Gomphotherium
Gomphotherium ist eine ausgestorbene Rüsseltiergattung aus der Familie der Gomphotherien (Gomphotheriidae). Sie war im Miozän und Pliozän über große Teile Eurasiens und Nordamerikas verbreitet, hat ihren Ursprung aber im Oligozän in Afrika. Fossilreste dieses Rüsseltiers werden häufig gefunden, in der Regel handelt es sich aber um Einzelfunde. Bedeutend ist das nahezu vollständige Skelett von Gweng bei Mühldorf östlich von München, das eine Widerristhöhe von über 3 m aufweist. Die meisten Vertreter blieben aber deutlich darunter. Der insgesamt elefantenähnliche Körperbau zeigt Besonderheiten am Schädel, der deutlich flacher und länger war als bei den heutigen Rüsseltieren und vier Stoßzähne aufwies. Als ausgesprochener Pflanzenfresser ernährte sich das Tier überwiegend von gemischter Pflanzenkost. Forschungsgeschichtlich wurde Gomphotherium in der Regel zu den „Mastodonten“ gestellt, einer heute nicht mehr anerkannten Gruppe urtümlicher Rüsseltiere. In der heutigen Systematik der Rüsseltiere bildet es einen Teil der Familie der Gomphotheriidae, einer den modernen Elefanten zeitlich und entwicklungsgeschichtlich vorangehenden Gruppe. Insgesamt war Gomphotherium eine sehr arten- und formenreiche Gattung, die recht häufig auftrat. Aufgrund des meist nur bruchstückhaften Fundmaterials gibt es aber noch Abgrenzungsprobleme zwischen den einzelnen Arten.
| Gomphotherium | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelettrekonstruktion von Gomphotherium | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oberes Oligozän bis Oberes Pliozän | ||||||||||||
| 28,1 bis 3,6 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Gomphotherium | ||||||||||||
| Burmeister, 1837 | ||||||||||||
Merkmale
Größe
Gomphotherium war ein großes Rüsseltier mit langgestrecktem, aber niedrigem Schädel, vier Stoßzähnen (je zwei im Ober- und im Unterkiefer) und einem allgemein elefantenähnlichen Körperbau. In seinen Ausmaßen erreichte es etwa die Größe heutiger Asiatischer Elefanten. Ein relativ vollständiges Skelett aus Gweng, östlich von München gelegen, das zu den größten bekannten dieser Gattung zählt, wird auf eine Schulterhöhe von 3,05 m und auf eine Kopf-Rumpf-Länge von rund 5 m rekonstruiert (jeweils ohne Weichteilgewebe).[1][2] Das Gewicht dieses Rüsseltiervertreters wird mit etwa 3,9 bis 4,7 t angegeben.[3]
Schädel- und Gebissmerkmale

Der Schädel von Gomphotherium war sehr groß, ist in der Regel aber meist nur fragmentiert überliefert. Charakteristisch war die gegenüber heutigen Elefanten längere Streckung, was weitgehend durch den niedrigeren und weniger domartig wirkenden Oberschädel hervorgerufen wurde. Die Knochen des Oberschädels besaßen luftgefüllte Kammern, die einerseits das Gewicht des gesamten Kopfes verringerten, andererseits auch durch die vergrößerte Oberfläche einer wesentlich kräftigeren Muskulatur als Ansatzstelle dienten. Dies war notwendig, um den enormen Kopf mit den vier Stoßzähnen zu stabilisieren. Das Hinterhauptsbein war kurz und stand senkrecht, in der Ansicht von hinten besaß es einen trapezförmigen Querschnitt und war bis zu 77 cm breit. Das Nasenbein saß relativ weit oben am Vorderschädel und besaß leicht erhöhte Knochenstellen am Nasenloch als Ansatzstellen des Rüssels. Die Zahnfächer der oberen Stoßzähne hatten abweichend von den heutigen Elefanten eine deutlich nach vorn gerichtete Position.[1]
Der charakteristische und äußerst massive Unterkiefer erreichte eine Länge von bis zu 129 cm und war dadurch wesentlich langgestreckter als bei den heutigen Elefanten. Die Längsstreckung erfolgte hauptsächlich durch die Ausbildung der unteren Stoßzähne, deren Zahnfächer seitlich an der Symphyse ansetzten und weitgehend horizontal verliefen. Die Symphyse selbst war äußerst robust und bis zu 52 cm lang und reichte bis zum Beginn der hinteren Bezahnung. Der Unterkiefer insgesamt war schmal, die beiden Äste trafen in einem Winkel von 40° aufeinander. Die Höhe des Unterkieferkörpers betrug am vorderen Bereich des Molaren rund 21,5 cm. Hier klafften die beiden Äste etwa 10 cm auseinander, die größte Breite erreichte der Unterkiefer an den beiden quer stehenden Gelenkenden, die an den Innenkanten einen Abstand von 26, an den Außenkanten von 62 cm aufwiesen.[1]

Wie die heutigen Elefanten verfügte Gomphotherium über einen horizontalen Zahnwechsel, der bewirkte, dass in der Regel nur jeweils ein funktionaler Backenzahn je Kieferbogen vorhanden war, insgesamt also vier. Dies setzt das Tier auch von den älteren großen Rüsseltiervertretern wie Deinotherium ab, die noch einen vertikalen, also den meisten heutigen Säugetieren gemeinen Zahnwechsel aufwiesen und so alle Zähne zur gleichen Zeit in Funktion hatten. Beim horizontalen Zahnwechsel schob sich ein neuer Zahn erst heraus, wenn der voranliegende weitgehend abgekaut war. Die Zahnformel für ausgewachsene Individuen von Gomphotherium lautet einschließlich der Stoßzähne: . Prämolaren waren in der Phase des Milchgebisses ausgebildet und umfassten insgesamt drei je Kieferbogen (dP2-4). Zusätzlich bestanden je Kieferbogen noch zwei dauerhafte Prämolaren, die sich zeitlich vor (P3) und nach (P4) dem ersten Molaren hervorschoben. Gomphotherium konnte also nach dem Durchbruch des ersten Backenzahnes insgesamt sieben Mal die Zähne wechseln. In ihrer Morphologie näherten sich die Prämolaren schon stark den späteren dauerhaften Molaren an und waren niederkronig (brachyodont) sowie durch höckerige Zahnschmelzbuckel auf der Kauoberfläche charakterisiert (bunodont). Jeweils ein Paar dieser Höcker bildeten eine Leiste auf der Kaufläche. Die ersten beiden Molaren besaßen jeweils drei Leisten (trilophodont), während der letzte Molar vier oder fünf Leisten trug (vier im Oberkiefer, fünf im Unterkiefer). Die letzten beiden Milchprämolaren wiesen eine rechteckige Form und ebenfalls je drei Leisten auf, dagegen waren die beiden dauerhaften Prämolaren eher rundlich in ihrer Gestalt und verfügten nur über zwei Leisten.[4] Die Größe der Backenzähne nahm kontinuierlich zu, während der letzte Prämolar etwa 4 cm lang war, konnte der letzte Molar bis zu 23 cm erreichen.[1] Der bunodonte Aufbau der Backenzähne ist ein markanter Unterschied zu den heutigen Elefanten mit ihren typischen, durch Schmelzlamellen aufgebauten und dadurch eher flach wirkenden Zähnen. Andere, stammesgeschichtlich zumeist ältere Rüsseltiere wie die etwa die Mammutidae und Deinotheriidae, heben sich mit einem deutlich lophodonten bis zygodonten Bau der Backenzähne ab, das heißt, diese Tiere besaßen markant erhöhte Schmelzleisten, die den Zähnen eine dachartigen Aufbau verliehen.[5]

Stoßzähne
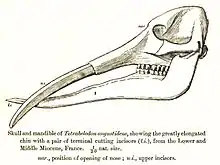
Gomphotherium besaß insgesamt vier Stoßzähne, also einen je Kieferast. Diese waren wie bei allen Rüsseltieren hypertrophierte Schneidezähne, die keine Wurzel ausgebildet hatten und ein Leben lang wuchsen. Im Gegensatz zu den Stoßzähnen der heutigen Elefanten, die weitgehend nur aus Zahnbein bestehen, war bei Gomphotherium ein schmales Band aus Zahnschmelz von 1 bis 3 mm Dicke ausgebildet, welches an den seitlichen Rändern des gesamten Zahns verlief.[6] Die oberen Stoßzähne wurden wie bei den heutigen Elefanten aus dem jeweils zweiten Schneidezahn (I2) gebildet und ragten in der Regel leicht gekrümmt nach unten. Der Querschnitt war an der Basis oval von etwa 13 mal 9 cm Größe, veränderte sich zu Spitze hin aber in eine runde Form, zudem verjüngten sich die Zähne kontinuierlich. Die Länge konnte bis zu 1,5 m betragen. Die Unterkieferstoßzähne dagegen wurden aus dem ersten Schneidezahn (I1) geformt[7] – lange Zeit hielt man analog zu den Stoßzähnen der oberen Zahnreihe ebenfalls den zweiten Schneidezahn als Ausgangszahn – und waren wesentlich kürzer und schlanker als die oberen. Auch diese hatten einen ovalen Querschnitt mit Durchmessern von 6,6 zu 4,8 cm an der Basis und verliefen mehr oder weniger gerade, sie verjüngten sich aber erst an der vordersten Spitze. Gelegentlich konnten sich die Enden der Unterkieferstoßzähne überlappen.[1]
Körperskelett
Das Körperskelett ist weitgehend vollständig bekannt. Die Wirbelsäule setzte sich aus 7 Hals-, 20 Brust-, 4 Lenden-, 3 Kreuzbein- und 21 Schwanzwirbel zusammen. Allein der erste Halswirbel, der Atlas war 48 cm breit und 22 cm hoch. Insgesamt zeigte die Wirbelsäule einen leicht gewölbten Verlauf, so dass die Rückenlinie von Gomphotherium wohl eher der des heutigen Asiatischen Elefanten entsprach, die größte Höhe des Tieres befand sich oberhalb der Vorderextremitäten. Die sehr hohen Dornfortsätze an den vorderen Brustwirbeln dienten als Ansatzstellen der kräftigen Nackenmuskulatur. Die Gliedmaßen standen senkrecht unter dem Körper und wirkten dadurch säulenartig. Die Gesamtlänge der vorderen Extremität betrug 219 cm, davon nahm der Oberarmknochen rund 100 cm ein, die Ulna 93 cm. Das Becken war wuchtig und maß in der Gesamtbreite über beide Schaufeln gemessen über 1,5 m. Den längsten Röhrenknochen stellte der Oberschenkelknochen mit 120 cm dar, das Schienbein erreichte 73 cm. Das Verhältnis der längeren Obergliedmaße zu den kürzeren Untergliedmaßen weist Gomphotherium als ein auf eine langsamere und schwerfälligere Fortbewegungsart angepasstes Tier aus, was typisch für die Rüsseltiere ist. Vorder- und Hinterfüße endeten in jeweils fünf Strahlen, die kurz und breit ausgebildet waren, der mittlere Metacarpus (Mc III) maß 22 cm, der mittlere Metatarsus (Mt III) 16 cm. Die gegenüber den Hinterfüßen größeren Vorderfüße geben an, dass die Hauptlast der Bewegung auf den vorderen Gliedmaßen lag.[1] Wie bei den heutigen Elefanten waren an Vorder- und Hinterfuß ein „sechster Zeh“ ausgebildet, ein eigentlich aus Knorpel bestehendes Gebilde, welches die Füße beim Stand und Lauf unterstützte und der sich an bestimmten Gelenkflächen der Hand- und Fußwurzelknochen ermitteln ließ. Diese Knorpelbildung entstand evolutionär mit der extremen Größenzunahme der Rüsseltiere und ist seit etwa dem Ende des Oligozän vor 30 Millionen Jahren bei verschiedenen Rüsseltiervertretern nachweisbar.[8]
Fossilfunde


Gomphotheriums ist aus weiten Bereichen Afrikas, Eurasiens und Nordamerikas bekannt. Aus Mitteleuropa liegen einige der bedeutendsten Fossilien vor, die weitgehend dem Molassebecken des Voralpenlandes entstammen. Hervorzuheben ist ein nahezu vollständiges und mehr als 170 Einzelknochen umfassendes Skelett, das 1971 in einer 50 cm mächtigen, graufarbenen Sandschicht am Ufer des Inns nahe der Ortschaft Gweng südwestlich von Mühldorf gefunden wurde. Es ist etwa 10 Millionen Jahre alt, lag am Fundort aber nicht mehr vollständig im Skelettverband. Nach seinem Fundort wird das Tier als „Mühldorfer Urelefant“ bezeichnet. Das Originalskelett wird in der Bayerischen Staatssammlung für Paläontologie und Geologie in München aufbewahrt, ein Abguss steht im Senckenbergmuseum. Bei dem Fund handelt es sich um ein männliches Individuum, das circa 50 Jahre alt war. Pathologisch-anatomische Veränderungen am rechten Hinterfuß deuten auf eine ernsthafte Verletzung des Tieres in seinen Jugendjahren hin. Das Fossil wurde systematisch in die Art Gomphotherium aff. steinheimense gestellt. Aufgrund der Größe des rekonstruierten Skelettes und des guten Erhaltungszustandes ist der Fund einer der wichtigsten überhaupt.[1] Bedeutend sind auch die Funde von Sandelzhausen, 70 km nördlich von München. Hier wurden über 200 Skelettreste, überwiegend Zähne und Stoßzähne, aber auch einige postcraniale Elemente, gefunden, die aber weitgehend nicht mehr im Skelettverband lagen.[9] Bei den meisten anderen Funden in Mitteleuropa handelt es sich häufig um Einzelstücke, wie aus der Umgebung von Ulm, wo mehrere Zahnfunde und Stoßzahnreste herstammen,[10] aber auch ein Stoßzahn aus Mettmach im Innviertel am Rande des Kobernaußerwaldes.
Auch außerhalb Mitteleuropas liegen sehr zahlreiche Funde vor, allerdings sind vollständige Skelettreste oder zusammengehörige Skelettelemente eher die Ausnahme; im Folgenden können nur beispielhafte Angaben gemacht werden. Allein von der Iberischen Halbinsel sind Fossilien von mehr als 125 Fundorten überliefert. Das Alter der Funde variiert von rund 17 bis 11 Millionen Jahren, herauszuheben sind hier ein vollständiger Schädel und Unterkiefer aus Cerámica Mirasierra in Madrid.[11] Herausragend sind weiterhin Funde aus En Pejouan nahe Simorre im Südwesten Frankreichs, wo aus dem Mitteleozän allein 10 Schädel und 18 Unterkiefer inklusive deren Fragmente unterschiedlichen Alters und Geschlechts aufgefunden wurden und als zu einer Population gehörig interpretiert werden.[12] In Ägypten kamen seit 1981 in mehreren Untersuchungen in der Moghara-Formation im Wadi Moghara im Norden des Landes Zahnfunde von Gomphotherium zum Vorschein, die wenigstens ein Dutzend Stücke umfassen. Mit einem Alter von 17 bis 18 Millionen Jahren sind diese ins Untere Miozän zu stellen und gehören möglicherweise mit zu den spätesten Nachweisen dieser Gattung auf dem afrikanischen Kontinent.[13] In China sind mehrere gute Fossilreste dokumentiert, so unter anderem ein vollständiger Schädel aus mitteleozänen Ablagerungen von Lantian in der Provinz Shaanxi und ein etwa gleichaltriger Fund eines Unterkiefers, dem die Stoßzähne fehlen aus Nanyucun in der Provinz Gansu.[14] Zu den jüngsten bisher nachgewiesenen Resten von Gomphotherium gehören Zahnfunde aus verschiedenen Steinbrüchen im zentralen Florida in Nordamerika. Sie entstammen den oberen Abschnitten der Bone-Valley-Formation und datieren ins beginnende Pliozän, dürften also rund 5 Millionen Jahre alt sein.[15] Generell war Gomphotherium in Nordamerika weit verbreitet und gehörte dort zu den häufigsten Rüsseltieren jener Zeit.[16]
Paläobiologie
Die aufgefundenen Fossilien von Gomphotherium zeigen einen gewissen Geschlechtsdimorphismus, der vor allem bei häufig nachgewiesenen Arten wie G. angustidens deutlicher herausgearbeitet ist als bei weniger häufigen Arten. Dieser spiegelt sich vor allem an Schädel und Unterkiefer wider und zeigt kräftigere und massivere Ausprägungen bei männlichen Tieren mit längeren Stoßzähnen als im Vergleich zu weiblichen. Auch am postcranialen Skelett sind solche Unterschiede zwischen den Geschlechtern erkennbar, die nur wenig von denen der heutigen Elefanten abweichen.[1] Da auch das Wachstum innerhalb der Jugendphase mit einem im Vergleich zum Afrikanischen Elefanten kaum abweichenden Zahndurchbruchsmuster relativ ähnlich erfolgte, beweist dies, dass der generelle Körperbau der Rüsseltiere evolutiv schon sehr früh in der Entwicklung dieser Säugetiergruppe festgelegt war und die späteren Veränderungen hauptsächlich über Variationen im Schädel- und Zahnbau stattfanden.[12]
Die niederkronigen Backenzähne mit ihren typisch buckligen (bunodonten) Kauflächenmuster sprechen dafür, dass das Tier sich hauptsächlich von weicher Pflanzenkost ernährte (browsing). Isotopenuntersuchungen an den Molaren ergaben aber ein etwas abweichendes Muster, dass auf eine Ernährung durch gemischte Pflanzenkost verweist. Allerdings scheint es gewisse regionale Unterschiede in der Ernährung gegeben zu haben, da einige untersuchte Individuen, die aus dem heutigen Texas stammen, einen höheren Konsum an Gräsern annehmen lassen. Diese Variationen werden jedoch als Unterschiede im Nahrungsangebot der einzelnen Regionen angesehen. Geschlossen wird dies aus der Tatsache, dass alle untersuchten Tiere unterschiedlichen geologischen Epochen vom Unteren bis zum Oberen Miozän angehören. Während dieser Zeit kam es zwar zu gravierenden naturräumlichen Umwandlungen hin zu offeneren Landschaften mit trockenerem bis kühlerem und stärker jahreszeitlich gegliedertem Klima, doch zeigte keines der untersuchten Tiere eine generelle Veränderung in der Bevorzugung bestimmter Pflanzen. Auch scheint es innerhalb der Wachstumsphase der Jungtiere nicht zu einer Umstellung auf bestimmte Nahrungspflanzen gekommen zu sein.[17][18] Diese Analysen sprechen aber auch dafür, dass sich Gomphotherium bevorzugt in offenen, teils trockenen Graslandschaften und Parkwäldern aufhielt und dichte, geschlossene Wälder eher mied.[19] Allerdings erbrachten wiederum Untersuchungen von Phytolithen an Fossilfunden aus dem heutigen Ostasien, dass zumindest einzelne Vertreter wie G. steinheimense bereits im Mittleren Miozän fast ausschließlich nur Gräser fraßen. Diese gehören damit zu den frühesten Nachweisen von Rüsseltieren mit einer derartig spezialisierten Ernährungsweise, da sie bereits rund 10 Millionen Jahre vor dem ersten Erscheinen der Elefanten auftraten.[20]
Dass Gomphotherium zu einer Zeit lebte, in der sich dramatische Änderungen des Klimas vollzogen, konnte auch anhand des Wachstums der Stoßzähne festgestellt werden. Da diese über das gesamte Leben eines Tieres an Länge zunahmen, zeigen sich bestimmte Wachstumsraten in Form dunklerer und hellerer Einlagerungen im Zahnbein, deren jeweilige Breite direkt mit dem Nahrungsangebot korreliert. Dabei bedeuten die helleren Bereiche Phasen intensiven, die dunkleren solche weniger intensiven Wachstums. Untersuchte Stoßzahnreste aus dem Mittleren Miozän wiesen ein eher irreguläres Wachstumsmuster auf, das erkennen lässt, dass es nur verschiedentlich zu Nahrungsengpässen kam, die nicht jahreszeitlich gebunden waren. Spätmiozäne Funde dagegen besaßen ein sehr regelmäßiges Wachstumsmuster mit abwechselnd breiten hellen und schmalen dunklen Streifen. Dies wird mit stärkeren jahreszeitlich bedingten klimatischen Schwankungen und damit einhergehenden variierenden Nahrungsressourcen in Verbindung gebracht.[6]
Stammesgeschichte
Der Ursprung von Gomphotherium liegt in Afrika, die ältesten Funde sind aus dem Oligozän von der Fundstelle Chilga im nördlichen Äthiopien bekannt und werden auf rund 28 Millionen Jahre datiert, allerdings umfassen die Funde nur einzelne Zähne.[21] Die Ausbildung von vier Stoßzähnen ist auch vom älteren Phiomia bekannt, als dessen Abkömmling er manchmal angesehen wird. In Eurasien erschien die Gattung erstmals im Unteren Miozän nach der Entstehung einer Landbrücke durch die Schließung der Tethys, das Auftreten von Rüsseltieren außerhalb Afrikas wird auch als Proboscidea datum event bezeichnet. Die ältesten Funde hier stammen aus der Kamlial-Formation der Bugti-Berge vom Indischen Subkontinent und datieren älter als 18,3 Millionen Jahre,[22] in Europa ist Gomphotherium erstmals vor rund 18 Millionen Jahren belegt.[5][14] Spätestens im mittleren Miozän vor 16,5 Millionen Jahren erreichte es auch Nordamerika, nach Südamerika, das damals noch nicht über den Isthmus von Panama mit dem nördlichen Kontinent verbunden war, drang es aber nicht mehr vor. Die südlichsten Fundstellen in Amerika liegen im heutigen Costa Rica.[23]
In Eurasien wurde Gomphotherium bereits im ausgehenden Mittel- und im beginnenden Obermiozän durch fortschrittlichere Rüsseltiere wie Tetralophodon ersetzt, einige Forscher sehen in dieser Rüsseltierform auch den direkten Nachfolger von Gomphotherium. Der Austauschprozess begann vor etwa 11 bis 12 Millionen Jahren. Zu den jüngsten Funden in Europa gehören jene von Gweng bei München mit einem Alter von rund 10 Millionen Jahren. In Nordamerika hielt sich der Rüsseltiervertreter noch bis ins Pliozän vor vier Millionen Jahren.[5][14]
Systematik
Innere Systematik der Gomphotheriidae nach Cozzuol et al. 2012[24]
|
Gomphotherium ist eine Gattung aus der Ordnung der Rüsseltiere (Proboscidea) und wird innerhalb dieser zur Familie der Gomphotheriidae gestellt, die wiederum zur Überfamilie Gomphotherioidea gehört. Die Gomphotherien umfassen entwickelte Rüsseltiere, die wie die heutigen Elefanten über das Merkmal des horizontalen Zahnwechsels verfügten, im Gegensatz zu den frühesten Vertretern der Ordnung, die noch einen säugetiertypischen vertikalen Zahnwechsel aufwiesen. Der horizontale Zahnwechsel entstand durch die Vergrößerung der Backenzähne – so ist im Laufe der Rüsseltierevolution eine Zunahme der Anzahl der Schmelzleisten von zwei auf sechs bis sieben je Molar zu verzeichnen – bei gleichzeitiger Verkürzung des Kiefers, so dass nicht mehr alle Zähne gleichzeitig Platz fanden. Erstmals nachgewiesen ist er mit der Gattung Eritreum vor rund 28 Millionen Jahren. Alle Rüsseltiere mit dem Merkmal des horizontalen Zahnwechsels werden in die Unterordnung der Elephantimorpha gestellt.[25] Innerhalb der Gomphotheriidae ist eine Zuweisung zur Unterfamilie der Gomphotheriinae üblich, die nächstverwandte Gattung stellt Serridentinus dar, die relativ ähnlich ist und lange Zeit mit Gomphotherium synonymisiert war.[26]
Insgesamt wurden im Laufe der Forschungsgeschichte mehr als 75 unterschiedliche Arten von Gomphotherium beschrieben, von denen ein Großteil allerdings Synonyme von einerseits anderen Arten von Gomphotherium, andererseits von anderen Rüsseltiergattungen darstellen, weitere werden als nomina nuda angesehen. Die genaue Anzahl der Arten lässt sich derzeit nicht bestimmen, da vor allem die afrikanischen und ostasiatischen Vertreter bisher keiner genaueren Revision unterlagen. Aufgeführt werden hier nur die als sicher zu Gomphotherium zu stellenden Arten:[1][27][14][28]
Innere Systematik der Gattung Gomphotherium nach Wang et al. 2017[28]
|
- G. angustidens Cuvier, 1817
- G. annectens Matsumoto, 1925
- G. browni Osborn, 1926
- G. connexum Hopwood, 1935
- G. cooperi Osborn, 1926
- G. hannibali Welcomme, 1994
- G. inopinatum Schlesinger, 1928
- G. lybicum Fourtau, 1918
- G. mongoliensis Osborn, 1924
- G. pasalarense Gaziry, 1987
- G. productum Cope, 1875
- G. pygmaeus Depéret, 1897
- G. republicanum Osborn, 1926
- G. shensiense Chang & Zhai, 1978
- G. simplicidens Osborn, 1923
- G. steinheimense Klähn, 1922
- G. subtapiroideum Borissiak & Beljaeva, 1917
- G. sylvaticum Tassy, 1985
- G. tassyi Wang, Li, Duangkrayom, Yang, He & Chen, 2017
- G. wimani Hopwood, 1935
Innerhalb von Gomphotherium wurden zwei größere Formenkreise unterschieden. Dabei umfasst die G. annectens-Gruppe morphologisch archaische Formen wie die namengebende Art oder G. cooperi aus Südasien und wird überwiegend in das Untere Miozän gestellt. Diese besitzen meist einfache bunodonte Zähne mit nur einer kleinen vierten Leiste am hintersten Molar. Eher moderne Formen werden in der G. angustidens-Gruppe zusammengefasst, die hauptsächlich in Eurasien auftritt und meist auch jünger datiert.[13][22] Insgesamt gibt es aufgrund der fehlenden Revisionen der Arten, aber auch aufgrund des häufig nur stark fragmentiert vorliegenden Fundmaterials noch viele Unterscheidungsschwierigkeiten sowohl zwischen den einzelnen Arten als auch zu nahe verwandten Gattungen hin. Dies zeigt sich auch an zahlreichen synonym geführten Gattungsnamen, so etwa Genomastodon, Ocalientinus, Tetrabelodon, Trilophodon, Trobelodon, Tatabelodon oder Hemilophodon.[1]
Forschungsgeschichte
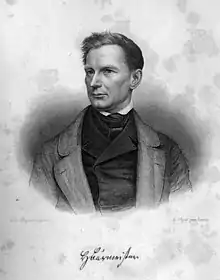
Die ersten heute zu Gomphotherium gestellten Fossilien wurden 1817 von Georges Cuvier als Mastodon angustidens bezeichnet, allerdings hatte er bereits 1806 den Begriff mastodonte à dents étroites geprägt.[29][30][1] Den Gattungsnamen Gomphotherium führte Hermann Burmeister im Jahr 1837 ein, wobei er in seiner kurzen Beschreibung „Stoßzähne in beiden Kiefern besaß die gleichfalls untergegangene Gatt. Gomphotherium.“ dieses eindeutig von den „Mastodonten“ abhob, welchen er zusätzlich den Namen „Ohiothier“ gab (gemeint war damit das Amerikanische Mastodon mit der heutigen wissenschaftlichen Bezeichnung Mammut americanum).[31] Der Name „Mastodon“ diente später als Bezeichnung für eine ganze Gruppe von Rüsseltieren, die durch ihre urtümlichere Morphologie von den heutigen Elefanten abwichen. Henry Fairfield Osborn (1857–1935) wies diesen stammesgeschichtlich älteren Formen im Jahr 1921 als „Mastodontoidea“ den Status einer Überfamilie zu.[32][33] Dieser Überfamilie, in der Regel als „Mastodonten“ bezeichnet, wurden mehrere Familien zugerechnet, wie die Mammutiden (ursprünglich als „Echte Mastodonten“ bezeichnet), die Gomphotherien und die Stegodonten. Aufgrund abweichender Zahnstrukturen unterschied man in der Regel die Formenkreise der „bunodonten“ (Gomphotherien) und der „zygodonten Mastodonten“ (Mammutiden), wobei innerhalb der bunodonten Formen eine Entwicklung von einer trilophodonten (die eigentlichen Gomphotherien) zu einer tetralophodonten Gruppe (Gattungen wie Anancus oder Tetralophodon, heute zu den Elephantoidea gestellt) erkannt wurde.[34] Die Bezeichnungen bezogen sich dabei auf die Anzahl der Paare an Zahnschmelzhöckern auf den ersten beiden Molaren. Allerdings erwies sich die Überfamilie „Mastodontoidea“ einerseits als polyphyletisch – so stammen die Mammutiden höchstwahrscheinlich von Palaeomastodon, die Gomphotherien und deren Nachfolger aber von Phiomia ab[25] –, andererseits wurde der Begriff „Mastodon“ zunehmend als synonym zur Gattung Mammut gesehen, welche bereits Johann Friedrich Blumenbach im Jahr 1799 anhand nordamerikanischer Fossilien als Mammut ohioticum[35] (heute mit Mammut americanum bezeichnet, das Amerikanische Mastodon) eingeführt hatte. Die tri- und tetralophodonten Gomphotherien und die Stegodonten wurden deshalb in den 1980er und 1990er Jahren in die modernere Überfamilie der Elephantoidea eingegliedert, der auch die heutigen Rüsseltiere angehören, die Mammutiden dagegen zur Überfamilie der Mammutoidea.[36] Der Begriff „Mastodon“ wird heute nicht mehr verwendet, mit Ausnahme als Teil eines Gattungsnamens beziehungsweise umgangssprachlich für das Amerikanische Mastodon.[37] Erst nach dem Jahr 2000 erhielten die trilophodonten und damit eigentlichen Gomphotherien einschließlich Gomphotherium einen Platz in der Überfamilie Gomphotherioidea, welche zwar bereits 1922 von Oliver Perry Hay etabliert, jedoch als Ordnungsgruppe nur selten benutzt worden war.[26]
Einzelnachweise
- Ursula B. Göhlich: Elephantoidea (Proboscidea, Mammalia) aus dem Mittel- und Obermiozän der Oberen Süßwassermolasse Süddeutschlands: Odontologie und Osteologie. Münchner Geowissenschaftliche Abhandlungen Reihe A 36, München, 1998
- Eva-Maria Natzer: Fossil des Jahres 2013: Gomphotherium von Gweng () auf der Webseite der Bayerischen Staatssammlung für Paläontologie und Geologie in München; auch Gertrud Rößner: Fossil des Jahres 2013: Gomphotherium von Gweng () auf der Webseite der Fakultät für Geowissenschaften der Ludwig-Maximilians-Universität München; beide zuletzt abgerufen am 14. April 2019
- Per Christiansen: Body size in proboscideans, with notes on elephant Metabolism. Zoological Journal of the Linnean Society 140, 2004, S. 523–549
- William J. Sanders: Horizontal tooth displacement and premolar occurrence in elephants and other elephantiform proboscideans. Historical Biology, 2018 doi:10.1080/08912963.2017.1297436
- Jan van der Made: The evolution of the elephants and their relatives in the context of a changing climate and geography. In: Harald Meller (Hrsg.): Elefantenreich - Eine Fossilwelt in Europa. Halle/Saale, 2010, S. 340–360
- David L. Fox: Growth increments in Gomphotherium tusks and implications for late Miocene climate change in North America. Palaeogeography, Palaeoclimatology, Palaeoecology 156, 2000, S. 327–348
- Cyrille Delmer: Reassessment of the generic attribution of Numidotherium savagei and the homologies of lower incisors in proboscideans. Acta Palaeontologica Polonica 54 (4), 2009, S. 561–580
- John R. Hutchinson, Cyrille Delmer, Charlotte E. Miller, Thomas Hildebrandt, Andrew A. Pitsillides und Alan Boyde: From Flat Foot to Fat Foot: Structure, Ontogeny, Function, and Evolution of Elephant “Sixth Toes”. Science 234, 2011, S. 1699–1703
- Ursula B. Göhlich: The Proboscidea (Mammalia) from the Miocene of Sandelzhausen (southern Germany). Paläontologische Zeitschrift 84, 2010, S. 163–204
- Volker J. Sach und Elmar P. J. Heizmann: Stratigraphie und Säugetierfaunen der Brackwassermolasse in der Umgebung von Ulm (Südwestdeutschland). Stuttgarter Beiträge zur Naturkunde Serie B 310, 2001, S. 1–95
- A. V. Mazo und Jan Van der Made: Iberian mastodonts: Geographic and stratigraphic distribution. Quaternary International 255, 2012, S. 239–256
- Pascal Tassy: Growth and sexual dimorhism among Miocene elephantoids: the example of Gomphotherium angustidens. In: Jeheskel Shoshani und Pascal Tassy (Hrsg.): The Proboscidea. Evolution and palaeoecology of the Elephants and their relatives. Oxford, New York, Tokyo, 1996, S. 92–100
- William J. Sanders und Ellen R. Miller: New Proboscideans from the Early Miocene of Wadi Moghara, Egypt. Journal of Vertebrate Paleontology 22 (2), 2002, S. 388–404
- Wang Shi-Qi, Liu Shan-Pin, Xie Guang-Pu, Liu Jia, Peng Ting-Jiang und Hou Su-Kuan: Gomphotherium wimani from Wushan County, China, and its implications for the Miocene stratigraphy of the Tianshui area. Vertebrata Palasiatica 51 (1), 2013, S. 71–84
- S. David Webb, Richard C. Hulburt, Jr., Gary S. Morgan und Helen F. Evans: Terrestrial mammals of the Palmetto Fauna (early Pliocene, latest Hemphillian) from the Central Florida Phosphate District. Science Series 41, 2008, S. 293–312
- W. David Lambert: The biogeography of gomphotheriid proboscidean of North America. In: Jeheskel Shoshani und Pascal Tassy (Hrsg.): The Proboscidea. Evolution and palaeoecology of the Elephants and their relatives. Oxford, New York, Tokyo, 1996, S. 143–148
- David L. Fox und Daniel C. Fisher: Dietary reconstruction of Miocene Gomphotherium (Mammalia, Proboscidea) from the Great Plains region, USA, based on the carbon isotope composition of tusk and molar enamel. Palaeogeography, Palaeoclimatology, Palaeoecology 206, 2004, S. 311–335
- Ivan Calandra, Ursula B. Göhlich und Gildas Merceron: Feeding preferences of Gomphotherium subtapiroideum (Proboscidea, Mammalia) from the Miocene of Sandelzhausen (Northern Alpine Foreland Basin, southern Germany) through life and geological time: evidence from dental microwear analysis. Paläontologische Zeitschrift 84, 2010, S. 205–215
- Mark T. Clementz. New insight from old bones: stable isotope analysis of fossil mammals. Journal of Mammalogy, 93 (2), 2012, S. 368–380
- Yan Wu, Tao Deng, Yaowu Hu, Jiao Ma, Xinying Zhou, Limi Mao, Hanweng Zhang, Jie Ye und Shi-Qi Wang: A grazing gomphothere in the Middle Miocene Central Asia, 10 million years prior to the origin of Elephantidae. Scientific Reports 8, 2018, S. 7640, doi:10.1038/s41598-018-25909
- William Sanders, John Kappelmann und D. Tab Rassmussen: New large-bodied mammals from the late Oligocene site of Chilga, Ethiopia. Acta Palaeontologica Polonica 49 (3), 2004, S. 365–392
- Pascal Tassy: The earliest Gomphotheres. In: Jeheskel Shoshani und Pascal Tassy (Hrsg.): The Proboscidea. Evolution and palaeoecology of the Elephants and their relatives. Oxford, New York, Tokyo, 1996, S. 89–91
- Spencer Georges Lucas und Guillermo E. Alvarado: Fossil Proboscidea from the Upper Cenozoic of Central America: taxonomy, evolutionary and palaeobiogeographic significance. Revista Geológica de America Central 42, 2010, S. 9–42
- Mario A. Cozzuol, Dimila Mothé und Leonardo S. Avilla: A critical appraisal of the phylogenetic proposals for the South American Gomphotheriidae (Proboscidea: Mammalia). Quaternary International 255, 2012, S. 36–41
- Jeheskel Shoshani, Robert C. Walter, Michael Abraha, Seife Berhe, Pascal Tassy, William J. Sander, Gary H. Marchant, Yosief Libsekal, Tesfalidet Ghirmai und Dietmar Zinner: A proboscidean from the late Oligocene of Eritrea, a ‘‘missing link’’ between early Elephantiformes and Elephantimorpha, and biogeographic implications. PNAS 103 (46), 2006, S. 17296–17301
- Jeheskel Shoshani und Pascal Tassy: Advances in proboscidean taxonomy & classification, anatomy & physiology, and ecology & behavior. Quaternary International 126-128, 2005, S. 5–20
- William J. Sanders, Emmanuel Gheerbrant, John M. Harris, Haruo Saegusa und Cyrille Delmer: Proboscidea. In: Lars Werdelin und William Joseph Sanders (Hrsg.): Cenozoic Mammals of Africa. University of California Press, Berkeley, London, New York, 2010, S. 161–251
- Shi-Qi Wang, Yu Li, Jaroon Duangkrayom, Xiang-Wen Yang, Wen He und Shan-Qin Chen: A new species of Gomphotherium (Proboscidea, Mammalia) from China and the evolution of Gomphotherium in Eurasia. Journal of Vertebrate Paleontology, 2017, S. e1318284 doi: 10.1080/02724634.2017.1318284
- Georges Cuvier: Sur différentes dents du genre des mastodontes, mais d'espèces moindres que celles de l'Ohio, trouvées en plusieurs lieux des deux continens. Annales du Muséum d'histoire naturelle 8, 1806, S. 401–424 ()
- Georges Cuvier: Le règne animal distribué d'après son organisation: pour servir de base a l'histoire naturelle des animaux et d'introduction a l'anatomie comparée. Paris, 1817, S. 1–540 (S. 232–233) ()
- Hermann Burmeister: Handbuch der Naturgeschichte. Zum Gebrauch bei Vorlesungen. Berlin, Enslin, 1837, S. 1–858 (S. 795)
- Henry Fairfield Osborn: The Evolution, Phylogeny, and Classification of the Mastodontoidea. Bulletin of the Geological Society of America 32 (3), 1921, S. 327–332
- Henry Fairfield Osborn: Adaptive radiation and classification of the Proboscidea. PNAS 7, 1921, S. 231–234
- Jeffrey J. Saunders: North American Mammutidae. In: In: Jeheskel Shoshani und Pascal Tassy (Hrsg.): The Proboscidea. Evolution and palaeoecology of the Elephants and their relatives. Oxford, New York, Tokyo, 1996, S. 271–279
- Johann Friedrich Blumenbach: Handbuch der Naturgeschichte. Göttingen, 1799 (S. 697–698)
- Jeheskel Shoshani: Understanding proboscidean evolution: a formidable task. Tree 13, 1998, S. 480–487
- Pascal Tassy: Who is who among the Proboscidea? In: Jeheskel Shoshani und Pascal Tassy (Hrsg.): The Proboscidea. Evolution and palaeoecology of the Elephants and their relatives. Oxford, New York, Tokyo, 1996, S. 40–48
Weblinks
- The Paleobiology Database Gomphotherium