Fossil
Ein Fossil (lateinisch fossilis ‚ausgegraben‘) ist jedes Zeugnis vergangenen Lebens der Erdgeschichte, das älter als 10.000 Jahre ist und sich somit einem geologischen Zeitalter vor dem Beginn des Holozäns zuordnen lässt.[1] Derartige erdgeschichtliche Dokumente können sowohl körperliche Überreste von Lebewesen (Körperfossilien) sein als auch Zeugnisse ihrer Aktivität (Spurenfossilien). Zum Beispiel werden auch versteinerte Trittsiegel und Exkremente (Koprolithe) zu den Fossilien gezählt. Die Entstehung von Fossilien nennt man Fossilisation. Die Erforschung der Fossilien erfolgt in erster Linie durch die Paläontologie.

Die Bezeichnung „Versteinerung“ oder veraltet „Petrefakt“ (lateinisch petra [von altgriechisch πέτρα] Stein, factum „gemacht“)[2] ist nicht gleichbedeutend, denn nicht jedes Fossil ist mineralisiert und liegt somit als eine Versteinerung vor.
Wissenschaftlich fundierten Schätzungen zufolge sind etwa eine Milliarde Tier- und Pflanzenarten seit dem Beginn des Phanerozoikums vor 541 Millionen Jahren entstanden und größtenteils wieder ausgestorben. Manche Schätzungen belaufen sich sogar auf 1,6 Milliarden. Weit unter ein Prozent dieses Artenreichtums ist fossil erhalten geblieben, nicht nur wegen der besonderen Umstände, die für die Fossilisation erforderlich sind, sondern auch, weil viele Fossilien im Laufe der Jahrmillionen infolge von Verwitterung, Erosion oder Gesteinsmetamorphose wieder zerstört wurden. Bis 1999 sind rund 250.000 fossile Arten wissenschaftlich beschrieben worden.[3]
Historisches

Funde von in Gesteinen enthaltenen marinen Lebewesen auf dem Festland erregten bereits im Altertum und im Mittelalter von China bis Europa Aufmerksamkeit, und sie wurden von mehreren Gelehrten (u. a. Xenophanes, Eratosthenes, Leonardo da Vinci) richtig als Überreste von Organismen bzw. als Hinterlassenschaften einer einstigen Meeresbedeckung gedeutet. Hingegen wurden sie vom altgriechischen Philosophen Aristoteles und den auf seinen Lehren aufbauenden Scholastikern als Launen der Natur (Lusus naturae) betrachtet.[5][6]
Die Bezeichnung Fossil wurde erstmals 1546 von Georgius Agricola in seinem Werk De natura fossilium verwendet. Benannte man, wie Agricola es tat, zunächst unterschiedslos alle „beim Ausgraben“ gefundenen Kuriositäten als Fossil, darunter auch Minerale, Artefakte, seltsam geformte Wurzeln oder Konkretionen, fand im Laufe der Zeit eine Bedeutungsverengung auf Objekte statt, die von der Existenz urzeitlichen Lebens zeugten. Maßgeblich trug dazu der Franzose Jean-Baptiste Lamarck mit dem Kapitel Sur les fossiles in seinem umfassenden Werk zur Systematik der wirbellosen Tiere bei, das 1801 erschien.[4][7] Gängige Bezeichnungen für Fossilien waren auch Petrefakt und Versteinerung, und tatsächlich entstehen in vielen Fällen Fossilien dadurch, dass organische durch mineralische Substanz ersetzt wird. Entsprechend wurde die Vorform der modernen Paläontologie bis weit ins 19. Jahrhundert hinein Petrefaktenkunde genannt. Der Däne Nicolaus Steno führte bereits 1667 als erster neuzeitlicher Gelehrter den Nachweis, dass es sich bei Fossilien nicht um Launen der Natur, sondern um Reste von Lebewesen aus früherer Zeit handelt.
Substantiv Fossil vs. Adjektiv fossil
Neben der Bezeichnung Fossil als Substantiv wird das Wort auch als Adjektiv verwendet, um damit Objekte oder Bildungen zu charakterisieren, die geologisch relativ alt sind. In diesem Sinne wird das Adjektiv fossil auch heute noch nicht ausschließlich für Überreste von Lebewesen verwendet, beispielsweise in der Wortkombination fossile Energieträger (Kohle, Erdöl, Erdgas) oder fossiles Wasser (sehr altes Tiefengrundwasser). Das Gegenteil zum Adjektiv fossil lautet rezent. Es bezeichnet alle Bildungen, die geologisch relativ jung sind. Geologische Zeugnisse, deren Alter im Grenzbereich zwischen fossil und rezent liegt, können als subfossil oder subrezent bezeichnet werden. Dies betrifft Bildungen aus der Zeit des Quartärs bis etwa 8000 v. Chr. (siehe auch → Prähistorie).[8] In der Paläobotanik bezieht sich subfossil aber auch oft auf eine unvollständige Fossilisation, also einen bestimmten Erhaltungszustand. In diesem Sinne können selbst Floren aus dem Paläogen (älter als 25 Millionen Jahre) noch als subfossil gelten.
In der Biologie und Paläontologie wird das Adjektiv fossil auch auf ausgestorbene Arten bezogen. Als „fossile Arten“ werden jene Arten bezeichnet, die vor der Wende vom Pleistozän zum Holozän ausstarben, also vor mehr als rund 12.000 Jahren. Den fossilen Arten können die rezenten Arten gegenübergestellt werden, die erst im Holozän ausstarben oder heute noch existieren. Mit den sogenannten lebenden Fossilien sind ebenfalls (rezente) Arten gemeint, keine Einzelexemplare. Die Gesamtheit aller aktuell der Wissenschaft bekannten fossilen Arten sowie deren Vorkommen wird Fossilbericht oder auch Fossilüberlieferung (englisch fossil record) genannt.
In der Geomorphologie werden Oberflächenformen als fossil bezeichnet, die zwar heute noch existieren, deren Bildungsprozess aber in der geologischen Vergangenheit zum Erliegen kam (sogenannte inaktive Strukturen). Dies betrifft auch Bildungen, die deutlich jünger sind als 10.000 Jahre, beispielsweise inaktive holozäne Kliffs in Anlandungsbereichen der deutschen Ostseeküste.[9]
Einteilung
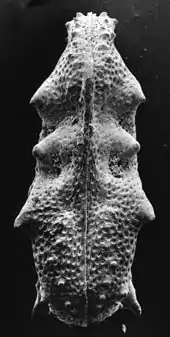





Körperfossilien
Als Körperfossilien werden Fossilien bezeichnet, die unmittelbar auf ganze Körper oder einzelne Körperteile von toten Lebewesen zurückgehen. Sie werden nach ihrer Größe weiter differenziert in
- Makrofossilien, die mit bloßem Auge sichtbar sind und bisweilen Hausgröße erreichen (in aller Regel sind Makrofossilien gemeint, wenn allgemein von „Fossilien“ die Rede ist),
- Mesofossilien, die nur mit Hilfe einer Lupe sichtbar sind (Bezeichnung wird selten angewendet),
- Mikrofossilien, die nur mit Hilfe eines Lichtmikroskops sichtbar sind (oft werden hier die Mesofossilien mit eingeordnet) und
- Nannofossilien, die nur mit Hilfe eines Elektronenmikroskops sichtbar sind.
Der Wortteil „Körper-“ steht bei Körperfossilien nicht zwangsläufig für das originale Körpergewebe, sondern die Überlieferung erfolgt oft durch Ersetzung von Teilen des Gewebes oder des gesamten Gewebes durch Minerale oder Sediment. Die Erhaltung unveränderter Körpersubstanz sowie die teilweise oder vollständige Ersetzung von Körpersubstanz durch Minerale wird auch Originalerhaltung genannt. Dies betrifft bei Tieren meistens nicht die Weichteile, sondern nur die mineralischen Hartteile (Hartteilskelette). Da viele Wirbellose Außenskelette haben, unterscheiden sich deren Fossilien im Aussehen oft nicht besonders stark vom lebenden Tier. Beispiele sind Mollusken mit Gehäusen aus Calcit oder känozoische Mollusken mit Aragonitgehäuse. Bei Wirbeltieren mit ihren Innenskeletten unterscheidet sich das Fossil stärker vom lebenden Tier. Ausnahmen bilden die paläozoischen Knochenfische mit ihren schon zu Lebzeiten stark mineralisierten Schuppen. Weichteile haben generell ein geringes Erhaltungspotenzial, und je älter eine Sedimentschicht ist, desto unwahrscheinlicher ist es, dass darin Weichteile überliefert sind. Daher ist beispielsweise die großflächige Originalerhaltung von Dinosaurierhaut in kreidezeitlichen Sedimenten äußerst selten.[10] Auch einige organische Substanzen haben ein relativ hohes Erhaltungspotenzial, zum Beispiel das Chitin der Arthropoden oder das Sporopollenin der Pollen und Pflanzensporen. Pflanzen werden oft inkohlt überliefert, das heißt, ihr Gewebe wird unter Luftabschluss entwässert und chemisch umgewandelt, so dass sich Kohlenstoff darin anreichert. Eine relativ seltene Form der Originalerhaltung ist der Einschluss in Bernstein. In dieser Form sind neben Pflanzenteilen vor allem kleine, wirbellose Landtiere, meistens Insekten, überliefert.[11]
Während als solche erkennbare Meso-, Mikro- und Nannofossilien in aller Regel in Originalerhaltung auftreten, unterscheidet man bei Makrofossilien verschiedene weitere Erhaltungsformen:
- Steinkern (engl. internal mould): Dieser Spezialfall eines Körperfossils ist typisch für wirbellose Tiere mit Außenskelett („Gehäuse“) aus in geologischen Zeiträumen instabilen Mineralen. Nach Ableben des Tieres und seiner Einbettung im Sediment zerfällt die organische Substanz im Inneren des Gehäuses, und ein Hohlraum bildet sich. In diesen sowie in eventuell schon vor dem Tod im Gehäuse vorhandene Hohlräume dringt nachfolgend das umgebende Sediment ein und füllt sie ganz oder teilweise aus. Wird das Außenskelett im Laufe der folgenden Millionen Jahre weggelöst, bleibt die Ausfüllung des Gehäuses erhalten und wird als Steinkern überliefert. Steinkerne sind typisch für prä-känozoische Mollusken mit aragonitischem Gehäuse, speziell Ammoniten, Schnecken und bestimmte Muscheln. Bei der Steinkernüberlieferung treten zwei Spezialformen auf:
- Beim Prägesteinkern wird die äußere Skulptur des Gehäuses, die im umschließenden Sediment einen Negativabdruck erzeugt, nach Weglösung des Gehäuses auf den Steinkern aufgeprägt. Der Prägesteinkern ist eine typische Überlieferungsform bei Ammoniten und bestimmten prä-känozoischen Muscheln.[12]
- Der Marksteinkern ist eine Überlieferungsform der Sprossachsen („Stängel“) schwach verholzter Pflanzen: Das Gewebe im Inneren der Sprossachsen, das sogenannte Mark, zerfällt nach dem Tod der Pflanze deutlich schneller als die äußere „Rinde“. Dadurch entsteht die sogenannte Markhöhle. Wird die Markhöhle mit Sediment verfüllt, bleibt nach dem Zerfall der „Rinde“ die Hohlraumfüllung übrig und kann als Marksteinkern überliefert werden. Marksteinkerne sind typische Erhaltungsformen der Stämme von Riesenschachtelhalmen (Calamiten).
- Abdruck (engl. impression, external mould): Abdrücke sind die Negativformen von Körperfossilien in Originalerhaltung und von Steinkernen. Bisweilen wird jedoch die originale Skelettsubstanz des eingebetteten Organismus im Laufe der Diagenese ohne vorherige Steinkernbildung komplett aus dem Gestein weggelöst, so dass nur ein Abdruck zurückbleibt. Insbesondere hierfür prädestiniert sind Wirbeltiere, weil sie aufgrund ihrer Innenskelette nur in Ausnahmefällen als Steinkerne erhalten werden können. Derartige Abdruckerhaltung kommt u. a. bei Mesosaurus-Skeletten in der Whitehill-Formation in Südafrika sowie bei Landwirbeltierskeletten im Mittleren Buntsandstein und in Kännelkohle des Oberkarbons von Nordamerika vor. Das Studium der Anatomie dergestalt erhaltener Tiere erfolgt in solchen Fällen nicht am Abdruck, sondern dieser wird mit Latex oder Silikon (früher Guttapercha) ausgegossen, und erst dieser Abguss wird dann untersucht und beschrieben.
Spurenfossilien

Als Spurenfossilien oder Lebensspuren gelten alle Zeugnisse von Lebewesen, die nicht auf deren tote Körper zurückzuführen sind, vor allem solche Hinterlassenschaften, die durch die Tätigkeit von Lebewesen erzeugt wurden. Dazu zählen beispielsweise fossil erhaltene Trittsiegel und Fährten, Grabgänge im Sediment (siehe auch → Bioturbation), Bohrspuren in fossilem Holz oder in Kalkstein sowie Fraß- und Weidespuren (z. B. Bissmarken an fossilen Knochen). Eine spezielle Form der Lebensspuren sind fossile Eierschalen in fossilen Gelegen. Auch Verdauungsrückstände zählen dazu, beispielsweise die Kotpillen von kleinen Fischen und Wirbellosen, die mit bloßem Auge im Gestein oft gar nicht sichtbar sind. Größere „Kotsteine“, sogenannte Koprolithen, sowie fossile Gewölle und Speiballen (letztgenannte oft nur in quartären Höhlensedimenten erhalten) stellen ein Bindeglied zu den Körperfossilien dar, weil sie nicht selten Reste des Originalmaterials der Beutetiere der Koprolith- und Gewöllproduzenten enthalten.
Bedeutung
Dokumente der Stammesgeschichte
Die enorme Anzahl und Vielfalt der im Fossilbericht enthaltenen Arten ist eines der wichtigsten Argumente für die Evolutionstheorie. Sie zeigt, dass im Laufe der Geschichte des Lebens unzählige Formen auf der Erde entstanden und wieder verschwunden sind. Diese ehemalige Vielfalt ist aus der Existenz heutiger Formen allein nicht ableitbar. So finden sich im Fossilbericht nicht nur relativ enge Verwandte von in der Gegenwart lebenden Organismen, sondern auch einst blühende, aber nachkommenslos erloschene Tier- und Pflanzengruppen (siehe auch Evolutionsgeschichte). Des Weiteren können in Kombination mit sedimentologischen Untersuchungen, insbesondere durch die Bestimmung des Ablagerungsmilieus fossilführender Schichten, evolutive Entwicklungen über ökologische Grenzen hinweg, wie der Landgang verschiedener Organismengruppen (höhere Pflanzen, Gliederfüßer, Wirbeltiere), nachvollzogen werden.
Zeugen vergangener Lebensräume
Fossilien dienen als Hinweise auf ehemalige geographische und ökologische Verhältnisse. Beispielsweise gelten einige fossile Arten und Ichnospezies als typisch für ein ganz bestimmtes Ablagerungsmilieu. Sie werden daher Zeigerfossilien oder Faziesanzeiger genannt. Zudem können bestimmte Assoziationen von Spurenfossilien mit Körperfossilien (z. B. Prädationsspuren), sowie von Körperfossilien mit Körperfossilien etwas über die trophischen Beziehungen („Nahrungsketten“) in urzeitlichen Ökosystemen verraten. Ein relativ spektakuläres Beispiel hierfür sind abgebrochene Tyrannosaurus-Zähne, die in Knochen von pflanzenfressenden Dinosauriern stecken.[13] Weniger bekannt, aber wesentlich überraschender waren Funde von Pflanzenresten in der Magengegend des permzeitlichen Reptils Protorosaurus, das anhand seines Gebisses zuvor als reiner Fleischfresser eingestuft worden war.[14]
Des Weiteren können geochemische Analysen von Fossilien in Originalerhaltung, vor allem Messungen der Verhältnisse der stabilen Isotope des Sauerstoffs und des Kohlenstoffs, Aufschluss über die lokalen, und bei entsprechend umfassender Beprobung auch über die globalen Umweltverhältnisse in vergangenen Erdzeitaltern geben, unter anderem über die Paläotemperaturen.[15][16] Eine morphometrische Methode zur Paläotemperaturbestimmung fußt auf einer vermuteten Abhängigkeit der maximalen Körpergröße wechselwarmer Wirbeltiere von der mittleren Umgebungstemperatur, das heißt, dass Größenunterschiede eng verwandter wechselwarmer Arten (z. B. boider Schlangen), die in verschieden alten Schichten einer bestimmten Region auftreten, relative Temperaturunterschiede zwischen den entsprechenden Lebewelten widerspiegeln.[17] Anhand der Mikrostrukturen verschiedener Meeresorganismen wurde ermittelt, dass die Tageslänge auf der Erde infolge von Wechselwirkungen im Erde-Mond-System in den vergangenen Jahrhundertmillionen kontinuierlich zugenommen hat[18] (siehe auch → Geoarchiv).
Zu den ältesten fossilen Belegen für Leben auf der Erde gehören nach heutigem Forschungsstand Stromatolithe aus dem Isua-Grünsteingürtel in Grönland, denen ein Alter von bis zu 3,7 Milliarden Jahren zugeschrieben wird.[19] Sie zeigen, dass die Schelfmeere schon auf der frühen Erde eine große Bedeutung als Lebensraum hatten. Mikroskopische mineralische Strukturen (sogenannte Hämatit-Filamente), die als Fossilien von primitiven Einzellern gedeutet werden, könnten sogar bis zu 4,28 Milliarden Jahre alt sein. Sie stammen aus Bändereisenerzen des Nuvvuagittuq-Grünsteingürtels, die aus absedimentierten Ausfällungen von untermeerischen hydrothermalen Quellen hervorgegangen sein sollen. Damit stützen diese Funde die Hypothese, dass das Leben an solchen Quellen in tieferen Regionen der Meere entstanden ist.[20]
Geologische Zeitskala

Das Gesetz von Steno besagt unter anderem, dass sich in einem ungestörten Schichtenstapel aus Sedimentgesteinen die ältesten Schichten ganz unten und die jüngsten Schichten ganz oben befinden. So lässt sich das Alter einer Schicht oder mehrerer aufeinander folgender Schichten (eines sogenannten Schichtintervalls) relativ zu einer anderen Schicht bzw. einem anderen Intervall angeben. Man spricht dabei auch vom relativen Alter.
Der Engländer William Smith entdeckte gegen Ende des 18. Jahrhunderts, dass Schichten eines bestimmten relativen Alters ganz bestimmte Fossilien aufweisen, die in Schichten eines anderen relativen Alters nicht vorkommen. Diese Beobachtungen machte er sich zunutze, indem er eine Tabelle erstellte, mit der die gesamte geologische Überlieferung Großbritanniens stratigraphisch kategorisiert werden konnte. Die Kategorisierung erfolgte in Form standardisierter Intervalle, die durch das Vorkommen bestimmter Fossilien definiert und nach dem für dieses Intervall typischen Gestein benannt waren (z. B. Chalk, Coal measures etc.).[21] So ließen sich reale Schichtintervalle, anhand ihres Fossilinhaltes einem der Standardintervalle zuordnen (Korrelation), selbst wenn sie nicht aus dem dafür typischen Gestein bestanden. Damit war Smith der Erfinder der Biostratigraphie und der Geologischen Zeitskala, wenngleich seine „Zeitskala“ nur regional anwendbar war und erst im weiteren Verlauf des 19. Jahrhunderts jene Standardskala entwickelt wurde, die im Wesentlichen noch heute weltweit verwendet wird.
Während sich mit Hilfe der geologischen Zeitskala anfangs nach wie vor ausschließlich das relative Alter einer Schicht innerhalb des gesamten geologisch überlieferten Schichtenstapels bestimmen ließ, eröffneten die im 20. Jahrhundert entwickelten Methoden der radiometrischen Datierung schließlich die Möglichkeit, die relativen Zeitintervalle mit absoluten Altersangaben, d. h. mit Zahlenwerten, zu versehen. Seither weiß man, wie alt die Erde tatsächlich ist und dass beispielsweise die Kreidezeit vor etwa 150 Millionen Jahren begann und vor etwa 65 Millionen Jahren endete. Allerdings werden in der Regel nicht die fossilführenden Sedimentgesteine absolut datiert, sondern magmatische, oft pyroklastische Gesteine, die sich ober- und unterhalb fossilführender Schichtintervalle befinden, wodurch sich das numerische Mindest- bzw. Höchstalter des fossilführenden Intervalls ermitteln lässt.
Leitfossilien

Fossilien, anhand derer das relative Alter einer sedimentären Abfolge, im besten Falle einer einzelnen Schicht, bestimmt werden kann, werden als Leitfossilien bezeichnet.
Ein gutes Leitfossil sollte folgende Ansprüche erfüllen:
- erdgeschichtlich kurzzeitiges Auftreten (geringe stratigraphische Reichweite)
- häufiges Auftreten im entsprechenden stratigraphischen Intervall
- weite geographische Verbreitung
- von der eng mit der Ablagerungsumwelt verknüpften Gesteinsausbildung (Fazies) weitgehend unabhängiges Auftreten („faziesbrechend“)
- leichte Kenntlichkeit
Durch diese Anforderungen soll sichergestellt werden, dass möglichst alle fossilhaltigen Schichten auf der Welt auch Leitfossilien enthalten und dass Schichten, die das gleiche Leitfossil aufweisen, im selben, relativ eng definierten Zeitabschnitt der Erdgeschichte gebildet worden sind, unabhängig davon wo auf der Welt sie sich heute befinden oder wo sie sich zum Ablagerungszeitraum befunden haben.
Wichtige Beispiele sind:
- verschiedene Arten von Trilobiten vom Unterkambrium (541 mya) bis zum Obersilur (419,2 mya), ausgestorben ab dem Perm
- verschiedene Arten von Ammoniten vom Unterdevon (419,2 mya) bis zur Kreide-Tertiär-Grenze (66 mya)
Beide Tiergruppen lebten ausschließlich im Meer, was kein Zufall ist, denn die meisten Leitfossilien der klassischen Biostratigraphie sind wirbellose Meerestiere, und Biostratigraphie findet hauptsächlich Anwendung auf marine Sedimente.
Fossilführende Gesteine und Fossilfundstätten

Fossilien finden sich vorwiegend in Sedimentgesteinen des Phanerozoikums, das heißt, in Sedimenten, die im Lauf der letzten etwa 540 Millionen Jahre abgelagert wurden. Aber nicht jedes Sediment ist reich an Fossilien. Einige führen so gut wie keine Körperfossilien, dafür aber viele Spurenfossilien. Schichten mit besonders vielen oder relativ vielen besonders gut erhaltenen Körperfossilien werden als Fossillagerstätten bezeichnet.
Gute Plätze zum Sammeln von Fossilien sind Aufschlüsse fossilführender Schichten, das heißt Stellen, an denen solche Gesteine offen zutage treten. Dies können natürliche Aufschlüsse sein, aber auch Steinbrüche oder Straßen- und Bahnanschnitte. Da das Betreten eines Bergwerks nicht ohne weiteres möglich und überdies recht gefährlich ist, sind bei Hobbysammlern aufgrund ihrer besseren Zugänglichkeit Bergehalden sehr beliebt, beispielsweise die der Kupferschieferbergwerke im Richelsdorfer Gebirge. Bisweilen werden auch Makrofossilien und Spurenfossilien in Bohrkernen gefunden.
Die bedeutendsten Fossilfundstellen sind in der Regel Lokalitäten mit besonders seltenen oder besonders spektakulären Körperfossilien. Meist sind dies Wirbeltierfossilien, aber auch Wirbellose, bei denen die Weichteile erhalten sind, zählen dazu. Zu den wichtigsten Fossilfundstellen in Deutschland gehören folgende Lokalitäten:
- Fundplatz Bilzingsleben (Thüringen), Pleistozän, bekannt durch Funde von Homo erectus
- Sandgrube Grafenrain in Mauer (Baden) bei Heidelberg (Baden-Württemberg), Pleistozän, bekannt durch den Unterkiefer von Mauer (Homo heidelbergensis)
- Steinheim an der Murr (Baden-Württemberg), Pleistozän, bekannt durch Funde des Homo steinheimensis
- Sandelzhausen bei Mainburg (Bayern), Mergel und Tone des Miozäns, besonders bekannt durch Wirbeltierfunde[23]
- Doberg bei Bünde (Nordrhein-Westfalen), marine Mergel des Oligozäns, besonders bekannt durch Funde mariner Wirbeltiere, siehe auch Dobergmuseum
- Geiseltal bei Halle (Saale) (Sachsen-Anhalt), braunkohleführendes Eozän, vor allem bekannt durch die Säugetierfunde
- Grube Messel bei Darmstadt (Hessen), Ölschiefer des Eozän, UNESCO-Weltnaturerbe, vor allem bekannt durch die Säugetierfunde
- Steinbrüche bei Solnhofen und Eichstätt (Bayern), lithographische Kalke des Tithoniums (Oberjura), vor allem durch die Funde des Urvogels Archaeopteryx bekannt
- Holzmaden (Baden-Württemberg), dunkle Tonsteine des Toarciums (Unterjura), vor allem durch Funde von Ichthyosauriern bekannt
- Kupferzell (Baden-Württemberg), bunte Mergel des Lettenkeuper (Mitteltrias), bekannt durch Massenvorkommen von Temnospondylen
- Bromacker (Thüringen), rote Sandsteine des Artinskiums (Unterperm), besonders bekannt durch Funde von „Ur-Sauriern“ (reptiliomorphe Amphibien und frühe Amnioten)
- Ziegeleigrube Vorhalle bei Hagen (Nordrhein-Westfalen), Tonsteine des Namuriums (Oberkarbon), besonders bekannt durch Funde terrestrischer Gliederfüßer
- Bundenbach (Rheinland-Pfalz), dunkle Tonschiefer des Emsiums (Unterdevon), bekannt durch detaillierte Erhaltung wirbelloser Meerestiere
Populäre Fossilfunde

Nicht nur Dinosaurierfunde professioneller Paläontologen erregten öffentliche Aufmerksamkeit, viele bekannte Funde sind auch Hobbypaläontologen zu verdanken. So zum Beispiel die verschiedenen Exemplare des „Urvogels“ Archaeopteryx aus dem Solnhofener Plattenkalk. Ein weiteres Beispiel für einen weithin bekannten Fund ist das ungewöhnlich vollständige Skelett „AL 288-1“ eines weiblichen Australopithecus afarensis, genannt „Lucy“.
Für seine ungewöhnliche Erhaltung bekannt wurde der im Eis konservierte Körper des Steppenbisons Blue Babe. Der weltweit größte gefundene Ammonit (Kopffüßer) Parapuzosia seppenradensis, mit über 170 Zentimeter Gehäusedurchmesser, befindet sich heute im Naturkundemuseum Münster.
Abgrenzung
Pseudofossilien
Als Pseudofossilien oder Scheinfossilien werden (natürliche) anorganische Bildungen bezeichnet, die an Organismen erinnern (vgl. Lusus naturae). Zu den bekanntesten Erscheinungen dieser Art gehören die an filigrane Korallenstrukturen oder Pflanzen erinnernden Mangandendriten, wie sie beispielsweise häufig im Solnhofener Plattenkalk anzutreffen sind, oder Faserkalk, der mitunter fossilisiertem Holz ähnelt.
Dubiofossilien
Dubiofossilien sind entweder echte Fossilien, deren taxonomische Stellung zweifelhaft ist,[24] oder aber Bildungen, deren Status als echtes Fossil umstritten ist. Sie stehen damit zwischen Fossilien mit taxonomisch klarem Status und Pseudofossilien.[24] Ein relativ bekanntes Beispiel geben die mikroskopischen Strukturen im Mars-Meteoriten ALH84001, bei denen es sich um fossile Bakterien handeln könnte, deren Ursprung aber auch anders erklärbar ist.[25]
Fossilienfälschungen
In der Geschichte der Paläontologie kam es immer wieder zu aufsehenerregenden Fälschungen von Fossilien. Einer der ältesten Fälle spielte sich im Franken des 18. Jahrhunderts ab: die sogenannten Würzburger Lügensteine, „Figurensteine“ aus gebranntem Ton, wurden einem angeblich arglosen Naturforscher untergeschoben.[26] Aus jüngerer Zeit stammte der sogenannte Piltdown-Mensch (Eoanthropus dawsoni), Fragmente eines vermeintlichen Frühmenschenschädels, die 1912 in einer Kiesgrube in England „gefunden“ wurden, sich aber letztlich als Bruchstücke des Craniums eines modernen Menschen, kombiniert mit dem unvollständigen Unterkiefer eines Orang-Utans herausstellten.[27] Noch jüngeren Datums ist der Fall des Archaeoraptor, eines angeblichen Bindeglieds zwischen Nicht-Vogel-Dinosauriern und Vögeln. Diese Fälschung war aus einem Vogelfossil und Teilen von Nicht-Vogel-Dinosaurierfossilien aus der gleichen Formation zusammengeleimt worden und beschädigte um das Jahr 2000 die Reputation des populärwissenschaftlichen Magazins National Geographic nachhaltig.[28][29]
Dies sind allerdings nur die spektakulärsten Fälle. Darüber hinaus existiert eine Vielzahl gefälschter Fossilien, die mitunter auf Fossilienbörsen angeboten werden, nicht zu verwechseln mit Kopien (Abgüssen) echter Fossilien, die als solche ausgewiesen werden.[30]
Lebende Fossilien
Ein sogenanntes lebendes Fossil ist eine heute existierende Tier- oder Pflanzenart, die sich in ihrem grundlegenden Körperbau von ihren Vorfahren nur unwesentlich unterscheidet. Schon aus der Eigenschaft „lebend“ geht hervor, dass es sich nicht um Fossilien im Wortsinn handelt, sondern um bildhaften Sprachgebrauch.
Fossilien als Wappentiere
Gehäuse bzw. Schalen von Trilobiten, Ammoniten und Schnecken kommen in der Heraldik selten als Wappentiere vor; ihre Verwendung deutet auf lokale Fundstätten hin.
 Trilobit im Wappen der Gemeinde Skryje, Tschechien
Trilobit im Wappen der Gemeinde Skryje, Tschechien Trilobit im Wappen der Gemeinde Jince, Tschechien
Trilobit im Wappen der Gemeinde Jince, Tschechien Trilobit im Wappen der Gemeinde Murero, Spanien
Trilobit im Wappen der Gemeinde Murero, Spanien Ammonit im Wappen der Ortschaft Heersum, Niedersachsen
Ammonit im Wappen der Ortschaft Heersum, Niedersachsen Ammonit im Wappen der Gemeinde Cremlingen, Niedersachsen
Ammonit im Wappen der Gemeinde Cremlingen, Niedersachsen Ammonit im Wappen der Gemeinde Erkerode, Niedersachsen
Ammonit im Wappen der Gemeinde Erkerode, Niedersachsen Ammonit im Wappen der Gemeinde Evessen, Niedersachsen
Ammonit im Wappen der Gemeinde Evessen, Niedersachsen Ammonit im Wappen des Gemeindeteils Destedt, Niedersachsen
Ammonit im Wappen des Gemeindeteils Destedt, Niedersachsen Ammonit im Wappen des Ortsteils Hemkenrode, Niedersachsen
Ammonit im Wappen des Ortsteils Hemkenrode, Niedersachsen
Literatur
- Bernhard Ziegler: Einführung in die Paläobiologie Teil 1: Allgemeine Paläontologie. 5. Auflage. E. Schweizerbart’sche Verlagsbuchhandlung, Stuttgart 1992, ISBN 3-510-65316-5.
- Richard Moody: Fossilien – Versteinerte Zeugnisse der Vergangenheit. Albatros Verlag, Zollikon 1977, ohne ISBN.
Weblinks
- Die deutschsprachige Community von Fossiliensammlern für Fossiliensammler
- Fossilien – das Weltkulturerbe schlechthin von Reinhold Leinfelder u. a. In: Messeheft der Münchner Mineralientage Oktober 2004 (PDF-Datei; 1,1 MB)
- The Life of a Vertebrate Fossil veröffentlicht vom: Smithsonian's National Museum of Natural History
- Beispiele für Fossilien aller Erdzeitalter (private Seite)
- Vergessen Sie Archaeopteryx! Ein Artikel zu gefälschten Fossilien von A. Kunkel & W. Werner, München. Publiziert in: avisio 1/2002 (Bayerisches Staatsministerium für Wissenschaft, Forschung und Kunst) (PDF-Datei)
- Eintrag zu Fossilien in Österreich im Austria-Forum
Einzelnachweise
- General Questions: What is a fossil? San Diego Natural History Museum.
- Vgl. Duden online: Petrefakt.
- Donald Prothero: Fossil Record. In: Ronald Singer (Hrsg.): Encyclopedia of Paleontology. Fitzroy Dearborn Publishers, Chicago 1999, ISBN 1-884964-96-6, S. 491.
- Martin Schmeisser: Erdgeschichte und Paläontologie im 17. Jahrhundert: Bernard Palissy, Agostino Scilla, Nicolaus Steno und Leibniz. In: Herbert Jaumann (Hrsg.): Diskurse und Gelehrtenkultur in der Frühen Neuzeit. Ein Handbuch. De Gruyter, Berlin/Boston 2010, S. 809–858, doi:10.1515/9783110241068.809
- Udo Kindermann: Conchae marinae. Marine Fossilien in der Fachliteratur des frühen Mittelalters. Geologische Blätter für Nordost-Bayern und angrenzende Gebiete. Bd. 31, 1981, S. 515–530.
- Erich Thenius: Evolution des Lebens – und der Mensch. Die erdgeschichtliche Dokumentation. Vortrag, Wien 1980, S. 79–139 (zobodat.at [PDF]).
- Jean-Baptiste Lamarck: Sur les fossiles. In: Systême des animaux sans vertèbres, ou Tableau géneral des classes, des ordres et des genres de ces animaux. Paris 1801, doi:10.5962/bhl.title.14255, S. 403–411.
- Ulrich Lehmann: Paläontologisches Wörterbuch. 4., durchgesehene und erweiterte Auflage. Ferdinand Enke Verlag, Stuttgart 1996, ISBN 3-432-83574-4, S. 232.
- diverse Beispiele hierfür finden sich in Ralf-Otto Niedermeyer, Reinhard Lampe, Wolfgang Janke, Klaus Schwarzer, Klaus Duphorn, Heinz Kliewe, Friedrich Werner: Die Deutsche Ostseeküste. Sammlung geologischer Führer, Band 105. 2., völlig neu bearbeitete Auflage. Borntraeger, Stuttgart 2011, ISBN 978-3-443-15091-4.
- Phillip L. Manning, Peter M. Morris, Adam McMahon, Emrys Jones, Andy Gize, Joe H. S. Macquaker, George Wolff, Anu Thompson, Jim Marshall, Kevin G. Taylor, Tyler Lyson, Simon Gaskell, Onrapak Reamtong, William I. Sellers, Bart E. van Dongen, Mike Buckley, Roy A. Wogelius: Mineralized soft-tissue structure and chemistry in a mummified hadrosaur from the Hell Creek Formation, North Dakota (USA). Proceedings of the Royal Society B. Bd. 276, 2009, S. 3429–3437, doi:10.1098/rspb.2009.0812, PMC 2817188 (freier Volltext).
- David A. Grimaldi: Amber – Window to the Past. Harry N. Abrams, New York 1996, ISBN 0-8109-1966-4.
- siehe z. B. Kapitel Muscheln (Birgit Niebuhr, Simon Schneider, Markus Wilmsen; S. 83–168; PDF 7,1 MB) und Ammoniten (Markus Wilmsen, Emad Nagm; S. 201–204; PDF 10,0 MB) in: Birgit Niebuhr, Markus Wilmsen (Hrsg.): Kreide-Fossilien in Sachsen, Teil 1. Geologica Saxonica. Bd. 60, Nr. 1, 2014.
- Robert A. DePalma, David A. Burnham, Larry D. Martin, Bruce M. Rothschild, Peter L. Larson: Physical evidence of predatory behavior in Tyrannosaurus rex. Proceedings of the National Academy of Science of the United States of America. Bd. 110, Nr. 31, 2013, S. 12560–12564, doi:10.1073/pnas.1216534110, PMC 3732924 (freier Volltext).
- Johannes Weigelt: Über die vermutliche Nahrung von Protorosaurus und über einen körperlich erhaltenen Fruchtstand von Archaeopodocarpus germanicus aut. Leopoldina, Reihe 2. Bd. 6, 1930, S. 269–280.
- Qiong Li: Belemnite Palaeoproxies and Dating of Mesozoic Carbonates. PhD Thesis, Faculty of Mathematics and Physical Sciences, University College London, 2011 (online), S. 30 ff.
- H.-G. Attendorn, R. N. C. Bowen: Radioactive and Stable Isotope Geology. ISBN 978-94-010-6467-5 (Kapitel Isotopes in palaeoclimatology and palaeoecology, S. 397–418).
- Jason J. Head, Jonathan I. Bloch, Alexander K. Hastings, Jason R. Bourque, Edwin A. Cadena, Fabiany A. Herrera, P. David Polly, Carlos A. Jaramillo: Giant boid snake from the Palaeocene neotropics reveals hotter past equatorial temperatures. Nature. Bd. 757, Nr. 7357, 2009, S. 715–717, doi:10.1038/nature07671.
- C. T. Scrutton: Periodic growth features in fossil organisms and the length of the day and month. In: Peter Brosche, Jürgen Sündermann (Hrsg.): Tidal Friction and the Earth’s Rotation. Springer-Verlag, Berlin/Heidelberg/New York 1978, S. 154–196, ISBN 978-3-540-09046-5.
- Allen P. Nutman, Vickie C. Bennett, Clark R. L. Friend, Martin J. Van Kranendonk, Allan Chivas: Rapid emergence of life shown by discovery of 3,700-million-year-old microbial structures. In: Nature. Bd. 537, 2016, S. 535–538, doi:10.1038/nature19355 (alternativer Volltextzugriff: ResearchGate)
- Matthew S. Dodd, Dominic Papineau, Tor Grenne, John F. Slack, Martin Rittner, Franco Pirajno, Jonathan O’Neil, Crispin T. S. Little: Evidence for early life in Earth’s oldest hydrothermal vent precipitates. In: Nature. Bd. 537, 2017, S. 60–64, doi:10.1038/nature21377; siehe dazu auch World's oldest fossils unearthed. Pressemitteilung auf der Website des University College London vom 1. März 2017.
- William Smith: Strata Identified by Organized Fossils, Containing Prints on Colored Paper of the Most Characteristic Specimens in Each Stratum. W. Arding, London 1816 (archive.org).
- M. J. Melchin, P. M. Sadler, B. D. Cramer, R. A. Cooper, F. M. Gradstein, O. Hammer: The Silurian Period. In: Felix M. Gradstein, James G. Ogg, Mark Schmitz, Gabi Ogg (Hrsg.): The Geologic Time Scale 2012. Elsevier, 2012 doi:10.1016/B978-0-444-59425-9.00021-4, S. 527 ff.
- Volker Fahlbusch, Renate Liebreich: Hasenhirsch und Hundebär: Chronik der tertiären Fossil-Lagerstätte Sandelzhausen bei Mainburg. Hrsg.: Freunde der Bayerischen Staatssammlung für Paläontologie und historische Geologie München e. V. Verlag Dr. Friedrich Pfeil, 1996, ISBN 978-3-931516-07-9.
- Ulrich Lehmann: Paläontologisches Wörterbuch. 4., durchgesehene und erweiterte Auflage. Ferdinand Enke Verlag, Stuttgart 1996, ISBN 3-432-83574-4, S. 70.
- Dirk Schulze-Makucha, Alberto G. Fairéna, Alfonso F. Davilaa: The case for life on Mars. International Journal of Astrobiology. Bd. 7, Nr. 2, 2008, S. 117–141, doi:10.1017/S1473550408004175.
- Birgit Niebuhr: Wer hat hier gelogen? Die Würzburger Lügenstein-Affaire. Fossilien. Nr. 1/2006, 2006, S. 15–19 (PDF (Memento vom 13. September 2014 im Internet Archive) 886 kB).
- Keith Stewart Thomson: Piltdown Man: The Great English Mystery Story. American Scientist. Bd. 79, Nr. 3, 1991, S. 194–201 (JSTOR 29774366).
- Timothy Rowe, Richard A. Ketcham, Cambria Denison, Matthew Colbert, Xing Xu, Philip J. Currie: The Archaeoraptor forgery. Nature. Bd. 410, 2001, S. 539–540, doi:10.1038/35069145.
- Zhonghe Zhou, Julia A. Clarke, Fucheng Zhang: Archaeoraptor’s better half. Nature. Bd. 420, 2002, S. 285, doi:10.1038/420285a.
- Joan Corbacho, Consuelo Sendino, M’Hamed Tahiri: Palaeontological fakes. Batalleria. Bd. 16, 2011, S. 37–45 (ResearchGate).