Palichnologie
Die Palichnologie (seltener auch Paläoichnologie; griech. palaios „alt“, íchnos „Fußspur“, logos „Lehre“) befasst sich als ein Teilgebiet der Paläontologie mit den fossilen Lebensspuren der erdgeschichtlichen Vergangenheit. Solche Spurenfossilien können, wie z. B. Trittsiegel (Fußspuren) und Fährten, eine ehemalige Sedimentoberfläche prägen oder, wie Grabgänge und Wohnbauten, ehemals oberflächennahe Horizonte tiefgehend durchdringen.

Spurenfossilien werden auch Ichnofossilien oder Ichnia (Sing. Ichnium) genannt.[2] Die Bezeichnung Ichnologie (Spurenkunde) wurde erstmals von William Buckland (1784–1856) um 1830 verwendet. Die Palichnologie wird der Neoichnologie gegenübergestellt, die sich mit rezenten Lebensspuren beschäftigt.[2]
Die bislang ältesten Spuren wurden auf einem 565 Millionen Jahre alten Felsen am Mistaken Point der Avalon-Halbinsel in Neufundland (Kanada) entdeckt. Die insgesamt 70, jeweils 13 mm breiten und zwischen 5 und 17 cm langen Spuren stammen von Tieren der Ediacara-Fauna mit einer vermutlich rundlichen Fußscheibe, wie sie die heutigen Seeanemonen besitzen.[3] Die wahrscheinlich ältesten Fährten eines bilateralsymmetrischen Tieres stammen aus 551 bis 541 Millionen Jahre alten Ediacara-Fundschichten im Süden der Volksrepublik China.[4]
Klassifikation fossiler Spuren
Spurentaxonomie (Ichnotaxonomie)


Da es sich bei Spurenfossilien nicht um die Überreste von Tierkörpern handelt und da unterschiedliche Organismengruppen morphologisch sehr ähnliche Spuren erzeugen können und umgekehrt ein und dieselbe Tierart unterschiedliche Spuren hinterlassen kann, klassifiziert die Palichnologie Spuren unabhängig von der zoologischen Taxonomie fossiler Organismen als Formtaxa, die in diesem Fall als Ichnotaxa bezeichnet werden. Hierbei werden in erster Linie Spurengattungen (Ichnogenera) und Spurenarten (Ichnospezies) unterschieden, deren Benennung nach der Linné’schen Nomenklatur erfolgt. Grundlage für die Einordnung von Spuren in die gleiche Spurengattung und die Untergliederung in verschiedene Spurenarten sind gemeinsame morphologische Merkmale, die auf die mehr oder weniger identische Verhaltensweisen des Spurenerzeugers zurückzuführen sind bzw. morphologische Unterschiede, die durch geringfügige Abweichungen im Verhalten der Spurenerzeuger zustande gekommen sind.[6] Dies betrifft die allgemeine Form der Spur (meist gattungsspezifisch), speziell bei Gängen im Sediment auch die Wandungsstruktur, die Raumlage und Besonderheiten bei der Sedimentfüllung (z. B. sogenannte Stopfgefüge). Spurenart- oder -unterartspezifische Unterschiede können aber auch durch Unterschiede im Körperbau der Erzeuger hervorgerufen sein (sogenanntes „Fingerprinting“), was nicht zwangsläufig daran liegen muss, dass die Erzeuger verschiedenen biologischen Arten angehörten, sondern beispielsweise auch ein grundsätzlich ähnliches Verhalten verschiedener Stadien in der Individualentwicklung (Ontogenese) ein und derselben Erzeugerart widerspiegeln kann.
Bei Landwirbeltierfährten besteht, im Gegensatz zu den Spuren vieler wirbelloser Meerestiere, generell ein relativ enger Zusammenhang zwischen der Morphologie der Spur, d. h. dem Abdruck der Extremität, und der Morphologie der Extremität des Erzeugers und damit dessen taxonomischer Stellung. Daher werden in dieser Spurengruppe oft höhere Taxa, wie Spurenfamilien und Spurenordnungen, unterschieden, deren Benamung sich an die Körperfossiltaxonomie anlehnt. Bisweilen werden Landwirbeltierfährtengattungen sogar direkt in die zoologische Systematik eingehängt, was von vielen Palichnologen abgelehnt wird.
Ein sehr bekanntes und zugleich das wissenschaftsgeschichtlich älteste Beispiel für ein Ichnotaxon ist Chirotherium barthii („Barth’sches Handtier“) aus der Trias. Friedrich Sickler veröffentlichte 1834 eine wissenschaftliche Beschreibung dieser Bewegungsspur auf der Grundlage von Fährten auf einer Sandsteinplatte aus dem mittleren Buntsandstein, die in Südthüringen bei Hildburghausen gefunden worden waren, und Johann Jakob Kaup verlieh ihr ein Jahr später den Namen. Über den Verursacher der Spuren kam es jedoch zu einer langanhaltenden wissenschaftlichen Kontroverse. Heute gelten Vertreter der frühen Archosaurier als Chirotherium-Ezeuger.[7]
Ethologische Klassifikation
Die ethologische Klassifikation unterscheidet Spurenfossilien grob nach dem Verhalten des erzeugenden Organismus, das zur Entstehung der entsprechenden Spur führte. Im Folgenden sind die verschiedenen ethologischen Klassen von Spurenfossilien aufgelistet. Die ersten fünf (Ruhespuren bis Wohnspuren) entstammen der traditionellen ethologischen Klassifikation nach Adolf Seilacher (1953). Die übrigen Klassen wurden in jüngeren Arbeiten aufgestellt. Es ist zu beachten, dass es Spuren gibt, die Übergangsformen zwischen zwei Klassen darstellen, die mehreren Klassen zugeordnet werden können oder bei denen nicht genau klar ist, welchem ethologischen Spurentyp sie zuzuordnen sind (z. B. Lumbricaria).
Hinsichtlich der Spurentaxonomie gilt allgemein, dass Spuren verschiedener ethologischer Klassen meist nicht in der gleichen Spurengattung stehen, während Spuren, die der gleichen Spurenart oder -gattung zugeordnet werden, in der Regel auch der gleichen ethologischen Klasse angehören.
Ruhespuren (Cubichnia)
Gräbt sich ein aktiv bewegliches (vagiles) Meerestier vorübergehend ein wenig im Sediment ein und verlässt diesen Platz dann wieder auf gleiche Weise, verursacht es dabei Sedimentstrukturen, die als Ruhespuren (Cubichnia) bezeichnet werden und den Körperumrissen des erzeugenden Tieres z. T. erstaunlich ähnlich sind. Dies geschieht u. a., um (wie z. B. bei Seesternen) eingegrabene Beutetiere zu fangen oder um zu fressen (z. B. bei Muscheln oder einigen Garnelen) oder auch zum Schutz vor Fressfeinden. Die Bezeichnung Ruhespur bezieht sich darauf, dass sie zustande kommt, indem ihr Erzeuger weitgehend stationär („in Ruhe“) bleibt, sich also nicht von A nach B bewegt (vgl. Bewegungsspuren). Typische Beispiele sind Rusophycus, Lockeia und Asteriacites.
Bewegungs- bzw. Kriechspuren (Repichnia)



Bewegungsspuren werden von Organismen verursacht, die sich an der Sedimentoberfläche kriechend, schreitend, dicht über Grund schwimmend o. ä. fortbewegen. Diese Spuren verlaufen üblicherweise in gerade bis leicht kurvig verlaufenden Linien und mit etwa parallelen Seitenrändern. Beispiele hierfür sind Cruziana, Diplichnites, Undichnus, Kouphichnium sowie alle Fährten von Landwirbeltieren (Tetrapoda) (zu Letztgenanntem siehe auch → Fährtensandstein).
Weidespuren (Pascichnia)
Vagile Sedimentfresser erzeugen Weidestrukturen, indem sie sich auf oder oberflächennah schichtparellel in einem Sediment fortbewegen und dabei angereicherten Detritus oder andere Nahrungsquellen „abweiden“, wobei sie meist ein eng mäandrierendes Muster zurücklassen. Typische Vertreter sind Helminthopsis, Nereites und Phycosiphon. Auch Cruziana und Rusophycus werden hier bisweilen mit eingeordnet.
Fressspuren (Fodinichnia)

Im Gegensatz zu den Pascichnia sind Fodinichnia nicht schichtparallel und werden auch in tieferen Niveaus des Meeresbodens durch Sedimentfresser angelegt. Fodinichnia sind meist zusammengesetzt aus einem oder mehreren senkrechten Gängen, die mit der Sedimentoberfläche in Verbindung stehen und der Frischwasserzufuhr dienen, sowie einem oder mehreren annähernd waagerechten Fressgängen. Diese Struktur wird vom Erzeuger sowohl zum Wohnen als auch zum Fressen genutzt. Als Beispiele sind hier Thalassinoides suevicus, Dactyloidites ottoi, Rhizocorallium irregulare, Palaeophycus, Planolites und auch einige Formen von Zoophycos zu nennen.
Wohnspuren (Domichnia)

Ein primär zum dauerhaften Wohnen angelegter Bau wird als Wohnspur (Domichnion) bezeichnet. Solche Bauten werden sowohl in so genanntem Weichsubstrat, meist das unverfestigte Sediment eines Meeres- oder Seebodens, als auch in so genanntem Hartsubstrat, in der Regel hartes Gestein (meist Kalkstein, z. B. an Felsenküsten) oder Außenskelette von Meerestieren, errichtet. Wohnspuren sind Bauten, in denen beispielsweise standortfeste (sessile) Suspensionsfresser, geringmobile Detritusfresser oder Lauerjäger die meiste Zeit ihres Lebens verbringen. Typische Vertreter dieses Spurentyps in (ehemaligen) Weichsubstraten sind z. B. Skolithos, Ophiomorpha und Arenicolites. Domichnia in Hartsubstraten, z. B. Trypanites, und – je nach Definition – auch in Holz (z. B. Teredolites) fallen unter den Oberbegriff Bioerosion.[8] Relativ selten sind Baue von Landwirbeltieren, zu denen die spiralförmige Daemonelix gehört, die vom tertiären Nager Palaeocastor angelegt wurde.
Fallen und Kultivierungsspuren (Agrichnia)
Agrichnia sind fossile Bauten, die Organismen zur Nahrungsbeschaffung als Fallen dienten, wie z. B. für Spirorhaphe und Cosmorhaphe angenommen, oder als „Gewächshaus“ zur Kultivierung von Mikroorganismen, wie z. B. für Paleodictyon und Helicolithes angenommen. Einige solcher Bauten sind möglicherweise extra in tiefen Sedimentschichten mit euxinischem Milieu angelegt worden, da deren Erzeuger mit schwefeloxidierenden Bakterien in Symbiose lebten, wie es z. B. für den Verursacher von Chondrites vermutet wird.
Raubspuren (Praedichnia)
Praedichnia heißen Strukturen, die durch räuberische Aktivitäten verursacht wurden, wie etwa Oichnus, bei dem es sich um Bohrlöcher handelt, die von z. B. Mondschnecken (Naticidae) in Muschelschalen erzeugt wurden, oder Phagophytichnus, Fraßspuren in Blättern. Weitere Beispiele sind von Räubern zerbrochene und zerbissene Außen- und Innenskelette. Praedichnia an Schalentieren und – im weitesten Sinne – auch Fraßspuren an Pflanzen fallen unter den Oberbegriff Bioerosion.[8]
Ausgleichsspuren (Equilibrichnia)
Wenn Organismen ihre Position im Sediment ändern müssen, um sich einer Veränderung der Sedimentoberfläche anzupassen, erzeugen sie Ausgleichsspuren. Dies geschieht, wenn die Sedimentoberfläche langsam abgetragen oder aufgeschüttet wird.
Fluchtspuren (Fugichnia)
Im Gegensatz zu den Equilibrichnia wurde bei diesem Spurentyp durch schnelle Grab-Aktivität des Erzeugers das ursprüngliche Sedimentgefüge gestört, was sich oft durch sichelförmige, feinlaminierte Strukturen äußert. Die Flucht wird durch eine rasche Aufschüttung oder Erosion des Sedimentes, in dem der Erzeuger lebt, ausgelöst. Auch ein Beutegreifer, der nach dem Fugichnium-Erzeuger grub, kann ursächlich sein.
Über dem Substrat angelegte Spuren (Aedificichnia)
Hierzu zählen Spurenfossilien, die auf und/oder über dem Substrat angelegt wurden und aus miteinander verkitteten Sedimentpartikeln geschaffen sind, z. B. Wespennester aus Schlamm, Ameisenhügel, Termitenbauten, Röhren von Köcherfliegenlarven und „Sandorgeln“, die von Röhrenwürmern errichtet werden.
Brutstrukturen (Calichnia)
Calichnia sind zu Fortpflanzungszwecken und zur Aufzucht von Jungtieren und Larven angelegte Strukturen. Dabei verwischen oft die Grenzen zu den Fodinichnia (z. B. Insektenlarven in und auf Blättern) und den Aedificichnia (z. B. hügelartige Dinosauriernester, Köcherfliegenlarvenröhren).
Verankerungsspuren (Fixichnia)
Sessile Meerestiere hinterlassen an der Oberfläche von karbonatischen Hartsubstraten charakteristische Ätznarben, die daher stammen, dass der Verursacher an entsprechender Stelle verankert war. Hierbei werden zwei Typen von Fixichnia unterschieden: Spuren, die auf Verankerung mittels Organen des Weichkörpers (z. B. die Byssusfäden von Muscheln) zurückgehen und Spuren, die dadurch entstanden, dass das Außenskelett des Verursachers am Untergrund festzementiert war. Beispiele für Erstgenanntes sind Podichnus und Centrichnus eccentricus, Beispiele für Letztgenanntes Leptichnus und Centrichnus concentricus. Fixichnia zählen zu den Bioerosionsstrukturen.[8]
Spurenfossilien-Vergesellschaftungen und Bioturbationsgrad
Die Gesamtheit aller Spuren eines bestimmten Raums oder Gesteins wird als Ichnozönose (fossil auch Paläoichnozönose) bezeichnet.[2] Oftmals zeigen Spurenvergesellschaftungen aufgrund unterschiedlicher Lebensweisen und Gewohnheiten seiner Erzeuger eine horizontale Abfolge innerhalb des Sediments, d. h. einen Stockwerkbau. Eine horizontale Abfolge unterschiedlicher fossiler Spuren im Gestein kann jedoch auch Sukzessionen, also eine zeitliche Abfolge unterschiedlicher Spurenfossilien-Vergesellschaftungen, repräsentieren.
Spurenfossilgemeinschaften, die im Verlauf der Erdgeschichte immer wiederkehrend für einen bestimmten Ablagerungsraum typisch waren, werden als Ichnofazies bezeichnet. Eine charakteristische Spuren-Vergesellschaftung kann demnach wichtig für die Interpretation der Genese eines Sedimentgesteins sein. Aussagen zu Umweltfaktoren wie Sauerstoffgehalt, Salinität und Sedimentationsrate in einem marinen Ablagerungsraum lassen sich aus der Analyse von Spurenfossilien-Vergesellschaftungen ableiten.
Die Zerstörung ursprünglicher Sedimentmerkmale durch grabende Organismen wird als Bioturbation bezeichnet. So kann z. B. ein ehemals geschichteter Sand durch Bioturbation homogenisiert werden, so dass die Schichtung ausgelöscht wird. Der Bioturbationsgrad bezeichnet in einer Schicht den prozentualen Anteil des bioturbierten Sediments.
Biostratigraphie
Im Gegensatz zu Körperfossilien besitzen die meisten Spurenfossilien nur begrenztes Potenzial für biostratigraphische Anwendungen. Ichnia konservieren meist nicht die anatomisch eindeutigen Merkmale des Erzeugers, da die Morphologie von Spuren im Regelfall von der Ethologie und nicht von der Anatomie eines Tieres bestimmt wird. So können viele Wirbellosenspuren auf verschiedene Erzeuger zurückgeführt werden. Typische Spurenfossilien ohne biostratigraphische Signifikanz sind Palaeophycus oder Planolites, welche in fast allen Faziesbereichen und stratigraphischen Einheiten zu finden sind. Wahrscheinlicher Verursacher sind Würmer und Gliedertiere (Arthropoden).
Cruziana – Rusophycus

Diese vornehmlich in marinen Sandsteinen des Paläozoikums verbreiteten Spuren werden überwiegend auf die Tätigkeit von Trilobiten zurückgeführt. Die in Fachkreisen weitgehend anerkannte Cruziana-Ichnostratigraphie wurde maßgeblich von Adolf Seilacher entwickelt.[9] Besonders in flachmarinen, körperfossilarmen Siliziklastika kann das Auftreten bestimmter Cruziana-Ichnospezies wertvolle Anhaltspunkte für das relative Alter des Gesteins liefern. Anders als bei den meisten von wirbellosen Tieren verursachten Spuren besteht bei Cruziana und Rusophycus ein enger Zusammenhang zwischen der Anatomie des Exoskeletts, insbesondere der „Beine“, des Erzeugers und der speziellen Morphologie des Spurenfossils („Fingerprinting“). Somit spiegelt sich die Evolution innerhalb der Organismengruppe der Spurenerzeuger in der Spurenfossilüberlieferung wider.
Tetrapodenfährten
In Siliziklastika kontinentaler Sedimentationsräume sind Trittsiegel von Landwirbeltieren (Tetrapoda) oftmals die einzigen fossilen Zeugnisse urzeitlicher Wirbeltiere. Aufgrund des Umstands, dass die Form der Lebensspuren im direkten Zusammenhang mit der Anatomie des Erzeugers steht, lassen sich evolutionäre Trends erkennen und zur zeitlichen Einordnung nutzen. Ein sehr bekanntes Beispiel für Tetrapodenspuren sind die Fährtenzüge aus dem Chiroteriensandstein der Solling-Folge (Mittlerer Buntsandstein). Die ältesten als solche interpretierten Spuren von Landwirbeltieren entstanden in einem Watt. Sie wurden in einem Steinbruch im Heiligkreuzgebirge (südliches Polen) entdeckt und auf ein Alter von 395 Millionen Jahren[10] (Grenzbereich Unter- zu Mitteldevon) datiert. Die ältesten Spuren von Landwirbeltieren aus dem Binnenland sind 318 Millionen Jahre alt und entstanden 500 km von der Küste des Superkontinents Pangaea entfernt.[11]
Treptichnus pedum
Obwohl dieses Spurenfossil keine wesentlichen Veränderung durch die geologische Zeitskala zeigt und lediglich ein schwach spezialisiertes Fressverhalten von Wirbellosen dokumentiert, wird dieser Struktur eine große Bedeutung beigemessen. Treptichnus pedum ist das Leitfossil für die Präkambrium/Kambrium-Grenze und dokumentiert die wohl bedeutendste Veränderung in der Geschichte des Lebens. An der Basis des Phanerozoikums vollzog sich ein Prozess, der gemeinhin als Kambrische Explosion bekannt ist: Innerhalb geologisch kurzer Zeit entstanden fast alle Baupläne der heutigen Organismengruppen. Da insbesondere im untersten Kambrium Körperfossilien selten und schwer stratigraphisch korrelierbar sind, wurde das FAD (englisch: First Appearance Datum, deutsch: Erstauftreten) von Treptichnus pedum zur Definition dieser entscheidenden geologischen Grenze herangezogen, da diese Struktur ein spezialisiertes Fressverhalten darstellt, welches auf die Tätigkeit von komplex gebauten mehrzelligen Tieren (Metazoa) zurückgeführt werden kann. Trotzdem ist die Eignung als Leitfossil umstritten. So wird argumentiert, dass Treptichnus pedum lediglich unterseitiges Abweiden von Algenmatten („undermatmining“) darstellt, eine Fraßtechnik, der sich auch niedere Organismen bedienten und die bereits im oberen Präkambrium weit verbreitet war. Darüber hinaus ist Treptichnus pedum, wie die meisten Spurenfossilien, stark faziesabhängig, eine Eigenschaft, die bei Leitfossilien unerwünscht ist.
Fossile Vormenschen- und Menschenfährten
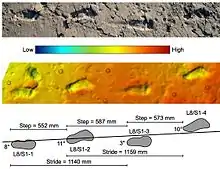


Eine besondere Form von Tetrapodenfährten sind solche, die von bereits aufrecht gehenden (bipeden) Vorfahren oder nahen Verwandten des anatomisch modernen Menschen (Homo sapiens), erzeugt worden sind. Die ältesten dieser Fährten (< 1 mya) kommen ausschließlich in Afrika vor. Bei einer auf Kreta entdeckten, 5,7 mya alten Fußspur ist aufgrund ihres hohen Alters und ihrer geographischen Position unklar, ob sie einem „echten“ Vormenschen (das heißt einem frühen Vertreter der Tribus Hominini) zuzuordnen ist, oder einer Entwicklungslinie, die konvergent eine menschenähnliche Fußanatomie entwickelt hat.[12]
Bei Laetoli in Tansania wurden 1978 entsprechende Fährten entdeckt, die auf ca. 3,6 mya datiert und vermutlich von Australopithecus afarensis erzeugt wurden.[13] Sie gelten als die ältesten von Hominini verursachten Spurenfossilien. Wie das renommierte Wissenschaftsmagazin Nature im April 2008 berichtete, sind diese Spuren vom Zerfall bedroht.[14]
Bei Ileret, östlich des Turkana-Sees in Kenia, wurden 1,51 bis 1,53 Millionen Jahre alte Fußabdrücke von Homo erectus freigelegt. Die Fährten belegen, dass der Bau der Füße im Wesentlichen mit dem des modernen Menschen übereinstimmte und dass der aufrechte Gang von H. erectus dem des modernen Menschen bereits stark geähnelt haben muss.[15] Ebenfalls rund 1,5 Millionen Jahre alt sind einige Fußabdrücke aus der Fundstelle Koobi Fora an der Nordostküste des Turkana-Sees.[16]
Seit 2008 werden rund 120.000 Jahre alte Fußspuren des frühen anatomisch modernen Menschen analysiert, die am damaligen Ufer des Natronsees in Tansania in frische Vulkanasche eingedrückt wurden. Insgesamt sind 350 Abdrücke auf 150 Quadratmetern nachgewiesen worden. Sie stammen von mehr als 30 Individuen (Männer, Frauen und Kinder), die vermutlich in zwei Gruppen unterwegs waren.[17] Ebenfalls 120.000 Jahre alt sind Fußspuren des frühen anatomisch modernen Menschen, die in der Nefud in Saudi-Arabien erhalten geblieben sind.[18]
117.000 Jahre alt sind Fußabdrücke, die in Südafrika, am Rande der sich südöstlich an die Saldanha Bay anschließende Langebaan Lagoon, entdeckt wurden[19][20] und – wie die gleich alten Spuren von Nahoon[21] – ebenfalls dem frühen Homo sapiens zuzurechnen sind. Die von dem Geologen David Roberts entdeckten Langebaan-Spuren wurden in von Regen befeuchtetem Sand am Hang einer Düne eingedrückt und kurze Zeit später mit Sand zugeweht. Durch nachfolgende Erosion befindet sich die Schicht mit der Fährte heute wieder an der Erdoberfläche.
Die ältesten Fußspuren des anatomisch modernen Menschen in Europa wurden 1965 in der Ciur-Izbuc-Höhle in den Westrumänischen Karpaten entdeckt und, basierend auf Begleitfunden von Höhlenbärenknochen und -spuren, zunächst auf ein Mindestalter von 10.000 bis 15.000 Jahren datiert. Eine Neudatierung der mittlerweile teilweise zerstörten Spuren mittels der Radiokarbonmethode, die 2014 publiziert wurde, wies ihnen jedoch ein deutlich höheres Alter von bis zu 36.000 Jahren zu.[22]
Die ältesten Fußspuren von „Urmenschen“ in Europa – vermutlich enge Verwandte der Vorfahren des Neandertalers – wurden in der ostenglischen Grafschaft Norfolk entdeckt. Forscher um Chris Stringer legten sie im Mai 2013 bei Happisburgh frei, datierten sie auf ein Alter von rund 800.000 Jahren und interpretierten sie aufgrund dieses Alters als Spuren von Homo antecessor, da diese Art die einzige bislang aus diesem Zeitraum in Europa beschriebene sei.[23][24]
Vermutlich von Homo heidelbergensis stammen drei 385.000 bis 325.000 Jahre alte Fährten, die in Süditalien (Roccamonfina) zusammen mit anderen Säugetierspuren in pyroklastischen Sedimenten entdeckt wurden. Die Spuren wurden von Individuen, die den Hang hinab liefen, erzeugt. Eine der Fährten ist Z-förmig, was vermutlich darauf zurückzuführen ist, dass ihr Erzeuger teilweise hangparallel lief, um den Abstieg leichter bewältigen zu können. Die zweite Fährte zeigt in ihrem unteren Teil deutliche Anzeichen dafür, dass ihr Erzeuger ins Rutschen geraten sein muss. Handabdrücke neben der Fährte werden als Versuch des Erzeugers gedeutet, durch Abstützen die Balance zu wahren.[25]
Rund 100.000 Jahre alt sind 87 Fußabdrücke von Neandertalern, die im Südwesten von Spanien entdeckt wurden. Sie stammen von zumeist Jugendlichen und einigen Erwachsenen, die – entlang einer Küstenlinie – gemeinsam über einen schlammigen Untergrund gelaufen sind.[26] Auf ein Alter von 97.000 Jahren wurden drei Fußabdrücke eines Neandertalers in einem karbonatischen, quellkalkähnlichen Höhlensediment (Mondmilch) datiert, die bereits 1974 in der Vârtop-Höhle im Bihor-Gebirge (Rumänien) entdeckt worden waren.[27] Rund 70.000 ± 10.000 Jahre alt ist eine Gruppe von mehr als 250 Fußabdrücken von 10 bis 13 vorwiegend sehr jungen und jugendlichen Neandertalern in einem Dünenbereich von Le Rozel im französischen Département Manche in der Region Normandie.[28][29]
23.000 bis 21.000 Jahre alt sind Fußspuren von Kindern und Jugendlichen, die im White-Sands-Nationalpark in New Mexico (Vereinigte Staaten) ausgegraben wurden; einer 2021 in Science veröffentlichen Studie zufolge sind sie der bislang älteste Beleg für die Anwesenheit von Menschen in Amerika.[30]
Siehe auch
Literatur
- R. G. Bromley: Spurenfossilien: Biologie, Taphonomie und Anwendungen. Springer, Berlin/Heidelberg 1999, ISBN 978-3-540-62944-3, 347 S.
- W. Miller, ed.: Trace Fossils: Concepts Problems Prospects. Elsevier, 2006, ISBN 978-0-444-52949-7, 632 S.
- A. Seilacher: Trace Fossils Analysis. Springer-Verlag, Berlin/Heidelberg 2007, ISBN 978-3-540-47225-4, 226 S.
- A. A. Ekdale et al.: Ichnology: trace fossils in sedimentology and stratigraphy. Society of Economic Paleontologists and Mineralogists, Short Course Notes, No. 15. Tulsa, Oklahoma 1984, ISBN 978-0-918985-42-2. 317 S.
Weblinks
- GeoLexikon
- Rätsel im Stein – Auf paläontologischer Spurensuche. Begleitartikel zur gleichnamigen Sonderausstellung des Paläontologischen Museums München – Sonderdruck aus dem offiziellen Katalog der 41. Münchner Mineralientage 2004. S. 134–143 Von Kurt Heißig u. a.
- „Spuren lesen – Vom Abdruck zum Erzeuger.“ (Memento vom 19. Oktober 2014 im Internet Archive) Von Lothar H. Vallon und Martin Röper. In: Paläontologie aktuell – Berichte aus Forschung und Wissenschaft. Internetpräsenz der Paläontologischen Gesellschaft (PDF; 513 kB)
- Paleoichnology (englisch)
Einzelnachweise
- Hartmut Haubold Saurierfährten. Die Neue Brehm-Bücherei Bd. 479. – A. Ziemsen Verlag, Wittenberg Lutherstadt, 1984. ISBN 3-89432-401-5.
- Ulrich Lehmann: Paläontologisches Wörterbuch. 4. Auflage. Ferdinand Enke Verlag, Stuttgart 1996, ISBN 3-432-83573-6.
- Alexander G. Liu, Duncan McIlroy, Martin D. Brasier: First evidence for locomotion in the Ediacara biota from the 565 Ma Mistaken Point Formation, Newfoundland. In: Geology. Band 38, Nr. 2, 2010, S. 123–126, doi:10.1130/G30368.1.
- Zhe Chen, Xiang Chen, Chuanming Zhou, Xunlai Yuan, Shuhai Xiao: Late Ediacaran trackways produced by bilaterian animals with paired appendages. In: Science Advances. Band 4, Nr. 6, eaao6691, doi:10.1126/sciadv.aao6691
- Andreas E. Richter: Handbuch des Fossiliensammlers. S. 405. Weltbild Verlag, Augsburg, 1999. ISBN 3-440-05004-1.
- Bromley: Spurenfossilien. 1999 (siehe Literatur), S. 173
- Martin Lockley: Auf den Spuren der Dinosaurier. Dinosaurierfährten – Eine Expedition in die Vergangenheit. Birkhäuser Verlag, Basel [u. a.] 1993, ISBN 978-3-7643-2774-3, S. 7.
- J. M. de Gibert, R. Domènech, J. Martinell: An ethological framework for animal bioerosion trace fossils upon mineral substrates with proposal of a new class, fixichnia. Lethaia, Bd. 37, 2004, S. 429–437, doi:10.1080/00241160410002144.
- Adolf Seilacher: Cruziana stratigraphy of “non-fossiliferous” Paleozoic sandstones. In: T. P. Crimes, J. C. Harper (Hrsg.): Trace Fossils. In: Geological Journal, Band 3, 1970, S. 447–476. ISBN 0-902354-09-4.
- Grzegorz Niedźwiedzki et al.: Tetrapod trackways from the early Middle Devonian period of Poland. In: Nature, Band 463, 2010, S. 43–48, doi:10.1038/nature08623.
- Howard J. Falcon-Lang et al.: Diverse tetrapod trackways in the Lower Pennsylvanian Tynemouth Creek Formation, near St. Martins, southern New Brunswick, Canada. In: Palaeogeography, Palaeoclimatology, Palaeoecology, Band 296, Nr. 1–2, 2010, S. 1–13. doi:10.1016/j.palaeo.2010.06.020.
- Gerard D. Gierliński, Grzegorz Niedźwiedzki, Martin Lockley, Athanassios Athanassiou, Charalampos Fassoulas, Zofia Dubicka, Andrzej Boczarowski, Matthew R. Bennett, Per Erik Ahlberg: Possible hominin footprints from the late Miocene (c. 5.7 Ma) of Crete? In: Proceedings of the Geologists’ Association. Band 128, Nr. 5–6, 2017, S. 697–710, doi:10.1016/j.pgeola.2017.07.006.
- Mary D. Leakey, R. L. Hay: Pliocene footprints in the Laetoli beds at Laetoli, northern Tanzania. In: Nature. Band 278, 1979, S. 317–323, doi:10.1038/278317a0.
- Anonymus: Tanzania takes steps to save ancient human prints. In: Nature. Band 452, 2008, S. 677, doi:10.1038/452677c.
- „The Ileret prints show that by 1.5 Ma, hominins had evolved an essentially modern human foot function and style of bipedal locomotion.“ Matthew R. Bennett, John W.K. Harris, Brian G. Richmond, David R. Braun, Emma Mbua, Purity Kiura, Daniel Olago, Mzalendo Kibunjia, Christine Omuombo, Anna K. Behrensmeyer, David Huddart, Silvia Gonzalez: Early Hominin Foot Morphology Based on 1.5-Million-Year-Old Footprints from Ileret, Kenya. In: Science. Band 323, Nr. 5918, 2009, S. 1197–1201, doi:10.1126/science.1168132.
- Anna K. Behrensmeyer, Léo F. Laporte: Footprints of a Pleistocene hominid in northern Kenya. In: Nature. Band 289, 1981, S. 167–169, doi:10.1038/289167a0.
- Ann Gibbons: Ancient Footprints Tell Tales of Travel. In: Science. Band 332, Nr. 6029, S. 534–535, doi:10.1126/science.332.6029.534-b.
- Mathew Stewart et al.: Human footprints provide snapshot of last interglacial ecology in the Arabian interior. In: Science Advances. Band 6, Nr. 38, 2020, eaba8940, doi:10.1126/sciadv.aba8940.
Alte menschliche Fußabdrücke in Saudi-Arabien ermöglichen Momentaufnahme der arabischen Ökologie vor 120.000 Jahren. Auf: mpg.de vom 18. September 2020. - David Roberts, Lee Berger: Last interglacial (c. 117 kyr) human footprints from South Africa. In: South African Journal of Science. Band 93, 1997, S. 349–350.
- nationalgeographic.com vom 14. August 1997: Footprints Found in South Africa Come From Dawn of Modern Humans.
Rick Gore: Tracking the First of Our Kind. In: National Geographic. September 1997, S. 92–99. - David L. Roberts: Last Interglacial Hominid and Associated Vertebrate Fossil Trackways in Coastal Eolianites, South Africa. In: Ichnos: An International Journal for Plant and Animal Traces. Band 15, Nr. 3–4, 2008, S. 190–207, doi:10.1080/10420940802470482.
- David Webb, Marius Robu, Oana Moldovan, Silviu Constantin, Bogdan Tomus, Ionel Neag: Ancient human footprints in Ciur-Izbuc Cave, Romania. In: American Journal of Physical Anthropology. Online-Vorabveröffentlichung vom 7. Juli 2014, doi:10.1002/ajpa.22561.
- Nick Ashton et al.: Hominin Footprints from Early Pleistocene Deposits at Happisburgh, UK. In: PLoS ONE. Band 9, Nr. 2, e88329, doi:10.1371/journal.pone.0088329.
- Ashleigh L. A. Wiseman, Chris B. Stringer, Nick Ashton et al.: The morphological affinity of the Early Pleistocene footprints from Happisburgh, England, with other footprints of Pliocene, Pleistocene, and Holocene age. In: Journal of Human Evolution. Band 144, 2020, 102776, doi:10.1016/j.jhevol.2020.102776.
- Paolo Mietto, Marco Avanzini, Giuseppe Rolandi: Human footprints in Pleistocene volcanic ash. In: Nature. Band 422, 2003, S. 133, doi:10.1038/422133a.
- Eduardo Mayoral et al.: Tracking late Pleistocene Neandertals on the Iberian coast. In: Scientific Reports. Band 11, 2021, Artikel-Nr. 4103, doi:10.1038/s41598-021-83413-8.
- Bogdan P. Onac, Iosif Viehmann, Joyce Lundberg, Stein-Erik Lauritzen, Chris Stringer, Vasile Popiţă: U–Th ages constraining the Neanderthal footprint at Vârtop Cave, Romania. In: Quaternary Science Reviews. Band 24, Nr. 10–11, 2005, S. 1151–1157, doi:10.1016/j.quascirev.2004.12.001.
- Norbert Mercier et al.: Dating the palaeolithic footprints of ‘Le Rozel’ (Normandy, France). In: Quaternary Geochronology. Band 49, 2019, S. 271–277, doi:10.1016/j.quageo.2017.12.005.
- Jérémy Duveau, Gilles Berillon, Christine Verna, Gilles Laisné und Dominique Cliquet: The composition of a Neandertal social group revealed by the hominin footprints at Le Rozel (Normandy, France). In: PNAS. Band 116, Nr. 39, 2019, S. 19409–19414, doi:10.1073/pnas.1901789116.
- Matthew R. Bennett et al.: Evidence of humans in North America during the Last Glacial Maximum. In: Science. Band 373, Nr. 6562, 2021, S. 1528–1531, doi:10.1126/science.abg7586.
Ancient footprints could be oldest traces of humans in the Americas. Auf: nature.com vom 23. September 2021.