abed El-Aziz, Mohammed Sameh M. Antar und Philip D. Gingerich: First remingtonocetid archaeocete (Mammalia, Cetacea) from the middle Eocene of Egypt with implications for biogeography and locomotion in early cetacean evolution. Journal of Paleontology 89 (5), 2015, S. 882–893, doi:10.1017/jpa.2015.57
- Abdullah S. Gohar, Mohammed S. Antar, Robert W. Boessenecker, Dalia A. Sabry, Sanaa El-Sayed, Erik R. Seiffert, Iyad S. Zalmout und Hesham M. Sallam: A new protocetid whale offers clues to biogeography and feeding ecology in early cetacean evolution. Proceedings of the Royal Society B: Biological Sciences 288 (1957), 2021, S. 20211368, doi:10.1098/rspb.2021.1368
- Philip D. Gingerich, Mohammed Sameh M. Antar und Iyad S. Zalmot: Aegicetus gehennae, a new late Eocene protocetid (Cetacea, Archaeoceti) from Wadi Al Hitan, Egypt, and the transition to tail-powered swimming in whales. PLoS ONE 14 (12), 2019, S. e0225391, doi:10.1371/journal.pone.0225391
- A. Mahdy, G. Abu El-Kheir, Abdullah S. Gohar und Maher I. El-Soughier: Palaeobiological assessment of some basilosaurid archaeocetes (mammalia, cetacea) and itsaffinity with recent odontocetes: new insights from Wadi El-Hitan, Fayoum, Egypt. Historical Biology 33 (83), 2021, S. 313–322, doi:10.1080/08912963.2019.1616292
- Philip D. Gringerich und Mark D. Uhlen: Ancalecetus simonsi, a new dorudontine archaeocete (Mammalia, Cetacea) from the Early Late Eocene of Wadi Hitan, Egypt. Contributions from the Museum of Paleontology University of Michigan 29 (13), 1996, S. 359–401
- Philip D. Gingerich: Early Evolution of Whales. A Century of Research in Egypt. In: J. G. Fleagle und C. C. Gilbert (Hrsg.): Elwyn Simons: A Search for Origins. Springer, 2008, S. 107–124
- Gebely A. Abu El-Kheir, Mohammed I. El Anbaawy, Sobhi A. Helal und Susan Gibbs: Dentary of Masracetus markgrafi in the north of Lake Qaroun, Fayoum Egypt. Journal of American Science 9 (12), 2013, S. 459–469
- Philip D. Gingerich: Stromerius nidensis, new archaeocete (Mammalia, Cetacea) from the Upper Eocene Qasr el-Sagha Formation, Fayum, Egypt. Contributions from the Museum of Paleontology University of Michigan 31 (13), 2007, S. 363–378
- Stéphane Ducrocq: The anthracotheriid genus Bothriogenys (Mammalia, Artiodactyla) in Africa and Asia during the Paleogene: phylogenetical and paleobiogeographical relationships. Stuttgarter Beiträge zur Naturkunde, Serie B Geologie und Paläontologie 250, 1997, S. 1–44
- Afifi H. Sileem, Hesham M. Sallam, Abdel Galil A. Hewaidy, Ellen R. Miller und Gregg F. Gunnell: A new anthracothere (Artiodactyla) from the early Oligocene, Fayum, Egypt, and the mystery of African ‘Rhagatherium’ solved. Journal of Paleontology 90 (1), 2016, S. 170–181
- Hesham M. Sallam, Afifi H. Sileem, Ellen R.Miller und Gregg F. Gunnell: Deciduous dentition and dental eruption sequence of Bothriogenys fraasi (Anthracotheriidae, Artiodactyla) from the Fayum Depression, Egypt. Palaeontologia Electronica 19 (3), 2016, S. 38A ()
- Henry Fairfield Osborn: New carnivorous mammals from the Fayum Oligocene, Egypt. Bulletin of the American Museum of Natural History 26, 1909, S. 415–424.
- Matthew R. Borths und Erik R. Seiffert: Craniodental and humeral morphology of a new species of Masrasector (Teratodontinae, Hyaenodonta, Placentalia) from the late Eocene of Egypt and locomotor diversity in hyaenodonts. PLoS ONE 12 (4), 2017, S. e0173527, doi:10.1371/journal.pone.0173527
- Matthew R. Borths, Patricia A. Holroyd und Erik R. Seiffert: Hyainailourine and teratodontine cranial material from the late Eocene of Egypt and the application of parsimony and Bayesian methods to the phylogeny and biogeography of Hyaenodonta (Placentalia, Mammalia). PeerJ 4, 2016, S. e2639, doi:10.7717/peerj.2639
- Patricia A. Holroyd: New Pterodontinae (Creodonta: Hyaenodontidae) from the late Eocene-early Oligocene Jebel Qatrani Formation, Fayum Province, Egypt. PaleoBios 19 (2), 1999, S. 1–18
- Daniel L. Gebo und David Tab Rasmussen: The Earliest Fossil Pangolin (Pholidota: Manidae) from Africa. Journal of Mammalogy 66 (3), 1985, S. 538–541
- Nancy B. Simmons, Erik R. Seiffert und Gregg F. Gunnell: A New Family of Large Omnivorous Bats (Mammalia, Chiroptera) from the Late Eocene of the Fayum Depression, Egypt, with Comments on Use of the Name “Eochiroptera”. American Museum Novitates 3857, 2016, S. 1–43
- Gregg F. Gunnell, Elwyn L. Simons und Erik R. Seiffert: New Bats (Mammalia: Chiroptera) from the Late Eocene and Early Oligocene, Fayum Depression, Egypt. Journal of Vertebrate Paleontology 28 (1), 2008, S. 1–11
- Gregg F. Gunnell, Sara R. Worsham, Erik R. Seiffert und Elwyn L. Simons: Vampyravus orientalisSchlosser (Chiroptera) from the Early Oligocene (Rupelian), Fayum, Egypt —Body Mass, Humeral Morphology and Affinities. Acta Chiropterologica 11 (2), 2009, S. 271–278
- Gregg F. Gunnell, Nancy B. Simmons und Erik R. Seiffert: New Myzopodidae (Chiroptera) from the Late Paleogene of Egypt: Emended Family Diagnosis and Biogeographic Origins of Noctilionoidea. PLoS ONE 9 (2), 2014, S. e86712, doi:10.1371/journal.pone.0086712
- Jean-Jacques Jaeger, Laurent Marivaux, Mustapha Salem, Awad Abolhassan Bilal, Mouloud Benammi, Yaowalak Chaimanee, Philippe Duringer, Bernard Marandat, Eddy Métais, Mathieu Schuster, Xavier Valentin und Michel Brunet: New rodent assemblages from the Eocene Dur At-Talah escarpment (Sahara of central Libya): systematic, biochronological, and palaeobiogeographical implications. Zoological Journal of the Linnean Society 160, 2010, S. 195–213
- Hesham M. Sallama, Erik R. Seiffert, Michael E. Steiper und Elwyn L. Simons: Fossil and molecular evidence constrain scenarios for the early evolutionary and biogeographic history of hystricognathous rodents. PNAS 106 (39), 2009, S. 6722–16727
- Hesham M. Sallam, Erik R. Seiffert und Elwyn L. Simons. Craniodental Morphology and Systematics of a New Family of Hystricognathous Rodents (Gaudeamuridae) from the Late Eocene and Early Oligocene of Egypt. PLoS ONE 6 (2), 2011, S. e16525, doi:10.1371/journal.pone.0016525
- Hesham M. Sallam, Erik R. Seiffert und Elwyn L. Simons: A basal phiomorph (Rodentia, Hystricognathi) from the late Eocene of the Fayum Depression, Egypt. Swiss Journal of Palaeontology 131, 2012, S. 283–301
- Sharouk F. Al-Ashqar, Erik R. Seiffert, Dorien de Vries, Sanaa El-Sayed, Mohamed S. Antar und Hesham M. Sallam: New phiocricetomyine rodents (Hystricognathi) from the Jebel Qatrani Formation, Fayum Depression, Egypt. PeerJ 9, 2021, S. e12074, doi:10.7717/peerj.12074
- Hesham M. Sallam und Erik R. Seiffert: New phiomorph rodents from the latest Eocene of Egypt, and the impact of Bayesian "clock"-based phylogenetic methods on estimates of basal hystricognath relationships and biochronology. PeerJ 4, 2016, S. e1717, doi:10.7717/peerj.1717
- Hesham M. Sallam und Erik R. Seiffert: Revision of Oligocene ‘Paraphiomys’ and an origin for crown Thryonomyoidea (Rodentia: Hystricognathi: Phiomorpha) near the Oligocene–Miocene boundary in Africa. Zoological Journal of the Linnean Society 190 (1), 2020, S. 352–371, doi:10.1093/zoolinnean/zlz148
- Hesham M. Sallam, Erik R. Seiffert, Elwyn L. Simons und Chlöe Brindley: A Large-Bodied Anomaluroid Rodent from the Earliest Late Eocene of Egypt: Phylogenetic and Biogeographic Implications. Journal of Vertebrate Paleontology 30 (5), 2010, S. 1579–1593
- Hesham M. Sallam, Erik R. Seiffert und Elwyn L. Simons: A highly derived anomalurid rodent (Mammalia) from the Earliest Late Eocene of Egypt. Palaeontology 53 (4), 2010, S. 803–813
- Erik R. Seiffert, Elwyn L. Simons und Yousry Attia: Fossil evidence for an ancient divergence of lorises and galagos. Nature 422, 2003, S. 421–424
- Elwyn L. Simons: Discovery of the smallest Fayum Egyptian primates (Anchomomyini, Adapidae). PNAS 94, 1997, S. 180–184
- Erik R. Seiffert, Elwyn L. Simons, Timothy M. Ryan und Yousry Attia: Additional remains of Wadilemur elegans, a primitive stem galagid from the late Eocene of Egypt. PNAS 102, 2005, S. 11396–11401
- Elwyn L. Simons: Diversity in the early Tertiary anthropoidean radiation in Africa. PNAS 89, 1992, S. 10743–10747
- Elwyn L. Simons und David Tab Rasmussen: A remarkable cranium of Plesiopithecus teras (Primates, Prosimii) from the Eocene of Egypt. PNAS 91, 1994, S. 9946–9950
- Marc Godinot: Lemuriform Origins as Viewed from the Fossil Record. Folia Primatologica 77 (6), 2006, S. 446–464
- Gregg F. Gunnell, Doug M. Boyer, Anthony R. Friscia, Steven Heritage, Fredrick Kyalo Manthi, Ellen R. Miller, Hesham M. Sallam, Nancy B. Simmons, Nancy J. Stevens und Erik R. Seiffert: Fossil lemurs from Egypt and Kenya suggest an African origin for Madagascar’s aye-aye. Nature Communications 9, 2018, S. 3193, doi:10.1038/s41467-018-05648-w
- Erik R. Seiffert, Jonathan M. G. Perry, Elwyn L. Simons und Doug M. Boyer: Convergent evolution of anthropoid-like adaptations in Eocene adapiform primates. Nature 461, 2009, S. 1118–1121
- Erik R. Seiffert, Doug M. Boyer, John G. Fleagle, Gregg F. Gunnell, Christopher P. Heesy, Jonathan M. G. Perry und Hesham M. Sallam: New adapiform primate fossils from the late Eocene of Egypt. Historical Biology 30 (1–2), 2018, S. 204–226, doi:10.1080/08912963.2017.1306522
- Erik R. Seiffert, Elwyn L. Simons, William C. Clyde, James B. Rossie, Yousry Attia, Thomas M. Bown, Prithijit Chatrath und Mark E. Mathison: Basal Anthropoids from Egypt and the Antiquity of Africa’s Higher Primate Radiation. Science 310, 2005, S. 300–304
- Elwyn L. Simons und Richard F. Kay: Qatrania, a new basal anthropoid primate from the Fayum, Oligocene of Egypt. Nature 304, 1983, S. 624–626
- Elwyn L. Simons: Crania of Apidium: Primitive Anthropoidean (Primates, Parapithecidae) from the Egyptian Oligocene. American Museum Novitates 3124, 1995, S. 1–10
- E. Christopher Kirk und Elwyn L. Simons: Diets of fossil primates from the Fayum Depression of Egypt: a quantitative analysis of molar shearing. Journal of Human Evolution 40, 2001, S. 203–229
- Timothy M. Ryan, Mary T. Silcox, Alan Walker, Xianyun Mao, David R. Begun, Brenda R. Benefit, Philip D. Gingerich, Meike Köhler, László Kordos, Monte L. McCrossin, Salvador Moyà-Solà, William J. Sanders, Erik R. Seiffert, Elwyn Simons, Iyad S. Zalmout und Fred Spoor: Evolution of locomotion in Anthropoidea: the semicircular canal evidence. Proceedings of the Royal Society of London B 279, 2012, S. 3467–3475
- Elwyn L. Simons: Discovery of the Oldest Known Anthropoidean Skull from the Paleogene of Egypt. Science 247, 1990, S. 1567–1569
- Erik R. Seiffert und Elwyn L. Simons: Last of the oligopithecids? A dwarf species from the youngest primate-bearing level of the Jebel Qatrani Formation, northern Egypt. Journal of Human Evolution 64, 2013, S. 211–215
- Elwyn L. Simons: New fossil apes from Egypt and the initial differntiation of Hominoidea. Nature 205, 1965, S. 135–139
- Elwyn L. Simons, Erik R. Seiffert, Timothy M. Ryan und Yousry Attia: A remarkable female cranium of the early Oligocene anthropoid Aegyptopithecus zeuxis (Catarrhini, Propliopithecidae). PNAS 104 (21), 2007, S. 8731–8736
- Erik R. Seiffert, Elwyn L. Simons, Doug M. Boyer, Jonathan M. G. Perry, Timothy M. Ryan und Hesham M. Sallam: A fossil primate of uncertain affinities from the earliest late Eocene of Egypt. PNAS 107 (21), 2010, S. 9712–9717
- Elwyn L. Simons und Thomas M. Bown: Afrotarsius chatrathi, first tarsiiform primate (?Tarsiidae) from Africa. Nature 313, 1985, S. 475–477
- Erik R. Seiffert: Early Primate Evolution in Afro-Arabia. Evolutionary Anthropology 21, 2012, S. 239–253
- Thomas M. Bown: Ichnofossils and rhizoliths of the nearshore fluvial Jebel Qatrani Formation (Oligocene), Fayum Province, Egypt. Palaeogeography, Palaeoclimatology, Palaeoecology 40, 1982, S. 255–309
- Scott L. Wing, Stephen T. Hasiotis und Thomas M. Bown: First ichnofossils of flank‐buttressed trees (late Eocene), Fayum Depression, Egypt. Ichnos 3, 1995, S. 281–286
- Wilhelm B. Dames: Über eine tertiäre Wirbelthierfauna von der westlichen Insel des Birket-el-Qurūn im Fajum, Aegypten. Sitzungsberichte der Königlich Preussischen Akademie der Wissenschaften zu Berlin Januar-Mai 1883, S. 129–153 ()
- Max Blanckenhorn: Neues zur Geologie und Paläontologie Aegyptens. II. Das Palaeogen. Zeitschrift der Deutschen Geologischen Gesellschaft 52, 1900, S. 403–479 ()
- Max Blanckenhorn: Neue geologisch-stratrigraphische Betrachtungen in Aegypten. Sitzungsberichte der Mathematisch-Physikalischen Classe der K.B. Akademie der Wissenschaften München 32, 1902, S. 353–433 ()
- Sobhi Helal und Katarína Holcová: Response of foraminiferal assemblages on the middle Eocene climatic optimum and following climatic transition in the shallow tropical sea (the south Fayoum area, Egypt). Arabian Journal of Geosciences 10, 2017, S. 43, doi:10.1007/s12517-016-2818-7
- John A. Van Couvering und Judith A. Harris: Late Eocene age of Fayum mammal faunas. Journal of Human Evolution 21, 1991, S. 241–260
- Phillip D. Gingerich: Oligocene age of the Gebel Qatrani Formation, Fayum, Egypt. Journal of Human Evolution 24, 1993, S. 207–218
- John G. Fleagle, Thomas M. Bown, John D. Obradovich und Elwyn L. Simons: Age of the earliest African anthropoids. Science 234 (4781), 1986, S. 1247–1249
- John Kappelman, Elwyn L. Simons und Carl C. Swisher III: New age determinations for the Eocene-Oligocene boundary sediments in the Fayum Depression, Northern Egypt. Journal of Geology 100 (6), 1992, S. 647–667
- Erik R. Seiffert: Revised age estimates for the later Paleogene mammal faunas of Egypt and Oman. PNAS 103 (13), 2006, S. 5000–5005
- Ashraf Mohamed: Die Krusten der Ränder der Fayoum-Depression – Geomorphologische Untersuchungen. Band 1. Ludwig-Maximilians-Universität München, 22003, S. 1–169
- A. El-Araby und A. N. El-Barkooky: Stratigraphic architecture and facies evolution of the Oligocene Gebel Qatrani Formation, Gebel Qatrani, North Fayum, Egypt. Proceedings of the 5th Geology of the Arab World Conference, Cairo University, Kairo, 2000, S. 1175–1208
- Lászlό Kocsis, Emmanuel Gheerbrant, Mustapha Mouflih, Henri Cappetta, Johan Yans und Mbarek Amaghzaz: Comprehensive stable isotope investigation of marine biogenic apatite from the late Cretaceous–early Eocene phosphate series of Morocco. Palaeogeography, Palaeoclimatology, Palaeoecology 394, 2014, S. 74–88
- Johan Yans, M'Barek Amaghzaz, Baâdi Bouya, Henri Cappetta, Paola Iacumin, László Kocsis, Mustapha Mouflih, Omar Selloum, Sevket Sen, Jean-Yves Storme und Emmanuel Gheerbrant: First carbon isotope chemostratigraphy of the Ouled Abdoun phosphate Basin, Morocco; implications for dating and evolution of earliest African placental mammals. Gondwana Research 25, 2014, S. 257–269
- Emmanuel Gheerbrant, Arnaud Schmitt und László Kocsis: Early African Fossils Elucidate the Origin of Embrithopod Mammals. Current Biology 28 (13), 2018, S. 2167–2173, doi:10.1016/j.cub.2018.05.032
- Mohammed Mahboubi, R. Ameur, Jean-Yves Crochet und Jean-Jacques Jaeger: El Kohol (Saharan Atlas, Algeria): A new Eocene mammal locality in northwestern Africa. Palaeontographica, Abteilung A 192, 1986, S. 15–49
- J. Sudre: Nouveaux mammifères éocènes du Sahara occidental. Palaeovertebrata 9, 1979, S. 83–115
- Mohammed Adaci, Mustapha Bensalah, Rodolphe Tabuce, Fateh Mebrouk, Laurent Marivaux, Olga Otero, Djamila Zaoui, Madani Benyoucef und Mahammed Mahboubi: L'Éocène continental du complexe de Gour Lazib (Sahara Nord-Occidental, Algérie). Mémoire du Service Géologique de l'Algérie 19, 2016, S. 63–89
- Jean-Louis Hartenberger, Claude Martinez und Ahmed Ben Said: Découverte de mammifères d’âge Éocène inférieur en Tunisie centrale. Comptes rendus de l'Académie des sciences. Série 2 301, 1985, S. 649–652 ()
- Rodolphe Tabuce, Laurent Marivaux, Mohammed Adaci, Mustapha Bensalah, Jean-Louis Hartenberger, Mohammed Mahboubi, Fateh Mebrouk, Paul Tafforeau und Jean-Jacques Jaeger: Early Tertiary mammals from North Africa reinforce the molecular Afrotheria clade. Proceedings of the Royal Society B 274, 2007, S. 1159–1166
- Emmanuel Gheerbrant: Les mammifères paléocènes du bassin d’Ouarzazate (Maroc). Teil 1: Introduction générale et Palaeoryctidae. Palaeontographica, Abteilung A 224, 1992, S. 67–132; Teil 2: Todralestidae (Proteutheria, Eutheria). Palaeontographica, Abteilung A 231, 1994, S. 133–188; Teil 3: Adapisoriculidae et autres mammifères (Carnivora, ?Creodonta, Condylarthra, ?Ungulata et incertae sedis). Palaeontographica, Abteilung A 237, 1995, S. 39–132
- Emmanuel Gheerbrant: Premières données sur les mammifères «insectivores» de l’Yprésien du bassin d’Ouarzazate. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 187, 1993, S. 225–242
- Emmanuel Gheerbrant, Jean Sudre, Sevket Sen, Claude Abrial, Bernard Marandat, Bernard Sigé und Moniue Vianey-Liaud: Nouvelles données sur les mammifères du Thanétien et de l’Yprésien du Bassin d’Ouarzazate (Maroc) et leur contexte stratigraphique. Palaeovertebrata 27, 1998, S. 155–202
- Rodolphe Tabuce, Sylvain Adnet, Henri Cappetta, Abdelmajid Noubhani und Frédéric Quillevere: Aznag (bassin d’Ouarzazate, Maroc), nouvelle localité à sélaciens et mammifères de l’Eocène moyen (Lutétien) d’Afrique. Bulletin de la Societe géologique de France 176 (4), 2005, S. 381–400
- Erik R. Seiffert: Chronology of the Paleogene mammal localities. In: Lars Werdelin und William Joseph Sanders (Hrsg.): Cenozoic Mammals of Africa. University of California Press, Berkeley, Los Angeles, London, 2010, S. 19–26
- Philippe-Émmanuel Coiffat, Brigitte Coiffat, Jean-Jacques Jaeger und Mohammed Mahboubi: Un nouveau gisement à mammifères fossiles d’âge Éocène supérieur sur le versant sud des Nementcha (Algérie orientale), découverte des plus anciens rongeurs d’Afrique. Comptes rendus de l'Académie des sciences. Série 2 299, 1984, S. 893–898 ()
- Rodolphe Tabuce, Brigitte Coiffait, Philippe-Émmanuel Coiffait, Mohamed Mahboubi und Jean-Jacques Jaeger: A new genus of Macroscelidea (Mammalia) from the Eocene of Algeria: a possible origin for elephant-shrews. Journal of Vertebrate Paleontology 21 (3), 2001, S. 535–546
- Sylvain Adnet, Henri Cappetta und Rodolphe Tabuce: A Middle–Late Eocene vertebrate fauna (marine fish and mammals) from southwestern Morocco; preliminary report: age and palaeobiogeographical implications. Geological Magazine 147 (6), 2010, S. 860–870, doi:10.1017/S0016756810000348
- Samir Zouhri, Philip D. Gingerich, Najia Elboudali, Samira Sebti, Abdelmajid Noubhani, Meriem Rahali und Saïd Meslouh: New marine mammal faunas (Cetacea and Sirenia) and sea level change in the Samlat Formation, Upper Eocene, near Ad-Dakhla in southwestern Morocco. Comptes Rendus Palevol 13, 2014, S. 599–610, doi:10.1016/j.crpv.2014.04.002
- Samir Zouhri, Bouziane Khalloufi, Estelle Bourdoni, France de Lapparent de Broin, Jean-Claude Rage, Leila M'Haïdrat, Philip D. Gingerich und Najia Elboudali: Marine vertebrate fauna from the late Eocene Samlat Formation of Ad-Dakhla, southwestern Morocco. Geological Magazine 15 (7), 2018, S. 1596–1620, doi:10.1017/S0016756817000759
- David Tab Rasmussen, Sefau O. Tshakreen, Miloud M. Abugares und Joshua B. Smith: Return to Dor al-Talha. Paleontological Reconnaissance of the Early Tertiary of Libya. In: J. G. Fleagle und C. C. Gilbert (Hrsg.): Elwyn Simons: A Search for Origins. Springer, 2008, S. 181–196
- Jean-Jacques Jaeger, K. Christopher Beard, Yaowalak Chaimanee, Mustafa Salem, Mouloud Benammi, Osama Hlal, Pauline Coster, Awad A. Bilal, Philippe Duringer, Mathieu Schuster, Xavier Valentin, Bernard Marandat, Laurent Marivaux, Eddy Métais, Omar Hammuda und Michel Brunet: Late middle Eocene epoch of Libya yields earliest known radiation of African anthropoids. Nature 467, 2010, S. 1095–1099
- Rodolphe Tabuce, Jean-Jaques Jaeger, Laurent Marivaux, Mustapha Salem, Awad Abolhassan Bilal, Mouloud Benammi, Yaowalak Chaimanee, Pauline Coster, Bernard Marandat, Xavier Valentin und Michel Brunet: New stem elephant-shrews (Mammalia, Macroscelidea) from the Eocene of Dur at-Talah, Libya. Palaeontology 55 (5), 2012, S. 945–955
- Herbert Thomas, Emmanuel Gheerbrant und Jean-Michel Pacaud: Découverte de squelettes subcomplets de mammifères (Hyracoidea) dans le Paléogène d’Afrique (Libye). Comptes Rendus Palevol 3, 2004, S. 209–217
- Pauline Coster, Mouloud Benammi, Vincent Lazzari, Guillaume Billet, Thomas Martin, Mustafa Salem, Awad Abolhassan Bilal, Yaowalak Chaimanee, Mathieu Schuster, Xavier Valentin, Michel Brunet und Jean-Jacques Jaeger: Gaudeamus lavocati sp. nov. (Rodentia, Hystricognathi) from the early Oligocene of Zallah, Libya: first African caviomorph? Naturwissenschaften 97 (8), 2010, S. 697–706
- Pauline M. C. Coster, K. Christopher Beard, Mustafa J. Salem, Yaowalak Chaimanee, Michel Brunet und Jean-Jacques Jaeger: A new early Oligocene mammal fauna from the Sirt Basin, central Libya: Biostratigraphic and paleobiogeographic implications. Journal of African Earth Sciences 104, 2015, S. 43–55, doi:10.1016/j.jafrearsci.2015.01.006
- Pauline M. C. Coster, K. Christopher Beard, Mustafa J. Salem, Yaowalak Chaimanee und Jean-Jacques Jaeger: New fossils from the Paleogene of central Libya illuminate the evolutionary history of endemic African anomaluroid rodents. Frontiers in Earth Science 3, 2015, S. 56, doi:10.3389/feart.2015.00056
- Spencer G. Mattingly, K. Christopher Beard, Pauline M. C. Coster, Mustafa J. Salem, Yaowalak Chaimanee und Jean-Jacques Jaeger: A new carnivoraform from the early Oligocene of Libya: Oldest known record of Carnivoramorpha in Africa. Journal of African Earth Sciences 172, 2020, S. 103994, doi:10.1016/j.jafrearsci.2020.103994
- Hendrik Jan van Vliet und Gebely Abu el-Kheir: A new Eocene marine mammal site in the Qattara depression (Egypt). Cainozoic Research 7 (1-2), 2010, S. 73–77
- Hendrik Jan van Vliet, Anne S. Schulp, Gebely A. M. M. Abu el-Kheir, Theo M. Paijans, Mark Bosselaers und Charles J. Underwood: A new Oligocene site with terrestrial mammals and a selachian fauna from Minqar Tibaghbagh, the Western Desert of Egypt. Acta Palaeontologica Polonica 62 (3), 2017, S. 509–525
- John Kappelman, David Tab Rasmussen, William J. Sanders, Mulugeta Feseha, Thomas Bown, Peter Copeland, Jeff Crabaugh, John Fleagle, Michelle Glantz, Adam Gordon, Bonnie Jacobs, Murat Maga, Kathleen Muldoon, Aaron Pan, Lydia Pyne, Brian Richmond, Timothy Ryan, Erik R. Seiffert, Sevket Sen, Lawrence Todd, Michael C. Wiemann und Alisa Winkler: Oligocene mammals from Ethiopia and faunal exchange between Afro-Arabia and Eurasia. Nature 426, 2003, S. 549–552
- William J. Sanders, John Kappelmann und D. Tab Rassmussen: New large-bodied mammals from the late Oligocene site of Chilga, Ethiopia. Acta Palaeontologica Polonica 49 (3), 2004, S. 365–392
- David Tab Rasmussen und Mercedes Gutiérrez: A Mammalian fauna from the Late Oligocene of Northwestern Kenya. Palaeontographica, Abteilung A 288 (1-3), 2009, S. 1–52
- Tom Heldal, Elizabeth G. Bloxam, Patrick Degryse, Per Storemyr und Adel Kelany: Gypsum quarries in the Northern Faiyum quarry landscape, Egypt: a geo-archaeological case study. In: N. Abu-Jaber, E. G. Bloxam, P. Degryse und T. Heldal (Hrsg.): QuarryScapes: ancient stone quarry landscapes in the Eastern Mediterranean. Geological Survey of Norway Special publication12, 2009, S. 51–66
- James A. Harrell und Thomas M. Bown: An Old Kingdom basalt quarry at Widan el-Farras and the quarry road to Laker Moeris. Journal of the American Research Center in Egypt 32, 1995, S. 71–91
- Arthur Bedford Orlebar: Some observations on the Geology of the Egyptian desert. Journal of the Bombay Branch of the Royal Society 2, 1845, S. 229–250 ()
- Karl Mayer-Eymar: Die Versteinerungen der tertiaeren Schichten von der westlichen Insel im Birket-el-Qurûn-See (Mittel-Egypten). Palaeontographica 30, 1883, S. 67–77 ()
- Wilhelm B. Dames: Amblypristis Cheops, nov. gen., nov. sp., aus dem Eocän Aegyptens. Sitzungsberichte der Gesellschaft naturforschender Freunde zu Berlin 1888, S. 106–109 ()
- Wilhelm B. Dames: Ueber Zeuglodonten aus Aegypten und die Beziehungen der Archaeoceten zu den übrigen Cetaceen. Geologische und Paläontologische Abhandlungen N. S. 1, 1894, S. 189–222 ()
- Georg Schweinfurth: Reise in das Depressiongebiet im Umkreise des Fajum im Januar 1886. Zeitschrift der Gesellschaft für Erdkunde Berlin 21, 1886, S. 96–149 ()
- Charles W. Andrews: Fossil Mammalia from Egypt. Part I. Geological Magazine 4 (6), 1899, S. 481–484 ()
- Charles W. Andrews: Notes on an Expedition to the Fayum, Egypt, with Descriptions of some new Mammals. Geological Magazine 4 (10), 1903, S. 337–343 ()
- Charles W. Andrews: Further notes on the mammals of the Eocene of Egypt. Geological Magazine, Part I. 5 (1), 1904, S. 109–116 (), Part II. 5 (1), 1904, S. 157–162 (), Part III. 5 (1), 1904, S. 211–215 ()
- Charles W. Andrews: Note on the Gigantic Land-Tortoise (Testudo ammon) from the Upper Eocene of Egypt. Geological Magazine 5 (1), 1904, S. 527–530 ()
- Hugh John Llewellyn Beadnell: A preliminary note on Arsinoitherium zitteli, Beadn. from the Uppere Eocene strata of Egypt. Egyptian Survey Department, Public Works Ministry, 1902 ()
- Charles William Andrews und Hugh John Llewellyn Beadnell: A preliminary note on some new mammals from the Upper Eocene of Egypt. Survey Department, Public Works Ministry, Kairo, 1902, S. 1–9 ()
- Hugh John Llewellyn Beadnell: The Fayum Depression; a Preliminary Notice of a District of Egypt containing a new Palaeogene Fauna. Geological Magazine 4 (8), 1901, S. 540–546 ()
- Charles W. Andrews: Notes on some vertebrate remains collected in the Fayum, Egypt. Geological Magazine 5 (4), 1907, S. 97–100 ()
- Ernst Stromer: Bericht über eine von den Privatdozenten Dr. Max Blanckenhorn und Dr. Ernst Stromer von Reichenbach ausgeführte Reise nach Aegypten. Sitzungsberichte der Mathematisch-Physikalischen Classe der K.B. Akademie der Wissenschaften München 32, 1902, S. 341–352 ()
- Ernst Stromer: Zeuglodon-Reste aus dem oberen Mitteleocän des Fajûm. Beiträge zur Paläontologie und Geologie Österreich-Ungarns und des Orients 15, 1903, S. 65–100 ()
- Ernst Stromer: Die Fischreste des mittleren und oberen Eocäns von Ägypten. Beiträge zur Paläontologie und Geologie Österreich-Ungarns und des Orients 18, 1905, S. 37–58 () und S. 163–192 ()
- Eberhard Fraas: Wüstenreise eines Geologen in Ägypten. Kosmos 3, 1906, S. 263–269
- Henry Fairfield Osborn: The Fayum Expedition of the American Museum. Science 25 (639), 1907, S. 513–516
- Henry Fairfield Osborn: Hunting the ancestral elephant in the Fayum desert. Century Magazine 74, 1907, S. 815–835 ()
- Rodolphe Tabuce: A mandible of the hyracoid mammal Titanohyrax andrewsi in the collections of the Muséum National d'Histoire Naturelle, Paris (France) with a reassessment of the species. Palaeovertebrata 40 (1), 2016, S. e4, doi: 10.18563/pv.40.1.e4
- Rodolphe Tabuce: Mars-Avril 1904, l’expédition oubliée du MNHN au Fayoum (Egypte) et révision de Titanohyrax andrewsi (Mammalia, Hyracoidea). In: Résumés des communications du congrès 2016 de l’Association Paléontologique Française. Elbeuf: 30. März bis 2. April 2016, Musée d’Elbeuf-Fabrique des savoirs, Métropole-Rouen-Normandie, 2016, S. 21
- Elwyn L. Simons: An Anthropoid Frontal Bone from the Fayum Oligocene of Egypt: the Oldest Skull Fragment of a Higher Primate. American Museum Novitates 1976, 1959, S. 1–16