Brychotherium
Brychotherium ist eine Gattung aus der Ordnung der Hyaenodonta, ausgestorbenen fleischfressenden Säugetieren, die möglicherweise den Raubtieren nahe stehen. Die einzigen bisher aufgefundenen Überreste der Gattung stammen aus der Fossillagerstätte des Fayyum im nördlichen Ägypten und datieren in das Obere Eozän vor rund 34 Millionen Jahren. Es handelt sich um mehrere Unterkiefer und Schädelfragmente. Sie repräsentieren relativ kleine Tiere mit einem eher generalisierten Gebiss. Die Gattung wurde im Jahr 2016 wissenschaftlich eingeführt. Der Name Brychotherium fand aber schon vorher teilweise Verwendung.
| Brychotherium | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
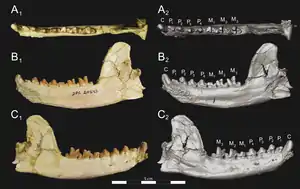
Unterkiefer von Brychotherium, Holotyp | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oberes Eozän (Priabonium) | ||||||||||||
| 34 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Brychotherium | ||||||||||||
| Borths, Holroyd & Seiffert, 2016 | ||||||||||||
Beschreibung
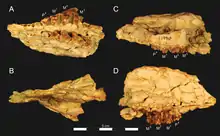
Brychotherium war ein eher kleiner Vertreter der Hyaenodonta mit einem angenommenen Körpergewicht von 5,2 bis 6,2 kg. Die Größe ist damit vergleichbar zu einem Rotfuchs oder Silberdachs. Die Gattung basiert auf mehreren Unterkieferfunden zuzüglich zwei fragmentierter Schädel. Von letzteren ist lediglich jeweils der vordere Teil überliefert. Der Mittelkieferknochen stieg nach hinten steil an, die gemeinsame Knochennaht mit dem Oberkiefer endete etwa auf Höhe des Eckzahns. Durch die Ausrichtung des Mittelkieferknochens war der Naseninnenraum wohl etwa zurückgesetzt, allerdings sind die Nasenbeine nur unvollständig erhalten. Der hintere Abschnitt verbreiterte sich aber deutlich. Der Gesichtsbereich des Oberkiefers war breit, oberhalb des zweiten Prämolaren öffnete sich das Foramen infraorbitale. Der Oberkiefer hatte keinen Anteil am vorderen Rand der Orbita. Dies übernahm stattdessen ein breiter Fortsatz des Tränenbeins. Der untere Rand des Augenfensters wurde durch das Jochbein gebildet. Dieses zeigte auch einen breiten Kontakt zum Tränenbein, beide Knochen schlossen somit den Oberkiefer von der Orbita aus. Den oberen Orbitarand bildete das Stirnbein. Insgesamt lag das Augenfenster mit seinem vorderen Rand oberhalb des ersten Molaren. Am hinteren Abschnitt des Stirnbeins setzte der Scheitelkamm an. Das Gaumenbein war vorn schmal und verbreiterte sich nach hinten. Die Choanen begannen hinter dem dritten Molaren. Die Symphyse des Unterkiefers reichte bis zum dritten Prämolaren, in keinem bisher bekannten Fund war sie verwachsen. Die Unterkante des Unterkiefers wölbte sich leicht konvex, etwa in der Mitte unterhalb des Kronenfortsatzes brach sie über einen leichten Knick und lief dann zum Winkelfortsatz aus. Die Vorderkante des Kronenfortsatzes stand in einem Winkel von rund 125° zur Alveolarfläche. Ein sehr großes Foramen mentale lag unterhalb zwischen dem dritten und vierten Prämolaren, davor befanden sich zwei weitere kleinere jeweils unterhalb des zweiten und ersten Vormahlzahns.[1]
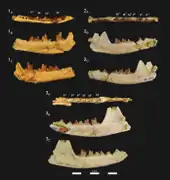
Vom Gebiss sind lediglich die Eckzähne, ein Großteil der Prämolaren und Molaren überliefert, die Position der Schneidezähne kann nur für die obere Gebisshälfte anhand der Alveolen bestimmt werden. Hier waren wenigsten der zweite und der dritte Schneidezahn ausgebildet, die in ihrer Größe von innen nach außen zunahmen. Der Eckzahn war groß und übertraf den letzten Schneidezahn um das Doppelte an Breite. Er hatte eine gepresste konische Gestalt. Die Wurzel des oberen Eckzahns reichte weit zurück in den Knochen, wo sie über dem ersten und zweiten Prämolaren lagerte. In der Regel waren keine Diastemata zwischen den einzelnen Backenzähnen ausgebildet, nur bei zwei Unterkiefern standen der zweite und dritte Prämolar nicht geschlossen. Im Oberkiefer ist lediglich die Krone des letzten Prämolaren erhalten. Die Unterkieferprämolaren zeichneten sich durch einen hohen, spitzen Höcker aus (Protoconid) und wirkten dadurch hoch dreieckig. Bei den beiden mittleren Vormahlzähnen hatte dieser einen etwa asymmetrischen Bau. Der letzte untere Prämolar besaß an den Längsseiten scharfe Scherkanten. An den Molaren, die einen typisch sectorialen Aufbau hatten, waren jeweils drei hohe, spitze Höcker auf der Kauoberfläche ausgeprägt, die im Oberkiefer durch den Para-, Meta- und Protoconus, im Unterkiefer durch den Para-, Meta- und Protoconid gebildet wurden. Wie bei allen Vertretern der Teratodontinae waren im Oberkiefer der Para- und Metaconus der ersten beiden Molaren an der Basis miteinander verschmolzen, die Spitzen standen aber im Unterschied zu den Hyainailourinae frei und deutlich auseinander. Ebenfalls abweichend von den Hyainailourinae ragte der Metaconus üblicherweise über den Paraconus hinaus. Bei Brychotherium war der Paraconus allerdings noch nicht so deutlich reduziert wie bei den nahe verwandten Dissopsalis und Anasinopa. In diesem Merkmal stimmte Brychotherium stärker mit Glibzegdouia übereinstimmt. Eine Ausnahme bildet hier Furodon, das einen höheren Paraconus hatte. Beide Höcker besaßen bei Brychotherium einen eher elliptischen Querschnitt und so eine stärker gepresste Gestalt als bei Anasinopa. Der Protoconus war breit und deutlich niedriger als die beiden anderen Höcker. Die hintere Scherkante des Metaconus (Postmetacrista) führte über eine tiefe Einbuchtung zur Scherkante des Metastyls, einem kleineren Buckel am Zahnende. Der letzte Oberkiefermahlzahn wies auffallende Reduktionen auf und bestand hauptsächlich aus dem Para- und dem Protoconus. Die Molaren des Unterkiefers charakterisierte ein relativ großes Talonid, ein tiefer liegender Bereich der Kauoberfläche. Es wies zwischen 25 und 33 % der Länge des gesamten Zahnes auf und erreichte dadurch nicht die Ausmaße wie bei Glibzegdouia. Der gut entwickelte Metaconid war kleiner als der Proto- und Paraconid, bei Glibzegdouia ragte er deutlich höher. Die Kante zwischen beiden letztgenannten bildete eine tiefe Einbuchtung. Während auf dem vordersten Mahlzahn der Protoconid noch höher wurde als der Paraconid, hatten beide Buckel auf dem letzten Molaren die gleiche Höhe. Sie waren dort aber insgesamt massiver als an den beiden voranliegenden Mahlzähnen. Im Oberkiefer vergrößerten sich die Zähne vom letzten Prämolaren mit 7,1 mm Länge zum zweiten Molaren mit 9,7 mm Länge, der letzte Molar wurde dann nur 3,7 mm lang. Dagegen wies im Unterkiefer der vorderste Prämolar eine Länge von 5,3 mm, der hinterste Molar eine Länge von 9,6 mm auf, die dazwischenliegenden Zähne nahmen graduell an Größe zu.[1]
Fossilfunde

Die bisher bekannten Funde von Brychotherium sind alle im Fayyum-Gebiet im nördlichen Ägypten in Nordafrika gefunden worden. Sie stammen von der Fundstelle L-41 im unteren Abschnitt der Gebel-Qatrani-Formation. Diese datiert paläomagnetischen Untersuchungen zufolge in das Obere Eozän mit absoluten Altersangaben um etwa 34 Millionen Jahren. Es handelt sich um die älteste Fundstelle innerhalb der Formation. Die Ablagerungen bestehen aus grünlich bis grünlich grau gefärbten Ton- und Schluffsteinen, die in eine Sequenz aus gebänderten Sanden eingebettet sind. Die feinkörnigen Ablagerungen stellen Relikte eines ehemaligen Sees dar. Sie werden durch einen hohen Anteil an Evaporiten und durch eine starke Konzentration an Natriumchlorid charakterisiert. In ihrer gesamten Struktur unterscheidet sich die Fundstelle so auffallend von den anderen, eher gröberklastischen, fluviatilen Ablagerungen der Gebel-Qatrani-Formation. Als weitere Besonderheit kann eine hohe Anzahl an Schädeln mit teils assoziierten Partien des Körperskeletts hervorgehoben werden. Die Fossilfunde von L-41 umfassen Reste von Fischen, Amphibien, Reptilien, Vögeln und Säugetieren. Unter anderem ist die hohe Vielfalt an Schliefern, Primaten, Nagetieren und Fledertieren bemerkenswert. Unter den Hyaenodonta sind bisher Akhnatenavus, Masrasector und Brychotherium dokumentiert. Aufgrund der guten Erhaltung von Überresten kleinerer Wirbeltiere hat die Fundstelle L-41 für die Fayyum-Region eine hohe Bedeutung. Ihre Entdeckung geht auf 1983 getätigte Felduntersuchungen eines Forscherteams um Elwyn L. Simons zurück, nachfolgend wurde sie unter anderem von D. Tab Rasmussen genauer vorgestellt.[2][3][4] Brychotherium ist an der Fundstelle bisher mit mehreren Unterkieferteilen und zwei unvollständigen Schädeln belegt. Einer der Schädel lag zusammen mit einem rechten Unterkieferast, hier wird vermutet, dass es sich um ein einzelnes Individuum handelt. Die beiden Schädel gehören zu den ältesten eines Vertreters der Teratodontinae im afroarabischen Raum.[1]
Systematik
Innere Systematik der Teratodontinae nach Solé & Mennecart 2019[5]
|
Brychotherium ist eine Gattung aus der ausgestorbenen Unterfamilie der Teratodontinae, die wiederum einen Teil der ebenfalls erloschenen Ordnung der Hyaenodonta bildet. Die Hyaenodonta formten ursprünglich einen Teil der Creodonta, die teilweise etwas irreführend auch als „Urraubtiere“ bezeichnet werden. Es wurde angenommen, dass die Creodonta die Schwestergruppe der heutigen Raubtiere (Carnivora) innerhalb der übergeordneten Gruppe der Ferae bildeten.[6] Allerdings erwiesen sich die Creodonta als in sich nicht geschlossene Gruppe, sie wurden daher in die Hyaenodonta und die Oxyaenodonta aufgespalten.[7][8] Beiden Gruppen gemeinsam ist die gegenüber den Raubtieren weiter nach hinten im Gebiss verlagerte Brechschere, bei den Hyaenodonten sind zumeist der zweite Oberkiefer- und der dritte Unterkiefermolar involviert. Die Hyaenodonten traten erstmals im Mittleren Paläozän vor rund 60 Millionen Jahren in Erscheinung und starben im Mittleren Miozän vor etwa 9 bis 10 Millionen Jahren aus. Innerhalb der Hyaenodonta stellen die Teratodontinae die Schwestergruppe der Familie der Hyainailouridae dar, beide werden zusammen in die übergeordnete Gruppe der Hyainailouroidea gestellt. Charakteristischerweise sind bei den Teratodontinae der Para- und der Metaconus der Oberkiefermolaren nur an der Basis miteinander verschmolzen, wobei letzterer ersteren überragt. Das unterscheidet sie unter anderem von den Hyainailouridae mit ihren zum Amphiconus verschmolzenen Para- und Metaconus, bei denen außerdem der Paraconus höher ist. Im Bezug auf die Höhenausprägung der beiden Höcker ähneln die Teratodontinae damit den Hyaenodontidae, diese weisen aber ebenfalls verschmolzene Höcker auf. Brychotherium bildet die Schwestergruppe einer Klade aus stammesgeschichtlich jüngeren Vertretern der Teratodontinae wie etwa Dissopsalis, Teratodon und Anasinopa. Mit in dieses nähere Verwandtschaftsverhältnis gehört auch Masrasector, ein sehr kleiner Angehöriger der Teratodontinae, der ebenfalls im Fayyum nachgewiesen wurde.[9][1][10]
Der Gattungsname Brychotherium wurde von Patricia A. Holroyd bereits 1994 in ihrer Doktorarbeit geprägt. In der Folgezeit verwendeten ihn einige Wissenschaftler mehrfach informativ,[11] teilweise wurde die Form als afrikanischer Vertreter einer Sinopa-Gruppe gewertet.[12][13] Das damals bekannte Fossilmaterial beschränkte sich aber auf einzelne Unterkiefer. Erst weiteres Material veranlasste Matthew R. Borths im Jahr 2016 zusammen mit Holroyd und Erik R. Seiffert die wissenschaftliche Erstbeschreibung durchzuführen, wodurch die Gattung ihre offizielle Anerkennung erhielt. Den Holotyp (Exemplarnummer CGM 83750) bildet ein rechter Unterkieferast aus dem Fayyum, der noch die Zahnreihe vom Eckzahn bis zum letzten Molaren aufweist. Der Name Brychotherium setzt sich aus den griechischen Wörtern bρύχω (brycho für „zähneknirschen“) und θηρίον (thēríon für „Tier“) zusammen. Gemeinsam mit der Gattung wurde eine Art eingeführt und mit Brychotherium ephalmos bezeichnet. Das Artepitheton stammt ebenfalls aus dem Griechischen (έϕαλμος ephalmos) und bedeutet „mariniert“. Es bezieht sich auf die stark salzhaltigen Eigenschaften der Fundstelle L-41.[1]
Literatur
- Matthew R. Borths, Patricia A. Holroyd und Erik R. Seiffert: Hyainailourine and teratodontine cranial material from the late Eocene of Egypt and the application of parsimony and Bayesian methods to the phylogeny and biogeography of Hyaenodonta (Placentalia, Mammalia). PeerJ 4, 2016, S. e2639, doi:10.7717/peerj.2639
Einzelnachweise
- Matthew R. Borths, Patricia A. Holroyd und Erik R. Seiffert: Hyainailourine and teratodontine cranial material from the late Eocene of Egypt and the application of parsimony and Bayesian methods to the phylogeny and biogeography of Hyaenodonta (Placentalia, Mammalia). PeerJ 4, 2016, S. e2639, doi:10.7717/peerj.2639
- Elwyn L. Simons: Description of two genera and species of Late Eocene Anthropoidea from Egypt. PNAS 86, 1989, S. 9956–9960
- D. Tab Rasmussen und Elwyn L. Simons: The oldest hyracoids (Mammalia: Pliohyracidae): new species of Saghatherium and Thyrohyrax from the Fayum. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 182, 1991, S. 187–209
- Hesham M. Sallam, Afifi H. Sileem, Ellen R.Miller und Gregg F. Gunnell: Deciduous dentition and dental eruption sequence of Bothriogenys fraasi (Anthracotheriidae, Artiodactyla) from the Fayum Depression, Egypt. Palaeontologia Electronica 19 (3), 2016, S. 38A ()
- Floréal Solé und Bastien Mennecart: A large hyaenodont from the Lutetian of Switzerland expands the body mass range of the European mammalian predators during the Eocene. Acta Palaeontologica Polonica 64, 2019, doi:10.4202/app.00581.2018
- Kenneth D. Rose: The beginning of the age of mammals. Johns Hopkins University Press, Baltimore, 2006, S. 1–431 (S. 122–126)
- Michael Morlo, Gregg Gunnell und P. David Polly: What, if not nothing, is a creodont? Phylogeny and classification of Hyaenodontida and other former creodonts. Journal of Vertebrate Paleontology 29 (3 suppl), 2009, S. 152A
- Floréal Solé: New proviverrine genus from the Early Eocene of Europe and the first phylogeny of Late Paleocene-Middle Eocene hyaenodontidans (Mammalia). Journal of Systematic Paleontology 11, 2013, S. 375–398
- Floréal Solé, Eli Amson, Matthew Borths, Dominique Vidalenc, Michael Morlo und Katharina Bastl: A New Large Hyainailourine from the Bartonian of Europe and Its Bearings on the Evolution and Ecology of Massive Hyaenodonts (Mammalia). PLoS ONE 10 (9), 2015, S. e0135698, doi:10.1371/journal.pone.0135698
- Matthew R. Borths und Nancy J. Stevens: The first hyaenodont from the late Oligocene Nsungwe Formation of Tanzania: Paleoecological insights into the Paleogene-Neogene carnivore transition. PLoS ONE 12 (10), 2017, S. e0185301, doi:10.1371/journal.pone.0185301
- Floréal Solé, Julie Lhuillier, Mohammed Adaci, Mustapha Bensalah, Mohammed Mahboubi und Rodolphe Tabuce: The hyaenodontidans from the Gour Lazib area (?Early Eocene, Algeria): implications concerning the systematics and the origin of the Hyainailourinae and Teratodontinae. Journal of Vertebrate Paleontology 12 (3), 2014, S. 303–322
- Naoko Egi, Patricia A. Holroyd, Takehisa Tsubamoto, Aung Naing Soe, Masanaru Takai und Russell L. Ciochon: Proviverrine hyaenodontids (Creodonta: Mammalia) from the Eocene of Myanmar and a phylogenetic analysis of the proviverrines from the para-Tethys area. Journal of Systematic Palaeontology 3 (4), 2005, S. 337–358
- Rajendra S. Rana, Kishor Kumar, Shawn P. Zack, Floreal Solé, Kenneth D. Rose, Pieter Missiaen, Lachham Singh, Ashok Sahni und Thierry Smith: Craniodental and Postcranial Morphology of Indohyaenodon raoi from the Early Eocene of India, and Its Implications for Ecology, Phylogeny, and Biogeography of Hyaenodontid Mammals. Journal of Vertebrate Paleontology 35 (5), 2015, S. e965308, doi:10.1080/02724634.2015.965308