Gastornis
Gastornis ist eine ausgestorbene Gattung großer, flugunfähiger Vögel aus der näheren Verwandtschaft der Gänsevögel. Sie war vom Mittleren Paläozän bis zum Mittleren Eozän vor 62 bis 43 Millionen Jahren sowohl im heutigen Eurasien als auch in Nordamerika verbreitet. Die ersten Fossilien der Gattung Gastornis wurden 1855 nahe Paris entdeckt und noch im selben Jahr erstbeschrieben. Die ältesten nordamerikanischen Funde stammen aus dem Jahr 1876 und wurden ursprünglich zur Gattung Diatryma gestellt. Über lange Zeit galten beide Gattungen als eigenständig, wofür zum Teil eine fehlerhafte Rekonstruktion aus dem Ende des 19. Jahrhunderts verantwortlich war. Erst Anfang der 1990er Jahre wurde erkannt, dass Diatryma und Gastornis nahe miteinander verwandt sind, neue Knochenfunde aus Frankreich führten schließlich zu Synonymisierung von Diatryma mit Gastornis.
| Gastornis | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelettrekonstruktion von Gastornis | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Seelandium (Paleozän) bis Lutetium (Eozän) | ||||||||||||
| 61,6 bis 43,4 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Gastornis | ||||||||||||
| Hébert, 1855 | ||||||||||||
Über lange Zeit wurde Gastornis als wendiger, auf Fleischnahrung spezialisierter Beutegreifer angesehen, der vor allem Jagd auf kleinere Säugetiere machte und in Verbindung mit seiner Größe, die bis knapp über 2 m betrug, einen Spitzenprädator seiner Zeit repräsentierte. Die Ansicht ging einher mit einem den südamerikanischen „Terrorvögeln“ (Phorusrhacidae) ähnelnden Skelettbau, der anhand eines nahezu vollständigen, 1916 entdeckten Skelettfundes aus Wyoming, erarbeitet wurde. Teilweise beruhte diese Ähnlichkeit aber auf fehlerhaften Annahmen zur Länge einzelner Knochen im Fußskelett. Erst in den 1970er Jahren konnte durch Neufunde von Fußknochen das Bild des wendigen Läufers korrigiert werden. Demnach stellte Gastornis aufgrund des Körperbaus einen sich eher langsam fortbewegenden, bodenbewohnenden Vogel dar. Die Ernährungsweise war lange Zeit ungeklärt. Neben der Spezialisierung auf Fleischnahrung wurde immer wieder eine rein pflanzliche Ernährung diskutiert, die sich vor allem aus anatomischen Erwägungen wie der Größe des Tiers und dem Bau des Schnabels ergab. Mehrere unabhängige Untersuchungen aus dem Jahr 2013 bestätigen vorerst die Ansicht der Spezialisierung auf Pflanzennahrung.
Merkmale
Habitus

Gastornis war ein sehr großer und robust gebauter, flugunfähiger Vogel, der eine Scheitelhöhe von 1,75 bis etwas über 2 m erreichte. Dabei waren europäische Vertreter im Durchschnitt etwas kleiner als nordamerikanische.[1][2] Das Körpergewicht betrug für die frühen Formen in Europa schätzungsweise etwa 100 kg, bei späteren zwischen 135 und 156 kg, teilweise auch bis 180 kg. Gewichtsspannen für die durchschnittlich größeren Individuen aus Nordamerika reichen von 160 bis 229 kg. Damit war er einer der größten und schwersten bekannten Vögel.[3][4][5] Besondere Merkmale der Vertreter von Gastornis stellten der große Kopf mit einem mächtigen Schnabel, der kurze, aber kräftige Hals und die massiven Beine dar, deren untere Abschnitte eher kurz gestaltet waren. Wie bei den heutigen großen flugunfähigen Vögeln wiesen die Flügel in ihrer Länge stark reduzierte Knochen auf.[6]
Schädel und Schnabel

Der Schädel war sehr groß und maß bei einem vollständigen Exemplar 43 cm, der mächtige Schnabel nahm davon 23 cm ein. Dieser war zudem sehr hoch, insgesamt 16,5 cm, und seitlich stark verschmälert. Die Schnabeloberseite wies eine deutlich konvexe Krümmung auf, während die Schnabelspitze nicht hakenartig verlängert war. Vor allem durch letzteres Merkmal unterscheidet sich Gastornis deutlich von den ähnlich gebauten Phorusrhacidae („Terrorvögel“). Die äußeren Nasenlöcher befanden sich etwa 5 cm vor der Orbita und nahe der Unterkante des Schnabels. Sie waren klein und nach vorn gerichtet. Auch hier bestehen Unterschiede zu den Phorusrhaciden, deren Nasenlöcher häufig direkt am Augenfenster ansetzen. Die Orbita selbst war von einem kräftigen und prominent erhöhten Wulst umgeben. Das für Vögel typische Quadratbein zeichnete sich durch seine Größe aus, war aber insgesamt eher kurz und gedrungen. Die Jochbeinbögen wiesen ebenfalls einen kurzen Bau auf und verliefen in einer S-Kurve. Der gesamte, hinter den Augenfenstern ansetzende Hinterschädel besaß im Gegensatz zum massiven Schnabel nur eine geringe Ausdehnung.[6][7]
Der Unterkiefer war massiv, bis zu 38 cm lang[8] und am Knochenkörper mit bis zu 10 cm sehr hoch. Beide Unterkieferhälften waren durch die Symphyse im vorderen Bereich fest miteinander verwachsen, das vordere Ende lief spitz aus. Die Verbindung reichte teils über eine Länge von 18 cm und nahm so fast die Hälfte der Gesamtlänge des Unterkiefers ein, was ungewöhnlich ist für Vögel.[6][7]
Körperskelett
Die Wirbelsäule ist nicht vollständig bekannt, allgemein zeichneten sich die Wirbel aber durch ihre Robustheit und ihren kurzen Wirbelkörper aus. Der Hals wurde aus 13 oder 14 Wirbeln gebildet. Am Rücken befanden sich sieben rippentragende Wirbel. Die dort ansetzenden Rippen waren dünn und weit gebogen. Der Schwanz umfasste zehn oder mehr Wirbel. Die Vordergliedmaßen waren stark verkümmert. Typisch für flugunfähige Vögel war der stumpfe Winkel zwischen Schulterblatt und Rabenbein, der 180° erreichte, ähnlich wie bei den zweibeinig laufenden Dinosauriern mit verkürzten Vordergliedmaßen, aber im Gegensatz zu den flugfähigen Vögeln, bei denen der Winkel unter 90° liegt.[9] Die Hinterbeine dagegen waren äußerst kräftig ausgebildet. Der Oberschenkelknochen erreichte über 38 cm Länge und war am unteren Gelenkende gut 10 cm breit. Der Tibiotarsus maß fast 60 cm und wies einen geraden Schaft auf. Das Wadenbein wirkte demgegenüber sehr grazil. Mit rund 25 cm Länge sehr kurz und eher breit war der Tarsometatarsus gestaltet. Die Beine endeten in vierstrahligen Füßen, von denen drei Strahlen nach vorn (II bis IV) ragten und einer (I) nach hinten stand. Von den drei nach vorn weisenden Zehen besaß der mittlere (III) die größte Länge, was unterschiedlich zu den ebenfalls riesigen Moas mit drei gleich langen Strahlen ist. Der kürzeste Strahl war der nach hinten zeigende. Die Endphalangen wiesen eine nur kurze Form und einen dreieckigen Querschnitt auf. In der Längsansicht waren sie nur wenig gekrümmt, was sie wiederum von den Phorusrhaciden unterscheidet.[6][7][2]
Biogeographie
Fossilfunde

Gastornis ist sowohl in Europa als auch in Nordamerika nachgewiesen. Die europäischen Funde datieren in die Zeit vom Mittleren Paläozän bis zum Mittleren Eozän vor 62 bis etwa 41 Millionen Jahren. Zu den ältesten Nachweisen gehören die spärlichen Überreste von Walbeck im westlichen Sachsen-Anhalt, die in das Seelandium vor rund 60 Millionen Jahren datieren. Unter ihnen befindet sich unter anderem ein 7 cm langes Rabenbein.[10] Die umfangreichsten Funde des Kontinents sind aus dem Geiseltal bekannt. Sie umfassen mehr als drei Dutzend Fundobjekte, darunter größtenteils Reste der Laufbeine und des Beckens, aber auch Flügelknochen und Teile des Schädels wie dem Unterkiefer. Sie können insgesamt neun Individuen zugewiesen werden und verteilen sich über die Unterkohle bis zur Oberen Mittelkohle der Fundstelle, datieren somit ins Mittlere Eozän vor rund 47 bis 43 Millionen Jahren. Die Funde der oberen Mittelkohle sind gleichzeitig die jüngsten Gastornis-Funde überhaupt.[11][1][2] Gleich alt zur Geiseltaler Unterkohle ist ein singulärer Fund in Form eines etwa 30 cm langen Oberschenkelknochens aus der Grube Messel in Hessen, der sich in einer Gesteinsknolle befand. Er war bereits Anfang des 20. Jahrhunderts während des Bergbaubetriebs gefunden worden, konnte aber erst in den 1960er Jahren sicher zugewiesen werden.[12][13] Dagegen sind vom Mont-de-Berru bei Reims im Ostteil des Pariser Becken wiederum zahlreiche Reste bekannt. Sie sind etwas jünger als jene von Walbeck und repräsentieren unter anderem zahlreiche Laufbeinreste.[14] Ein Unterkiefer von der gleichen Lokalität ist mit 31 cm Länge noch deutlich kleiner ist als der späterer Vertreter.[8] Aus der gleichen Region und ebenfalls dem ausgehenden Paläozän angehörend kamen Funde, vor allem Reste des Bewegungsapparates, aus Louvois zu Tage.[15] Zu den südlichsten europäischen Resten sind des Weiteren einige Beinknochen, Wirbel und ein 18 cm langer und 17 cm hoher Rest eines Oberschnabels aus Saint-Papoul im Département Aude im südlichen Frankreich zu rechnen.[16] Darüber hinaus fanden sich Fossilien von Gastornis in London und in Belgien.[17][4][5]
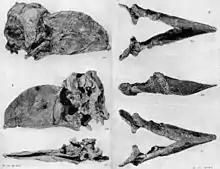
Die bekannten nordamerikanischen Funde sind alle dem Unteren Eozän vor 56 bis 49 Millionen Jahren zuzuweisen und verteilen sich auf über 50 Fundstellen.[9] Hervorzuheben ist dabei eines der wenigen vollständigen Skelette, das rekonstruiert bis zu 2,1 m hoch ist. Dieses stammt aus dem Bighorn-Becken im US-Bundesstaat Wyoming. Es wurde bereits Anfang des 20. Jahrhunderts entdeckt und ist das Resultat zahlreicher vorangegangener, tiefgründiger Untersuchungen, bei denen schon zuvor einige Bein- und Fußknochen zum Vorschein gekommen waren.[6] Aus dem gleichen Becken stammt aus der Willwood-Formation eine größere Kollektion, die auch Schädel- und Beinreste sowie Wirbelfunde beinhaltet.[7] Weitere Funde kamen darüber hinaus in New Mexico zum Vorschein. Hier ist vor allem ein Tarsometatarsus von Bedeutung, der im Jahr 1876 zur Aufstellung der Gattung Diatryma führte.[18] Wenige Knochenreste konnten auch aus der untereozänen Margaret-Formation auf Ellesmere Island im arktischen Norden Kanadas beschrieben werden. Diese stellen die nördlichsten bekannten Funde von Gastornis überhaupt dar.[19][20] Aus Asien sind nur wenige Reste bekannt, die ebenfalls ein untereozänes Alter aufweisen. Hierzu gehört ein unteres Ende eines großen Tibiotarsus aus der Yuhuangding-Formation in der chinesischen Provinz Henan. Dieser wurde 1980 als zur nahe mit Gastornis verwandten Gattung Zhongyuanus beschrieben,[21] eine Revision im Jahr 2013 ergab aber keine bedeutenden morphologischen Unterschiede zu Gastornis.[22][5]
Weichteile, Eier und Spurenfossilien
Neben fossilen Knochenresten sind nur wenige Überbleibsel des Weichteilgewebes bekannt. Bereits in den 1920er Jahren waren Federfunde vom Loan Creek im westlichen US-Bundesstaat Colorado beschrieben worden, die gut 20 cm lang waren und aus der Green-River-Formation stammten.[23] Diese erwiesen sich nach genaueren Untersuchungen als Reste von Pflanzenfasern.[24] Tatsächliche Federn in Form unter anderem eines 24 cm langen Exemplars, kamen dagegen im Fossil Butte Member, ebenfalls der Green-River-Formation zugehörig, zu Tage. Da aber weitere fossile Überreste fehlen, können sie aufgrund ihrer Größe nur bedingt Gastornis zugewiesen werden.[25]
Aus der Provence und aus Languedoc im südöstlichen und südlichen Frankreich wurden in Ablagerungen des Oberen Paläozäns (Thanetium) und des Unteren Eozäns (Sparnacium), zahlreiche fragmentierte Eier gefunden, deren Entdeckung bis in die 1950er Jahre zurückreicht. Die Eierschalen waren bis zu 3,1 mm dick. Einige wenige, teils vollständig erhaltene Eier erreichten Dimensionen von 24 × 15 cm, womit sie größer als jene des Afrikanischen Straußes (Struthio) aber kleiner als die der madagassischen Elefantenvögel (Aepyornis) sind. Einige andere Eier besaßen sogar Durchmesser von bis zu 40 cm. Anhand der Kurvatur großer Schalenfragmente konnte für einzelne Eier ein Volumen von bis zu 1300 cm³ berechnet werden, das ursprünglich Gewicht betrug wohl 1,4 kg. Da die Eierschalen von unterschiedlicher Größe und Struktur sind, werden sie als Hinterlassenschaften verschiedener Vogelarten gedeutet, die, da kein weiteres Fossilmaterial vorliegt, dem Oogenus Ornitholithus zugewiesen werden, wobei Ornitholithus arcuatus die größere Art darstellt und mit Gastornis übereinstimmen könnte. Die nächstgelegene Fundstelle mit Gastornis-Resten liegt mit Saint-Papoul nur einige Dutzend Kilometer entfernt. Allerdings ist es durchaus möglich, dass auch andere frühe Vögel, etwa der straußartige Palaeotis, einen Teil der Eier gelegt haben könnten.[17] Weitere Funde von Eierschalen, die dem gleichen Oogenus zugewiesen werden, stammen aus der Tremp-Formation im nordöstlichen Spanien.[4][5]
Zu den ebenfalls seltenen Funden gehören Spurenfossilien. In der Chuckanut-Formation im Nordwesten des US-Bundesstaates Washington wurden in untereozänen Sandsteinlagen eingetiefte Trittsiegel eines großen Vogels mit drei nach vorn weisenden Zehen gefunden. Die größten Siegel haben Längen von 28,5 und Breiten von 24,5 cm. Aufgrund der Größe der Fußspuren kann Gastornis als Verursacher angesehen werden. Da keine weiteren Fossilien vorliegen, werden sie aber der Ichnospezies Rivavipes giganteus zugewiesen. Bereits zuvor war aus dem King County des gleichen Bundesstaates über eine ähnliche, aber singuläre Spur berichtet worden, die dem Mittleren bis Oberen Eozän angehört und damit außerhalb der zeitlichen Reichweite von Gastornis in Nordamerika liegt. Heute wird diese Spur als Pseudofossil angesehen.[26] Auch aus Europa, aus Gipsablagerungen von Montmorency nördlich von Paris, wurden fossile Spuren großer Vögel bekannt und mit Gastornis in Verbindung gebracht. Diese gehören allerdings dem Oberen Eozän an, wofür es keine Nachweise dieser Vogelgattung in Europa gibt.[3][4]
Paläobiologie
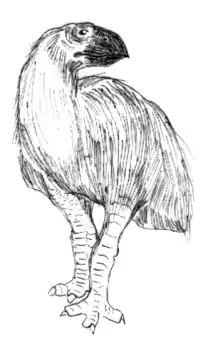
Ernährungsweise
Die Ernährungsweise von Gastornis war und ist Gegenstand wissenschaftlicher Kontroversen. Bereits sehr früh schlossen Wissenschaftler aus dem mächtigen und sehr kräftigen Schnabel, dass es sich bei Gastornis vermutlich um einen Fleischfresser gehandelt haben muss. Dieser entwickelte sich konvergent zu den südamerikanischen, heute ebenfalls ausgestorbenen Phorusrhacidae („Terrorvögel“), bei denen sich unter anderem ein ähnlich gearteter Schnabel findet. Beide Linien entstanden demzufolge unabhängig voneinander nach dem Aussterben der Dinosaurier vor rund 65 Millionen Jahren und besetzten die dadurch frei gewordene Nische der großen Beutegreifer.[9] Untersuchungen zur Biomechanik des Unterkiefers in Verbindung mit der massiven Symphyse und dem mächtigen Oberkiefer ergaben, dass der Schnabel und die Kiefergelenke darauf ausgelegt waren, eine hohe Beißkraft zu entwickeln, vor allem im hinteren Teil des Schnabels. Dabei hielt die Symphyse großen, sowohl quer als auch vertikal wirkenden Beanspruchungen stand. Die daraus resultierende Kraft, so wurde errechnet, reichte aus, um jede bekannte Nuss zu knacken. Für eine rein pflanzliche Ernährung wurde der Schnabel daher als zu gut ausgestattet angesehen, auch wenn er im Bau jenen von Papageien und anderen Samen- und Nussfressern ähnelt. Er diente nach dieser Ansicht somit dem Beutefang, dem Zerfleischen von Aas und dem Zerbrechen von Knochen oder von Krustentieren.[7] Als Nahrungsgrundlage dienten demnach kleinere Säugetiere wie das unter anderem im Geiseltal häufig auftretende Propalaeotherium, aber auch kleinere Reptilien und Weichtiere. Dabei wirkte Gastornis beim Beutefang als starker „Buschbrecher“.[11] Als Spitzenprädator seiner Zeit hätte Gastornis lediglich mit den großen, zumeist Wasser bewohnenden Krokodilen konkurriert.[2]
Als alternatives Szenario wurde bereits in den 1970er Jahren eine überwiegend pflanzliche Ernährung angenommen, wobei diese Ansicht zu Beginn der 1990er Jahre neuen Aufschwung erhielt. Dabei wurde argumentiert, dass dem Oberschnabel das für Greifvögel typische hakenförmige Ende fehlt – lediglich ein juveniles Skelett aus dem Bighorn-Becken in Wyoming weist einen leicht nach unten gerichteten Mittelkieferknochen auf.[27] Auch besaßen die Füße nicht die charakteristischen Greifvogelkrallen, die ebenfalls bei den Phorusrhacidae („Terrorvögel“) nachgewiesen sind und deren Abwesenheit man auch bei den fossil erhaltenen, Gastornis zugeordneten, Fußabdrücken beobachten kann. Dieser Ansicht zufolge ernährte sich Gastornis von zahlreichen Pflanzen der dichten tropischen Regenwälder, die er mit dem Schnabel zerbiss oder zerschnitt. Als eine in Erscheinung und möglicherweise in der Ernährungsweise zu Gastornis analoge Vogelgattung wird darüber hinaus der heute noch lebende Takahe (Porphyrio) angesehen. Dieser heute auf Neuseeland verbreitete, flugunfähige Vogel verfügt über einen kräftigen, hohen Schnabel, einen kurzen Hals und ebenfalls kurze Beinknochen, besitzt aber nur ein Gewicht von maximal 3 kg und ist ein Verwandter der Rallen. Eine vollständig auf Pflanzen basierende Ernährungsweise ist bei heutigen flugfähigen Vögeln wenig bekannt, da die für den Flug benötigte Energie unmittelbar aus der Nahrung bereitgestellt werden muss. Bei großen flugunfähigen Vögeln kommt eine derartige Ernährungsweise aber vor. Da Blätter eine eher energiearme Kost sind, die zudem lange verdaut wird, hätte Gastornis bei einer deutlich blätterhaltigen Pflanzenkost sehr viel Zeit mit dem Fressen verbringen müssen. Die Umstellung auf eine rein pflanzliche Nahrung bei Gastornis wurde aber teilweise mit dessen enormer Körpergrößenzunahme im Zusammenhang gesehen.[9][26] Eine im Jahr 2013 durchgeführte Analyse von Kalzium-Isotopen in den fossilen Knochen von Gastornis-Funden aus dem Geiseltal ergab ähnliche Isotopenverhältnisse wie bei pflanzenfressenden Säugetieren. Diese unterscheiden sich wiederum deutlich von denen der fleischfressenden Säugetiere und den sich ebenso ernährenden Dinosauriern wie Tyrannosaurus, da Tiere mit einer derartigen Ernährungsweise einen deutlich höheren Anteil an flüchtigen Isotopen (42Ca) aufweisen.[28][29][30] Zu vergleichbaren Ergebnissen kamen Untersuchungen von Gastornis-Resten aus mehreren Fundstellen Frankreichs. Diese wurden mit verschiedenen carnivor und herbivor lebenden Vögeln und Säugetieren verglichen, ebenso erfolgten erneute biomechanische Untersuchungen des Schnabels. Hierbei stellte sich heraus, dass vor allem der Musculus adductor mandibulae vergleichbar ähnlich stark entwickelt war wie bei den heutigen, samenknackenden Formen der Darwinfinken, während er bei räuberisch lebenden Greifvögeln weniger deutlich hervortritt. Der Muskel steht in Verbindung mit der Nahrungsaufnahme und dient dem Schließen des Schnabels und damit dem Zerteilen der aufgenommenen Nahrung, wobei für Gastornis ebenfalls ein Zerbeißen von harten Samen angenommen wird. Dadurch ist es nun wahrscheinlicher, dass die Vertreter der Gattung Gastornis Pflanzenfresser waren.[31]
Fortbewegung
Die Laufbeine sind ähnlich aufgebaut wie die des heutigen Afrikanischen Straußes (Struthio). Wie bei diesem konzentriert sich auch bei Gastornis ein Großteil der Muskelmasse der Beine direkt unterhalb des Körpers, was an dem eher kurzen und sehr kompakten Oberschenkelknochen erkennbar ist. Dadurch wirken die Beine wie ein Pendel unter dem Rumpf und ermöglichen die Fortbewegung. Als schneller Läufer besitzt der Strauß einen langen und schlanken Tarsometatarsus und zwei nach vorn ragende Zehenstrahlen an den Füßen. Andere heute lebende bodenbewohnende Vögel, unter anderem Kasuare (Casuarus) und Nandus (Rhea), verfügen neben einem ebenfalls langen Tarsometatarsus über Füße mit drei nach vorn zeigenden Strahlen. Diese Reduktion der Anzahl der Zehenstrahlen gegenüber vier bei vielen flugfähigen Vögeln ist eine funktionale Anpassung an das schnelle Laufen in offenen Landschaften und reduziert den Reibungswiderstand mit dem Boden. Gastornis hingegen besitzt einen vergleichsweise kurzen und breiten Tarsometatarsus. Das Laufbein endet zudem in vierstrahligen Füßen, von denen ein Strahl nach hinten zeigt (anisodactyl). Diese Merkmalskombination ist als sehr urtümlich anzusehen, auch im Vergleich zu anderen heute lebenden Laufvögeln. Aufgrund des kurzen Fußknochens und des nach hinten zeigenden vierten Zehs wird für Gastornis nur eine geringe Laufgeschwindigkeit rekonstruiert. Da der überwiegende Lebensraum der Vogelgattung dichte tropische und subtropische Regenwälder umfasst, wird ein langsamer Laufgang angenommen, der möglicherweise von kurzen schnelleren Gangarten unterbrochen wurde. Auch die wenig gebogenen Krallen der Füße stellen eine Anpassung an stärkere Bodenaktivitäten und langsames Gehen dar und sind weniger für eine schnelle Jagd nach Beutetieren geeignet.[9][2][32] Ob dabei die hintere kurze Zehe funktional den Fuß bei der Fortbewegung unterstützte, ist unklar. Die Trittsiegel aus der Chuckanut-Formation weisen keinen hinteren, vierten Zehenabdruck auf, so dass dieser Zeh möglicherweise zu stark reduziert gewesen sein könnte.[26]
Geschlechtsdimorphismus
Anhand der Funde von Louvois südlich von Reims, die zahlreiche Reste der Fuß- und Beinknochen umfassen, ließ sich eine gewisse Größendifferenz feststellen. So wird die erste Phalanx des vierten Zehenstrahls bei großen Individuen bis zu 7,6 cm lang, bei kleineren nur bis zu 4,9 cm. Entsprechende Maße des zweiten Zehengliedes belaufen sich auf 3,1 beziehungsweise 2,3 cm. Vergleichbare Unterschiede ließen sich auch bei anderen Funden ermitteln. Die Größenabweichungen betragen zwischen 30 und 65 %. Im Aufbau der einzelnen Knochen besteht kein Unterschied, so dass nicht von der Anwesenheit verschiedener Arten auszugehen ist. Die Wissenschaftler vermuten daher einen Geschlechtsdimorphismus. Dieser ist auch bei anderen großen bodenlebenden Vögeln belegt. Bei den Moas etwa wurden weibliche Tiere um bis zu 150 % größer und bis zu 280 % schwerer als männliche. Bei den heutigen Straußen ist das Verhältnis dagegen umgekehrt, da die männlichen Individuen größer als die weiblichen werden. Für Gastornis kann vorerst nicht bestimmt werden, welches Geschlecht größer war.[15] Über einen Größenunterschied zwischen männlichen und weiblichen Tieren wurde bereits bei der Aufarbeitung der Funde aus dem Geiseltal und aus Saint-Papoul nachgedacht.[2][16]
Systematik
Nähere Verwandtschaft von Gastornis nach Worthy et al. 2017[33]
|
Gastornis ist eine Gattung aus der Familie der Gastornithidae. Diese stellen große flugunfähige Vögel dar, die durch einen kräftigen, seitlich verschmälerten Schnabel charakterisiert sind, der eine konvex gebogene Oberseite (Culmen) aufweist, im Gegensatz zu den ähnlich gebauten Phorusrhacidae aber keine hakenartige Spitze besitzt. Die Herkunft der Gastornithidae ist nicht vollständig geklärt. Möglicherweise können sie mit großen Vögeln aus der Oberkreide in Verbindung gebracht werden, wie sie mit Gargantuavis, anhand eines rund 6,5 cm langen Synsacrums aus der Nähe von Fox-Amphous im französischen Département Var in der Provence erstmals für diese Zeitperiode nachgewiesen wurden.[34] Weitere Funde, so ein Becken sowie ein Oberschenkelknochen und mehrere Wirbel, stammen ebenfalls aus dem südlichen Frankreich. Diese Fossilreste weisen auf einen etwa straußengroßen Vogel hin, der möglicherweise bis zu 140 kg wog, und besitzen Ähnlichkeiten zu jenen von Gastornis,[35][36] allerdings stellen diese spätkreidezeitlichen Vögel höchstwahrscheinlich nicht die direkten Vorfahren dar.[4] Anhand der Merkmale der Gliedmaßenknochen und des Kiefergelenks ist aber deutlich, dass Gastornis und seine in der Ordnung Gastornithiformes zusammengefassten Verwandten mit den Gänsevögeln (Anseriformes) und den Hühnervögeln (Galliformes) verwandt sind.[3] Laut einer Untersuchung aus dem Jahr 2017 bilden die Gastornithiformes möglicherweise die Schwestergruppe zu den Hühnervögeln. Sie schließen hierin außerdem noch die einst in Australien heimischen Donnervögel (Dromornithidae) mit ein.[33] Eine vier Jahre später veröffentlichte Analyse ordnet dieser umfassenderen Gruppe zusätzlich noch die aus Südamerika bekannten Brontornithidae mit Brontornis zu. Bei letzteren ist die genaue phylogenetische Position jedoch umstritten.[37]
Im Laufe der Forschungsgeschichte wurden zahlreiche Arten von Gastornis beschrieben, heute anerkannt sind folgende:[2][22][38]
- G. geiselensis Fischer, 1976
- G. gigantaeus Cope, 1876
- G. laurenti Mourer-Chauviré & Bourdon, 2020
- G. parisiensis Hébert, 1855
- G. russelli Martin, 1992
- G. sarasini Schaub, 1929
- G. xichuanensis Hou, 1980
G. russelli stellt die stammesgeschichtlich älteste und kleinste Art dar, die nur die Hälfte der Größe eines heutigen Straußes erreichte. Sie steht als Schwesterform den anderen Arten von Gastornis gegenüber. Weitere beschriebene Arten wie G. steini, G. edwardsi und G. klaasseni sind heute nicht mehr gültig. Für Nordamerika wird neben G. gigantaeus teilweise auch noch G. regens als eigenständige Art anerkannt, die in der Proportion der Zehenglieder etwas abweicht. Andere Forscher halten G. geiselensis für synonym zu G. sarasini, was aber nur teilweise anerkannt ist.[2] Generell wird eine neue Revision der Gattung angemahnt.[3]
Stammesgeschichte
Da die ältesten Nachweise von Gastornis aus dem heutigen Europa stammen, wird die Entstehung der Gattung und Familie hier vermutet. Allerdings ist der Ursprung bisher nicht geklärt. Während des Paläozän stellte das heutige Europa eine Insel dar mit endemischer Fauna, die hier nach dem Massensterben an der Kreide-Paläogen-Grenze, dem ein Großteil der Landwirbeltiere über 25 kg Körpergewicht zum Opfer fiel, zahlreiche ökologische Nischen vorfand. Zur Zeit der frühesten Nachweise von Gastornis vor rund 62 Millionen Jahren erreichten die größten Säugetiere dieser Insel nur ein Gewicht von rund 45 kg. Möglich ist, dass sich die Gattung unter einem derartigen isolierten Inselökosystem zur Riesenform entwickelte, ähnlich wie dies aus späterer Zeit mit den Aepyornithidae auf Madagaskar überliefert ist, die dort zur größten einheimischen Tiergruppe heranwuchs, die alle vorhandenen Säugetiere an Größe übertraf. Erst im Übergang zum Unteren Eozän entstanden Verbindungen zu Nordamerika (via Grönland), wodurch auch ein erheblicher Faunenaustausch begann. Dies führte einerseits dazu, dass Gastornis als größter Bewohner von einigen großen Säugetieren wie etwa Coryphodon aus der Gruppe der Pantodonta abgelöst wurde, andererseits ermöglichte es ihm, ebenfalls neue Regionen zu erreichen. Als wahrscheinlichste Route gilt die über Grönland, was auch die spärlichen Funde von Ellesmere Island aufzeigen.[39] Nach Asien gelangte die Gattung entweder über Nordamerika oder eventuell direkt über die Turgai-Straße, die den Kontinent von Europa trennte. Auf beiden Landmassen lebten aber bereits große pflanzenfressende Säugetiere, die im Fall der Pantodonten bis zu 600 kg wogen. Annehmbar ist deshalb, dass Gastornis in Konkurrenz zu diesen in Nordamerika und Asien bereits zum Ende des Unteren Eozän ausstarb. In Europa hielt sich die Gattung noch erfolgreich bis zum ausgehenden Mitteleozän vor etwa 43 Millionen Jahren.[31][4]
Gastornis und Diatryma – Zur Forschungsgeschichte

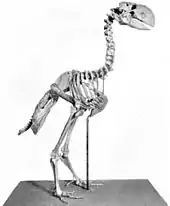
Die Erforschung der Gattung Gastornis reicht bis in die Mitte des 19. Jahrhunderts zurück. Im Jahr 1855 entdeckte der französische Physiker Gaston Planté (1834–1889), der spätere Erfinder des Bleiakkumulators, in Ablagerungen des Oberen Paläozäns und Unteren Eozäns, dem „Conglomérat de Meudon“ in einem westlichen Vorort von Paris, die ersten Knochen eines riesigen Vogels, die einen Femur und einen Tibiotarsus umfassten. Diese wurden im gleichen Jahr von Edmond Hébert als zur neu geschaffenen Gattung Gastornis gehörig beschrieben, die damals zu den ältesten bekannten Vogelgattungen überhaupt zählte. Der Name Gastornis leitet sich dabei vom Vornamen Plantés, Gaston, und der griechischen Bezeichnung ὄρνις (órnis) für „Vogel“ ab. Nur kurze Zeit später kamen einige wenige weitere Funde bei Passy, ebenfalls nahe Paris gelegen, zum Vorschein. Die systematische Position von Gastornis war zu jener Zeit völlig ungeklärt. Hébert selbst schlussfolgerte (1855), dass Gastornis zu schwer zum Fliegen war und stellte die Gattung wie auch Alphonse Milne-Edwards (1867) nahe zu den Entenvögeln (Anatidae), Édouard Armand Lartet (1855) dachte an eine Verwandtschaft mit den Regenpfeiferartigen (Charadriiformes), Achille Valenciennes (1855) sah sogar Übereinstimmungen mit Albatrossen (Diomedeidae) und Charles Lucien Jules Laurent Bonaparte (1856) hielt Gastornis für einen Verwandten des großen, madagassischen Vogels Aepyornis. Richard Owen wiederum gab 1859 eine erste genaue Beschreibung des Tibiotarsus wieder und hielt Gastornis für einen terrestrisch lebenden Vogel.[40] Sehr umfangreiches Material entdeckte man nach einem Zeitabstand von rund 20 Jahre im unterpaläozänen „Conglomérat de Cernay“ bei Reims, welche damals die älteste bekannte Fossilfundstelle des Tertiärs in Europa war. Dieses Fundmaterial veranlasste Victor Lemoine (1823–1911) im Jahr 1881 dazu, eine Skelettrekonstruktion anzufertigen, die vom heute bekannten, tatsächlichen Aussehen von Gastornis aber beträchtlich abwich. So erarbeitete er einen großen, grazil wirkenden Vogel mit langem Hals, kurzen Flügeln und langen Beinen mit einem ebenfalls langen und schlanken Tarsometatarsus. Die paddelartig gestalteten Füße suggerierten zudem Schwimmfähigkeit. Lemoine hatte sich bei seiner Rekonstruktion von kürzlich entdeckten Vögeln wie Archaeopteryx oder Hersperornis inspirieren lassen. Diese war über die nächsten Jahrzehnte bildbestimmend für das Aussehen von Gastornis. In der darauffolgenden Zeit wurden nur wenige weitere Funde zu Gastornis gestellt, so die Fossilreste von Croydon in England und Walbeck in Deutschland.[14][41][40]
Anhand eines großen Tarsometatarsus aus dem Untereozän des US-Bundesstaates New Mexico beschrieb Edward Drinker Cope (1840–1897) im Jahr 1876 die Gattung Diatryma, wobei der Name sich auf ein den Knochenschaft am oberen Ende durchdringendes Foramen, eine Öffnung im Knochen, bezieht. Cope erkannte zwar Ähnlichkeiten zum europäischen Gastornis, favorisierte aber einen eigenen Gattungsnamen.[18] Allerdings waren in der Folgezeit kaum anatomische Vergleiche der beiden Vogelgattungen möglich, da aus Nordamerika nur spärliche Fossilreste bekannt wurden. Einen Zehenknochen aus New Jersey verwies Othniel Charles Marsh 1894 zu Barornis, der später mit Diatryma vereint wurde. Die Situation änderte sich erst 1916, als im Bighorn-Becken von Wyoming ein nahezu vollständiges Skelett gefunden wurde, welches William Diller Matthew (1871–1930) und Walter W. Granger (1872–1941) im folgenden Jahr umfänglich beschrieben.[6] Ihre Skelettrekonstruktion erwies sich aber als völlig unterschiedlich zu Lemoines, was durch den untersetzt-robusten Körper und den riesigen Kopf mit mächtigem Schnabel verdeutlicht wurde. Dadurch fiel den beiden Forschern ein Vergleich mit dem europäischen Gastornis relativ schwer, sie äußerten auch vorsichtig Zweifel an der Zugehörigkeit aller Knochen in Lemoines Rekonstruktion zu Gastornis. Weiterhin bescheinigten sie beiden Vogelvertretern einen jeweils eigenständigen Gattungsstatus, einer Ansicht, der später zahlreiche Wissenschaftler folgten.[6] Während der Rekonstruktion des Diatryma-Skelettes hatten Matthew und Granger aufgrund des teils unvollständigen Fußskeletts den Tarsometatarsus verlängert rekonstruiert, möglicherweise beeinflusst durch die langen Beine heutiger Laufvögel, in dessen Folge nicht nur der mächtige Schnabel, sondern auch die Gliedmaßen ähnlich denen der Phorusrhacidae („Terrorvögel“) wirkten. Diese Vogelgruppe war seit den 1880er Jahren mit den intensiv einsetzenden Forschungen in Patagonien in den Focus der Öffentlichkeit gelangt. Hauptsächlich in der Santa-Cruz-Formation waren zahlreiche „Terrorvögel“ wie Phorusrhacos, Psilopterus und Patagornis entdeckt worden.[42][43] Dabei handelte es sich um große, flugunfähige Vögel mit langen Gliedmaßen, massigen Schnabel mit greifvogelartigem, spitzen Ende und Füße mit kräftigen, gebogenen Krallen, was die Tiere als schnelle und effektive Beutegreifer charakterisierte. Beide Forscher befürworteten eine Trennung von Diatryma und den Phorusrhaciden, nahmen aber aufgrund der annähernd übereinstimmenden Größe und den Ähnlichkeiten im Körperbau einschließlich der langen unteren Gliedmaßenabschnitte auch eine vergleichbare Lebensweise an.[6] In der Folgezeit bekamen auch europäische Neufunde, so aus dem Geiseltal und der Grube Messel in Deutschland oder von Lyon in Frankreich eine Zuweisung zur Gattung Diatryma.[14][41][40]

Obwohl sich Matthew und Granger in ihrer Erstveröffentlichung des Skeletts von Diatryma neben dessen prinzipiellen äußerlichen Übereinstimmung mit den „Terrorvögeln“, teilweise hervorgerufen durch die falsche Rekonstruktion eines langen Tarsometatarsus, nur wenig über dessen Lebensweise äußerten, kam in der Mitte der 1920er Jahre erstmals die Sichtweise über eine fleischliche Ernährung dieses riesigen Vogels auf, die auf kleinen Reptilien und Säugetieren basieren sollte. Die extrem populäre Vorstellung einer Jagd auf die kleinen, am Beginn ihrer Stammesgeschichte stehenden Urpferdchen wie Hyracotherium oder Paarhufer wie Diacodexis etablierte sich dabei in den 1930er Jahren, maßgeblich unterstützt durch Alfred Romer. Selbst die Entdeckung eines Teilskeletts eines noch juvenilen Diatryma-ähnlichen Vogels aus dem Jahr 1928 im Bighorn-Becken von Wyoming mit nahezu vollständigem Fuß, der einen kurzen und breiten Tarsometatarsus anstatt des langschmalen zeigte, beeinflusste diese Vorstellung kaum (beschrieben wurde das Teilskelett als Omorhamphus, heute ein Synonym zu Gastornis).[27] Zahlreiche Forscher vertraten die Meinung eines schnell laufenden, jagenden Riesenvogels im Eozän, so etwa George Gaylord Simpson 1950 in seinem Vergleich von Diatryma mit dem südamerikanischen „Terrorvögeln“ und dem europäischen Gastornis nach Lemoine.[44] Auch in Europa war die Ansicht verbreitet, so etwa auch für die Diatryma-Funde aus dem Geiseltal, wobei als Beute des Vogels das hier nachgewiesene Propalaeotherium angenommen wurde.[2] Dabei war es Karlheinz Fischer, der 1978 erstmals anhand von vollständigen Funden des Tarsometatarsus aus dem Geiseltal im Vergleich zu anderen Beinknochen die korrekten Proportionen in den Gliedmaßen von Diatryma darstellte. So erreichte der Tarsometatarsus rund 24 bis 25 cm Länge, deutlich abweichend von den von Matthew und Granger projizierten 38 cm, und nahm knapp 21 % der Beinlänge ein, womit der Fuß deutlich kürzer war als vorher angenommen. Damit stellte sich Diatryma nicht als schnell laufender, sondern mehrheitlich langsam schreitender Vogel heraus.[1][40]
Noch bis in die ausgehenden 1980er Jahre wurden die beiden Vogelgattungen Gastornis und Diatryma unterschiedlichen Familien zugewiesen, erstere zu den Gastornithidae und letztere zu den Diatrymidae. Die Erkenntnis, dass beide Vogelgattungen sich wesentlich ähnlicher waren, erwuchs erst zu Beginn der 1990er Jahre. Während einer Revision der Skelettrekonstruktion von Lemoine und der Aufnahme von neuem Fundmaterial im Jahr 1992 erkannte L. D. Martin, dass die Rekonstruktion nur zu einem geringen Teil auf Fossilresten von Vögeln basierte, nämlich nur auf einem Wirbel und Teilen der Laufbeine sowie einige Fußknochen, der weitaus größere Teil dagegen von Fischen und Reptilien entstammte, wodurch die urtümliche Gestalt von Gastornis hervorgerufen worden war. Martin stellte dabei auch morphologische Ähnlichkeiten von Gastornis zu Diatryma her und verwies beide zur Familie Gastornithidae. Neufunde aus Reims, so ein oberes Ende eines Tibiotarsus, der von Gastornis bis dahin noch nicht beschrieben worden war, von Diatryma aber bereits mehrfach vorlag, ließ 1997 die Frage aufkommen, ob beide Gattungen nicht identisch seien.[14] Nur wenige Jahre später, 2002, wurden Gastornis und Diatryma formal synonymisiert, was 2008 durch den Neufund eines weiteren Tibiotarsus aus Saint-Papoul bestätigt werden konnte. Dabei hatte Éric Buffetaut diesen Fund zur Typusart G. parisiensis gestellt, bemerkte aber gleichzeitig sowohl Ähnlichkeiten zu vergleichbaren Knochen von G. geiselensis, das ursprünglich als Diatryma beschrieben worden war, als auch zu ähnlichen Funden aus Nordamerika. Die einzelnen, bestehenden Unterschiede werden als so klein angesehen, dass Diatryma nun als synonym zu Gastornis gilt.[17][2] Die Funde aus Saint-Papoul gehören seit dem Jahr 2020 nach einer Neubeschreibung zur Art G. laurenti.[38]
Literatur
- Allison V. Andors: Diatryma among the Dinosaurs. Natural History 1995 (6), S. 68–72
- Delphine Angst und Eric Buffetaut: Palaeobiology of Giant Flightless Birds. Oxford, 2017, S. 1–282 (S. 173–214)
- Eric Buffetaut und Delphine Angst: „Terror cranes“ or peaceful plant eaters: changing interpretations of the palaeobiology of gastornithid birds. Revue de Paléobiologie 32 (2), 2013, S. 413–422
- Eric Buffetaut und Delphine Angst: Stratigraphic distribution of large flightless birds in the Palaeogene of Europe and its palaeobiological and palaeogeographical implications. Earth-Science Reviews 32 (2), 2014, S. 394–408
- Meinolf Hellmund: Reappraisal of the bone inventory of Gastornis geiselensis (Fischer, 1978) from the Eocene “Geiseltal Fossillagerstätte” (Saxony-Anhalt, Germany). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 269 (2), 2013, S. 203–220
Einzelnachweise
- Karlheinz Fischer: Neue Reste des Riesenlaufvogels Diatryma aus dem Eozän des Geiseltales bei Halle (DDR). Mitteilungen Zoologisches Museum Berlin, Suppl., 54, Annalen für Ornithologie 2, 1978, S. 133–144.
- Meinolf Hellmund: Reappraisal of the bone inventory of Gastornis geiselensis (Fischer, 1978) from the Eocene “Geiseltal Fossillagerstätte” (Saxony-Anhalt, Germany). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 269 (2), 2013, S. 203–220.
- Gerald Mayr: Gastornithidae. In: Gerald Mayr: Paleogene Fossil Birds. Springer-Verlag, Berlin und Heidelberg, 2009, S. 44–47.
- Eric Buffetaut und Delphine Angst: Stratigraphic distribution of large flightless birds in the Palaeogene of Europe and its palaeobiological and palaeogeographical implications. Earth-Science Reviews 32 (2), 2014, S. 394–408.
- Delphine Angst und Eric Buffetaut: Palaeobiology of Giant Flightless Birds. Oxford, 2017, S. 1–282 (S. 173–214).
- William Diller Matthew und Walter W. Granger: The skeleton of Diatryma, a gigantic bird from the lower Eocene of Wyoming. Bulletins of the American Museum Natural History, 37, 1917, S. 307–326.
- Lawrence M. Witmer und Kenneth D. Rose: Biomechanics of the jaw apparatus of the gigantic Eocene bird Diatryma: implications for diet and mode of life. Paleobiology 17 (2), 1991, S. 95–120.
- Delphine Angst und Eric Buffetaut: The first mandible of Gastornis Hébert, 1855 (Aves, Gastornithidae) from the Thanetian (Paleocene) of Mont-de-Berru (France). Revue de Paléobiologie, Genève 138, 2013, S. 423–432.
- Allison V. Andors: Diatryma among the Dinosaurs. Natural History 1995 (6), S. 68–72.
- Gerald Mayr: The birds from the Paleocene fissure filling of Walbeck (Germany). Journal of Vertebrate Paleontology 27 (2), 2007, S. 394–408.
- Karl-Heinz Fischer: Der Riesenlaufvogel Diatryma aus der eozänen Braunkohle des Geiseltales. Hallesches Jahrbuch für Mitteldeutsche Erdgeschichte 4, 1962, S. 26–33.
- Dietrich E. Berg: Nachweis des Riesenlaufvogels Diatryma im Eozän von Messel bei Darmstadt/Hessen. Notizblätter des hessischen Landesamtes für Bodenforschung 93, 1965, S. 68–72.
- Norbert Micklich: Laufvögel. In: Gabrielle Gruberg und Norbert Micklich (Hrsg.): Messel – Schätze der Urzeit. Stuttgart, 2007, S. 85–93.
- Eric Buffetaut: New remains of the giant bird Gastornis from the Upper Paleocene of the eastern Paris Basin and the relationships between Gastornis and Diatrjtma. Neues Jahrbuch für Geologie und Paläontologie Monatshefte 1997 (3), S. 179–190.
- Cécile Mourer-Chauviré und Estelle Bourdon: The Gastornis (Aves, Gastornithidae) from the Late Paleocene of Louvois (Marne, France). Swiss Journal of Palaeontology 135, 2016, S. 327–341, doi:10.1007/s13358-015-0097-7.
- Estelle Bourdon, Cecilie Mourer-Chauvire und Yves Laurent: Early Eocene birds from La Borie, southern France. Acta Palaeontologia Polonica 61 (1), 2016, S. 175–190, doi:10.4202/app.00083.2014.
- Eric Buffetaut: First evidence of the giant bird Gastornis from southern Europe: a tibiotarsus from the Lower Eocene of Saint-Papoul (Aude, southern France). Oryctos, 7, 2008, S. 75–82.
- Edward Drinker Cope: On a gigantic bird from the Eocene of New Mexico. Proceedings of the Academy of Natural Sciences of Philadelphia 28, 1876, S. 10–11.
- R. M. West und Mary R. Dawson: Vertebrale Paleontology and the Cenozoic History of the North Atlantic Region. Polarforschung 48 (1/2), 1978, S. 103–119.
- Jaelyn J. Eberle und David R. Greenwood: Life at the top of the greenhouse Eocene world — A review of the Eocene flora and vertebrate fauna from Canada’s High Arctic. Geological Society of America Bulletin; January/February 124 (1/2), 2012, S. 3–23.
- Hou Lian-hai: New form of the Gastornithidae from the Lower Eocene of the Xichuan, Honan. Vertebrata Palasiatica 18 (2), 1980, S. 111–115.
- Eric Buffetaut: The giant bird Gastornis in Asia: A revision of Zhongyuanus xichuanensis Hou, 1980, from the Early Eocene of China. Paleontological Journal 47 (11), 2013, S. 1302–1307.
- T. D. A. Cockerell: The supposed plumage of the Eocene bird Diatryma. American Museum Novitates 62, 1923, S. 1–4.
- Alexander Wetmore: The Supposed Plumage of the Eocene Diatryma. Auk 47 (4), 1930, S. 579–580.
- Lance Grande: The lost world of Fossil Lake. Snapshot from deep time. University of Chicago Press, Chicago und London, 2013, S. 1–425 (S. 216 und 225).
- George E. Mustoe, David S. Tucker, Keith L. Kemplin: Giant Eocene bird footprints from Northwest Washington, USA. Palaeontology 55 (6), 2012, S. 1293–1305, doi:10.1111/j.1475-4983.2012.01195.x.
- William J. Sinclair: Omorhamphus, a New Flightless Bird from the Lower Eocene of Wyoming. Proceedings of the American Philosophical Society 67 (1), 1928, S. 51–65.
- Max-Planck-Gesellschaft: Harmlosser Terrorvogel. Der Urzeitvogel Gastornis war vermutlich kein Raubvogel, sondern ein Pflanzenfresser. MPG vom 30. August 2013 ().
- Thomas Tütken, P. Held und S. J. G. Galer: Isotopes in vertebrate bioapatite: proxies for climate,pCO2 and diet. Miner Mag 77, 2013, S. 2368 (Conference abstracts) ().
- Alexander K. Hastings und Meinolf Hellmund: Aus der Morgendämmerung: Pferdejagende Krokodile und Riesenvögel. Neueste Forschungsergebnisse zur eozänen Welt Deutschlands vor ca. 45 Millionen Jahren. Halle (Saale), 2015, S. 1–120.
- Delphine Angst, C. Lécuyer, R. Amiot, Eric Buffetaut, F. Fourel, F. Martineau, S. Legendre, A. Abourachid und A. Herrel: Isotopic and anatomical evidence of an herbivorous diet in the Early Tertiary giant bird Gastornis. Implications for the structure of Paleocene terrestrial ecosystems. Naturwissenschaften 101, 2014, S. 313–322.
- Delphine Angst, Eric Buffetaut, Christophe Lécuyer und Romain Amiot: A new method for estimating locomotion type in large ground birds. Palaeontology 59 (2), 2016, S. 217–223, doi:10.5061/dryad.609j4.
- Trevor H. Worthy, Federico J. Degrange, Warren D. Handley und Michael S. Y. Lee: The evolution of giant flightless birds and novel phylogenetic relationships for extinct fowl (Aves, Galloanseres). Royal Society open sciience 4, 2017, S. 170975, doi:10.1098/rsos.170975
- Eric Buffetaut, Jean Le Loeuff, Patrick Mechin und Annie Mechin-Salessy: A large French Cretaceous bird. Nature 377, 1995, S. 110.
- Eric Buffetaut und Jean Le Loeuff: Gargantuavis philoinos: Giant bird or giant pterosaur? Annales de Paléontologie 96 (2010), S. 135–141.
- Eric Buffetaut: Les oiseaux fossiles du Crétacé Supérieur de L'Herault. Bulletin de la Societe d'Etude des Sciences Naturelles de Béziers 66, 2012, S. 34–39.
- Federico L. Agnolin: Reappraisal on the Phylogenetic Relationships of the Enigmatic Flightless Bird (Brontornis burmeisteri) Moreno and Mercerat, 1891. Diversity 13, 2021, S. 90, doi:10.3390/d13020090.
- Cécile Mourer-Chauviré und Estelle Bourdon: Description of a new species of Gastornis (Aves, Gastornithiformes) from the early Eocene of La Borie, southwestern France. Geobios 63, 2020, S. 39–46, doi:10.1016/j.geobios.2020.10.002.
- Eric Buffetaut: The unfinished story of the Early Tertiary giant bird Gastornis. DGF On Line Series 1, 1997 ().
- Eric Buffetaut und Delphine Angst: „Terror cranes“ or peaceful plant eaters: changing interpretations of the palaeobiology of gastornithid birds. Revue de Paléobiologie 32 (2), 2013, S. 413–422.
- Eric Buffetaut: L’oiseau géant Gastornis : interprétation, reconstitution et vulgarisation de fossiles inhabituels dans la France du XIXe siècle. Bulletin de la Société géologique de France 168, 1997, S. 805–811.
- Eric Buffetaut: Tertiary ground birds from Patagonia (Argentina) in the Tournouër collection of the Muséum National d’Histoire Naturelle, Paris. Bulletin de la Société Géologique de France 185 (3), 2014, S. 207–214.
- Federico J. Degrange, Jorge I. Noriega und Juan I. Areta: Diversity and paleobiology of the Santacrucian birds. In: Sergio F. Vizcaíno, Richard F. Kay und M. Susana Bargo (Hrsg.): Early Miocene paleobiology in Patagonia: High-latitude paleocommunities of the Santa Cruz Formation. Cambridge University Press, New York, 2012, S. 138–155.
- George Gaylord Simpson: Are Nonflying Wings Functionless? Science 112, 1950, S. 342.
Weblinks
- Thomas Tütken: Vom Top-Räuber zum Vegetarier. Steinmann-Institut für Geologie, Mineralogie und Paläontologie der Universität Bonn, 30. August 2013
- Eric Buffetaut: The unfinished story of the Early Tertiary giant bird Gastornis. Geological Society of Denmark