Seeigel
Die Seeigel (Echinoidea, von griechisch ἐχῖνος echĩnos: Igel, Seeigel) bilden eine der Klassen, die dem Stamm der Stachelhäuter angehören. Es sind in allen Meeren lebende, wirbellose Tiere.
| Seeigel | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
_and_Echinometra_viridis_(Reef_Urchin_-_bottom).jpg.webp)
Die beiden regulären Seeigel Tripneustes ventricosus (oben) und Echinometra viridis (unten) in einem Riff | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Echinoidea | ||||||||||||
| Leske, 1778 |
Die Klasse wird in 14 Ordnungen aufgeteilt. Bis heute sind etwa 950 Arten bekannt. Sie bewohnen die Meere seit rund 480 Millionen Jahren.[1]
Bau

„Gehäuse“
Das Skelett von Seeigeln besteht aus Kalziumkarbonat („kohlensaurer Kalk“, CaCO3) in der Modifikation Calcit. Der Teil des Skelettes, der die inneren Weichteile schützend umschließt, wird zwar oft als „Gehäuse“ oder „Schale“ bezeichnet, jedoch liegt dieses Gehäuse, anders als das Kalkgehäuse von Schnecken oder die Klappen von Muscheln, unterhalb der dünnen Epidermis.[2] Deshalb handelt es sich, im Gegensatz zu den Gehäusen und Schalen der meisten anderen wirbellosen Tiere, um ein Kalk-Innenskelett.
Das Gehäuse der regulären Seeigel („Regularia“), die die typischen langstacheligen Formen stellen, zeichnet sich durch eine regelmäßige fünfzählige (pentamere) Radialsymmetrie aus. Je nach Art kann es fast kugelförmig oder deutlich ellipsoid abgeplattet sein. Es besteht im Wesentlichen aus 5 Doppelreihen von Ambulakralplatten (Ambulacralia) und 5 Doppelreihen von Interambulakralplatten (Interambulacralia). Die einzelnen Gehäuseplatten, bei denen es sich um Calcit-Einkristalle handelt, haben jeweils die Form unregelmäßiger Fünf- oder Sechsecke. Die Ambulakralplatten sind kleiner als die Interambulakralplatten und enthalten kleine Löcher (Ambulakralporen), durch die die inneren Anteile der Ambulakralfüßchen hindurchtreten. Sowohl Ambulakral- als auch Interambulakralplatten tragen runde Gelenkhöcker („Warzen“), die als Lager für die Stacheln dienen.
.jpg.webp)
Die als irreguläre Seeigel (Irregularia) zusammengefassten Formen, zu denen unter anderem die Sanddollars und die Herzigel gehören, unterscheiden sich sehr von den regulären Seeigeln. Ihr Skelett ist oft abgeflacht (extrem bei den Sanddollars) und nicht radialsymmetrisch, sondern bilateralsymmetrisch, wenngleich die ursprüngliche Pentamerie noch erkennbar ist. Es gibt bei diesen Vertretern somit ein Vorne und Hinten sowie ein Links und Rechts, und die Fortbewegung findet immer mit einer Tendenz nach vorne statt. Stacheln sind in ihrer Größe stark reduziert, nicht aber in ihrer Anzahl, sodass das Stachelkleid entfernt an ein Haarkleid erinnert. Diese Unterschiede im Körperbau sind als Anpassung an eine grabende Lebensweise entstanden.[3]
Stacheln
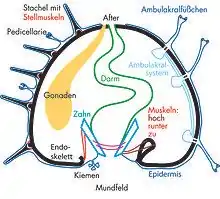
Die Stacheln der Seeigel bestehen ebenfalls aus Calcit und liegen zumeist ebenfalls unterhalb der Epidermis. Sie sitzen auf kleinen Gelenkhöckern und sind durch Muskeln in alle Richtungen beweglich. Die Ausbildung der Stacheln, ihre Größe und Funktion können je nach Art sehr unterschiedlich sein. Bei den regulären Seeigeln werden drei Haupttypen unterschieden, bei den irregulären gar zehn bis zwölf. Die Stacheln dienen vor allem zum Schutz vor Räubern wie Seesternen, großen Schnecken und Fischen und können bei einigen Arten beim Stechen sogar Gift abgeben. Besonders giftig sind die Lederseeigel (Feuerseeigel). Es gibt Seeigel, die sich mit ihren Stacheln in Hartsubstrate wie Korallenriffe und Felsen einbohren. Manche irreguläre Seeigel holen mit den Stacheln Nahrung heran. Bewohner weicher Böden benutzen ihre Stacheln auch zur Fortbewegung.
Tritt ein Mensch auf einen Stachel, bricht dieser unter Umständen ab und bleibt im Fuß stecken, was zu schmerzhaften eitrigen Entzündungen führen kann. Zudem sind die Stacheln einiger Arten schwierig zu entfernen.
Pedicellarien
Pedicellarien sind kleine zangenförmige, evolutiv aus Stacheln hervorgegangene Anhänge. Ihre Funktion besteht darin, die Oberfläche des Seeigels zu reinigen und Parasiten zu entfernen. Einige der Pedicellarien verfügen über Giftdrüsen.
Periprokt (und Peristom)
Der Bereich um die Afteröffnung wird bei Seeigeln aus mehreren, unregelmäßig angeordneten Kalkplatten gebildet und als Analfeld oder Periprokt bezeichnet.[4] Bei den radialsymmetrischen regulären Seeigeln befindet sich das Analfeld zentral auf der Oberseite des Gehäuses, gegenüber der Mundöffnung (Peristom), die zentral auf der Unterseite liegt.
Bei den bilateralsymmetrischen irregulären Seeigeln markiert das Periprokt hingegen das hintere Ende des Gehäuses. Meist weist es direkt nach hinten, aber bei einigen Formen liegt es sogar im hinteren Bereich der Gehäuseunterseite. Das Peristom liegt bei irregulären Seeigeln nicht zentral auf der Unterseite, sondern ist nach vorn versetzt.
Mundapparat

Zum Zerkleinern ihrer Nahrung wie Seetang oder Aas besitzen die regulären Seeigel einen speziellen, komplex aufgebauten Kieferapparat mit calcitischen Hartteilen, die Laterne des Aristoteles. Bei den irregulären Seeigeln ist dieser Kieferapparat entweder teilweise oder vollständig reduziert.
Ambulakralsystem
.jpg.webp)
Der Innenraum der Seeigel ist durch eine Reihe schlauchförmiger, flüssigkeitsgefüllter Hohlräume – das Ambulakralsystem – gegliedert. Allen Seeigeln (und auch Seesternen) gemeinsam sind die Ambulakralfüßchen, dünne schlauchartige Fortsätze, deren innere Anteile Ausstülpungen des Ambulakralsystems und deren äußere Anteile Ausstülpungen der Epidermis sind. Sie können durch Veränderung des Druckes der in den inneren Anteilen enthaltenen Flüssigkeit (Hämolymphe) bewegt werden, enden meistens in einer kleinen epidermalen Saugscheibe und werden unter anderem zur Fortbewegung, Befestigung am Untergrund oder zum Transport von Nahrungspartikeln zum Mund benutzt.
Fressfeinde
Zu den Fressfeinden der Seeigel gehören manche Seesterne. Da die Art Pycnopodia helianthoides (Sonnenblumen-Seestern) bedingt durch die Überhitzung des Lebensraums im Rahmen des Klimawandels und aufgrund eines Virus gegenwärtig von einem Massensterben betroffen ist, verlieren Seeigel einen bedeutenden Fressfeind und breiten sich stellenweise explosionsartig aus, was sich wiederum auf die Bestände von Seetang auswirkt, der für Seeigel als Nahrungsquelle dient.[5]
Fortpflanzung
Die Seeigel sind getrenntgeschlechtlich. Ei- und Samenzellen werden in großen Mengen ins Wasser abgegeben. Die planktonisch lebenden, bilateralsymmetrischen Sekundärlarven der Seeigel werden als Plutei (Einzahl Pluteus) bezeichnet. Bei Seeigeln vor Elba im Mittelmeer wurde in ca. 40 m Tiefe die Bildung von Fortpflanzungsgruppen beobachtet. Dabei finden sich mehrere (10–30) Individuen an einem Ort zusammen, zum Großteil so nah, dass sie sich mit den Stacheln berühren. In diesen Gruppen werden dann gleichzeitig von vielen Tieren Ei- und Samenzellen abgegeben und damit die Wahrscheinlichkeit einer erfolgreichen Befruchtung deutlich erhöht.






Forschungsgeschichte
Entwicklungsbiologie
Oscar Hertwig beobachtete 1875 die Befruchtung des Seeigeleies; die Ergebnisse verhalfen ihm zur Habilitation an der Universität Jena.[6] Den Anfang neuer Individuen wollte er auch experimentell verstehen. In Triest und Spezia hatte er Zugriff vor allem auf Strongylocentrotus lividus und Echinus microtuberculatus als Untersuchungsmaterial. Weniger Beachtung fand Sphaerechinus granularis.[7]
Zu jener Zeit war die Vereinigung von Ei und Spermium ein aktuelles Thema, das mehrere Forscher faszinierte.[8] Auch Theodor Boveri verdankte den Seeigeln einen Teil seines wissenschaftlichen Rufes als Zellbiologe und Chromosomen-Forscher.[9][10] Er untersuchte experimentell die Pathologie der Zellteilung und verwies früh auf das Problem der Tumorentstehung.[11][12]
Molekularbiologie
Die Gentechnik verdankt auch Seeigeln ihren Anfang. Mit dem Restriktionsenzym HindIII gelang es, DNA eines Histongens von Psammechinus miliaris in einen zurecht gemachten Lambda-Phagen einzusetzen.[13] Mit Seeigeln hat man gelernt, dass die Histone jene Proteine sind, die die DNA zum Chromatin beziehungsweise zu Chromosomen verpacken.[14] Histone machen den strukturellen Unterschied zwischen Euchromatin und Heterochromatin.[15]
Die molekular begründete Biologie erhellt, dass die Gestaltwerdung eines Seeigels (und anderer Organismen) von einem genetischen Netzwerk gesteuert ist. Die zitierte Untersuchung erklärt die Gestaltbildung von den Furchungsstadien über Gastrulation bis zum Larvenstadium, dem Pluteus.[16]
Die Molekularbiologie deckt auf, wie die Seeigel mit den anderen Echinodermen verwandt sind.[17] Es braucht molekulare Genetik, um zu verstehen, wie die embryonale zweiseitige Symmetrie sich in Fünfstrahligkeit wandelt.[18] Hier ist auch die Antwort zu erwarten, was den genetischen Unterschied zwischen Seeigeln und Seesternen ausmacht.[19]
Systematik
- Unterklasse: Perischoechinoidea
- Unterklasse: Cidaroidea
- Ordnung: Lanzenseeigel (Cidaroida)
- Familie: Cidaridae
- Familie: Psychocidaridae
- Ordnung: Lanzenseeigel (Cidaroida)
- Unterklasse: Euechinoidea
- Ordnung: Lederseeigel oder Feuerseeigel (Echinothurioida)
- Familie: Echinothuriidae
- Familie: Kamptosomatidae
- Familie: Phormosomatidae
- Ordnung: Micropygoida
- Familie: Micropygidae
- Ordnung: Aspidodiadematoida
- Familie: Aspidodiadematidae
- Ordnung: Diadematoida
- Familie: Diadematidae
- Ordnung: Pedinoida
- Familie: Pedinidae
- Überordnung: Echinacea
- Ordnung: Arbacioida
- Familie: Arbaciidae
- Ordnung: Echte Seeigel (Echinoida)
- Familie: Echinidae
- Familie: Echinometridae
- Familie: Parasaleniidae
- Familie: Strongylocentrotidae
- Ordnung: Temnopleuroida
- Familie: Temnopleuridae
- Familie: Toxopneustidae
- Ordnung: Arbacioida
- Überordnung: Calycina
- Ordnung: Salenioida
- Familie: Saleniidae
- Ordnung: Phymosomatoida
- Familie: Phymosomatidae
- Familie: Stomechinidae
- Ordnung: Salenioida
- Ordnung: Lederseeigel oder Feuerseeigel (Echinothurioida)
- Infraklasse: Irregularia
- Ordnung: Holectypoida †
- Ordnung: Echinoneoida
- Familie: Echinoneidae
- Überordnung: Neognathostomata
- Ordnung: Cassiduloida
- Familie: Apatopygidae
- Familie: Cassidulidae
- Familie: Echinolampadidae
- Familie: Neolampadidae
- Familie: Pliolampadidae
- Ordnung: Sanddollars (Clypeasteroida)
- Familie: Arachnoididae
- Familie: Clypeasteridae
- Familie: Fibulariidae
- Familie: Laganidae
- Familie: Rotulidae
- Familie: Astriclypeidae
- Familie: Dendrasteridae
- Familie: Echinarachniidae
- Familie: Mellitidae
- Ordnung: Cassiduloida
- Überordnung: Atelostomata
- Ordnung: Holasteroida
- Familie: Calymnidae
- Familie: Holasteridae
- Familie: Pourtalesiidae
- Familie: Urechinidae
- Ordnung: Herzseeigel (Spatangoida)
- Familie: Asterostomatidae
- Familie: Aeropsidae
- Familie: Hemiasteridae
- Familie: Pericosmidae
- Familie: Schizasteridae
- Familie: Brissidae
- Familie: Loveniidae
- Familie: Spatangidae
- Familie: Toxasteridae
- Ordnung: Holasteroida
Nutzung
.jpg.webp)

In Südfrankreich, Süditalien, Griechenland, Marokko, Chile und Korea sowie zahlreichen anderen Ländern sind die fünf Geschlechtsdrüsen der Seeigel, roh oder in Salzwasser gekocht, vor allem als Vorspeise beliebt. In der japanischen Küche sind die Eier von Seeigeln unter der Bezeichnung „Uni“ auf der Speisekarte zu finden und werden in unterschiedlichen Gerichten verwendet.
Einzelnachweise
- Der Seeigel − Steckbrief auf biologie-schule.de
- W. Westheide und R. Rieger: Spezielle Zoologie Teil 1, S. 328, ISBN 3-437-20515-3.
- Smith, Andrew B.: Echinoid palaeobiology. Allen & Unwin, London 1984.
- Stichwort „Periprokt.“ In: Herder-Lexikon der Biologie. Spektrum Akademischer Verlag GmbH, Heidelberg 2003, ISBN 3-8274-0354-5.
- C. D. Harvell u. a. (2019). Disease epidemic and a marine heat wave are associated with the continental-scale collapse of a pivotal predator (Pycnopodia helianthoides). Science Advances, 5 (1), eaau7042. https://doi.org/10.1126/sciadv.aau7042
- Oscar Hertwig: Beiträge zur Kenntnis der Bildung, Befruchtung und Teilung des tierischen Eies. 1. Teil. In: Morphologisches Jahrbuch: Eine Zeitschrift für Anatomie und Entwickelungsgeschichte 1, 1875.
- Oscar Hertwig: Experimentelle Studien am Tierischen Ei – vor, während und nach der Befruchtung. Erster Teil. Gustav Fischer, Jena 1890.
- Emil Selenka: Zoologische Studien I: Befruchtung des Eies von Toxopneustes variegatus. Leipzig 1878.
- Theodor Boveri: Zellen-Studien: Über das Verhalten der chromatischen Substanz bei der Bildung der Richtungskörper und bei der Befruchtung. In: Jenaische Zeitschrift für Naturwissenschaft 24, 1890: 314–401. Kapitel 5: Echinus microtuberculatus. 343–348.
- Theodor Boveri: Ueber die Polarität des Seeigeleies. In: Verh Phys-med Ges Würzburg NF 34, 1901: **–**.
- Theodor Boveri: Ueber mehrpolige Mitosen als Mittel zur Analyse des Zellkerns. In: Verh Phys-med Ges Würzburg NF 35, 1902: 67–90.
- Theodor Boveri: Zellenstudien 6: Die Entwicklung dispermer Seeigeleier. Ein Beitrag zur Befruchtungslehre und zur Theorie des Kerns. In: Jenaische Zeitschr Naturw 43, 1908: 1–292.
- Stuart G Clarkson, Hamilton O Smith, Walter Schaffner, Ken W Gross, Max L Birnstiel: Integration of eukaryotic genes for 5S RNA and histone proteins into a phage lambda receptor. In: Nucleic Acids Res 3, 10, 1976: 2617–2632. PDF.
- J Salik, L Herlands, H P Hoffmann, D Poccia: Electrophoretic analysis of the stored histone pool in unfertilized sea urchin eggs: Quantification and identification by antibody binding. In: J Cell Biol 90, 2, 1981: 385–395. PDF.
- Shoshana Levy, Irmingard Sures, Larry Kedes: The nucleotide and amino acid coding sequence of a gene for H1 histone that interacts with euchromatin. The early embryonic H1 gene of the sea urchin Strongylocentrotus purpuratus. In: J Biol Chem 257, 16, 1982: 9438–9443. PDF.
- Deirdre C Lyons, Stacy L Kaltenbach, David R McClay: Morphogenesis in sea urchin embryos: Linking cellular events to gene regulatory network states. In: Wiley Interdiscip Rev Dev Biol 1, 2, 2012: 231–252. PDF.
- Nicolás Mongiardino Koch, Simon E Coppard, Harilaos A Lessios, Derek E G Briggs, Rich Mooi, Greg W Rouse: A phylogenomic resolution of the sea urchin tree of life. In: BMC Evol Biol 18, 2018: 189. PDF.
- Yongxin Li, Akihito Omori, Rachel L Flores, Sheri Satterfield, Christine Nguyen, Tatsuya Ota, Toko Tsurugaya, Tetsuro Ikuta, Kazuho Ikeo, Mani Kikuchi, 20 weitere Autoren sowie Brian Livingston,Cynthia Bradham, Wen Wang, Naoki Irie: Genomic insights of body plan transitions from bilateral to pentameral symmetry in Echinoderms. In: Commun Biol 3, 2020: 37. PDF.
- Gregory A Cary, Brenna S McCauley, Olga Zueva, Joseph Pattinato, William Longabaugh, Veronica F Hinman: Systematic comparison of sea urchin and sea star developmental gene regulatory networks explains how novelty is incorporated in early development. In: Nat Commun 11, 1, 2020: 6235. PDF.
Weblinks
- Seeigel im National Center for Biotechnology Information (NCBI)
- Integrated Taxonomic Information System Echinoidea Leske, 1778
- Dr. Andrew B. Smith, The Natural History Museum, London The Echinoid Directory
- Echinus microtuberculatus, synonym: Psammechinus microtuberculatus.
- Bilder zu Strongylocentrotus lividus.