Birkamys
Birkamys ist eine ausgestorbene Gattung aus der Gruppe der Stachelschweinverwandten. Innerhalb dieser wird sie als eine sehr ursprüngliche Form der Phiomorpha betrachtet, zu denen unter anderem die heutigen Rohrratten und Felsenratten gehören. Funde von Birkamys stammen aus der bedeutenden Fossillagerstätte des Fayyum-Gebiet im nördlichen Ägypten sowie von einzelnen anderen Fundpunkten in der Region. Es liegen überwiegend Teile des Schädels und des Gebisses vor, die allerdings stark fragmentiert überliefert sind. Die Tiere waren vergleichsweise klein, charakteristisch sind der an Stachelschweine erinnernde Schädel- und Unterkieferbau sowie eine große Öffnung am Gaumen. Als Besonderheit kam es wie bei einigen anderen heutigen Phiomorpha nicht zum Austausch der Vormahlzähne während der Individualentwicklung, so dass die Milchzähne dauerhaft im Gebiss verblieben. Die Gattung wurde im Jahr 2016 eingeführt.
| Birkamys | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Unterkiefer von Birkamys | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oberes Eozän bis Unteres Oligozän | ||||||||||||
| 34 bis 28 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Birkamys | ||||||||||||
| Sallam & Seiffert, 2016 | ||||||||||||
Merkmale
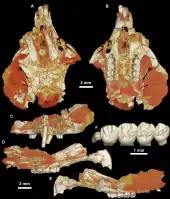
Birkamys ist ein kleiner Vertreter der Hystricognathi und einer der kleinsten bisher bekannten aus dem Paläogen des afro-arabischen Raumes. Es liegen hauptsächlich fragmentierte Schädel- und Gebissreste vor, von den bekannten Schädelfunden sind aber die meisten stark zerdrückt. Das Rostrum war lang und schmal. Der Mittelkieferknochen nahm den größten Teil des vorderen Bereiches ein und erstreckte sich bis zur Hälfte des Diastemas zwischen der vorderen und hinteren Bezahnung. Auf dem Gaumen befand sich eine große Öffnung, die aus dem Foramen incisivum entstanden war und den gesamten Raum bis zu den letzten beiden Prämolaren einnahm. Sie wird teilweise als „vorderes Gaumenfenster“ bezeichnet.[1] Wie bei anderen Angehörigen der Hystricognathi auch war das Foramen infraorbitale sehr groß und fungierte zusätzlich als Ankerpunkt für den Masseter-Muskel. Es lag etwa auf der Höhe des dritten Prämolaren. Direkt darunter erhob sich ein kleiner Höcker, seitlich davon setzte der vordere Jochbogenabschnitt an. An den bisher aufgefundenen Unterkiefern fehlt ein Teil des aufsteigenden Astes. Allgemein besaß er einen schlanken Bau. Durch den seitlich abstehenden, großen Winkelfortsatz wirkte er vollständig hystricognath. Die Unterkante des horizontalen Knochenkörpers war stark ausgebogen und wurde vollständig von der Alveole des ersten Schneidezahns eingenommen. Vom Winkelfortsatz setzten sich zwei massive Knochenrippen ab, die die Fossa masseterica oben und unten einrahmten und ebenfalls dem Masseter-Muskel als Ansatzstelle dienten. Das Foramen mentale besaß eine ovale Form und öffnete sich etwa auf Höhe des letzten Prämolaren. Der Gelenkfortsatz überragte die Kaufläche der Molaren nur wenig, war aber wiederum niedriger als der Kronenfortsatz. Das Foramen mandibulae saß im Bereich zwischen dem Gelenk- und dem Kronenfortsatz.[2]
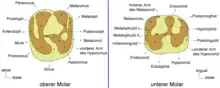
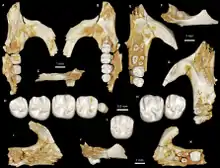
Das Gebiss setzte sich je Kieferbogen aus einem Schneidezahn, zwei (oben) beziehungsweise einem (unten) Prämolaren und drei Molaren zusammen. Die Schneidezähne waren als typische Nagezähne ausgebildet, der untere hatte einen ovalen Querschnitt mit seitlichen Verschmälerungen. Zahnschmelz war nur auf der Vorderseite ausgebildet. Zur hinteren Bezahnung bestand ein weites Diastema, das im Unterkiefer die Länge des halben horizontalen Knochenkörpers einnahm und tief eingedellt war. Mit Hilfe computertomographischen Untersuchungen konnte belegt werden, dass bei Birkamys die Milchprämolaren dauerhaft im Gebiss bestanden und somit nicht im Verlauf der Individualentwicklung ausgewechselt wurden. Das Merkmal ist auch von einigen heutigen Phiomorpha bekannt. Andere frühe Phiomorpha wie etwa Turkanamys oder Acritophiomys wiesen allerdings noch einen Zahnwechsel bei den Vormahlzähnen auf. Die Backenzähne der Nagetiere haben einen komplexen Aufbau mit mehreren Buckeln und dazwischen verlaufenden Leisten oder Jochen (Lophen genannt), die einzelnen Formen unterscheiden sich durch die jeweilige Anzahl an Höckern und Leisten.[3][4][5] Der vordere obere Prämolar (dp3) war klein und nagelartig, der letzte Prämolar (dp4) hatte oben einen quadratischen Umriss und vier Höckerchen, unten einen rechteckigen Umriss und fünf Höckerchen auf der Kauoberfläche. Die oberen Molaren zeigten sich alle mehr oder weniger rechteckig geformt. Auch sie wiesen vier Höckerchen auf (Para-, Meta-, Proto- und Hypoconus), die paarig zusammenstanden und durch rechtwinklig zur Zahnlängsachse orientierte Leisten miteinander verbunden waren. Zwischen dem Para- und Protoconus verlief demnach das Protoloph, zwischen dem Meta- und Hypoconus das Hypoloph. Seitlich außen am Zahnrand erstreckte sich jeweils noch eine weitere Leiste, das Anteroloph vor dem Protoloph und das Posteroloph hinter dem Metaloph. Dadurch wirkten die oberen Mahlzähne tetralophodont (mit vier Leisten). An den unteren Molaren saßen ebenfalls vier große Höcker (Meta-, Ento-, Proto- und Hypoconid) paarig angeordnet, ein fünfter Höcker, das Hypoconulid, erhob sich am hinteren Zahnrand. Die zungenseitigen Höcker waren kleiner als die lippenseitigen. Zwischen den Höckern verliefen mit dem Metalophulid (zwischen dem Meta- und Protoconid), dem Hypolophid (zwischen dem Hypo- und Entoconid) und dem Posterolophid (am Hypoconulid) insgesamt drei Leisten. Ein zusätzliches zweites Metalophulid zwischen dem eigentlichen Metalophulid und dem Hypolophid war bei Birkamys nur rudimentär und unvollständig ausgebildet, anders als etwa bei Phiomys und Acritophiomys mit einer kräftigen und vollständigen zusätzlichen Leiste; zudem besaß Acritophiomys mit dem Mesolophid noch eine weitere Leiste auf dem vordersten Molar. Im oberen Gebiss war der erste Molar am größten ausgebildet, seine Ausmaße betrugen 1,07 mm in der Länge und 1,13 mm in der Breite. Dagegen stellte im unteren Gebiss der zweite Molar den größten Zahn der hinteren Zahnreihe dar. Er wurde 1,1 mm lang und 1,06 mm breit. Der jeweils hinterste Molar war relativ klein, was von Phiomys abweicht.[2]
Fossilfunde
Umfangreiches Fundmaterial von Birkamys kam im Fayyum-Gebiet im nördlichen Ägypten in Nordafrika zu Tage und stammt dort von der Fundstelle L-41 im unteren Abschnitt der Gebel-Qatrani-Formation. Die Fundstelle liegt etwa 46 m oberhalb der Basis der Gesteinseinheit und datiert paläomagnetischen Untersuchungen zufolge in das Obere Eozän, die absoluten Altersangaben betragen um die 34 Millionen Jahre. L-41 stellt die älteste Fundstelle innerhalb der Formation dar. Entdeckt wurde sie im Jahr 1983 bei Felduntersuchungen durch ein Forscherteam um Elwyn L. Simons, eine genauere Vorstellung folgte dann später unter anderem durch D. Tab Rasmussen. Sie besteht aus grünlich bis grünlich grau gefärbten Ton- und Schluffsteinen. Diese sind wiederum in eine Sequenz aus gebänderten Sanden eingebettet. Die feinkörnigen Ablagerungen können als Relikte eines ehemaligen Sees aufgefasst werden. Charakteristisch sind ein hoher Anteil an Evaporiten und eine starke Konzentration an Natriumchlorid. In ihrer gesamten Struktur weicht die Fundstelle dadurch auffallend von den anderen, eher gröberklastischen, fluviatilen Ablagerungen der Gebel-Qatrani-Formation ab. Als weitere Besonderheit kann eine hohe Anzahl an Schädeln mit teils assoziierten Partien des Körperskeletts hervorgehoben werden. L-41 lieferte bisher Fossilreste von Fischen, Amphibien, Reptilien, Vögeln und Säugetieren. Unter anderem ist die hohe Vielfalt an Schliefern, Primaten, Nagetieren und Fledertieren bemerkenswert. Aufgrund der guten Erhaltung von Überresten kleinerer Wirbeltiere hat die Fundstelle L-41 für die Fayyum-Region eine hohe Bedeutung. Die Nagetiere können hunderte Skelettelemente vorweisen, darunter neben Fossilien von Birkamys auch solche von Mubhammys, Acritophiomys und Gaudeamus. Auffallenderweise gehören bisher alle Reste zu den Hystricognathi, nur aus der älteren Qasr-el-Sagha-Formation im Fayyum sind auch Reste von Dornschwanzhörnchen belegt. Allerdings fehlen hier Nachweise der Phiomorpha. Von Birkamys sind hauptsächlich Teile des Schädels, Unterkiefers und des Gebisses gefunden worden.[6][7][4][2]
Abseits des Fayyums ist Birkamys möglicherweise aus der Samlat Formation am äußersten Westrand der Sahara nahe der Stadt Ad-Dakhla in Westsahara dokumentiert. Die dortigen sandigen Ablagerungen sind Hinterlassenschaften eines Ästuars, der zu Beginn des Unteren Oligozän vor etwa 33 Millionen Jahren bestand. Die überwiegend isolierten Zähne sind aber im Vergleich zu denen aus dem Fayyum durchschnittlich etwas größer, weswegen der Verweis zu Birkamys nicht eindeutig ist.[8]
Systematik
Innere Systematik der Hystricognathi nach Sallam und Seiffert 2016[2]
Zur Verdeutlichung der stammesgeschichtlichen Position wurden die Caviomorpha und Hystricidae nach Sallam et al. 2011[9] nachgetragen |
Innere Systematik der Hystricognathi nach Marivaux und Boivin 2019[10]
|
Birkamys ist eine ausgestorbene Gattung aus der Gruppe der Hystricognathi innerhalb der Unterordnung der Stachelschweinverwandten (Hystricomorpha). Die Hystricognathi bilden eine vielfältige Gruppe eurasischer, afrikanischer und amerikanischer Vertreter der Nagetiere (Rodentia), die unter anderem die Stachelschweine (Hystricidae), die Meerschweinchenverwandten (Caviomorpha) und die Phiomorpha einschließen. Charakteristische Kennzeichen finden sich vor allem im Bau des Schädels und des Unterkiefers, bei letzterem etwa durch die Stellung des Winkelfortsatzes. Die gesamte Gruppe hat ihren Ursprung möglicherweise in Asien, wo sie bereits im Mittleren Eozän vor knapp 50 Millionen Jahren nachgewiesen ist. Die heutigen afrikanischen und amerikanischen Verbreitungsgebiete erreichten die ersten Einwanderer am Übergang zum Oberen Eozän vor rund 40 Millionen Jahren.[11][5][2]
Durch die Gestaltung der Backenzähne mit drei beziehungsweise vier Leisten gehört Birkamys zu den entwickelten Vertretern der Hystricognathi, deren ursprüngliches Zahnmuster mehr Leisten umfasste. Dies drückt sich unter anderem durch die Reduktion der mittleren Leiste aus, dem Mesoloph/Mesolophid, was unter anderem charakteristisch für die Phiomorpha ist.[12][11][13] Hinzu kommt der Verbleib der Milchprämolaren im Gebiss bei Birkamys, was ebenfalls mit einigen anderen Phiomorpha übereinstimmt. Dadurch kann eine nähere Beziehung zu den heutigen Rohrratten (Thryonomyidae), Felsenratten (Petromuridae) und Sandgräbern (Bathyergidae) angenommen werden. Die Ansicht ist kongruent zu molekulargenetischen Analysen, nach denen sich die Phiomorpha im Oberen Eozän vor rund 36 Millionen Jahren von den Caviomorpha abgespalten hatten, während die Hystricidae bereits zuvor einen eigenständigen evolutiven Weg beschritten.[14] Als nächster Verwandter wird phylogenetischen Studien zufolge Mubhammys angesehen, deren Fossilreste ebenfalls im Fayyum-Gebiet gefunden wurden. Eine genaue Zuweisung zu einer bestimmten Familie innerhalb der Phiomorpha erfolgte bisher noch nicht. Die beiden Gattungen bilden die Schwestergruppe zu den späteren Phiomorpha einschließlich der rezenten Formen.[14][9][2]
Nach dieser Ansicht gehört Birkamys neben Mubhammys und Acritophiomys zu den ältesten bekannten Vertretern der Phiomorpha. Die Entdeckung von Mubhammys und Birkamys zeigt an, dass die Unterdrückung des permanenten Prämolaren als Merkmal bei den Phiomorpha bereits in der Wende vom Eozän zum Oligozän und somit stammesgeschichtlich sehr früh einsetzte. Außerdem belegt sie, dass das außerordentliche große Gaumenfenster ein inklusives Charakteristikum verschiedener früher Pliomorpha darstellt und somit ebenfalls sehr früh in der Entwicklung der Hystricognathi auftrat. Die geringe Größe von Birkamys lässt annehmen, dass es innerhalb der Gruppe im Verlauf des Eozäns zu rapiden Größenveränderungen einschließlich Verzwergungen gekommen sein muss.[2] Laut einer phylogenetischen Studie aus dem Jahr 2019 ist aber auch eine Stellung von Birkamys gemeinsam mit Mubhammys in einem Verwandtschaftskreis um Phiocricetomys möglich. Die gemeinsame Gruppe wird als "phiocricetomyins" bezeichnet und charakterisiert sich über extrem niederkronige Backenzähne mit flachen Querleisten. Ursprünglich als Mitglieder der Myophiomyidae betrachtet, ordnet die Untersuchung die "phiocricetomyins" außerhalb der Phiomorpha als basale Hystricognathi ein, die Unterdrückung der permanenten Prämolaren wäre demnach mehrfach innerhalb der Stachelschweinverwandten entstanden.[10]
Die Gattung Birkamys wurde im Jahr 2016 von Hesham M. Sallam und Erik R. Seiffert wissenschaftlich erstbeschrieben. Die Basis dafür bildete das Fundmaterial aus der Fundstelle L-41 im Fayyum-Gebiet in Ägypten. Der Holotyp (Exemplarnummer CGM 66000) ist ein Teilschädel bestehend aus dem Rostrum, in dem noch die Schneidezähne sowie beide Zahnreihen vom dritten Prämolaren bis zum dritten Molaren erhalten sind. Der Name Birkamys leitet sich vom arabischen Wort birka für „Sumpf“ oder „See“ und vom griechischen Wort μῦς (mŷs) für „Maus“ ab. Birka bezieht sich dabei auf die Fundstelle L-41. Einzige bekannte Art ist B. korai. Das Artepitheton ehrt Mahmoud Kora für seine Beiträge zur Paläontologie und Stratigraphie Ägyptens.[2] Einige Forscher vermuten, dass Birkamys synonym zu Neophiomys ist, einem frühen Vertreter phiocricetomyiner Nagetiere. Allerdings bedarf es zur Bestätigung der Annahme an aufschlussreicherem Fossilmaterials von Neophiomys.[10]
Literatur
- Hesham M. Sallam und Erik R. Seiffert: New phiomorph rodents from the latest Eocene of Egypt, and the impact of Bayesian "clock"-based phylogenetic methods on estimates of basal hystricognath relationships and biochronology. PeerJ 4, 2016, S. e1717 doi:10.7717/peerj.1717
Einzelnachweise
- Albert E. Wood: The African Oligocene Rodentia. In: Elwyn L. Simons und Albert E. Wood (Hrsg.): Early Cenozoic Mammalian Faunas Fayum Province, Egypt. Bulletin of the Peabody Museum of Natural History 28, 1968, S. 23–105
- Hesham M. Sallam und Erik R. Seiffert: New phiomorph rodents from the latest Eocene of Egypt, and the impact of Bayesian "clock"-based phylogenetic methods on estimates of basal hystricognath relationships and biochronology. PeerJ 4, 2016, S. e1717 doi:10.7717/peerj.1717
- Albert E. Wood und R. W. Wilson: A Suggested Nomenclature for the Cusps of the Cheek Teeth of Rodents. Journal of Paleontology 10 (5), 1936, S. 388–391
- Hesham M. Sallam, Erik R. Seiffert und Elwyn L. Simons: A basal phiomorph (Rodentia, Hystricognathi) from the late Eocene of the Fayum Depression, Egypt. Swiss Journal of Palaeontology 131, 2012, S. 283–301
- Laurent Marivaux, El Mabrouk Essid, Wissem Marzougui, Hayet Khayati Ammar, Sylvain Adnet, Bernard Marandat, Gilles Marzeraud, Rodolphe Tabuce und Monique Vianey Liaud: A new and primitive species of Protophiomys (Rodentia, Hystricognathi) from the late middle Eocene of Djebel el Kébar, Central Tunisia. Palaeovertebrata 38 (1), 2014, S. e2
- Elwyn L. Simons: Description of two genera and species of Late Eocene Anthropoidea from Egypt. PNAS 86, 1989, S. 9956–9960
- D. Tab Rasmussen und Elwyn L. Simons: The oldest hyracoids (Mammalia: Pliohyracidae): new species of Saghatherium and Thyrohyrax from the Fayum. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 182, 1991, S. 187–209
- Laurent Marivaux, Sylvain Adnet, Mohamed Benammi, Johan Yans und Mouloud Benammi: Earliest Oligocene hystricognathous rodents from the Atlantic margin of Northwestern Saharan Africa (Dakhla, Marocco): systematics, paleobiogeographical, and paleoenvironmental implications. Journal of Vertebrate Paleontology 37 (5), 2017, S. e1357567 doi:1080/02724634.2017
- Hesham M. Sallam, Erik R. Seiffert und Elwyn L. Simons: Craniodental Morphology and Systematics of a New Family of Hystricognathous Rodents (Gaudeamuridae) from the Late Eocene and Early Oligocene of Egypt. PLoS ONE 6 (2), 2011, S. e16525 doi:10.1371/journal.pone.0016525
- Laurent Marivaux und Myriam Boivini: Emergence of hystricognathous rodents: Palaeogene fossil record, phylogeny, dental evolution and historical biogeography. Zoological Journal of the Linnean Society, 2019, S. 1–36
- Laurent Marivaux, Monique Vianey-Liaud, Jean-Loup Welcomme und Jean-Jaques Jaeger: The role of Asia in the origin and diversification of hystricognathous rodents. Zoologica Scripta 31, 2002, S. 225–239
- René Lavocat: Rodentia and Lagomorpha. In: Vincent J. Maglio und H. B. S. Cooke (Hrsg.): Evolution of African Mammals. Harvard University Press, 1978, S. 69–89
- Pauline Coster, Mouloud Benammi, Vincent Lazzari, Guillaume Billet, Thomas Martin, Mustafa Salem, Awad Abolhassan Bilal, Yaowalak Chaimanee, Mathieu Schuster, Xavier Valentin, Michel Brunet und Jean-Jacques Jaeger: Gaudeamus lavocati sp. nov. (Rodentia, Hystricognathi) from the early Oligocene of Zallah, Libya: first African caviomorph? Naturwissenschaften 97 (8), 2010, S. 697–706
- Hesham M. Sallam, Erik R. Seiffert, Michael E. Steiper und Elwyn L. Simons: Fossil and molecular evidence constrain scenarios for the early evolutionary and biogeographic history of hystricognathous rodents. PNAS 106 (39), 2009, S. 16722–16727 doi:10.1073/pnas.0908702106