Gaudeamus
Gaudeamus ist eine ausgestorbene Gattung aus der Gruppe der Stachelschweinverwandten. Innerhalb dieser gehört sie zu den Hystricognathi und zur Familie der Gaudeamuridae, deren genauen Beziehungen noch ungeklärt sind. Die Vertreter der Gattung kamen im Zeitraum vom Oberen Eozän bis zum Unteren Oligozän vor 34 bis 28 Millionen Jahren im heutigen Nordafrika vor. Fossilfunde wurden im Fayyum in Ägypten und vereinzelt an anderen Fundstellen der Region entdeckt. Sie umfassen weitgehend Schädel- und Gebissmaterial, die mehreren Arten zugewiesen werden können. Die Tiere aus der weiteren Verwandtschaft der Stachelschweine waren relativ groß. Besondere Kennzeichen finden sich neben dem allgemein an die Stachelschweine erinnernden Schädel- und Unterkieferbau vor allem in der Gestaltung der Zähne mit querverlaufenden Leisten, ebenso in dem Wechsel der Prämolaren vom Milch- zum Dauergebiss wie auch in einer großen Öffnung am Gaumen. Die Gattung wurde im Jahr 1968 eingeführt. Sie erhielt im Jahr 2011 einen eigenen Familienstatus, was sich mit der charakteristischen Morphologie der Tiere erklärt.
| Gaudeamus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
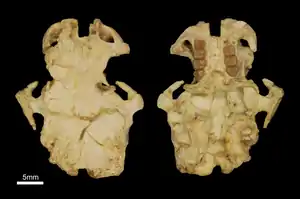
Schädel von Gaudeamus, Holotyp der Art Gaudeamus aslius | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oberes Eozän bis Unteres Oligozän | ||||||||||||
| 34 bis 28 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name der Familie | ||||||||||||
| Gaudeamuridae | ||||||||||||
| Sallam, Seiffert & Simons, 2011 | ||||||||||||
| Wissenschaftlicher Name der Gattung | ||||||||||||
| Gaudeamus | ||||||||||||
| Wood, 1968 | ||||||||||||
Merkmale
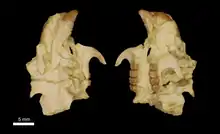
Gaudeamus war ein relativ großer Vertreter der Hystricognathi. Die überlieferten Schädel sind weitgehend vollständig, durch Lagerungsdruck jedoch flachgepresst und teilweise zerbrochen. Das Rostrum war moderat lang. Der Mittelkieferknochen enthielt die beiden Schneidezähne, deren Alveolen den Großteil des Daches ausfüllten. Die seitlichen Fortsätze des Mittelkieferknochens verbanden sich mit dem Nasenbein und dem Oberkiefer. Das Nasenbein reichte nach hinten bis zum letzten Prämolaren oder ersten Molaren. Der Oberkiefer war hoch und barg ein typisch hystricognathes, großes Foramen infraorbitale, an welchem gleichzeitig noch der Musculus masseter ansetzte. Es öffnete sich vor der Zahnreihe und wurde gleichzeitig vom vorderen Ansatz des Jochbogens begrenzt. Möglicherweise war es dadurch ursprünglich am Schädel senkrecht orientiert, was seine Entsprechung bei den heutigen Rohrratten findet, allerdings von den Meerschweinchen und den Quastenstachlern abweicht. Das Stirnbein nahm das mittlere Drittel des Schädels ein. Hier führte der Processus postorbitalis seitlich weg. Er hatte eine dreieckige Form und war größer als bei den meisten heutigen Hystricognathi. Auf dem Stirnbein verliefen zwei Temporallinien. Sie vereinigten sich etwa in der Mitte des Scheitelbeins zu einem schwachen Scheitelkamm. Nach hinten nahm der Scheitelkamm an Stärke zu, vergleichbar zu den Quastenstachlern. Das Scheitelbein selbst formte das hintere Drittel des Schädels. Das Hinterhauptsbein ist bisher nur stark beschädigt überliefert, es wies aber große und nach hinten gestreckte Gelenkflächen für die Verbindung mit der Halswirbelsäule auf. Das Schläfenbein bildete den hinteren Rand der Orbita. An der Schädelbasis war eine große Paukenblase ausgebildet, die etwa ein Viertel der Schädellänge beanspruchte und vollständig aus dem Schläfenbein bestand. Der Gaumen charakterisierte sich durch eine große Öffnung, die bis zum ersten Molaren reichte und als „großes Gaumenfenster“ bezeichnet wird.[1][2]
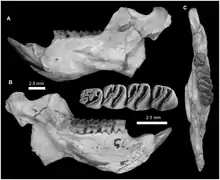
Der Unterkiefer war robust und durch den im Vergleich zur Zahnreihe seitlich abstehenden Winkelfortsatz hystricognath geformt. Dadurch entstand in der Seitenansicht eine deutliche Einbuchtung zwischen dem Winkelfortsatz und dem horizontalen Knochenkörper. An dieser Einbuchtung war wiederum der Massetermuskel verankert. Dagegen schwang der horizontale Knochenkörper deutlich nach unten aus, seine Form wurde durch die Alveolen der Schneidezähne vorgegeben. Die Symphyse endete am vierten Prämolaren. Das Foramen mentale lag unterhalb des letzten Prämolaren oder kurz davor am Ende des Diastemas. Der vordere Rand des aufsteigenden Astes erhob sich seitlich hinter dem letzten Molaren und war schräg nach hinten gerichtet bei einem geschwungenen Verlauf. Noch zwischen dem letzten Molaren und dem Astansatz befand sich eine markante Eindellung, wie sie auch bei den Quastenstachlern vorkommt. Der Kronenfortsatz übertraf den Gelenkfortsatz, letzterer lag wiederum weit oberhalb der Kauebene. Die Gelenkfläche war oval geformt mit der größeren Länge in Längsrichtung des Unterkiefers. Seitlich am aufsteigenden Ast zeichnete sich die Fossa masseterica ab, sie war hinten breit und verengte sich nach vorn, wo sie etwa am Übergang vom letzten Prämolaren zum ersten Molaren ihren Abschluss fand. Oben und unten wurde sie jeweils durch einen kräftigen Knochenwulst flankiert. Der Winkelfortsatz endete spitz.[1][2]
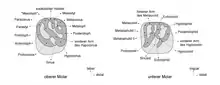
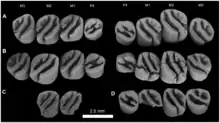

Das Gebiss bestand sowohl oben als auch unten aus je einem Schneidezahn, einem Prämolaren (den letzten) und drei Molaren. Die Schneidezähne repräsentierten Nagezähne. Sie waren stark gebogen und oval im Querschnitt mit der Längsachse quer zum Schädel. Der obere maß 3,57 mm in der Breite und 1,82 mm in der Länge. Zahnschmelz kam nur auf der jeweiligen Vorderseite vor, er bedeckte zusätzlich noch die Hälfte der Seitenflächen. Die Schneidefläche war dreieckig geformt. Die hintere Bezahnung trennte ein langes, jeweils eingebuchtetes Diastema ab. Es erreichte im Unterkiefer die Hälfte der Länge der Zahnreihe, war aber kürzer als im oberen Gebiss. Hier wurde es zu zwei Dritteln vom Mittelkieferknochen gebildet. Die Mahlzähne hatten eine weitgehend rechteckige Form, eine Ausnahme bildete der dreieckige letzte Molar. Generell waren die hinteren Zähne hochkronig (hypsodont) gestaltet. Die Kauoberfläche der Molaren von Gaudeamus besaß die charakteristische Strukturierung, die von den Nagetieren bekannt ist, mit mehreren Höckern und Leisten beziehungsweise Jochen (als Lophen bezeichnet) dazwischen. Die jeweilige Anzahl an Höckern und Leisten hat taxonomischen Wert.[3][4][5] Im oberen Gebiss wiesen die Molaren jeweils vier Höcker auf der Kaufläche auf (Para-, Meta-, Proto- und Hypoconus). Diese standen paarig beisammen und wurden jeweils durch Schmelzleisten verbunden. Im Unterschied zu vielen anderen Verwandten der Stachelschweine verliefen diese Leisten aber nicht rechtwinklig zur Zahnachse, sondern waren diagonal orientiert, wodurch die Zähne prinzipiell denen der Rohrratten ähnelten. Der lippenseitige Paraconus war so über das Protoloph nicht mit dem zungenseitigen Protoconus verbunden. Vielmehr traf es auf einen Ausläufer des benachbarten Hypoconus. An der Zahnvorderseite verlief das Anteroloph direkt zum Protoconus, an der Zahnrückseite saß das Posteroloph zwischen dem Metaconus und dem Hypoconus. Bei anderen Hystricognathi übernahm dies üblicherweise das Metaloph, welches bei Gaudeamus aber extrem kurz ausfiel. Dafür kam bei den meisten Arten von Gaudeamus ein zusätzliches Mesoloph vor, das seitlich vom Metaloph verlief. Insgesamt wirkten die oberen Molaren dadurch tetralophodont (mit vier Leisten). Ähnliches kann zu den Unterkiefermolaren gesagt werden. Auch diese wiesen vier Haupthöcker auf (Meta-, Ento-, Proto- und Hypoconid), ein bei den Hystricognathi häufig ausgebildeter fünfter Höcker, das Hypoconulid am Zahnende, fehlte hier. Das Hypolophid orientierte sich vom zungenseitigen Entoconid schräg in Richtung zum lippenseitigen Protoconid, auf den es über eine Verlängerung traf. Die eigentliche Verbindung zum Hypoconid bestand nicht. Am vorderen Zahnrand verlief das Metalophulid, am hinteren das Posterolophid, das sich vom Hypoconid zum Entoconid erstreckte. Diese drei Leisten gaben den unteren Molaren von Gaudeamus ihr prägnantes Aussehen. Sie teilten die Kauoberfläche außerdem in zwei nahezu gleich große Senken auf. Bei manchen Individuen wurde das vordere Becken durch ein zusätzliches Metalophulid noch einmal halbiert. Im Gegensatz zu den Rohrratten mit ihrer ähnlichen Zahngestaltung kam es bei Gaudeamus zu einem Austausch der Prämolaren vom Milch- zum Dauergebiss. Überliefert sind bisher nur die unteren Milchprämolaren. Diese besaßen typisch für Nagetiere und aufgrund ihrer mahlenden Funktion im ersten Gebiss eine höhere Anzahl an Leisten als die permanenten Prämolaren, bei Gaudeamus waren es bis zu fünf. Sowohl im oberen als auch im unteren Gebiss bildete der jeweils zweite Molar den größten Zahn. Die Größe variierte von 1,88 bis 2,38 mm in der Länge und von 2,00 bis 2,88 mm in der Breite.[1][2]
Fossilfunde
Funde von Gaudeamus sind bisher nur aus Nordafrika überliefert. Die bedeutendste Fundstelle bildet hier die Fayyum-Region im nördlichen Ägypten. Die Gattung konnte an mehreren Fundstellen innerhalb der Gebel-Qatrani-Formation beobachtet werden, die sich auf unterschiedlichen stratigraphischen Abschnitte verteilen. Das bisher älteste und umfangreichste Material kam in L-41 zu Tage. Die Fundstelle, die 1983 von einem Forscherteam um Elwyn L. Simons entdeckt und später unter anderem von D. Tab Rasmussen genauer vorgestellt wurde, befindet sich etwa 46 m über der Basis, im unteren Abschnitt der Gesteinseinheit. Sie gehört paläomagnetischen Untersuchungen zufolge in das Obere Eozän mit absoluten Altersangaben um etwa 34 Millionen Jahren. Aufgrund der eher feinkörnigen Ablagerungen, die wohl auf einen ehemaligen See zurückgeführt werden können, und des hohen Anteils an kleineren Wirbeltieren unterscheidet sich L-41 deutlich von den anderen Fundstellen der Gebel-Qatrani-Formation.[6][7] Allein die Nagetiere umfassen hier hunderte Fossilfunde, darunter neben Gaudeamus auch Birkamys, Mubhammys und Acritophiomys.[8] Auf Gaudeamus entfallen mehrere zerdrückte Schädel zuzüglich Schädelteilen, des Weiteren Unterkiefer nebst Fragmenten und isolierten Zähnen.[2] Zusätzliche Fossilreste wurden an den Fundstellen A und B entdeckt, die stratigraphisch rund 12 m oberhalb von L-41 liegen und aus eher gröberen, durch Flüsse zusammengetragenen Sedimenten bestehen. Sie datieren in das Untere Oligozän. Die hier gemachten Funde von Gaudeamus, mehrere Unterkiefer, wurden bereits in den 1960er Jahren geborgen, sie bildeten die Basis für die wissenschaftliche Benennung der Gattung.[1] Forschungsgeschichtlich älter ist ein nicht genau lokalisierter Unterkiefer, der von Max Schlosser im Jahr 1911 bei der Aufarbeitung von Fundmaterial deutscher Expeditionen in das Fayyum vorgestellt wurde. Schlosser verwies ihn damals aber zu Phiomys,[9] seine tatsächliche Natur wurde erst in den 1960er Jahren erkannt.[1][2] Alle aufgefundenen Nagetiere der Gebel-Qatrani-Formation repräsentieren momentan Vertreter der Hystricognathi. Andere Linien der Nagetiere wie die Dornschwanzhörnchen kamen im Fayyum zum gegenwärtigen Stand nur in der älteren Qasr-el-Sagha-Formation zum Vorschein, hier fehlen allerdings Nachweise der Phiomorpha.[8]
Außerhalb des Fayyum sind Reste von Gaudeamus von der Fundstelle Zallah nahe der gleichnamigen Oase im Sirte-Becken im zentralen Libyen berichtet worden. Der Fundstellenkomplex stellt das Relikt eines ehemaligen Flussdeltas dar und datiert in das Untere Oligozän vor rund 32 Millionen Jahren. Gefunden wurden hier einzelne Zähne.[10] Möglicherweise ebenfalls zu der Gattung müssen isolierte Zähne aus der Samlat-Formation in Westsahara auf dem Festland gegenüber der Halbinsel von Dakhla gezählt werden. Die reichhaltigen Kleinvertebratenfunde waren in eine sandige Matrix eingebettet, die auf einen ehemaligen Ästuar schließen lassen. Er bestand zu Beginn des Unteren Oligozän.[11]
Systematik
Innere Systematik der Hystricognathi nach Sallam und Seiffert 2016[8]
Zur Verdeutlichung der stammesgeschichtlichen Position wurden die Caviomorpha und Hystricidae nach Sallam et al. 2011[2] nachgetragen |
Innere Systematik der Hystricognathi nach Marivaux und Boivin 2019[12]
|
Gaudeamus ist eine ausgestorbene Gattung aus der Gruppe der Hystricognathi innerhalb der Unterordnung der Stachelschweinverwandten (Hystricomorpha). Die sehr vielfältige Gruppe der Hystricognathi kommt sowohl in Eurasien als auch in Afrika und Amerika vor und bildet einen Teil der Ordnung der Nagetiere (Rodentia). Sie schließt unter anderem die Stachelschweine (Hystricidae), die Meerschweinchenverwandten (Caviomorpha) und die Phiomorpha ein. Als charakteristisches Kennzeichen gilt der besondere Bau des Schädels und des Unterkiefers, bei letzterem ist etwa die Stellung des Winkelfortsatzes von Bedeutung. Der Ursprung der Gruppe liegt möglicherweise in Asien, die ältesten Funde dort datieren in das Mittlere Eozän vor knapp 50 Millionen Jahren. Afrika und Amerika erreichten die Hystricognathi wohl im Übergang zum Oberen Eozän vor rund 40 Millionen Jahren.[13][5][8]
Innerhalb der Hystricognathi wird Gaudeamus in die eigene Familie der Gaudeamuridae gestellt und ist dort gegenwärtig ihr einziges Mitglied. Die genauen Verwandtschaftsverhältnisse sind zur Zeit noch ungeklärt, in der Vergangenheit wurden drei unterschiedliche Möglichkeiten diskutiert. Durch den charakteristischen Zahnbau mit den schräg verlaufenden und in ihrer Anzahl reduzierten Leisten zeigt Gaudeamus deutliche Ähnlichkeiten zu den Rohrratten (Thryonomyidae), die den Phiomorpha zugeordnet werden.[14] Einige Forscher sahen beide deshalb auch als näher verwandt an.[1] Allerdings bestehen markante Unterschiede zu den Rohrratten. So ist die Zahnstruktur bei Gaudeamus komplexer, zudem kam es zu einem Austausch der Prämolaren in der Individualentwicklung, so dass mit den Milch- und den dauerhaften Zähnen zwei Generationen vorliegen. Prinzipiell verfügen die Phiomorpha über einen reichhaltigen Fossilnachweis in Nordafrika, der bis in das Obere Eozän zurückreicht.[2] Andere Autoren vermuteten wiederum Gaudeamus in einer engeren Bindung zu den Meerschweinchenverwandten, da beispielsweise die sehr frühen Caviomorpha wie Eoincamys ebenfalls eine vergleichbare Zahngestaltung aufweisen.[10] Als problematisch erweisen sich die bisher generell jüngeren Altersdaten der Caviomorpha gegenüber Gaudeamus, ebenso wie ein mehrfaches Wanderungsgeschehen zwischen Afrika und Südamerika erforderlich wäre. Als eine dritte mögliche Verwandtschaftsgruppe kommen die Stachelschweine in Betracht. Ihr erster Fossilnachweis fällt allerdings in das Obere Miozän vor rund 11 Millionen Jahren, womit eine große zeitliche Lücke bestehen würde.[2] Möglicherweise handelt es sich bei der übereinstimmenden Zahnmorphologie von sowohl Gaudeamus und Eoincamys als auch den Rohrratten um eine konvergente Bildung. Molekulargenetische Analysen machen eine Nahverwandtschaft zu letzteren eher unwahrscheinlich. Diesen zufolge trennten sich die Phiomorpha im Oberen Eozän vor rund 36 Millionen Jahren von den Caviomorpha ab. Eine stärkere Diversifizierung der heutigen Phiomorpha fand aber erst im Zeitraum vom Oberen Oligozän bis zum Unteren Miozän statt. Demnach bildeten sich die Rohrratten und die Felsenratten (Petromuridae) als Schwestergruppen vor rund 18 Millionen Jahren heraus.[15] Die Stachelschweine hatten sich dagegen bereits vor etwa 39 Millionen Jahren von der gemeinsamen Linie gelöst. Zur Untermauerung oder Überprüfung der möglichen Verwandtschaftsbeziehungen bedarf es weitaus besserer Schädelreste, als sie momentan vorliegen.[15][2][16][8] Eine weitere phylogenetischen Studie aus dem Jahr 2019 sieht die Gaudeamuridae ebenfalls in der Nahverwandtschaft zu den Caviomorpha.[12]
Es sind vier Arten aus der Gattung Gaudeamus bekannt:[1][10][2]
- G. aegyptius Wood, 1968
- G. aslius Sallam, Seiffert & Simons, 2011
- G. hylaeus Sallam, Seiffert & Simons, 2011
- G. lavocati Coster, Benammi, Lazzari, Billet, Martin, Salem, Bilal, Chaimanee, Schuster, Valentin, Brunet & Jaeger, 2010
Die vier Arten unterscheiden sich weitgehend in den Merkmalen der Zähne, so sind die Molaren von G. aegyptius mit weniger Leisten ausgestattet als die der anderen Arten, bei denen zusätzlich noch das Mesoloph vorkommt. Größenunterschiede zwischen den Arten lassen sich kaum nachweisen. G. aegyptius, die Nominatform, ist an den Fundstellen A und B im Fayyum belegt und trat somit im Unteren Oligozän auf. G. aslius und G. hylaeus sind von der Fundstelle L-41 im gleichen Raum bekannt, die in das Oberen Eozän datiert. Von beiden Vertretern liegen möglicherweise auch Nachweise aus den etwas jüngeren Ablagerungen von Dakhla in Westsahara vor.[11] Als weitere Art konnte G. lavocati außerhalb des Fayyums an der unteroligozänen Fundstelle Zallah in Libyen dokumentiert werden.[10] Wahrscheinlich kommen noch weitere Arten vor, da einige Fossilreste des Fayyums abweichende Merkmale zeigen. Vorläufig wurden diese jedoch den bekannten Arten zugesprochen. Aus der gegenwärtigen Sicht hatten Gaudeamus und die Gaudeamuridae eine nur kurze stammesgeschichtliche Entwicklungsphase. Funde der Gattung oder Familie aus der gegenüber L-41 rund 3 Millionen Jahre älteren Fundstelle BQ-2 im Fayyum liegen nicht vor, ebenso fehlen Hinweise aus etwa zeitgleichen Fundstellen wie Dor el-Talha in Libyen. Dem gegenüber sind auch aus jüngeren Fundstellen keine Hinweise auf die Gaudeamuridae bekannt, wobei der Fossilbericht aus dem afrikanischen Oligozän eher spärlich ist.[10][2]
Die Gattung Gaudeamus wurde im Jahr 1968 von Albert E. Wood wissenschaftlich erstbeschrieben. Der Aufstellung gingen mehrere Jahre Forschungsarbeit im Fayyum unter Leitung von Elwyn L. Simons voraus. Das Fundmaterial, das Wood für seine neue Art verwendete, stammt aus den Fundstellen A und B der Gebel-Qatrani-Formation. Es bestand aus mehreren Unterkiefern und isolierten Zähnen. Davon bestimmte Wood einen linken Unterkieferast mit der Zahnreihe vom vierten Milchprämolaren bis zum zweiten Molaren zuzüglich eines noch nicht durchgebrochenen letzten Prämolaren als Holotypen (Exemplarnummer CGM 26920). Die lokalen Arbeiter, die während der Forschungsarbeiten vor Ort gruben, waren nur bedingt der englischen Sprache mächtig. So bezeichneten sie die bei den Grabungen entdeckten Unterkiefer (englisch jaw) mit joy (englisch für „Freude“). Aufgefundene Nagetierunterkiefer nannten sie dadurch allgemein joy mouse, übersetzt also „Freudenmaus“. Die Wortverbindung wurde später von Mary R. Dawson mit Gaudeamus latinisiert (der lateinische Ausspruch gaudeamus (zu deutsch „freuen wir uns“) ist vor allem von einem Studentenlied bekannt; Dawson deutete die Endsilbe nach lateinisch mūs für „Maus“ um). Wood übernahm dies für seine Erstbeschreibung.[1]
In seiner Erstbeschreibung ordnete Wood Gaudeamus in die Familie der Phiomyidae innerhalb der Phiomorpha ein. Als damals einzige Art definierte er G. aegyptium.[1] Später, 1978, verwies René Lavocat die Gattung zu den Thryonomyidae und sah sie damit als direkte Verwandte der Rohrratten an.[17] Die Ansicht wurde später verschiedentlich übernommen.[18] In ihrer Dissertationsarbeit von 1994 stufte Patricia A. Holroyd den Familienstatus für Gaudeamus als incertae sedis ein. Unter Berufung auf neuem Fossilmaterial von der Fundstelle L-41 im Fayyum kreierte sie mit G. hylaeus eine weitere Art. Da ihre Arbeit nie öffentlich publiziert wurde, blieb die Art inoffiziell,[18] einige Studien gaben aber den Namen wieder.[19] Erst im Jahr 2011 publizierten Hesham M. Sallam und Forscherkollegen eine gültige Artbeschreibung für G. hylaeus. Holroyds Art erwies sich in ihren Analysen aber als hochvariabel, weshalb sie mit G. aslius eine zusätzliche ausgliederten. Gleichzeitig schufen sie mit der von Gaudeamus abgeleiteten Bezeichnung Gaudeamuridae eine neue Familie, die sie aufgrund der deutlich abweichenden Merkmale außerhalb der Phiomorpha einstuften.[2]
Literatur
- Hesham M. Sallam, Erik R. Seiffert und Elwyn L. Simons: Craniodental Morphology and Systematics of a New Family of Hystricognathous Rodents (Gaudeamuridae) from the Late Eocene and Early Oligocene of Egypt. PLoS ONE 6 (2), 2011, S. e16525 doi:10.1371/journal.pone.0016525
- Albert E. Wood: The African Oligocene Rodentia. In: Elwyn L. Simons und Albert E. Wood (Hrsg.): Early Cenozoic Mammalian Faunas Fayum Province, Egypt. Bulletin of the Peabody Museum of Natural History 28, 1968, S. 23–105
Einzelnachweise
- Albert E. Wood: The African Oligocene Rodentia. In: Elwyn L. Simons und Albert E. Wood (Hrsg.): Early Cenozoic Mammalian Faunas Fayum Province, Egypt. Bulletin of the Peabody Museum of Natural History 28, 1968, S. 23–105
- Hesham M. Sallam, Erik R. Seiffert und Elwyn L. Simons: Craniodental Morphology and Systematics of a New Family of Hystricognathous Rodents (Gaudeamuridae) from the Late Eocene and Early Oligocene of Egypt. PLoS ONE 6 (2), 2011, S. e16525 doi:10.1371/journal.pone.0016525
- Albert E. Wood und R. W. Wilson: A Suggested Nomenclature for the Cusps of the Cheek Teeth of Rodents. Journal of Paleontology 10 (5), 1936, S. 388–391
- Hesham M. Sallam, Erik R. Seiffert und Elwyn L. Simons: A basal phiomorph (Rodentia, Hystricognathi) from the late Eocene of the Fayum Depression, Egypt. Swiss Journal of Palaeontology 131, 2012, S. 283–301
- Laurent Marivaux, El Mabrouk Essid, Wissem Marzougui, Hayet Khayati Ammar, Sylvain Adnet, Bernard Marandat, Gilles Marzeraud, Rodolphe Tabuce und Monique Vianey Liaud: A new and primitive species of Protophiomys (Rodentia, Hystricognathi) from the late middle Eocene of Djebel el Kébar, Central Tunisia. Palaeovertebrata 38 (1), 2014, S. e2
- Elwyn L. Simons: Description of two genera and species of Late Eocene Anthropoidea from Egypt. PNAS 86, 1989, S. 9956–9960
- D. Tab Rasmussen und Elwyn L. Simons: The oldest hyracoids (Mammalia: Pliohyracidae): new species of Saghatherium and Thyrohyrax from the Fayum. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 182, 1991, S. 187–209
- Hesham M. Sallam und Erik R. Seiffert: New phiomorph rodents from the latest Eocene of Egypt, and the impact of Bayesian "clock"-based phylogenetic methods on estimates of basal hystricognath relationships and biochronology. PeerJ 4, 2016, S. e1717 doi:10.7717/peerj.1717
- Max Schlosser: Beiträge zur Kenntnis der Oligozänen Landsäugetiere aus dem Fayum, Ägypten. Beiträge zur Paläontologie und Geologie Österreich-Ungarns und des Orients 24, 1911, S. 51–167 (S. 90–94 und Tafel 5 (13):7 und 7a) ()
- Pauline Coster, Mouloud Benammi, Vincent Lazzari, Guillaume Billet, Thomas Martin, Mustafa Salem, Awad Abolhassan Bilal, Yaowalak Chaimanee, Mathieu Schuster, Xavier Valentin, Michel Brunet und Jean-Jacques Jaeger: Gaudeamus lavocati sp. nov. (Rodentia, Hystricognathi) from the early Oligocene of Zallah, Libya: first African caviomorph? Naturwissenschaften 97 (8), 2010, S. 697–706
- Laurent Marivaux, Sylvain Adnet, Mohamed Benammi, Johan Yans und Mouloud Benammi: Earliest Oligocene hystricognathous rodents from the Atlantic margin of Northwestern Saharan Africa (Dakhla, Marocco): systematics, paleobiogeographical, and paleoenvironmental implications. Journal of Vertebrate Paleontology 37 (5), 2017, S. e1357567 doi:1080/02724634.2017
- Laurent Marivaux und Myriam Boivini: Emergence of hystricognathous rodents: Palaeogene fossil record, phylogeny, dental evolution and historical biogeography. Zoological Journal of the Linnean Society, 2019, S. 1–36
- Laurent Marivaux, Monique Vianey-Liaud, Jean-Loup Welcomme und Jean-Jaques Jaeger: The role of Asia in the origin and diversification of hystricognathous rodents. Zoologica Scripta 31, 2002, S. 225–239
- Albert E. Wood: The juvenile tooth pattern of certain African rodents. Journal of Mammalogy 43 (3), 1952, S. 310–322
- Hesham M. Sallam, Erik R. Seiffert, Michael E. Steiper und Elwyn L. Simons: Fossil and molecular evidence constrain scenarios for the early evolutionary and biogeographic history of hystricognathous rodents. PNAS 106 (39), 2009, S. 16722–16727 doi:10.1073/pnas.0908702106
- Frank Barbière und Laurent Marivuax: Phylogeny and evolutionary history of hystricognathous rodents from the Old World during the Tertiary: new insights into the modern "phiomorph" families. In: P. G. Cox und L. Hautier (Hrsg.): Evolution of the rodents: Advances in phylogeny, functional morphology and development. Cambridge University Press, 2015, S. 87–119
- René Lavocat: Rodentia and Lagomorpha. In: Vincent J. Maglio und H. B. S. Cooke (Hrsg.): Evolution of African Mammals. Harvard University Press, 1978, S. 69–89
- Alisa J. Winkler, Christiane Denys und D. Margaret Avery. Rodentia. In: Lars Werdelin und William Joseph Sanders (Hrsg.): Cenozoic Mammals of Africa. University of California Press, Berkeley, Los Angeles, London, 2010, S. 263–304
- Patrick J. Lewis und Elwyn L. Simons: Morphological trends in the molars of fossil rodents from the Fayum Depression, Egypt. Palaeontologica Africana 42, 2007, S. 37–42