Goldmulle
Die Goldmulle (Chrysochloridae) sind eine im südlichen und teilweise im mittleren Afrika lebende Familie der Säugetiere. Die Tiere ähneln in ihrem Körperbau den Maulwürfen und führen auch eine ähnliche unterirdische Lebensweise, sind mit diesen jedoch nicht näher verwandt.
| Goldmulle | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
.JPG.webp)
Riesengoldmull (Chrysospalax trevelyani) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Chrysochloridae | ||||||||||||
| Gray, 1825 |
Der Körper zeigt spezielle Anpassungen an eine grabende Lebensweise. Er ist spindel- bis rautenförmig gestaltet, äußerlich sichtbare Ohren und ein Schwanz fehlen, die Augen liegen unter dem Fell verborgen. Vor allem die Vordergliedmaßen sind besonders gut zum Graben geeignet. Sie haben einen kurzen und kräftigen Bau, an den Händen sind große Grabklauen ausgebildet. Der Unterarm verfügt über einen dritten Knochen, die Anzahl der Finger- und Handwurzelknochen ist durch charakteristische Verwachsungen reduziert. Ein kennzeichnendes Merkmal der Goldmulle stellt auch das seidige Fell dar, das teilweise bei bestimmtem Lichteinfall metallisch glänzt.
Goldmulle bewohnen offene bis wüstenartige Regionen und geschlossene Wälder, sie sind sowohl in Tief- als auch in Hochländern anzutreffen. Aufgrund ihrer grabenden Lebensweise stellen sie Habitatspezialisten dar, die teilweise nur eng begrenzte, an zumeist lockeren Untergrund gebundene Lebensräume besiedeln. Über die Lebensweise der Tiere liegen nur in wenigen Fällen genauere Informationen vor. Sie sind nachtaktiv und einzelgängerisch.
Die Nahrung der Goldmulle besteht hauptsächlich aus Wirbellosen wie Regenwürmern und Insekten. Stark vergrößerte Gehörknöchelchen befähigen sie, die Beutetiere anhand der von diesen erzeugten Vibrationen aufzuspüren. Auffallend ist die labile Körpertemperatur, die sich der Umgebung anpasst und daher eine energiesparende Funktion hat. Die Fortpflanzung ist bisher wenig untersucht, pro Wurf kommen ein bis drei Jungtiere zur Welt, die als Nesthocker im unterirdischen Bau verbleiben. Die Individualentwicklung läuft vermutlich sehr langsam ab.
Die Stammesgeschichte der Familie ist nur spärlich belegt. Sie reicht aber bis in das Mittlere Eozän vor etwa 47 Millionen Jahren zurück. Fossilfunde umfassen mit Ausnahmen nur einzelne Schädel- und Gebissreste. Goldmulle sind bereits seit dem 18. Jahrhundert bekannt, sie wurden aber anfangs mit den Maulwürfen gleichgesetzt. Erst in der Wende zum 19. Jahrhundert erfolgte eine Abtrennung von der Gruppe, wobei die Benennung der Familie in das Jahr 1825 datiert.
Aufgrund ähnlicher Lebensweise und vergleichbarer Körpermerkmale galten die Goldmulle bis zum Ende des 20. Jahrhunderts als Mitglied der Ordnung der Insektenfresser (Lipotyphla). Jüngere molekulargenetische Untersuchungen stellen sie zusammen mit den Tenreks in eine gänzlich eigene Säugetiergruppe, die Tenrekartigen (Afrosoricida) innerhalb der Afrotheria. Gegenwärtig umfasst die Familie zehn Gattungen mit 21 Arten, von denen mehr als zwei Drittel in unterschiedlichen Maßen in ihrem Bestand gefährdet sind.
Beschreibung
Habitus
Die Goldmulle sind relativ kleine Vertreter der Säugetiere. Zu den kleinsten Formen gehört der Wüstengoldmull (Eremitalpa granti) mit einer Kopf-Rumpf-Länge von 7,6 bis 8,5 cm, der Riesengoldmull (Chrysospalax trevelyani) wird dagegen 20,8 bis 23,5 cm lang. Das Körpergewicht schwankt dementsprechend zwischen 15 und 40 g beziehungsweise zwischen 410 und 500 g. Die meisten Vertreter besitzen aber Gesamtlängen von 9 bis 14 cm und ein Gewicht von 20 bis 100 g. Die einzelnen Arten weisen starke Ähnlichkeiten im Körperbau auf, unterscheiden sich aber zum Teil erheblich in Bezug auf ihre Fellfärbung, die trotz des Familiennamens der Goldmulle nicht nur goldgelb gefärbt erscheint, sondern auch zwischen schwarzen, grauen oder bräunlichen Farbtönen variiert. Der Körperbau der Goldmulle ist an eine grabende, unterirdische Lebensweise angepasst. Äußerlich ähnelt er etwa dem der Maulwürfe, mit denen die Goldmulle aber nicht näher verwandt sind. Der kompakte Rumpf zeigt einen spindelförmigen bis rautenförmigen Bau, ein Schwanz ist äußerlich nicht sichtbar. Die Nase bedeckt ein derbes, lederartiges Polster, das unterstützend beim Graben eingesetzt wird. Die Nasenlöcher liegen in einer Hautfalte an der Unterseite der Nasenspitze. Wie viele andere grabend lebende Säugetiere haben die Goldmulle keine äußeren Ohrmuscheln, auch sind die Augen von Fell bedeckt. Häufig treten im Gesicht und vor allem im Bereich der Augen akzentuierende helle Farbflecken auf. Die Gliedmaßen sind kurz und kräftig und liegen eher unterhalb des Rumpfes, im Gegensatz zu den Maulwürfen, bei denen sie eher seitlich angebracht sind. Die Vordergliedmaßen weisen vier Zehen auf – der äußere fünfte Strahl ist reduziert – und sind zu Grabwerkzeugen umgebildet. Die Kralle am Mittelfinger (Strahl III) ist stark vergrößert und kann länger als der Unterarm sein. Am ersten und zweiten Finger (Strahl I und II) tragen die Goldmulle jeweils eine kürzere, spitze Kralle, deren Länge aber je nach Art unterschiedlich ausfällt. Lediglich der Wüstengoldmull besitzt auch einen kleinen, krallenartigen Nagel am markant verkürzten vierten Finger (Strahl IV).[1] Die Hinterbeine enden in fünf Zehen, die mit kleinen scharfen Krallen ausgestattet sind. Die einzelnen Zehen werden durch kleine Membranen miteinander verbunden, die Fußsohle ist unbehaart. Weibliche Tiere besitzen zwei Paar Zitzen.[2][3][4]
Schädel- und Gebissmerkmale
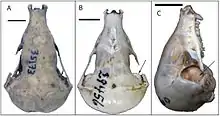
Der Kopf ist kegel- bis tropfenförmig gestaltet, die größte Breite befindet sich im Bereich des Hirnschädels. Das Rostrum ist meist verlängert. Das Verhältnis der größten Breite des Schädels zur größten Länge variiert von Art zu Art. Bei langen und schmalen Schädeln erreicht die größte Schädelbreite nur 60 % der größten Schädellänge, bei kurzen und breiten Schädeln liegt der entsprechende Wert bei bis zu 96 %. Zu den besonders schmalschädeligen Formen gehören die Kupfergoldmulle (Amblysomus), Arends’ Goldmull (Carpitalpa), die Riesengoldmulle (Chrysospalax) oder die Vertreter der Gattungen Neamblysomus und Chlorotalpa. Zu den breitschädeligen zählen dagegen die Kapgoldmulle (Chrysochloris), der Wüstengoldmull oder die Angehörigen der Gattung Cryptochloris. Auch das Rostrum ist dementsprechend unterschiedlich kräftig gestaltet und kann schmal (mit einer Gaumenbreite von circa 28 % der größten Schädellänge) oder breit (mit einem entsprechenden Wert von bis zu 35 %) sein. Auffälligerweise fehlen bei den Goldmullen Einschnürungen im Bereich der Augen. Die Jochbögen sind vollständig geschlossen, das Jochbein ist allerdings zurückgebildet und der Bogen besteht aus einem Knochenfortsatz des Oberkiefers, der mit dem Schläfenbein artikuliert.[5] Als Besonderheit findet sich bei den Goldmullen eine Gelenkverbindung zwischen dem Unterkiefer und dem Zungenbein. Die Gelenkverbindung besteht zwischen dem Winkelfortsatz des Unterkiefers und dem Stylohyale, welches zur Aufhängevorrichtung des Zungenbeins gehört. Eine ähnliche Gelenkung ist von anderen Höheren Säugetieren nicht bekannt. Sie unterstützt eventuell die Zunge beim Zerkauen der Nahrung.[6][2][3]
Eine weitere Auffälligkeit findet sich im Bereich des Mittelohres. Bei einigen Arten sind die Gehörknöchelchen stark vergrößert. Dies betrifft vor allem den Hammer (Malleus), dessen Kopf kugelartig aufgebläht oder keulenartig verlängert sein kann.[7][8] Die Vergrößerungen sind teilweise enorm: beim Wüstengoldmull (Körpergewicht rund 40 g) kann der Malleus ein Gewicht von 70 mg erreichen, beim Riesengoldmull (Körpergewicht rund 500 g) bis zu 185 mg (im Vergleich: beim Menschen mit einem durchschnittlichen Körpergewicht von 70 kg wiegt der Malleus 28 mg). In einigen Fällen geht die Vergrößerung des Hammers mit einer Zunahme der Knochendichte einher, die beim Wüstengoldmull einen Wert von 2,44 g/cm³ erreicht. Dies stellt einen der höchsten Werte bei terrestrisch lebenden Säugetieren dar und wird nur von einigen Walen und Seekühen übertroffen.[9] Zur Aufnahme des aufgeblähten Hammerkopfes ist an der Oberseite der Paukenhöhle eine weitere Nebenhöhle ausgebildet, die sich bei einigen Arten äußerlich sichtbar als Knochenblase über der Schläfengrube hinter der Orbita aufwölbt.[10] Die knöcherne Blase wird aufgrund ihrer Lage am Schläfenbein als temporal bulla (etwa „Schläfenblase“) bezeichnet.[11] Darüber hinaus weist die Hörschnecke einen komplexen Bau auf und besitzt wenigstens vier Windungen mit insgesamt kombiniert 1200° Drehwinkel, was mehr ist als bei zahlreichen anderen, unterirdisch lebenden Säugetieren.[12] Die besonderen Ausprägungen sowohl der Gehörknöchelchen als auch der Hörschnecke stehen in Verbindung mit der auditiven Wahrnehmung der Goldmulle. Die Goldmulle können stark niederfrequente Töne von weniger als einigen hundert Hertz wahrnehmen, ebenso vermögen sie seismische Schwingungen und Vibrationen zu empfangen, etwa von den Bewegungen von Beutetieren. Die Übertragung erfolgt mittels Knochenleitung, die durch die stark asymmetrische Form des Malleus aufgrund des vergrößerten Kopfes und einer damit einhergehenden Verschiebung des Schwerpunktes ausgelöst wird. Die Befähigung ist bei Tieren mit vergrößertem Hammer besser ausgeprägt als bei solchen mit normal gebautem.[13][14][9][15][16]
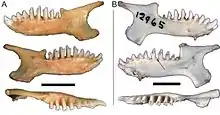
Die Goldmulle haben ein leicht reduziertes Gebiss mit 36 bis 40 Zähnen. Das vordere Gebiss ist vollständig, lediglich im hinteren sind einzelne Backenzähne zurückgebildet. Die Zahnformel lautet demzufolge: . Der vorderste Schneidezahn in der oberen Gebissreihe und der zweite Schneidezahn im Unterkiefer sind vergrößert, die übrigen Schneidezähne haben eine eckzahnähnliche und kleine Gestalt. Der vordere Prämolar kann variabel geformt sein – so kommen sectoriale (mit scharfen Schneidkanten), bicuspide oder tricuspide (mit zwei oder drei Höckerchen) Typen vor. Die Molaren selbst haben auffallend hohe (hypsodonte) Kronen und ein tricuspides Kauflächenmuster. Die drei Haupthöcker stellen der Paraconus, der Metaconus und der Protoconus dar (bezogen auf die Oberkieferzähne). Der Protoconus ist häufig, aber nicht bei allen Arten, in seiner Größe stark reduziert, der Metaconus und der Paraconus stehen eng beieinander – letzterer bildet den Haupthöcker der Mahlzähne. Zusätzlich verläuft eine V-förmige Scherleiste (Ectoloph) über die Zahnfläche, deren Spitze der Paraconus darstellt. Aufgrund der gesamten Konfiguration kann die Zahnstruktur als typisch zalambdodont angesehen werden. An den Unterkiefermolaren ist bei einigen Arten ein Talonid ausgebildet, ein tiefliegender Vorsprung der Kaufläche, in welchen der Protoconus der oberen Backenzähne bei Gebissschluss greift. Sofern der hinterste Molar auftritt, ist dieser überwiegend sehr klein und besitzt entweder die Form der anderen Mahlzähne oder ähnelt einem Stift. Das Merkmal kann aber stark durch die Abkauung der Zähne überprägt sein.[17][18][2][3]

Skelettmerkmale
Die Wirbelsäule setzt sich aus 7 Hals-, 16 Brust-, 3 Lenden-, 5 Kreuzbein- und 9 Schwanzwirbeln zusammen. Die Halswirbelsäule verläuft nach unten gebogen. Die Dornfortsätze der ersten zwölf Brustwirbel stehen senkrecht, die der hinteren Brust- und der Lendenwirbel zeigen nach hinten. Von den sechzehn Rippenpaaren sind acht freistehend. Die vorderen, mit dem Brustbein verbundenen und dieses selbst sind nach innen gerichtet, wodurch Raum für die komplexe Vorderbeinmuskulatur entsteht.[19][3]
Vor allem der vordere Bewegungsapparat zeigt besondere Anpassungen an die grabende Lebensweise. Der gesamte Schultergürtel ist vorverlagert, so dass sich die Schultern etwa auf der Höhe des hinteren Kopfabschnittes befinden. Der so nach vorn verschobene Körperschwerpunkt erlaubt es den Tieren, eine kraftvolle Vorwärtsbewegung beim Graben auszuführen. Schulterblatt und Brustbein sind markant verlängert; ersteres verläuft etwa parallel zur Wirbelsäule und ist zudem außerordentlich schmal. Die stark vergrößerte Schultergräte dient als Ansatzstelle der massigen Schultermuskulatur. Ein Schlüsselbein ist vorhanden, ihm fehlt aber die typische geschwungene Form. Durch die vorgelagerte Position am Körper nimmt es einige Energie beim Graben auf. Der Oberarmknochen fällt durch seine kurze kompakte Form und den gedrehten Schaft auf. Diesem entlang verläuft eine massive deltopectorale Leiste, an der Teile der Arm- und Schultermuskulatur ansetzen. Das Ellenbogengelenk ist stark asymmetrisch geformt; hier sticht der seitlich stark ausladende innere (mediale) Knochenvorsprung des Humerus (Epicondylus medialis) hervor. Dadurch kann die Breite des unteren Gelenkendes des Oberarmknochens 65 % und mehr der gesamten Knochenlänge betragen. Der Grad der Ausbildung des Epicondylus medialis hängt mit der Intensität der Grabungsaktivität der einzelnen Arten zusammen.[20] Ebenso ist das Olecranon, der obere Gelenkfortsatz der Elle, stark ausgezogen und auffallend geschwungen. Es macht etwa 35 % der gesamten Knochenlänge aus. An beiden Gelenkfortsätzen setzt die Armstreckermuskulatur des Unterarmes an, beide Knochenvorsprünge verhindern aber auch zu starke seitliche Bewegungen des Arms. Hervorzuheben ist die Bildung eines stabförmigen Knochens, der gewissermaßen einen dritten Unterarmknochen darstellt. Er entstand wohl aus der Verknöcherung einer Sehne eines Unterarmmuskels, möglicherweise des Musculus flexor digitorum profundus, welcher die Mittelfinger bewegt. Aus diesem Grund wird er häufig als „Flexor“-Knochen bezeichnet. Der „Flexor“-Knochen liegt unterhalb der Elle und besitzt etwa die Größe der Speiche. Die Anzahl der Handwurzelknochen und der Fingerglieder ist reduziert. Die Handwurzelknochen zeichnen sich durch eine abgeflachte Form aus, was die Rotationsfähigkeit der Hand einschränkt. Von den vier Strahlen der Hand bestehen der erste, zweite und vierte aus je einem Mittelhandknochen und zwei Phalangen, wobei die jeweils vordere Phalanx ein Resultat der Verwachsung des ersten und zweiten Fingerglieds ist. Der hypertrophierte dritte Strahl stellt eine Fusion aus dem Mittelhandknochen und den ersten beiden Fingergliedern dar und wird daher auch teilweise „Triplex“-Knochen genannt. Eine weitere charakteristische Bildung liegt am zweiten Fingerstrahl vor. Hier ist der entsprechende Mittelhandknochen mit dem Großen Vieleckbein und dem Kleinen Vieleckbein zum sogenannten Trapezium-trapezoid-metacarpal-II-Komplex (auch „ttm“- beziehungsweise „Totem“-Knochen genannt) verwachsen. Die einzelnen Knochenverwachsungen bilden sich im Verlauf der Embryonalentwicklung heraus. Das jeweils letzte Fingerglied ist vergrößert und am Ende gespalten, was das Bestehen der Kralle anzeigt. Eine Ausnahme bildet der erste Finger (Daumen), dessen letztes Glied nicht eingekerbt ist.[21][22][19][23]
Gegenüber den vorderen Gliedmaßen finden sich nur wenige charakteristische Merkmale an den hinteren. Schien- und Wadenbein sind oben und im unteren Drittel miteinander verwachsen. Dazwischen befindet sich durch die deutlichen Schaftkrümmungen beider Knochen ein weiter Zwischenraum. Ebenso sind die ersten beiden Phalangen der Strahlen II bis IV miteinander verwachsen, so dass alle fünf Fußstrahlen jeweils nur zwei Zehenglieder aufweisen.[21][23]
Fell
Das Fell besteht aus dem Deckhaar und einer dichten Unterwolle. Es ist zumeist seidig und weich. Die Leithaare sind nach hinten gerichtet und wasserabweisend. Die Einzelhaare des Deckfells werden zwischen 7 und 21 mm lang, der Durchmesser liegt bei 78 bis 190 μm. An der Basis sind sie schmaler als im oberen Drittel. Im Querschnitt weisen sie häufig im oberen Drittel eine flache bis gepresste, an der Basis eine rundliche Form auf, die Haarschuppen zeigen eine wellige Anordnung, die Medulla ist gitterartig strukturiert.[24] Bei einigen Arten tritt ein metallischer Glanz auf, der von rötlich über gelblich, grünlich, bräunlich bis hin zu silbern reicht. Der Glanz entsteht in dem abgeplatteten, oberen Drittel der Haare, wo sehr flache Schuppen in mehreren Schichten ausgebildet sind. Das Licht trifft somit auf eine große ebene Fläche zur Reflexion und wird aufgrund der übereinander angeordneten Schuppenschichten mehrfach gebrochen. Irisierende Farben bei Lebewesen bieten häufig einen Vorteil bei der Partnerwerbung während der Fortpflanzung; dies kann aber bei den Goldmullen aufgrund ihrer blinden Natur ausgeschlossen werden. Möglicherweise ist der Farbschimmer ein Nebeneffekt, der durch die Abflachung und stärkere Schichtung der Haaroberflächen entstand. Was diese Haarveränderungen verursachte ist unbekannt, sie stehen aber eventuell im Zusammenhang mit der Fortbewegung im Untergrund.[25]
Weichteilanatomie
Der Verdauungstrakt ist einfach und schlauchförmig gebaut. Er besitzt bei mittelgroßen Arten wie dem Hottentotten-Goldmull (Amblysomus hottentotus) ein Gewicht von 4,9 g und eine Länge von 43,9 cm. Beim Riesengoldmull wiegt er 24,5 g. Der Magen nimmt insgesamt 11 bis 20 % der Länge des Verdauungstraktes ein, letzterer selbst macht zwischen 9 und 12 % der Gesamtkörpermasse aus. Wie bei vielen anderen insektenfressenden Säugetieren fehlt ein Blinddarm, wodurch eine Trennung zwischen Dickdarm und Dünndarm kaum möglich ist. Der Bereich des Magenpförtners (Pylorus) dehnt sich sehr weit aus. Der gesamte Darmabschnitt ist mit fadenförmigen Fortsätzen (Mikrovilli) bedeckt.[26][27] Das Urogenitalsystem endet wie bei den Kloakentieren in einer einzigen Körperöffnung, der Kloake.[28] Bei den Männchen liegen die Hoden in der Bauchhöhle, ihr Gewicht beträgt zusammen etwa 23 mg. Der Penis ist relativ kurz und misst nur 3 bis 4 mm, Penisstacheln fehlen. Das Acrosom der Spermien ist zurückgebildet, dafür sind am Kopf kleine Widerhaken ausgebildet. Die Weibchen haben eine zweihörnige Gebärmutter (Uterus bicornis). Die Kloake und die im Körperinneren liegenden Hoden stellen Gemeinsamkeiten mit den Tenreks dar und können als Anzeichen für die Verwandtschaft beider Taxa gewertet werden.[29][30] Die Nieren sind einfach gebaut und besitzen eine relativ große Medulla und große Markkegel, die bis in den Harnleiter hineinreichen. Beides spricht dafür, dass die Niere Urin hoch konzentrieren kann.[31] Das Gehirn erreicht bei mittelgroßen Arten ein Volumen von 700 bis 736 mg und ist damit nur etwa halb so groß wie bei vergleichbar großen Rüsselspringern.[32]
Die Augen sind bei den Goldmullen zurückgebildet und unter der Haut verborgen. Bei erwachsenen Tieren ist allerdings noch ein Augapfel vorhanden, der rund 0,5 mm lang und 0,4 mm tief wird. Ebenso sind der Bindehautsack und die Tränendrüsen sowie die Tränenwege entwickelt, jedoch fehlen die Linse, die Iris und die gesamte Augenmuskulatur. Möglicherweise hat der Tränenapparat bei den Goldmullen die gleiche Funktion wie bei anderen Säugetieren und hält das Bindehautgewebe frei von Fremdkörpern. Insgesamt ist das Auge weiter zurückentwickelt als vergleichbar bei den Maulwürfen, aber nicht ganz so stark wie bei den Beutelmullen. Die Anlage des Auges beginnt bei den Goldmullen analog zu den übrigen Säugetieren im Embryonalstadium, eine Weiterentwicklung und ein Wachstum finden aber kaum statt.[33][34] Am Aufbau der Nase sind wenigstens fünf Muskeln beteiligt, die teils am Jochbogen ansetzen und bis zur Nasenspitze reichen. Trotz der kurzen und breiten Form der Nase bei den Goldmullen ist diese somit sehr beweglich und kann als taktiles Organ fungieren, mit dem die Tiere nach Nahrung suchen und graben.
Verbreitung und Lebensraum
Die Goldmulle sind ausschließlich in Afrika südlich der Sahara beheimatet, wobei der Schwerpunkt ihres Verbreitungsgebietes im südlichen Afrika liegt. Mehr als die Hälfte der Arten kommt dabei endemisch in der Republik Südafrika vor; einzelne Vertreter bewohnen auch Gebiete in Namibia, in Lesotho und in Mosambik. Außerhalb des südlichen Afrika sind bislang nur drei Arten belegt: der Kongo-Goldmull (Huetia leucorhina) im zentralen, Stuhlmanns Goldmull (Chrysochloris stuhlmanni) im zentralen und östlichen und der Somalia-Goldmull (Huetia tytonis) im nordöstlichen Afrika. Aufgrund ihrer unterirdischen Lebensweise stellen die Goldmulle Habitatspezialisten dar. Ihre Lebensräume sind dadurch stark zersplittert oder eng begrenzt. Häufig lassen sich die einzelnen Arten nur an wenigen, lokal eng begrenzten Fundpunkten feststellen – die wenigsten Vertreter sind aus einem größeren Verbreitungsgebiet bekannt. Andere Goldmulle wie etwa der Somalia-Goldmull, aber auch Visagies Goldmull (Chrysochloris visagiei), De Wintons Goldmull (Cryptochloris wintoni) oder Van Zyls Goldmull (Cryptochloris zyli) wurden bisher nur über einige wenige Exemplare nachgewiesen.[2][3][4]
Die verschiedenen Arten der Goldmulle lassen sich in zwei unterschiedliche ökologische Gruppen teilen:
- Bewohner trockener halbwüstenartiger bis teils wüstenartiger Regionen, dazu gehören der Wüstengoldmull (Eremitalpa), die Vertreter der Kapgoldmulle (Chrysochloris) des südlichen Afrikas und die Arten der Gattung Cryptochloris
- Bewohner von offenen Gras- und Savannenlandschaften sowie von Wäldern, dazu gehören die Kupfergoldmulle (Amblysomus), Arends’ Goldmull (Carpitalpa), die Riesengoldmulle (Chrysospalax) sowie die Vertreter der Gattungen Neamblysomus, Calcochloris und Chlorotalpa.
Die Tiere sind dabei sowohl in küstennahen Tiefländern als auch in Hochgebirgen in Höhenlagen bis zu 4000 m verbreitet. Grundvoraussetzungen für die Anwesenheit von Goldmullen stellen neben einem ausreichenden Nahrungsangebot auch lockere, durchdringbare Böden dar. Ausbreitungsgrenzen bilden Felslandschaften und Flüsse. Die Anpassungsfähigkeit an durch Menschen veränderte Landschaften ist bei den einzelnen Arten unterschiedlich ausgeprägt. Kommen einige Formen in einer gemeinsamen Region vor, wie das etwa beim Fynbos-Goldmull (Amblysomus corriae) und bei Duthies Goldmull (Chlorotalpa duthieae) sowie beim Highveld-Goldmull (Amblysomus septentrionalis) und bei Sclaters Goldmull (Chlorotalpa sclateri) der Fall ist, bestehen in der Regel unterschiedliche Biotopansprüche.[2][3][4]
Lebensweise



Über die Lebensweise der Goldmulle ist generell relativ wenig bekannt. Das liegt zum einen an der scheuen und unterirdischen Lebensweise und zum anderen an der Seltenheit vieler Arten. Einzelne Aspekte der Lebensweise sind nur bei wenigen Arten wie dem Wüstengoldmull, dem Hottentotten-Goldmull, dem Riesengoldmull oder Stuhlmanns Goldmull sowie dem Kap-Goldmull genauer erforscht, weitere Informationen gehen häufig auf zufällige Beobachtungen zurück.[2][3][4]
Aktivitätszeiten, Grabetätigkeit und Sozialverhalten
Die Goldmulle leben weitgehend unterirdisch. Da das Graben im Untergrund etwa 26-mal kostenintensiver ist als eine oberirdische Fortbewegung,[36] haben sich bei den Goldmullen einige energiesparende Eigenschaften ausgebildet. Der Großteil der Arten ist nachtaktiv, einige wenige können auch tagsüber gesichtet werden. Die Aktivitätszeit richtet sich dabei häufig nach der Umgebungstemperatur. Ursache dafür ist die geringe Befähigung der Tiere, eine stabile Körpertemperatur aufrechtzuerhalten. Die durchschnittliche Körpertemperatur liegt bei 29 bis 33 °C und ist damit sehr gering. Da die Goldmulle bei Ruhe nicht thermoreregulieren und die Wärmeleitfähigkeit hoch ist, kann die Körpertemperatur abhängig von der Aktivität stark schwanken. Sie variiert beispielsweise beim Hottentotten-Goldmull von 27 bis 38 °C, beim Kap-Goldmull von 26 bis 36 °C und beim Wüstengoldmull von 18 bis 32 °C. Dabei bestehen optimale Bedingungen für eine stärkere Aktivität bei Außentemperaturen in einem Bereich der mittleren 20-Grad bis mittleren 30-Grad-Marke, was in etwa der thermoneutralen Zone entspricht. Außerhalb von dieser Temperaturspanne schwankt die Körpertemperatur erheblich und liegt dann teilweise nur wenige Grad über der des umgebenden Bodens, beim Wüstengoldmull beträgt diese Differenz nur 0,7 °C. Bei sehr hohen und sehr niedrigen Außentemperaturen fallen einige Arten in einen Torpor (Starrezustand), etwa der Hottentotten-, der Kap- und der Wüstengoldmull. Darüber hinaus kennzeichnet die Goldmulle eine vergleichsweise niedrige Stoffwechselrate, die deutlich niedriger ist als bei vergleichbar großen insektenfressenden Säugetieren. Der Stoffwechsel steigert sich mit fallenden Außen- und Körpertemperatur, fällt beim Eintreten in den Torpor aber wieder rapide ab. Die niedrige Stoffwechselrate in Verbindung mit der Thermolabilität (schwankende Körpertemperatur) führt zu einer adaptiven Hypothermie, womit die Tiere befähigt sind, ihre Körpertemperatur aus Gründen der Energieersparnis der Umgebungstemperatur anzupassen.[37][38][39][36] In diesem Sinne dient daher die häufig nachtaktive Lebensweise der Goldmulle nicht primär der Vermeidung von Begegnungen mit Fressfeinden, sondern stellt zumindest bei den Bewohnern trockener Habitate eine Anpassung an eine energiesparende Lebensweise dar.[40][2][3][4]
Im Gegensatz zu den Maulwürfen, die ihre Grabetätigkeit durch eine seitliche Rotation des Oberarmes verrichten, sind Goldmulle Scharrgräber und graben sich mit nach vorn gerichteten Armbewegungen vorwärts. Ein Grabungszyklus besteht aus einer Anpressphase mit dem Kopf und dem gesamten Körper, bei der das Bodensubstrat verdichtet wird. Darauf folgen mehrere schaufelartige Grabbewegungen. Die vergrößerten Klauen fungieren ähnlich wie eine Spitzhacke, das lockere Bodenmaterial wird unter dem Bauch geschoben und dann mit den Hinterbeinen weggescharrt. Dieser Ablauf der Grabetätigkeit wiederholt sich zyklisch,[22][19] der Hottentotten-Goldmull beispielsweise kann auf diese Weise je nach Beschaffenheit des Untergrundes täglich 4 bis 12 m weit graben.[37] Viele Arten legen Gänge auf zwei Ebenen an: in Tunneln knapp unterhalb der Erdoberfläche suchen die Tiere nach Nahrung, diese bestehen oft nur temporär und sind oberflächlich als schmale Rippeln erkennbar. Tieferreichende Gänge besitzen einzelne Kammern, die oft durch Tunnel verbunden werden und teilweise mit Pflanzenmaterial wie Gräsern nestartig ausgekleidet werden. Sie dienen als Ruheplätze und zur Aufzucht der Jungen. Der Wüstengoldmull, dessen Lebensraum aus Sanddünen besteht, errichtet hingegen – soweit bekannt – keine dauerhaft beständigen Gänge, was dem lockeren Dünensand geschuldet ist. Die instabilen Grabgänge brechen in der Regel hinter den Tieren ein, so dass der Eindruck entsteht, dass sie durch den Sand „schwimmen“.[19] Von einigen Arten ist bekannt, dass sie manchmal die Baue mit Strandgräbern, Graumullen oder Lamellenzahnratten teilen. Dies könnte ein symbiotisches Verhalten sein, da die Goldmulle mit diesen Tieren nicht in Nahrungskonkurrenz stehen und so der Grabaufwand des einzelnen Tieres verringert wird.[37][2][3][4]
Die meisten Goldmulle leben einzelgängerisch und sind territorial, jedes Tier unterhält ein eigenes Revier. Begegnungen zwischen zwei Tieren resultieren häufig in heftigen Kämpfen, die durch Ringen mit den kräftigen Vorderpfoten oder mit Bissen ausgetragen werden. Dabei stoßen sie hohe Quietschtöne aus. Der eigene Bau wird vehement gegen Artgenossen des gleichen und teilweise auch des anderen Geschlechts verteidigt, an den Rändern können sich die Reviere allerdings überlappen. Lediglich beim Riesengoldmull wird aufgrund von allerdings unbestätigten Berichten über Beobachtungen von mehreren Tieren, die in einem Bau überwinterten, ein sozialeres Verhalten angenommen.[2][3][4]
Ernährung
Die Nahrung der Goldmulle besteht vorwiegend aus Wirbellosen wie Regenwürmern sowie Insekten und deren Larven. Hinzu kommen Hundert- und Tausendfüßer, Asseln, Weichtiere und Spinnen. Manche Arten nehmen auch kleine Wirbeltiere wie Skinke zu sich.[37][41][42] Die Nahrungssuche erfolgt sowohl oberirdisch als auch unterirdisch. Hier spielt wahrscheinlich die Ausprägung des Gehörs und somit des Hammers im Mittelohr eine große Rolle. Arten mit aufgeblähtem Malleus kommen häufiger an die Oberfläche als solche mit normal gebautem. Erstere können verstärkt niederfrequente Töne wahrnehmen und besser Vibrationen orten, was ihnen bei der Nahrungssuche, aber auch bei der Vermeidung von Begegnungen mit Fressfeinden hilft.[43] Viele Arten verlassen aber nach ergiebigen Regenfällen ihre Baue und suchen an der Erdoberfläche nach Fressbarem.[2][3][4]
Aufgrund der niedrigen Stoffwechselrate und einer effizienten Nierenfunktion, wodurch Urin im Wasser hochkonzentriert angereichert werden kann, brauchen die meisten Arten keine Flüssigkeit zu sich zu nehmen. Insbesondere bei Arten, die in trockenen Habitaten leben wie dem Wüstengoldmull, ist diese Fähigkeit entwickelt. Weitere Faktoren zur Reduzierung des Wasserverlustes stellen die nächtliche und unterirdische Lebensweise dar.[31][2][4]
Fortpflanzung
Über die Fortpflanzung der Goldmulle ist wenig bekannt. Bei Arten, die in Regionen mit stark jahreszeitlich schwankenden Klimaverhältnissen leben, gibt es eine feste Paarungszeit, bei anderen kann sie das ganze Jahr über erfolgen. Die Weibchen zumindest mancher Arten können mehrere Würfe im Jahr austragen, die meisten Geburten fallen jedoch in die Regenzeit. Während der Balz geben die Männchen des Hottentotten-Goldmull zwitschernde Laute von sich, heben und senken den Kopf und stampfen mit den Füßen auf den Boden. Unter Laborbedingungen konnte beobachtet werden, wie die Männchen die Weibchen verfolgen, um die Paarung zu erzwingen, wobei es auch zu Todesfällen bei den Weibchen kommen kann.[37] Da die Tiere blind sind, erfolgt die Suche nach einem bereitwilligen Paarungspartner wohl über Rufe und Sekretmarkierungen. Das Weibchen wählt seinen Partner möglicherweise über die Länge des Penis aus. Das Fehlen der Penisstacheln beim Männchen bedingt, dass Weibchen spontan ovulieren.[29][4]
Die Dauer der Tragzeit ist unbekannt. In der Regel werden zwischen ein und drei (meist zwei) Jungtiere zur Welt gebracht. Sie verbleiben zunächst in einer Kammer, die mit Gras ausgelegt ist. Die Neugeborenen sind Nesthocker und zunächst unbehaart und haben weiche Krallen. Über die Individualentwicklung liegen kaum Daten vor. Entsprechende Beobachtungen beim Riesengoldmull lassen annehmen, dass diese recht langsam erfolgt.[44] Vom Kap-Goldmull wird angenommen, dass die Jungen zwei bis drei Monate im Bau des Muttertiers verbleiben. Bemerkenswert ist dabei auch der sehr späte Durchbruch der Dauerzähne, was erst kurz vor dem Erreichen des Erwachsenenalters erfolgt.[11] Informationen zur Lebenserwartung freilebender Tiere liegen nicht vor.[2][3][4]
Natürliche Feinde
Zu den bedeutendsten natürlichen Feinden der Goldmulle zählt die Schleiereule, in deren Gewöllen regelmäßig Reste der Tiere gefunden werden. Weitere Predatoren stellen Greif- und Rabenvögel sowie Schlangen dar. Unter den Säugetieren sind das Weißnackenwiesel oder der Schabrackenschakal zu nennen. Häufig erlegen auch Hauskatzen und -hunde einzelne Tiere, teilweise aber ohne diese zu fressen. Vor allem an der Erdoberfläche sind die Tiere stets alarmiert und fliehen bei geringen Bedrohungen in den nächstgelegenen Bau. Angehörige einiger Arten stellen sich tot, wenn sie berührt werden, etwa die des Rauhaar-Goldmulls.[2][3]
Systematik
Äußere Systematik
Innere Systematik der Afrotheria nach Heritage et al. 2021[45]
|
Die Goldmulle sind eine Familie innerhalb der Ordnung der Tenrekartigen (Afrosoricida), ihre nächsten Verwandten stellen die Tenreks (Tenrecidae) und Otterspitzmäuse (Potamogalidae) dar. Die Tenrekartigen wiederum bilden einen Teil der Überordnung der Afrotheria, einer der vier Hauptlinien innerhalb der Höheren Säugetiere. Bei den Afrotheria handelt es sich um eine überwiegend aus Afrika stammende oder dort heimische Verwandtschaftsgemeinschaft, deren Zusammengehörigkeit vor allem auf molekulargenetischen Untersuchungen beruht, weniger auf anatomischen Gemeinsamkeiten. Es können innerhalb der Afrotheria zwei größere Linien unterschieden werden, die Paenungulata und die Afroinsectiphilia. Erstere umfassen die heutigen Elefanten, die Schliefer und die Seekühe, die Gruppe galt schon länger als eine gemeinsame Abstammungseinheit. Zur zweiten Linie werden neben den Tenrekartigen auch die Rüsselspringer und teilweise das Erdferkel gestellt. Einige genetische Untersuchungen unterstützen eine Schwestergruppenbeziehung der Afrosoricida zu den Rüsselspringern, wobei beide dann unter dem Taxon der Afroinsectivora vereint werden.[46][47][48] Andere wiederum sehen die Tenrekartigen in einer Schwestergruppenposition zu allen anderen Afrotheria, während die Rüsselspringer eine engere Beziehung zum Erdferkel formen.[49] Den molekulargenetischen Untersuchungen zufolge entstanden die Afrotheria in der Oberkreide vor 90,4 bis 80,9 Millionen Jahren. Die Aufspaltung der beiden Hauptgruppen fand etwa 15 Millionen Jahre später statt. Die Afrosoricida traten dann kurz vor der Kreide-Tertiär-Grenze vor etwa 68 Millionen Jahren auf.[48] Dem gegenüber diversifizierten sich die Goldmulle erst im Verlauf des Oligozäns vor rund 28 Millionen Jahren.[50] Der bisher älteste Fossilnachweis von Goldmullen stammt aber bereits aus dem Eozän und wird auf ein Alter von 48 bis 41 Millionen Jahre geschätzt.[51]
Innere Systematik
Innere Systematik der Goldmulle nach Asher et al. 2010[52]
|
Die innere systematische Gliederung der Goldmulle wurde im Lauf der Jahrzehnte unterschiedlich bewertet, was auf einer abweichenden Wichtung von Merkmalen beruhte. Nachdem bereits im 19. Jahrhundert mehrere Gattungen beschrieben worden waren, führte diese Robert Broom Anfang des 20. Jahrhunderts allesamt innerhalb von Chrysochloris, unterteilte die Gattung aber anhand des Vorkommens oder Fehlens einer äußerlich sichtbaren Knochenblase an der Schläfengrube des Schädels und der Anzahl der Zähne (36 oder 40) in vielfacher Weise.[11] In den 1920er Jahren gliederte Austin Roberts die Goldmulle auf Gattungsebene auf, führte mehrere neue Gattungen ein und etablierte einige vorher beschriebene neu,[53] was sich in der Folgezeit durchsetzte. Allerdings kam es in späteren Jahrzehnten zu einem häufigen Zusammenführen und Auftrennen von einzelnen Gattungen, was wiederum aus der unterschiedlichen Betonung von Einzelmerkmalen resultierte. In den meisten Fällen lagen diesen Gliederungsversuchen keine statistischen Erhebungen zugrunde.[54][55] Dies änderte sich erst Mitte der 1990er Jahre mit den Untersuchungen von Gary N. Bronner, der intensive morphometrische[56][57] und zytogenetische[58] Analysen vornahm. Auf Bronner basiert auch weitgehend die heutige systematische Gliederung der Goldmulle auf Art- und Gattungsebene.[59][2][3]
Die genauen verwandtschaftlichen Verhältnisse der einzelnen Gattungen der Goldmulle zueinander blieben lange Zeit ungeklärt und wurden nur in Einzelfällen untersucht. Im Jahr 1957 legte dann Alberto M. Simonetta eine Aufteilung der Familie in drei Unterfamilien vor. Als Basis dafür diente ihm die Ausbildung des Hammers am Mittelohr. Zwar hatten schon im 19. und im beginnenden 20. Jahrhundert zahlreiche Wissenschaftler auf die teils auffallend vergrößerten Gehörknöchelchen hingewiesen, doch war es Clive Forster Cooper, der diese 1928 genauer beschrieb, er erkannte ihren möglichen taxonomischen Wert aber zu diesem Zeitpunkt nicht.[60] Simonetta gliedert die Goldmulle folgendermaßen:[54]

- Amblysominae Simonetta, 1957: mit einem normal gebauten Malleus (Amblysomus, Neamblysomus, Calcochloris, Huetia)
- Chrysochlorinae Gray, 1825: mit einem stark verlängerten Kopf des Malleus (Carpitalpa, Chlorotalpa, Chrysochloris, Cryptochloris)
- Eremitalpinae Simonetta, 1957: mit einem kugelig aufgeblähten Kopf des Malleus (Chrysospalax, Eremitalpa)
Simonetta nahm dabei eine graduelle Entwicklung des Kopfes des Hammers von einem relativ kleinen bei den Amblysominae hin zu einem relativ großen bei den Eremitalpinae an.[54] Andere Autoren sahen später dagegen den kugelig aufgeblähten oder keulenartig verlängerten Malleus-Kopf nur als Variation eines Merkmals, dass sich auch innerhalb der einzelnen Arten und Gattungen nicht eindeutig abtrennen lässt. Sie fassten daher die Eremitalpinae mit den Chrysochlorinae in eine Unterfamilie, den Chrysochlorinae, zusammen.[61][59][3]
Die Methode der Molekulargenetik erbrachte zu Beginn des 21. Jahrhunderts ein von dieser traditionellen Gliederung der Goldmulle etwas abweichendes Bild. Demzufolge lassen sich innerhalb der Familie zwei große Hauptlinien erkennen, die aber nur bedingt mit den Unterfamilien übereinstimmen. Eine Gruppe setzt sich aus den Gattungen Amblysomus und Neamblysomus sowie Carpitalpa zusammen, die ersten beiden verfügen über einen kleinen Hammerkopf, letztere über einen leicht vergrößerten. Sie werden informell als „amblysomine Gruppe“ bezeichnet. Dieser gegenüber steht eine „chrysochlorine Gruppe“ mit Gattungen und Arten mit vergrößertem Kopf des Malleus. Die „chrysochlorine Gruppe“ schließt aber mit Calcochloris und Huetia auch Formen mit normal großem Hammer ein. Eine eher mittlere Stellung zwischen diesen beiden Kladen hält Chlorotalpa, deren Hammerkopf ebenfalls vergrößert ist. Es ist daher anzunehmen, dass die besonderen Ausprägungen im Bereich des Mittelohres nicht auf ein einzelnes Ereignis innerhalb der Stammesgeschichte der Goldmulle zurückgehen, sondern ein stärker homoplastisches Merkmal darstellen, das mehrfach innerhalb der Familie entstand.[52] Im Jahr 2018 wurde ein neuer vorläufiger Gliederungsversuch der Goldmulle vorgestellt. Dieser beinhaltet die drei vorher schon etablierten Unterfamilien. Die Amblysominae würden sich demnach aus den Gattungen Amblysomus, Neamblysomus, Carpitalpa, Chlorotalpa, Chrysospalax und Calcochloris zusammensetzen, die Chrysochlorinae wiederum aus der Gattung Chrysochloris (einschließlich Kilimatalpa und Cryptochloris) und die Eremitalpinae aus der Gattung Eremitalpa. Zusätzlich sieht er mit den Huetinae bestehend aus der Gattung Huetia eine vierte eigenständige Unterfamilie vor.[62]
Überblick über die rezenten und fossilen Gattungen der Goldmulle
Die Familie der Goldmulle gliedert sich in zehn heute lebende Gattungen, zu denen gegenwärtig insgesamt 21 Arten gehören:[2][3][52]
- Familie: Chrysochloridae Gray, 1825
- Amblysomus Pomel, 1848 (Kupfergoldmulle; fünf rezente Arten)
- Calcochloris Mivart, 1867 (Gelber Goldmull; eine rezente Art)
- Carpitalpa Lundholm, 1955 (Arends’ Goldmull; eine rezente Art)
- Chlorotalpa Roberts, 1924 (zwei rezente Arten)
- Chrysochloris Lacépède, 1799 (Kapgoldmulle; drei rezente Arten)
- Chrysospalax Gill, 1883 (Riesengoldmulle; zwei rezente Arten)
- Cryptochloris Shortridge & Carter, 1938 (zwei rezente Arten)
- Eremitalpa Roberts, 1924 (Wüstengoldmull; eine rezente Art)
- Huetia Forcart, 1942 (zwei rezente Art)
- Neamblysomus Roberts, 1924 (zwei rezente Arten)
Studien aus dem Jahr 2015 lassen vermuten, dass innerhalb der Goldmulle eine höhere Artenvielfalt zu erwarten ist. Dies resultiert aus der starken Anpassung an bestimmte Habitate mit Herausbildung von Subpopulationen, die teilweise isoliert voneinander bestehen.[63] Bestätigt wird die Ansicht durch weitere genetische Analysen aus dem Jahr 2018. Deren Ergebnisse befürworten die Anerkennung weiterer Arten, die vor allem die Gattungen Eremitalpa und Amblysomus betreffen. Die Gesamtartanzahl der Goldmulle könnte so auf wenigstens rund zwei Dutzend ansteigen. Darüber hinaus sprechen sich die Studien für einen eigenständigen Gattungsstatus für Kilimatalpa aus, welche bisher innerhalb von Chrysochloris geführt wurde, während Cryptochloris wiederum nur als Untergattung von Chrysochloris aufzufassen wäre.[62]
Fossil sind folgende Gattungen anerkannt:[64][20][65][51][23][66]
- Diamantochloris Pickford, 2015
- Damarachloris Pickford, 2019
- Namachloris Pickford, 2015
- Prochrysochloris Butler & Hopwood, 1957
- Proamblysomus Broom, 1941
Eine weitere Gattung, Eochrysochloris Seiffert, Simons, Ryan, Bown & Attia, 2007, gehört wohl nicht direkt zu den Goldmullen, sondern in ein weiteres Verwandtschaftsfeld, den Chrysochloroidea.[67][51]
Forschungsgeschichte
Taxonomie


Die erste wissenschaftliche Benennung eines Vertreters der Goldmulle erfolgte durch Linnaeus (1707–1778) in seinem Systema Naturae aus dem Jahr 1758. Er benannte den heutigen Kap-Goldmull als Talpa asiatica, womit er diesen einerseits zu den Eurasischen Maulwürfen stellte, andererseits seine Verbreitung in Asien vermutete. Die Angaben, die Linnaeus über den Kap-Goldmull zur Verfügung standen, entnahm er aus Albert Sebas 1734 erschienenem Werk Thesaurus.[68][69] Bernard Germain Lacépède (1756–1825) führte im Jahr 1799 den Gattungsnamen Chrysochloris für den Kap-Goldmull ein und benannte somit erstmals einen Gattungsvertreter der Goldmulle. Der Name Chrysochloris ist dem Griechischen entlehnt und besteht aus den Wörtern χρύσεος (chryseos „golden“) und χλωρός (chlōrós „hellgrün“ oder „frisch“). Allerdings bezeichnet Chloris auch die griechische Göttin der Blüte, die in Flora ihr römisches Gegenstück findet. Der Gattungsname ist dabei nicht als Verweis auf die Fellfarbe zu verstehen, die sehr unterschiedlich sein kann, sondern bezieht sich auf den metallischen Glanz des Fells unter bestimmten Lichtverhältnissen.[2][3] Von Chrysochloris leitet sich auch der Familienname Chrysochloridae ab. Allgemein wird dieser heute John Edward Gray (1800–1875) zugesprochen und auf eine Publikation von ihm aus dem Jahr 1825 verwiesen, in der Gray einen Versuch der Gliederung der Säugetiere in Familien und Triben unternahm. In dieser verwendete Gray allerdings die Bezeichnung Chrysochlorina, mit der er die Goldmulle von den Maulwürfen (Talpina), den Spitzmäusen (Soricina), der Igeln (Erinacina), den Tenreks (Tenrecina) und den Spitzhörnchen (Tupaina) absetzte und alle unter der Familie Talpidae vereinigte.[70] Die heute gültige und richtige Schreibweise Chrysochloridae für die Familie der Goldmulle geht auf St. George Jackson Mivart zurück, der sie im Jahr 1868 erstmals verwendete.[71] Er galt lange Zeit auch als Erstbenenner der Familie.[72]
Zur Stellung der Goldmulle innerhalb der Höheren Säugetiere
Linnaeus hatte 1758 die Goldmulle mit den eurasischen Maulwürfen in Verbindung gebracht. Er ordnete sie in die Gruppe der „Bestiae“, in der er unter anderem auch die Spitzmäuse und die Igel, aber auch die Gürteltiere, die Opossums und Schweine sah.[68] Auch nachdem Lacépède vierzig Jahre später die Goldmulle durch die Etablierung der Gattung Chrysochloris von den Maulwürfen abgesetzt hatte, blieb die enge Gruppierung zu anderen insektenfressenden Tieren bestehen. So sah Johann Karl Wilhelm Illiger sowohl die Goldmulle als auch die Maulwürfe, Spitzmäuse und Igel sowie die Tenreks gemeinsam in einer von ihm als „Subterranea“ benannten Gruppe,[73] womit er erstmals alle jene Taxa zusammenfasste, die später als „Insektenfresser“ bezeichnet werden sollten. Informell benutzte bereits Georges Cuvier im Jahr 1817 den Begriff Les insectivores mit einer ähnlichen Zusammensetzung wie Illiger,[74] die offizielle Bezeichnung Insectivora stammt von Thomas Edward Bowdich aus dem Jahr 1821. Bowdich unterschied innerhalb der Insectivora zwei Gruppen, eine mit langen inneren Schneidezähnen wie bei den Goldmullen und Igeln und eine zweite mit großen Eckzähnen, etwa bei den Maulwürfen und den Tenreks.[75]
Die Zugehörigkeit der Goldmulle zu den Insektenfressern wurde in den folgenden eineinhalb Jahrhunderten nur selten in Frage gestellt, zur Debatte standen weitgehend nur die genaueren verwandtschaftlichen Verhältnisse innerhalb der Ordnungseinheit. Wilhelm Peters arbeitete im Jahr 1863 zwei namenlose Gruppen heraus, eine mit Blinddarm, zu denen er die Rüsselspringer und Spitzhörnchen verwies, und eine ohne, welche alle anderen Insectivora inklusive der Goldmulle einschloss.[76] Diesen beiden Gruppen gab Ernst Haeckel drei Jahre später im zweiten Band seines Werkes Generelle Morphologie der Organismen die formellen Namen Menotyphla (mit Blinddarm) und Lipotyphla (ohne Blinddarm).[77] Einen anderen Ansatz verfolgte Theodore Gill Mitte der 1880er Jahre, indem er innerhalb der Insectivora zwischen den Zalambdodonta und den Dilambdodonta unterschied. Die Aufteilung erfolgte anhand der Zahngestaltung, wobei die Zalambdodonta eine V-förmige Zahnschmelzleiste auf den Backenzähnen besitzen (von griechisch ζα (za-) für eine Übersteigerung und dem griechischen Buchstaben Λ (Lambda)), die Dilambdodonta eine W-förmige (von griechisch δι (di „zwei“) und dem griechischen Buchstaben Λ (Lambda)). Die Goldmulle bildeten dabei zusammen mit den Tenreks und den Schlitzrüsslern die zalambdodonten Insektenfresser, während die Igel, Maulwürfe und Spitzmäuse (und zudem die Rüsselspringer und Spitzhörnchen) zu den dilambdodonten gehörten.[78] Gill erkannte damit eine nähere Verwandtschaft der Goldmulle mit den Tenreks. Die Ansichten von Haeckel und Gill wurden 1910 teilweise von William K. Gregory in seinem Werk The orders of Mammals übernommen. Er führte die eigentlichen Insectivora unter dem Begriff Lipotyphla, die Menotyphla positionierte er dagegen außerhalb der Insektenfresser. Dabei behielt er die Goldmulle, Tenreks sowie die Schlitzrüssler in den Zalambdodonta, teilte die verbliebenen Dilambdodonta aber in die beiden Großgruppen Erinaceomorpha (Igel) und Soricomorpha (Spitzmäuse und Maulwürfe) auf.[79] Abweichend von anderen Autoren schloss Robert Broom die Goldmulle aufgrund der Schädelgestaltung in den Jahren 1915 und 1916 komplett aus den Insektenfressern aus und verneinte auch eine nahe Verwandtschaft dieser zu den Tenreks, er schuf für die Goldmulle eine Ordnungsgruppe unter der Bezeichnung Chrysochloridea.[80][5] Dem widersprach aber George Gaylord Simpson in seiner 1945 veröffentlichten generellen Taxonomie der Säugetiere und sah diesen Schritt als zu radikal an. Er untergliederte die damaligen Insectivora in fünf rezente Überfamilien, von denen eine die Goldmulle unter der Bezeichnung Chrysochloroidea repräsentierten, während er die Tenreks als weitere Überfamilie (Tenrecoidea) sah und diese in relative Nähe zu den Goldmullen stellte.[72] Eine engere Beziehung der beiden Gruppen innerhalb der Insektenfresser wurde später immer wieder thematisiert, etwa von Percy M. Butler 1956, der die Gliederung von Gregory in die beiden Großgruppen der Erinaceomorpha und der Soricomorpha übernahm und die Goldmulle und Tenreks in die Soricomorpha verwies. Für ihn stellten die Goldmulle aufgrund des stark überprägten Schädels infolge der unterirdischen Lebensweise abgeleitete Formen innerhalb der Soricomorpha dar.[81] Zu einem ähnlichen Ergebnis, aber unter anderer Schlussfolgerung, kam Malcolm C. McKenna im Jahr 1975. Er ordnete die Goldmulle ebenfalls den Soricomorpha zu, die sich von diesen aber durch einen vollständigen Jochbogen unterschieden. Da aber das Jochbein fehlt, sah McKenna den geschlossenen Bogen bei den Goldmullen als erneute, sekundäre Bildung an.[82] In seiner Klassifikation der Säugetiere, 1997 zusammen mit Susan K. Bell erschienen, schloss McKenna die Goldmulle allerdings aus den Soricomorpha aus und etablierte sie in einer eignen Ordnungsgruppe innerhalb der Insektenfresser.[83]
Erst Ende der 1990er Jahre ordneten molekulargenetische Untersuchungen die Verwandtschaftsverhältnisse der ursprünglichen Insektenfresser (Insectivora, später Lipotyphla) neu. Demnach bilden die Goldmulle keine monophyletische Einheit mit den Maulwürfen oder den Spitzmäusen, sondern formen zusammen mit den Tenreks eine in Afrika endemisch auftretende Gruppe insektenfressender Tiere, die daraufhin in die eigenständige Ordnung der Tenrekartigen (Afrosoricida; eigentlich übersetzt „afrikanische Spitzmausartige“) verwiesen wurde. Zudem ließ sich eine nähere Verwandtschaft der Tenrekartigen mit den Rüsselspringern, dem Erdferkel, den Elefanten, den Schliefern und den Seekühen und somit zu zumeist afrikanisch beheimateten Tieren erkennen, insgesamt eine äußerlich stark heterogene Gruppe, die die Bezeichnung Afrotheria erhielt.[84][85][46] Nachfolgende Analysen untermauerten dieses Ergebnis, unter anderem durch die Isolierung eines spezifischen Retroposons, des sogenannten AfroSINEs, das alle Vertreter der Afrotheria gemein haben.[86][87] Aus diesem Grund beruhen die Ähnlichkeiten der Goldmulle mit Maulwürfen und anderen Insektenfressern lediglich auf Konvergenz.[85]
Stammesgeschichte
Die fossile Überlieferungsgeschichte der Goldmulle ist spärlich. Alle ausgestorbenen Vertreter ähneln in ihrem Körperbau schon weitgehend den heutigen Arten. Sie besitzen einen tropfenförmigen Schädel mit zehn Zähnen je Kieferhälfte und zeigen, soweit das Körperskelett überliefert ist, Anpassungen an eine grabende Lebensweise. Unterschiede lassen sich häufig nur in verschiedenen Zahnmerkmalen und in der zunehmenden Reduktion des letzten Molaren erkennen.[65]
Eozän
Die bisher frühesten bekannten Funde reichen bis in das Eozän zurück und konnten im südlichen Afrika entdeckt werden. Einige isolierte Molaren, ein Unterkiefer und möglicherweise auch einzelne Oberkieferfragmente stammen aus der Fundlokalität Black Crow im Diamantensperrgebiet von Namibia und werden der Gattung Diamantochloris zugewiesen. Mit einem Alter von etwa 48 bis 41 Millionen Jahren gehören die Funde dem Mittleren Eozän an. Die unteren Zähne zeigen bereits die für Goldmulle typische Verschmelzung von zwei der drei Haupthöcker auf der Kauoberfläche (Paraconid und Metaconid).[51][88] Da zusätzlich noch ein besonders großes Talonid ausgebildet ist, sehen andere Autoren Diamantochloris nicht in der unmittelbaren Vorgängerlinie der heutigen Goldmulle.[89] Die gleiche Fundstelle barg auch einen Oberkieferrest und einzelne isolierte Unterkieferzähne. Sie gehören zu einem Tier mit vergleichbaren Zahnmerkmalen, das jedoch deutlich kleiner als Diamantochloris war. Im Jahr 2019 wurden die Funde als zur Gattung Damarachloris gehörig beschrieben.[66] Etwas jünger ist Namachloris, dessen Nachweis auf mehr als 100 Funden beruht. Entdeckt wurde die Gattung am Eocliff, ebenfalls im Sperrgebiet liegend. Die Entstehung der Formation liegt etwa 41 bis 38 Millionen Jahre in der Vergangenheit, was dem Oberen Eozän entspricht. Der aufgefundenen Säugetierfauna zufolge wäre auch ein etwas jüngeres Alter möglich.[89] Das Fossilmaterial setzt sich neben vollständigen Schädeln und Unterkiefern aus nahezu allen Teilen des Körperskelettes zusammen. Erstmals konnte für einen fossilen Goldmull der „Flexor“-Knochen nachgewiesen werden, ebenso sind der „Triplex“- und der „Totem“-Knochen belegt. Die Gliedmaßen zeigen Anpassungen an eine sandgrabende Lebensweise. Das Fundmaterial geht wahrscheinlich auf Beutereste von Greifvögeln oder Eulen zurück, die hier am Ufer eines ehemaligen Sees auf Jagd gingen.[23]
Oligozän
Aus dem nachfolgenden Oligozän sind Reste von Goldmullen oder deren nächsten Verwandten bisher nur aus der Gebel-Qatrani-Formation des Fayyum-Beckens im nördlichen Ägypten überliefert. Gefunden wurden zwei Unterkieferfragmente, die der Gattung Eochrysochloris angehören. Das geschätzte Alter von 34 bis 30 Millionen Jahren entspricht dem unteren Abschnitt der geologischen Serie. Ein auffälliges Kennzeichen stellen die drei Haupthöcker der Molaren dar, die alle nahezu gleich groß und eigenständig sind. Möglicherweise steht Eochrysochloris dadurch nicht in der direkten Verwandtschaftslinie mit den heutigen Goldmullen, sondern gehört einem Seitenzweig an.[67][51]
Miozän
Danach sind Goldmulle erst wieder mit Prochrysochloris im Verlauf des Miozäns fassbar. Das bisher umfangreichste Material kam in westlichen Kenia im Bereich des Victoriasees zu Tage und setzt sich aus nahezu einem halben Dutzend Schädeln und einigen Unterkieferresten zusammen. Bedeutende Lagerstätten stellen hier die Legetet-Formation und die Kapurtay-Agglomerate dar, die beide dem Unteren Miozän angehören und zwischen 20 und 17,5 Millionen Jahre alt sind. Letztere beinhaltet unter anderem die bedeutende Fundstelle von Songhor. Wie auch bei einigen älteren Fossilfunden der Goldmulle war der hinterste Mahlzahn von Prochrysochloris in seiner Größe noch weitgehend unreduziert, eine markante knöcherne Aufwölbung an der Schläfengrube wie bei den Kapgoldmullen bestand nicht.[64][90] Dieselbe Gattung konnte auch mit einem Unterkieferfragment in Arrisdrift im südlichen Namibia nachgewiesen werden. Der Fund ist etwas jünger als die aus Ostafrika, die fossilführenden Schichten datieren an den Beginn des Mittleren Miozän. Aus der gleichen Region liegen aus den mehr oder weniger gleichalten Fundstellen von Elisabethfeld und Langental weitere Unterkieferfragmente und einzelnen Zähne von Prochrysochloris vor.[91][92]
Pliozän und Pleistozän
Im Pliozän treten erstmals Vorläufer der heutigen Arten in Erscheinung. Aus Langebaanweg im südwestlichen Südafrika, das mit einem Alter von 5 Millionen Jahren am Beginn dieser Phase steht, wurden zwei Vertreter von Chrysochloris dokumentiert. Anhand des umfangreichen Fossilmaterials, das aus Schädel- und Unterkieferresten sowie Teilen des Körperskeletts besteht, kann auf sandgrabende Tiere vergleichbar dem Wüstengoldmull geschlossen werden.[20] In den Übergang zum Pleistozän vor etwa 2,5 Millionen Jahren datieren einige wenige Funde von Amblysomus, Neamblysomus, Chlorotalpa und Chrysospalax. Sie wurden in den bedeutenden südafrikanischen Höhlenfundstellen von Makapansgat[93] und Sterkfontein[94][95][96][97] entdeckt. Aus der Nähe von Sterkfontein, von Bolts Farm, stammt ein 29 mm langer und 20 mm breiter Schädel, der aufgrund seines schmalen Baus an den der Kupfergoldmulle erinnert. Abweichend von diesen besitzt er aber eine aufgewölbte, knöcherne Blase an der Schläfengrube, was bei den heutigen Vertretern der Gattung nicht vorkommt. Die Aufwölbung zeigt zudem seitliche Erweiterungen, was wiederum bei keiner anderen Form der Goldmulle mit derartigen Strukturen belegt ist. Der Schädel wurde daher zu Proamblysomus verwiesen.[94]
Bedrohung und Schutz
Mehrere Arten werden von der IUCN als gefährdet oder bedroht gelistet. Hauptursachen der Bedrohung sind der Verlust des Lebensraumes durch die Ausdehnung der menschlichen Siedlungen allgemein, durch Umwandlung in Felder oder Weidegebiete, durch Bergbau oder durch Infrastrukturmaßnahmen infolge von touristischen Erschließungen bestimmter Regionen. Lokal hat auch die Nachstellung durch Haushunde und Hauskatzen einen Einfluss auf die Populationen. Hinzu kommt, dass viele Arten nur ein kleines Gebiet bewohnen und so besonders anfällig für Störungen sind. Zwölf der 21 Arten sind laut IUCN „vom Aussterben bedroht“ (critically endangered), „stark gefährdet“ (endangered), „gefährdet“ (vulnerable) oder „potentiell gefährdet“ (near threatened), für drei weitere Arten fehlen genaue Daten, diese dürften jedoch auch bedroht sein. Ein größerer Teil der Arten ist in Naturschutzgebieten präsent, in vielen Fällen fehlen aber genauere Informationen zur Biologie der Tiere.[98]
Literatur
- Gary N. Bronner: Family Chrysochloridae Golden-moles. In: Jonathan Kingdon, David Happold, Michael Hoffmann, Thomas Butynski, Meredith Happold und Jan Kalina (Hrsg.): Mammals of Africa Volume I. Introductory Chapters and Afrotheria. Bloomsbury, London, 2013, S. 223–257
- Gary N. Bronner und Nigel C. Bennett: Order Afrosoricida. In: John D. Skinner und Christian T. Chimimba (Hrsg.): The Mammals of the Southern African Subregion. Cambridge University Press, 2005, S. 1–21
- Ronald M. Nowak: Walker’s Mammals of the World. The Johns Hopkins University Press, Baltimore 1999, ISBN 0-8018-5789-9
- Gerhard Storch: Lipotyphla, Insektenfresser. In: Wilfried Westheide, Reinhard Rieger (Hrsg.): Spezielle Zoologie. Teil 2: Wirbel- oder Schädeltiere. Spektrum Akademischer Verlag, Heidelberg – Berlin 2004, 712 Seiten, ISBN 3-8274-0307-3
- William A. Taylor, Samantha Mynhardt und Sarita Maree: Chrysochloridae (Golden moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos. Lynx Edicions, Barcelona 2018, S. 180–203 ISBN 978-84-16728-08-4
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. 3. Ausgabe. The Johns Hopkins University Press, Baltimore 2005, ISBN 0-8018-8221-4
Einzelnachweise
- Robert Broom: Some new and some rare Golden moles. Annals of the Transvaal Museum 20, 1946, S. 329–335
- Gary N. Bronner: Family Chrysochloridae Golden-moles. In: Jonathan Kingdon, David Happold, Michael Hoffmann, Thomas Butynski, Meredith Happold und Jan Kalina (Hrsg.): Mammals of Africa Volume I. Introductory Chapters and Afrotheria. Bloomsbury, London, 2013, S. 223–257
- Gary N. Bronner und Nigel C. Bennett: Order Afrosoricida. In: John D. Skinner und Christian T. Chimimba (Hrsg.): The Mammals of the Southern African Subregion. Cambridge University Press, 2005, S. 1–21
- William A. Taylor, Samantha Mynhardt und Sarita Maree: Chrysochloridae (Golden moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos. Lynx Edicions, Barcelona 2018, S. 180–203 ISBN 978-84-16728-08-4
- Robert Broom: On the structure of the skull in Chrysochloris. Proceedings of the Zoological Society of London 1916, S. 449–459
- Gary N. Bronner, Elizabeth Jones und D. J. Coetzer: Hyoid-dentary articulations in golden moles (Mammalia: Insectivore; Chrysochloridae). Zeitschrift für Säugetierkunde 55, 1990, S. 11–15
- Matthew J. Mason: Morphology of the middle ear of golden moles (Chrysochloridae). Journal of Zoology 260, 2003, S. 391–403
- Matthew J. Mason: Functional Morphology of the Middle Ear in Chlorotalpa Golden Moles (Mammalia, Chrysochloridae): Predictions From Three Models. Journal of Morphology 261, 2004, S. 162–174
- Matthew J. Mason, Sarah J. Lucas, Erica R. Wise, Robin S. Stein und Melinda J. Duer: Ossicular density in golden moles (Chrysochloridae). Journal of Comparative Physiology A 192, 2006, S. 1349–1357
- Joseph Hyrtl: Vergleichende anatomische Untersuchungen über das innere Gehörorgan. Prag, 1845, S. 1–139 (S. 10–14 und 59–60) ()
- Robert Broom: A contribution to the knowledge of the cape golden moles. Transactions of the South African Philosophical Society 18, 1907, S. 283–311 ()
- Nick Crumpton, Nikolay Kardjilov und Robert J. Asher: Convergence vs. Specialization in the Ear Region of Moles (Mammalia). Journal of Morphology 276, 2015, S. 900–914
- Matthew J. Mason und Peter M. Narins: Seismic sensitivity in the Desert golden mole (Eremitalpa granti): A review. Journal of Comparative Psychology 116 (2), 2002, S. 258–263
- Matthew J. Mason: Bone conduction and seismic sensitivity in golden moles (Chrysochloridae). Journal of Zoology 260, 2003, S. 405–413
- U. B. Willi, G. N. Bronner und P. M. Narins: Ossicular differentiation of airborne and seismic stimuli in the Cape golden mole (Chrysochloris asiatica). Journal of Comparative Physiology A 192, 2006, S. 267–277
- Matthew J. Mason: Of mice, moles and guinea pigs: Functional morphology of the middle ear in living mammals. Hearing Research 301, 2013, S. 4–18.
- Robert Broom: Some observations on the dentition of Chrysochloris and on the Tritubercular Theory. Annals of the Natal Museum 2, 1916, S. 129–139 ()
- Robert J. Asher und Marcelo R. Sánchez-Villagra: Locking Yourself Out: Diversity Among Dentally Zalambdodont Therian Mammals. Journal of Mammalian Evolution. 12 (1/2), 2005, S. 265–282
- J. P. Gasc, F. K. Jouffroy, S. Renous und F. von Blottnitz: Morphofunctional study of the digging system of the Namib Desert Golden mole (Eremitalpa granti namibensis): cinefluorographical and anatomical analysis. Journal of Zoology 208, 1986, S. 9–35
- Robert J. Asher und D. Margaret Avery: New Golden Moles (Afrotheria, Chrysochloridae) from the Early Pliocene of South Africa. Palaeontologia Electronica 13 (1), 2010, S. 3A ()
- Märtha Kindahl: The embryonic development of the hand and foot of Eremitalpa (Chrysochloris) granti (Broom). Acta Zoologica 30, 1949, S. 133–152
- Gillian M Puttick und Jennifer U M Jarvis: The Functional Anatomy of the Neck and Forelimbs of the Cape Golden Mole, Chrysochloris Asiatica (Lipotyphla: Chrysochloridae). Zoologica Africana 12, 1977, S. 2, 445–458
- Martin Pickford: Late Eocene Chrysochloridae (Mammalia) from the Sperrgebiet, Namibia. Communications of the Geological Survey of Namibia 16, 2015, S. 153–193
- Michael F. Schneider und Victorino a Buramuge: Atlas of the microscopic hair structure of Southern African shrews, hedgehogs, goldem moles and elephant-shrews (Mammalia). Bonner zoologische Beiträge 54 (3), 2006, S. 103–172
- Holly K. Snyder, Rafael Maia, Liliana D’Alba, Allison J. Shultz, Karen M. C. Rowe, Kevin C. Rowe und Matthew D. Shawkey: Iridescent colour production in hairs of blind golden moles (Chrysochloridae). Biology Letters 8, 2012, S. 393–396
- Julia Boonzaier, Elizabeth L. Van der Merwe, Nigel C. Bennett und Sanet H. Kotzé: Comparative Gastrointestinal Morphology of Three Small Mammalian Insectivores: Acomys spinosissimus (Rodentia), Crocidura cyanea (Eulipotyphla), and Amblysomus hottentotus (Afrosoricida). Journal of Morphology 274 (6), 2013, S. 615–626
- Cassandra Pinheiro, Michaela L. Venter, Lauren Sahd, Gary N. Bronner und Sanet H. Kotzé: Comparative gastrointestinal morphology of seven golden mole species (Mammalia: Chrysochloridae) from South Africa. Journal of Morphology 279, 2018, S. 1776–1786
- Matthew R. E. Symonds: Phylogeny and life histories of the ‘Insectivora’: controversies and consequences. Biological Reviews 80, 2005, S. 93–128
- Tarryn A. Retief, Nigel C. Bennett, Anouska A. Kinahan und Philip W. Bateman: Sexual selection and genital allometry in the Hottentot golden mole (Amblysomus hottentotus). Mammalian Biology 78, 2013, S. 356–360
- J. Michael Bedford, Orin B. Mock und Steven M. Goodman: Novelties of conception in insectivorous mammals (Lipotyphla), particularly shrews. Biological Reviews 79, 2004, S. 891–909
- Laura J. Fielden, Michael R. Perrin und G. C. Hickman: Water metabolism in the Namib desert golden mole, Eremitalpa granti namibiensis (Chrysochloridae). Comparative Biochemistry and Physiology 96A (1), 1990, S. 227–234
- Jason A. Kaufman, Gregory H. Turner, Patricia A. Holroyd, Francesco Rovero und Ari Grossman: Brain Volume of the Newly-Discovered Species Rhynchocyon udzungwensis (Mammalia: Afrotheria: Macroscelidea): Implications for Encephalization in Sengis. PlosOne 8 (3), 2013, S. e58667, doi:10.1371/journal.pone.0058667
- Georgina Sweet: The Eyes of Chrysochloris hottentota and C. asiatica. Journal of Cell Science 53 (2), 1909, S. 327–338
- V. Gubbay: A comparison of the development of the rudimentary eye of Eremitalpa granti (Broom) with that of the normal eye of Elephantulus myurus jamesoni (Chubb). South African Journal of Science 52 (8), 1956, S. 182–186 und 193–198
- R. S. Seymour, P. C. Withers und W. W. Weathers: Energetics of burrowing, running, and free-living in the Namib Desert golden mole (Eremitalpa namibensis). Journal of Zoology 244, 1998, S. 107–117
- M. A. Kuyper: The ecology of the golden mole Amblysomus hottentotus. Mammal Review 15 (1), 1985, S. 3–11
- Laura J. Fielden, J. P. Waggoner, Michael R. Perrin und G. C. Hickman: Thermoregulation in the Namib Desert golden mole, Eremitalpa granti namibensis (Chrysochloridae). Journal of Arid Environments 18, 1990, S. 221–237
- N. C. Bennett und A. C. Spinks: Thermoregulation and metabolism in the Cape golden mole (Insectivora: Chrysochloris asiatica). Journal of Zoology 76, 1995, S. 957–971
- Laura J. Fielden, G. C. Hickman und Michael R. Perrin: Locomotory activity in the Namib Desert golden mole Eremitalpa granti namibensis (Chrysochloridae). Journal of Zoology 226, 1992, S. 329–344
- Laura J. Fielden, Michael R. Perrin und G. C. Hickman: Feeding ecology and foraging behaviour of the Namib Desert golden mole, Eremitalpa granti namibensis (Chrysochloridae). Journal of Zoology 220, 1990, S. 367–389
- Jennifer U. M. Jarvis: Notes on the golden mole, Chrysochloris stuhlmanni Matschie, from the Ruwenzori Mountains, Uganda. East African Wildlife Journal 12, 1974, S. 163–166
- Matthew J. Mason und Peter M. Narins: Seismic Signal Use by Fossorial Mammals. American Zoologist 41 (5), 2001, S. 1171–1184
- Gary N. Bronner: Notes on the early post-natal development of a giant golden mole (Günther, 1875) born in captivity (Mammalia: Insectivora; Chrysochloridae). Koedoe 35 (2), 1992, S. 57–58
- Steven Heritage, Erik R. Seiffert und Matthew R. Borths: Recommended fossil calibrators for time-scaled molecular phylogenies of Afrotheria. Afrotherian Conservation 17, 2021, S. 9–13
- Peter J. Waddell, Hirohisa Kishino und Rissa Ota: A Phylogenetic Foundation for Comparative Mammalian Genomics. Genome Informatics 12, 2001, S. 141–154
- Matjaž Kuntner, Laura J. May-Collado und Ingi Agnarsson: Phylogeny and conservation priorities of afrotherian mammals (Afrotheria, Mammalia). Zoologica Scripta 40 (1), 2011, S. 1–15
- Robert W. Meredith, Jan E. Janečka, John Gatesy, Oliver A. Ryder, Colleen A. Fisher, Emma C. Teeling, Alisha Goodbla, Eduardo Eizirik, Taiz L. L. Simão, Tanja Stadler, Daniel L. Rabosky, Rodney L. Honeycutt, John J. Flynn, Colleen M. Ingram, Cynthia Steiner, Tiffani L. Williams, Terence J. Robinson, Angela Burk-Herrick, Michael Westerman, Nadia A. Ayoub, Mark S. Springer und William J. Murphy: Impacts of the Cretaceous Terrestrial Revolution and KPg Extinction on Mammal Diversification. Science 334, 2011, S. 521–524
- Erik R Seiffert: A new estimate of afrotherian phylogeny based on simultaneous analysis of genomic, morphological, and fossil evidence. BMC Evolutionary Biology 7, 2007, S. 224, doi:10.1186/1471-2148-7-224
- C. Gilbert, P. C. O’Brien, G. Bronner, F. Yang, A. Hassanin, M. A. Ferguson-Smith und T. J. Robinson: Chromosome painting and molecular dating indicate a low rate of chromosomal evolution in golden moles (Mammalia, Chrysochloridae). Chromosome Research 14, 2006, S. 793–803
- Martin Pickford: Chrysochloridae (Mammalia) from the Lutetian (Middle Eocene) of Black Crow, Namibia. Communications of the Geological Survey of Namibia 16, 2015, S. 105–113
- Robert J. Asher, Sarita Maree, Gary Bronner, Nigel C Bennett, Paulette Bloomer, Paul Czechowski, Matthias Meyer und Michael Hofreiter: A phylogenetic estimate for golden moles (Mammalia, Afrotheria, Chrysochloridae). BMC Evolutionary Biology 10, 2010, S. 69, doi:10.1186/1471-2148-10-69
- Austin Roberts: Some additions to the list of South African mammals. Annals of the Transvaal Museum 10 (2), 1924, S. 59–76
- Alberto M. Simonetta: A new golden mole from Somalia with an appendix on the taxonomy of the family Chrysochloridae (Mammalia, Insectivora). Monitore Zoologico Italiano NS Supplement 2, 1968, S. 27–55
- F. Petter: Remarques sur la systematique des Chrysochlorides. Mammalia 45 (1), 1981, S. 49–53
- Gary N. Bronner: Geographic patterns of morphometric variation in the Hottentot golden mole, Amblysomus hottentotus (Insectivora: Chrysochloridae). A multivariate analysis. Mammalia 60 (4), 1996, S. 729–751
- Gary N. Bronner: Non-eographic variation in morphological characteristics of the Hottentot golden mole, Amblysomus hottentotus (Insectivora: Chrysochloridae). Mammalia 60 (4), 1996, S. 707–727
- Gary N. Bronner: Cytogenetic Properties of Nine Species of Golden Moles (Insectivora: Chrysochloridae). Journal of Mammalogy 76 (3), 1995, S. 957–971
- Gary N. Bronner: Systematic revision of the Golden mole genera Amblysomus, Chlorotalpa and Calcochloris (Insectivora, Chrysochloromorpha, Chrysochloridae). University of Natal, Pretoria, 1995, S. 1–346 (S. 268)
- Clive Forster Cooper: On the ear region of certain of the Chrysochloridae. Philosophical Transactions of the Royal Society of London. Series B 435, 1928, S. 265–282
- Acacia von Mayer, G. O’Brian und E. E. Sarmiento: Functional and systematic implications of the ear in golden moles (Chrysochloridae). Journal of Zoology 236m 1995, S. 417–430
- Gary Bronner: An imminent updated (2017) taxonomy for golden moles. Afrotherian Conservation 14, 2018, S. 57–59
- Samantha Mynhardt, Sarita Maree, Illona Pelser, Nigel C. Bennett, Gary N. Bronner, John W. Wilson und Paulette Bloomer: Phylogeography of a Morphologically Cryptic Golden Mole Assemblage from South-Eastern Africa. PlosONE 10 (12), 2015, S. e0144995, doi:10.1371/journal.pone.0144995
- Percy M. Butler und A. Tindell Hopwood: Insectivora and Chiroptera from the Miocene rocks of Kenya Colony. Fossil Mammals of Africa 13, 1957, S. 1–35 ()
- Robert J. Asher: Tenrecoidea. In: Lars Werdelin und William Joseph Sanders (Hrsg.): Cenozoic Mammals of Africa. University of California Press, Berkeley, London, New York, 2010, S. 99–106
- Martin Pickford: New Chrysochloridae (Mammalia) from the middle Eocene of Black Crow, Namibia. Communications of the Geological Survey of Namibia 21, 2019, S. 40–47
- Erik R. Seiffert, Elwyn L. Simons, Timothy M. Ryan, Thomas M. Bown und Yousry Attia: New remains of Eocene and Oligocene Afrosoricida (Afrotheria) from Egypt, with implications for the origin(s) of afrosoricid zalambdodonty. Journal of Vertebrate Paleontology 27 (4), 2007, S. 963–972
- Carl von Linné: Systema naturae. 10. Auflage, 1758, Band 1, S. 53 ()
- Oldfield Thomas: On the probable identity of certain specimens, formerly in the Lidth de Jeude Collection, and now in the British Museum, with those figured by Albert Seba in his 'Thesaurus' of 1734. Proceedings of the Zoological Society 1892, S. 309–318 ()
- John Edward Gray: An outline of an attempt at the disposition of Mammalia into tribes and families, with a list of the genera apparently appertaining to each tribe. Annals of Philosophy N. S. 10, 1825, S. 337–344 ()
- St. George Jackson Mivart: Notes on the Osteology of the Insectivora. Journal of Anatomy and Physiology 2 (1), 1868, S. 117–154 (S. 150) ()
- George Gaylord Simpson: The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History 85, 1945, S. 1–350 (S. 49 und 175–179)
- Johann Karl Wilhelm Illiger: Prodromus Systematis Mammalium et Avium. Berlin, 1811, S. 1–301 (S. 126) ()
- Georges Cuvier: Le regne animal distribué d’apres son organisation pour servir de base a l’histoire naturelle des animaux et d’introduction à l’anatomie comparée. Paris, 1817, S. 1–540 (S. 131–138) ()
- Thomas Edward Bowdich: An analysis of the natural classifications of Mammalia for the use of students and travellers. Paris, 1821, S. 1–115 (S. 31–33) ()
- Wilhelm Peters: Über die Säugethiergattung Selenodon. Abhandlungen der Königlichen Akademie der Wissenschaften zu Berlin 1863, S. 1–22 ()
- Ernst Haeckel: Generelle Morphologie der Organismen. Band 2. Berlin, 1866, S. 1–462 (S. CLX) ()
- Theodore Gill: Insectivora. In: John Sterling Kingsley (Hrsg.): Standard natural history. Volume 5. Boston, 1885, S. 1–535 (S. 134–158) ()
- William King Gregory: The orders of mammals. Bulletin of the American Museum of Natural History 27, 1910, S. 1–524 (S. 464–465)
- Robert Broom: On the Organ of Jacobson and its relations in the “Insectivora”. Part II. Talpa, Centetes and Chrysochloris. Proceedings of the Zoological Society of London 1915, S. 347–354 ()
- Percy M. Butler: The skull of Ictops and the classification of the Insectivora. Proceedings of the Zoological Society of London 126, 1956, S. 453–481
- Malcolm C. McKenna: Toward a phylogenetic classification of the Mammalia. In W. Patrick Luckett und Frederick S. Szalay (Hrsg.): Phylogeny of the primates: a multidisciplinary approach. New York, London, 1975, S. 21–46
- Malcolm C. McKenna und Susan K. Bell: Classification of mammals above the species level. Columbia University Press, New York, 1997, S. 1–631 (S. 272–295)
- Mark S. Springer, Gregory C. Cleven, Ole Madsen, Wilfried W. de Jong, Victor G. Waddell, Heather M. Amrine und Michael J. Stanhope: Endemic African mammals shake the phylogenetic tree. Nature 388, 1997, S. 61–64
- Michael J. Stanhope, Victor G. Waddell, Ole Madsen, Wilfried de Jong, S. Blair Hedges, Gregory C. Cleven, Diana Kao und Mark S. Springer: Molecular evidence for multiple origins of Insectivora and for a new order of endemic African insectivore mammals. PNAS 95, 1998, S. 9967–9972 ()
- Masato Nikaido, Hidenori Nishihara, Yukio Hukumoto und Norihiro Okada: Ancient SINEs from African Endemic Mammals. Molecular Biology and Evolution 20 (4), 2003, S. 522–527
- Hidenori Nishihara, Yoko Satta, Masato Nikaido, J. G. M. Thewissen, Michael J. Stanhope und Norihiro Okada: A Retroposon Analysis of Afrotherian Phylogeny. Molecular Biology and Evolution 22 (9), 2005, S. 1823–1833
- Martin Pickford: Diamantochloris mandible from the Ypresian/Lutetian of Namibia. Communications of the Geological Survey of Namibia 19, 2018, S. 51–65
- Robert J. Asher: Recent additions to the fossil record of tenrecs and golden moles. Afrotherian Conservation 15, 2019, S. 4–13
- Percy M. Butler: Macroscelidea, Insectivora and Chiroptera from the Miocene of East Africa. Palaeovertebrata 14 (3), 1984, S. 117–200
- Pierre Mein und Martin Pickford: Insectivora from Arrisdrift, a basal Middle Miocene locality in southern Namibia. Memoir of the Geological Survey of Namibia 19, 2003, S. 143–146
- Pierre Mein und Martin Pickford: Early Miocene Insectivores from the Northern Sperrgebiet, Namibia. Memoir of the Geological Survey of Namibia 20, 2008, S. 169–183
- G. de Graaff: A new chrysochlorid from Makapansgat. Palaeontologia Africana 5, 1958, S. 21–27
- Robert Broom: On two Pleistocene Golden moles. Annals of the Transvaal Museum 20, 1941, S. 215–216
- Diana M. Avery: An assessment of the Lower Pleistocene micromammalian fauna from Swartkrans Members 1-3, Gauteng, South Africa. Geobios 31 (3), 1998, S. 393–414
- Diana M. Avery: Notes on the systematics of micromammals from Sterkfontein, Gauteng, South Africa. Palaeontologia Africana 36, 2000, S. 83–90
- Diana M. Avery: The Plio-Pleistocene vegetation and climate of Sterkfontein and Swartkrans, South Africa, based on micromammals. Journal of Human Evolution 41, 2001, S. 113–132
- IUCN: The IUCN Red List of Threatened Species. Version 2015. (), abgerufen am 24. Juni 2016
Weblinks
- Goldmulle bei der Afrotheria Specialist Group
- Goldmulle bei Animal Diversity Web: Informationen (englisch) und Abbildungen