Deinonychus
Deinonychus (altgriechisch δεινός ‚schrecklich‘ und ὄνυξ, Genitiv ὄνυχος ‚Klaue/Kralle‘) war eine Gattung fleischfressender Dinosaurier aus der Gruppe Dromaeosauridae. Die einzige beschriebene Art ist Deinonychus antirrhopus.
| Deinonychus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelettrekonstruktion von Deinonychus antirrhopus | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Unterkreide (Mittleres Aptium bis Unteres Albium) | ||||||||||||
| 123 bis 110,2 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Deinonychus | ||||||||||||
| Ostrom, 1969 | ||||||||||||
| Art | ||||||||||||
| ||||||||||||
Dieser bis zu 3,4 Meter lange Theropode lebte während der frühen Kreidezeit vor etwa 123 bis 110 Millionen Jahren (vom Mittleren Aptium bis zum Unteren Albium). Fossilien wurden in den US-amerikanischen Bundesstaaten Montana, Wyoming und Oklahoma in Gesteinen der Cloverly-Formation und der Antlers-Formation entdeckt. Zähne, die möglicherweise zu Deinonychus gehören, stammen aus dem viel weiter östlich gelegenen Maryland.
Studien des Paläontologen John Ostrom über Deinonychus revolutionierten in den späten 1960er-Jahren die Vorstellungen der Wissenschaftler über Dinosaurier, was zur „Dinosaurier-Renaissance“ führte und die Debatte entzündete, ob Dinosaurier gleichwarme („warmblütige“) Tiere waren. Zuvor galten Dinosaurier als schwerfällige Giganten, die heutigen Reptilien glichen. Der kleine Körper, die elegante, horizontale Körperhaltung und insbesondere die großen sichelartigen Fußkrallen zeigten, dass es sich bei Deinonychus um einen aktiven, agilen Prädator handelte.[1]
Merkmale
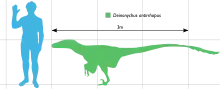
Wie die größten bekannten Exemplare zeigen, konnte Deinonychus eine Länge von 3,4 Metern erreichen. Ein Individuum dieser Größe besaß eine Schädellänge von 41 Zentimeter, eine Hüfthöhe von 87 Zentimeter und eine Masse von etwa 73 Kilogramm.[2] Der Schädel war mit kräftigen Kiefern ausgestattet, die etwa 70 gebogene, klingenartige Zähne trugen. Alle Zähne waren auf der Vorder- sowie auf der Rückseite gesägt, wobei die Zähnchen (Dentikel) auf der Rückseite der Zähne fast doppelt so groß waren wie die auf der Vorderseite.[3] John Ostroms erste, noch auf sehr fragmentarischen Überresten beruhende Schädelrekonstruktion zeigt einen relativ kompakten, breiten Schädel, der dem des Allosaurus ähnelt. Neuere Schädelfunde von Deinonychus und nahe verwandten Gattungen mit guter dreidimensionaler Erhaltung zeigen jedoch, dass der Gaumen gewölbter war, als John Ostrom vermutet hatte,[4] was zur Rekonstruktion einer weit schmaleren und längeren Schnauze führte. Gleichzeitig verbreiterte das paarige Jochbein (Jugale) den Schädel in der Augenregion, was ein gutes stereoskopisches Sehen ermöglichte. Der Schädel von Deinonychus unterschied sich von dem des Velociraptor unter anderem durch die tieferen, robusteren Unterkiefer[5] und durch die robustere Schädeldecke, die der von Dromaeosaurus ähnlich war. Des Weiteren war das Nasenbein nicht abgeflacht wie das des Velociraptor.[6] Sowohl der Schädel als auch der Unterkiefer zeigten Schädelfenster, welche das Gewicht des Schädels reduzierten. Bei Deinonychus war das vor den Augen gelegene Antorbitalfenster besonders groß.[4]

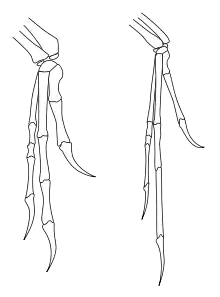
Wie bei allen Dromaeosauriden waren die Vordergliedmaßen sehr lang und erreichten bei Deinonychus 70 Prozent der Länge der Hinterbeine. Die große Hand machte dabei 40 Prozent der Gesamtlänge der Vordergliedmaßen aus. Der erste der drei Finger war der kürzeste, während der zweite der längste war. Jedes Hinterbein trug eine sichelförmige, ungewöhnlich große Kralle am zweiten Zeh. Im Leben des Tieres waren die Krallen größer, als sie fossil überliefert sind, da Dinosaurier ebenso wie heute lebende Archosaurier einen Überzug aus Horn über ihre Krallen besaßen. John Ostrom verglich eine Sichelkralle von Deinonychus (Exemplarnummer YPM 5205) mit Krallen heutiger Krokodile und Vögel und kam zu dem Ergebnis, dass die Kralle beim lebenden Tier über 120 Millimeter Länge maß.[1] Sowohl anatomische Nachweise[1] als auch Nachweise aus fossilen Fußspuren[7] zeigen, dass bei der Fortbewegung lediglich die dritte und die vierte Zehe den Boden berührten, während die die Sichelkralle tragende zweite Zehe bei der Fortbewegung über dem Boden gehalten wurde. Das Sitzbein (Ischium) war im Verhältnis zum Schambein (Pubis) kürzer als bei anderen Dromaeosauriden.[5] Die Schwanzwirbel waren mit einer Serie von verknöcherten Sehnen und stark verlängerten Wirbelfortsätzen ausgestattet – den Präzygapophysen an der Oberseite und den Chevron-Knochen an der Unterseite der Wirbelkörper. John Ostrom vermutete, dass diese Vorrichtung den Schwanz zu einem steifen Gegengewicht gemacht hat. Ein Fossil des eng verwandten Velociraptor mongoliensis (IGM 100/986) hat jedoch einen Schwanz im anatomischen Verbund, der horizontal S-förmig gebogen war. Dies deutet auf eine beträchtliche seitliche Beweglichkeit des Schwanzes hin.[8]
Obwohl bisher keine Federn in Verbindung mit Deinonychus-Fossilien entdeckt wurden, spricht der Nachweis von Federn in einer Vielzahl anderer Dromaeosauriden dafür, dass alle Vertreter der Gruppe einschließlich Deinonychus gefiedert waren (Prinzip der phylogenetischen Umklammerung).[9] So ist die gefiederte Dromaeoauriden-Gattung Microraptor sowohl geologisch älter als auch phylogenetisch ursprünglicher als Deinonychus.[10] Mehrere Fossilien von Microraptor zeigen Konturfedern an Armen, Beinen und Schwanz, die denen heutiger Vögel entsprechen.[9] Eine Elle (Ulna) des mit Deinonychus eng verwandten Velociraptor zeigt Ansatzstellen für Federn (englisch quill knobs), was auf lange Armfedern bei Velociraptor schließen lässt.[11]
Systematik
Deinonychus ist einer der am besten bekannten Dromaeosauriden[5] und ein enger Verwandter des kleineren Velociraptor, der aus jüngeren Schichten der Oberkreide Nordchinas und der Mongolei stammt.[12][13] Zwei weitere Gattungen der Oberkreide, Tsaagan aus der Mongolei[12] und der wenig bekannte Saurornitholestes aus Nordamerika,[2] könnten ebenfalls enge Verwandte von Deinonychus gewesen sein.
Velociraptor und Deinonychus bilden zusammen die Gruppe Velociraptorinae, die 1983 von Rinchen Barsbold aufgestellt wurde und ursprünglich Velociraptor als einzige Gattung enthielt.[14] Die langschnauzigen Velociraptorinen benutzten wahrscheinlich mehr ihre Klauen und weniger ihre Schädel zum Überwältigen von Beutetieren, im Unterschied zu anderen Dromaeosauriden wie den Dromaeosaurinen, die gedrungenere Schädel besaßen.[15] 1995 schrieb Philip Currie Deinonychus sowie eine Reihe weiterer Dromaeosauriden den Velociraptorinae zu, was allgemeine Akzeptanz fand.[16]
Eine neuere Studie von Nicholas Longrich und Philip Currie aus dem Jahr 2009 klassifiziert Deinonychus jedoch außerhalb der Velociraptorinae und stellt eine Eudromaeosauria genannte Gruppe auf, die Deinonychus, die Velociraptorinae und die Dromaeosaurinae enthalten soll.[17] Zusammen mit den Troodontiden bilden die Dromaeosauriden die Deinonychosauria, die nach Ansicht vieler Forscher die Schwestergruppe der Vögel und damit die mit den Vögeln am engsten verwandte Dinosauriergruppe ist.[18]
Entdeckungsgeschichte und Namensgebung
Frühe Entdeckungen und Erstbeschreibung
Fossile Überreste von Deinonychus stammen aus der Cloverly-Formation in Montana und Wyoming[1] sowie aus der etwa gleich alten Antlers-Formation in Oklahoma.[19] Die Cloverly-Formation wurde auf das späte Aptium bis frühe Albium datiert und ist damit etwa 123 bis 110 Millionen Jahre alt.[20][21] Darüber hinaus sind Zähne bekannt, die aus den Arundel Clay Facies (mittleres Aptium) der Potomac-Formation in Maryland stammen und möglicherweise dieser Gattung zugeordnet werden können.[22]

Die ersten Überreste wurden 1931 im südlichen Montana entdeckt, in der Nähe der Stadt Bridger. Barnum Brown, der Leiter der Ausgrabungen, hatte die Absicht, Überreste des Tenontosaurus auszugraben und zu präparieren. In seinem Grabungsbericht an das American Museum of Natural History berichtete er jedoch über die Entdeckung eines kleinen fleischfressenden (karnivoren) Dinosauriers in unmittelbarer Nähe zu einem Tenontosaurus-Skelett, der aber „von Kalk umschlossen und schwer zu präparieren“ war.[23] Barnum Brown nannte das Tier informell „Daptosaurus“ und bereitete die Präparation vor, um es beschreiben und ausstellen zu können, hat seine Arbeit aber niemals beendet.[24] Neben diesem Skelett entdeckte Barnum Brown in der Cloverly-Formation das Skelett eines noch kleineren Theropoden mit anscheinend übergroßen Zähnen, das er informell „Megadontosaurus“ nannte. Wie John Ostrom Jahrzehnte später feststellte, stammen die Zähne von Deinonychus – das Skelett aber stammt von einem ganz anderen Tier, das er Microvenator nannte.[24]
Im August 1964 entdeckte eine von John Ostrom geleitete Expedition des Peabody Museum of Natural History weiteres Skelettmaterial. Expeditionen während der folgenden zwei Sommer gruben mehr als 1000 Knochen aus, unter denen mindestens drei Individuen kleiner, fleischfressender Dinosaurier waren. Die Knochen wurden größtenteils unartikuliert vorgefunden, befanden sich also nicht mehr im Skelettverband, was es unmöglich machte, festzustellen, wie viele verschiedene Individuen vorlagen. Das Holotypus-Exemplar (YPM 5205) von Deinonychus umfasst daher nur einen vollständigen linken Fuß und Teile des rechten Fußes, die definitiv zum selben Individuum gehörten.[25] Die übrigen Fundstücke wurden im Peabody Museum of Natural History der Yale University unter 50 verschiedenen Inventarnummern katalogisiert.
Eine spätere Studie von John Ostrom und Grant E. Meyer analysierte dieses Material sowie Barnum Browns Daptosaurus und kam zu dem Schluss, dass es sich bei diesen Funden um ein und dieselbe Art handelt. John Ostrom veröffentlichte seine Ergebnisse 1969 und schrieb alle Überreste dem neuen Taxon Deinonychus antirrhopus zu. Deinonychus bedeutet so viel wie „schreckliche Klaue“ und weist auf die ungewöhnlich große, sichelartige Kralle am zweiten Zeh hin. Der Artname antirrhopus bedeutet „Gegengewicht“ und verweist auf John Ostroms Hypothese zur Funktion des Schwanzes.[25]
Obwohl bereits 1969 unzählige Knochen zur Verfügung standen, fehlten wichtige Teile des Skeletts oder waren schwierig zu interpretieren. So waren nur wenige postorbitale (hinter den Augen gelegene) Schädelknochen bekannt, und Oberschenkelknochen (Femur), Kreuzbein (Sacrum), Gabelbein (Furcula) und Brustbein (Sternum) fehlten ebenso wie einige Wirbel. Ein dünnes Knochenfragment beschrieb John Ostrom fälschlicherweise als Rabenbein (Coracoid). Seine erste Skelettrekonstruktion zeigt zudem ein sehr ungewöhnliches Schambein (Pubis), das trapezförmig, flach und so lang wie das Sitzbein (Ischium) war.
Weitere Entdeckungen

1974 veröffentlichte John Ostrom eine weitere Monographie, diesmal über den Schultergürtel von Deinonychus. Er stellte fest, dass das von ihm beschriebene Schambein tatsächlich ein Rabenbein war, ein Schulterelement.[26] Im selben Jahr wurde ein weiteres Deinonychus-Exemplar in Montana von einer Expedition der Harvard University ausgegraben, die von Farish Jenkins geleitet wurde. Dieses Exemplar zeigt eine Reihe von bis dahin unbekannten Skelettelementen, wie gut erhaltene Oberschenkelknochen, Schambein, ein Kreuzbein sowie ein besser erhaltenes Darmbein (Ilium), sowie Fuß- und Mittelhandknochen (Metatarsus). John Ostrom beschrieb dieses neue Exemplar und überarbeitete seine Rekonstruktion des Deinonychus-Skeletts. Diesmal zeigte die Rekonstruktion das sehr lange Schambein, das, so begann Ostrom zu spekulieren, wie bei Vögeln leicht nach hinten gebogen gewesen sein könnte.[27]
Das von Barnum Brown im Juli 1931 entdeckte Exemplar AMNH 3015 gilt heute als eines der vollständigsten Skelette. Einige kleine Kalksteinblöcke dieses Fundes mit Knochen und Knochenfragmenten wurden jedoch nicht präpariert und verblieben lange Zeit unbeachtet im Bestand des American Museums. Eine Untersuchung dieser Blöcke (Exemplarnummer AMNH 3015) von Gerald Grellet-Tinner und Peter Makovicky im Jahr 2000 brachte interessante, bislang übersehene Merkmale ans Licht. Einige lange, dünne Knochen, die zuerst für verknöcherte Sehnen gehalten wurden, waren tatsächlich Bauchrippen. Noch bedeutsamer ist die Entdeckung einer großen Zahl von fossilen Eierschalen sowie ein fragmentarisches Ei in der die Knochen umgebenden Gesteinsmatrix.[28]
In einer nachfolgenden, detaillierteren Studie schlossen Gerald Grellet-Tinner und Peter Makovicky, dass die Eierschalen fast mit Sicherheit zu Deinonychus gehörten; damit sind sie die ersten Überreste von Eiern, die von einem Dromaeosauriden identifiziert wurden.[23] Die Außenfläche einer der Eierschalen befand sich im engen Kontakt mit den Bauchrippen, was darauf hinweist, dass Deinonychus die Eier bebrütet haben könnte. Dies könnte zeigen, dass Deinonychus Körperwärme zum Bebrüten von Eiern eingesetzt hat, was eine Warmblütigkeit wie bei heutigen Vögeln bedeuten würde.[29] Weitere Untersuchungen von Gregory Erickson und Kollegen haben ergeben, dass dieses Individuum 13 oder 14 Jahre alt war, als es starb. Im Gegensatz zu anderen Theropoden, die in Verbindung mit Eiern oder Nestern entdeckt wurden, hatte das Deinonychus-Exemplar das Wachstum bereits beendet, als es starb.[30]
John Ostroms Beschreibung des Deinonychus im Jahr 1969 wurde als die wichtigste Einzelentdeckung der Dinosaurier-Paläontologie des mittleren 20. Jahrhunderts bezeichnet.[31] Die Entdeckung dieses eindeutig aktiven, flinken Prädators trug entscheidend zum Wandel der wissenschaftlichen und öffentlichen Auffassung zur Natur der Dinosaurier bei und entfachte Spekulationen, dass Dinosaurier gleichwarm gewesen sein könnten. Diese Entwicklung ist als Dinosaurier-Renaissance bekannt. Einige Jahre später stellte John Ostrom Ähnlichkeiten zwischen den Vordergliedmaßen von Deinonychus und denen der Vögel fest und schloss, dass Vögel von Dinosauriern abstammen.[32] Dreißig Jahre später wird diese Idee von fast allen Forschern akzeptiert.
Paläoenvironment und Faunengemeinschaft
Deinonychus bewohnte ein auen- oder sumpfähnliches Habitat.[5] So zeigen die Schichten der oberen Cloverly-Formation und der Antlers-Formation Hinweise auf subtropische Wälder, Flussdeltas und Lagunen.[33][34] Deinonychus teilte seinen Lebensraum mit dem gepanzerten Ankylosaurier Sauropelta sowie den Ornithopoden Zephyrosaurus und Tenontosaurus. Die Antlers-Formation in Oklahoma barg darüber hinaus den großen Theropoden Acrocanthosaurus, den riesigen Sauropoden Sauroposeidon, das Krokodil Goniopholis und den Knochenhecht Lepisosteus.[34] Die Potomac-Formation in Maryland, die einige möglicherweise zu Deinonychus gehörende Zähne barg, beherbergte den Sauropoden Astrodon und den Nodosaurier Priconodon, der lediglich von Zähnen bekannt ist.[35]
Paläobiologie
Jagdverhalten und Ernährung
Deinonychus-Zähne, die in Verbindung mit Fossilien des Ornithopoden Tenontosaurus gefunden werden, sind in der Cloverly-Formation relativ häufig. In zwei Steinbrüchen wurden Deinonychus-Exemplare zusammen mit Knochen des Tenontosaurus gefunden. Der erste dieser Steinbrüche, der Yale-Steinbruch in Montana (Cloverly-Formation), enthielt zahlreiche Zähne sowie die Überreste von fünf Deinonychus-Individuen und den Schwanz eines Tenontosaurus-Individuums. Diese Ansammlung von Individuen in einem einzigen Steinbruch könnte darauf hinweisen, dass Deinonychus am Tenontosaurus-Kadaver gefressen und diesen Pflanzenfresser vielleicht sogar gejagt hat. Ostrom und Maxwell (1995) vermuten sogar, dass Deinonychus in Gruppen gelebt und gejagt hat und dass Tenontosaurus das wichtigste Beutetier gewesen war.[36] Carpenter (1998) hält die Nachweise aus dem Yale-Steinbruch für zweifelhaft, weil von Tenontosaurus lediglich ein Schwanz gefunden wurde und weil die langen Knochen des Fundorts bevorzugt in nordwest- bis nordöstlicher Richtung ausgerichtet waren, was es möglich erscheinen lässt, dass die Knochen vor der Ablagerung transportiert wurden. Auch die relativ häufigen Funde von Deinonychus-Zähnen in Verbindung mit Tenontosaurus-Fossilien geben laut Carpenter keine Nachweise für eine aktive Jagd, da Deinonychus hier mit gleich großer Wahrscheinlichkeit lediglich als Aasfresser aufgetreten sein könnte.[37]

Paul Gignac und Kollegen beschrieben 2010 die Knochen eines Tenontosaurus, welcher Bissspuren aufweist, wahrscheinlich von Deinonychus stammend. Eine biomechanische Simulation kam zu dem Ergebnis, dass solche Bissspuren im Knochen eine Beißkraft von mindestens 3315 N erfordert haben. Gignac und Kollegen gehen außerdem von einer maximalen Beißkraft von mindestens 8200 N aus, womit Deinonychus eine sowohl absolut als auch im Verhältnis zur Körpergröße sehr große Beißkraft gehabt hätte. Allerdings finden sich selten Bissspuren des Theropoden in Knochen von Tenontosaurus, daher geht die Studie davon aus, dass Deinonychus nicht wie ein Aasfresser regelmäßig seine Bissspuren an Knochen hinterließ, sondern seinen starken Biss zur aktiven Jagd und Verteidigung einsetzte.[38]
In einem Steinbruch der Antlers-Formation in Oklahoma wurden sechs unterschiedlich große Teilskelette von Tenontosaurus zusammen mit einem Teilskelett und vielen Zähnen von Deinonychus gefunden. Ein Oberarmknochen (Humerus) von Tenontosaurus zeigt sogar Eindrücke, bei denen es sich um Bissspuren von Deinonychus handeln könnte. Brinkman und Kollegen (1998) stellen fest, dass Deinonychus 70 bis 100 Kilogramm schwer wurde, während ein erwachsener Tenontosaurus ein Gewicht von einer bis vier Tonnen hatte. Sie folgern, dass ein einzelner Deinonychus somit keinen erwachsenen Tenontosaurus hätte töten können, weshalb ein Jagen in Gruppen bei Deinonychus möglich sei.[39]
Roach und Brinkman (2007) sehen Deinonychus als solitären oder bestenfalls im lockeren Verband operierenden Jäger und fanden in den Deinonychus-Tenontosaurus-Fundstellen Hinweise auf ein Fressverhalten, das dem heutiger diapsider Reptilien gleicht. Versammeln sich beispielsweise Komodowarane um einen Kadaver, fressen die größten Individuen als erstes und attackieren kleinere Individuen, die dem Kadaver zu nahe kommen – falls das kleinere Tier dabei getötet und gefressen wird, handelt es sich um Kannibalismus. Roach und Brinkman geben an, dass das Verhalten der Komodowarane mit der Taphonomie der Deinonychus-Tenontosaurus-Fundstellen übereinstimmt. So stammen die unvollständigen Deinonychus-Überreste dieser Fundstellen von noch nicht ausgewachsenen Tieren, wobei das Fehlen von Skelettteilen anzeigt, dass andere Deinonychus an den Kadavern der getöteten Tiere gefressen haben.[40] Andererseits beschreiben Li und Kollegen (2007) fossile Fußspuren von Dromaeosauriden, die in parallelen Fährtenfolgen verlaufen, was diese Forscher als deutlichen Hinweis auf ein Leben in Gruppen deuten.[7]
Obwohl oft vermutet wird, dass Dromaeosauriden wie Deinonychus große Beutetiere wie Ornithopoden jagen konnten, ernährten sie sich wahrscheinlich wie heutige Raubtiere sowohl von kleinen als auch von großen Beutetieren.[41] Diese Annahme wurde jüngst durch die Entdeckung von Darminhalten in zwei Deinonychus-Skeletten bestätigt, die aus der Cloverly-Formation in Montana stammen. Dabei handelt es sich um kleine Klümpchen mit dicht gepacktem Knochenmaterial, das von Enzymen verätzt wurde. Während einige der stark fragmentierten Knochen wahrscheinlich von einem mittelgroßen Dinosaurier stammen, zeigen viele andere Fragmente wie beispielsweise eine kleine Vogelkralle, dass Deinonychus auch auf wesentlich kleinere Beute Jagd machte. Obwohl keine Gastrolithen (Magensteine) gefunden wurden, deuten die sehr stark fragmentierten Überreste darauf hin, dass im Magen Steine bei der Zerkleinerung der Nahrung halfen. Parsons hält Deinonychus für einen opportunistischen Räuber, der – wie die Vogelkralle zeigt – vielleicht in der Lage war, selbst flugfähige Beutetiere zu fangen.[42]
Funktion der Sichelkralle

Ostrom vermutete, dass Deinonychus Beutetiere mit seinen langen Armen festhalten konnte, um sie mit der Sichelkralle aufzuschlitzen.[1] Einige Forscher spekulierten, dass mit Hilfe der Sichelkralle die Eingeweide aus großen Ceratopsiern herausgerissen worden sein könnten.[43] Andere Studien ergaben, dass die Sichelkralle nicht zum Aufschlitzen, sondern zum gezielten Durchstechen der Halsadern oder der Luftröhre geeignet war.[37] Manning und Kollegen (2005) führten Versuche mit einem Robotermodell des Fußes von Deinonychus durch, indem sie die Auswirkungen von hydraulischen Stößen der Sichelkralle des Modells auf eine Schweinehaut untersuchten. Die Kralle machte in diesen Tests lediglich kleine rundliche Punktstiche und war nicht fähig, die Haut zu schneiden oder gar aufzuschlitzen. Stattdessen wurde die Haut unterhalb der Einstichlöcher stark zusammengeschoben, was der Kralle zusätzlichen Halt verschaffte. Die Forscher vermuten daher, dass die Kralle wie ein Steigeisen fungierte und dem Tier sicheren Halt auf großen Beutetieren verschaffte, während es mit seinen Zähnen eine Vielzahl von Wunden zufügen konnte.[44]
Ostrom verglich Deinonychus mit dem Strauß und dem Kasuar. Er bemerkte, dass diese Vögel mit ihrer verlängerten Kralle an der zweiten Zehe ernsthafte Verletzungen zufügen konnten.[1] Der Kasuar ist mit bis zu 12,5 Zentimeter langen Krallen ausgestattet.[45] Ostrom zitierte Gilliard (1958), der angab, dass Kasuare fähig seien, einen Arm abzutrennen oder einem Menschen die Eingeweide herauszureißen.[46] Kofron (1999 und 2003) studierte 241 dokumentierte Kasuar-Attacken, bei denen auch ein Mensch und zwei Hunde getötet wurden, fand aber keine Hinweise, dass ein Kasuar andere Tiere zerlegen oder aufschlitzen kann.[47][48]
Manning und Kollegen (2009) röntgten eine Handkralle von Velociraptor und verglichen sie mit Krallen des Uhus, um die Verteilung von Belastungen zu untersuchen, und schlussfolgerten, dass die Krallen von Dromaeosauriden deutliche Anpassungen an ein Klettern auf Bäumen zeigen.[49] Parsons (2009) vermutet ebenfalls, dass Deinonychus und andere Dromaeosauriden auch auf Bäumen gelebt haben könnten. Denn das vorletzte Zehenglied des die Sichelkralle tragenden zweiten Zehs hat einen Kiel, der sich in ähnlicher Form auch bei heutigen Spechten findet. Auch die Krümmung der Sichelkralle entspricht der Krümmung der Krallen der Spechte. Diese Krümmung ist bei den einzelnen Individuen sehr unterschiedlich. So hat das von Ostrom 1969 beschriebene Typusexemplar (YPM 5205) eine stark gekrümmte Sichelkralle, während ein neueres, 1976 beschriebenes Exemplar (MCZ 4371) eine Sichelkralle mit nur schwach entwickelter Krümmung hat, die mehr den „normalen“ Klauen der übrigen Zehen ähnelt.[27] Parsons vermutet, dass es sich um altersspezifische Variationen handelt und dass der Grad der Krümmung mit zunehmendem Alter abnimmt. So könnten junge Deinonychus mit noch stark gekrümmter Sichelkralle baumbewohnend gewesen sein, während Individuen mit zunehmendem Alter zu einer mehr terrestrischen Lebensweise wechselten.[50]
Funktion der Arme
Biomechanische Studien von Kenneth Carpenter (2002) bestätigen, dass die langen Arme während der Jagd zum Greifen und Festhalten von Beutetieren benutzt wurden. Weiter untermauert wird diese Interpretation durch das große und verlängerte Rabenbein, das auf starke Armmuskeln hinweist.[51] Carpenter zeigt mithilfe von Knochennachbildungen, dass Deinonychus nicht in der Lage war, seine Arme wie ein Vogel gegen den Körper zu falten, und widerspricht damit früheren Beobachtungen von Jacques Gauthier (1985)[52] und Gregory Paul (1988).[2]
Wie Studien von Phil Senter (2006) zeigen, konnte Deinonychus Objekte zwischen den Handflächen halten und tragen, jedoch nur mit gebeugtem Handgelenk. Durch eine Streckung des Handgelenks rotierten die Handflächen zwangsläufig nach oben (Supination), wodurch beispielsweise lebende Beute hätte entkommen können. Des Weiteren konnte Deinonychus Objekte durch Drücken gegen die Brust einhändig halten und tragen. Falls die Arme von Deinonychus wie bei Velociraptor mit langen Federn ausgestattet waren, hätten sie die Beweglichkeit der Arme zu einem gewissen Grad eingeschränkt. Stochern nach kleinen Beutetieren in Spalten hätte beispielsweise bei geflügelten Armen nur mit dem senkrecht zum Flügel ausgestreckten dritten Finger vorgenommen werden können. Senter hält ein derartiges Stochern nach Beutetieren für unwahrscheinlich, aber nicht für ausgeschlossen.[41]
Parsons zeigte, dass juvenile und subadulte Deinonychus-Exemplare einige morphologische Unterschiede zu erwachsenen Tieren zeigen. So waren die Arme der jüngeren Individuen proportional länger als die der erwachsenen, was ein Hinweis auf Unterschiede im Verhalten zwischen Jungtieren und erwachsenen Tieren sein könnte.[53]
Geschwindigkeit
Dromaeosauriden, insbesondere Deinonychus, werden in den populären Medien oft als ungewöhnlich schnelle Tiere dargestellt. John Ostrom vermutete in seiner Erstbeschreibung, dass Deinonychus „moderately, but not unusually fast” („mäßig, aber nicht ungewöhnlich schnell“) war.[25] Diese Vermutung basierte auf der Annahme, dass Oberschenkelknochen und Schienbein etwa gleich lang waren, obwohl ihm bei der Erstbeschreibung noch kein Oberschenkelknochen vorlag. Erst 1976 konnte ein Skelett mit Oberschenkelknochen (MCZ 4371) beschrieben werden. Wie Ostrom nun bemerkte, war der Oberschenkelknochen tatsächlich um zehn Prozent kürzer als das Schienbein – ähnlich wie bei dem an schnelles Laufen angepassten Ornithomimosaurier Struthiomimus. Gleichzeitig merkte er jedoch an, dass das Längenverhältnis zwischen Mittelfuß (Metatarsus) und Schienbein für Geschwindigkeitsschätzungen aussagekräftiger ist als das von ihm zuvor herangezogene Oberschenkel-Schienbein-Verhältnis. Beim Strauß ist das Mittelfuß-Schienbein-Verhältnis 0,95, während Struthiomimus ein Verhältnis von 0,68 aufweist. Deinonychus jedoch hatte einen kurzen Mittelfuß und ein Verhältnis von lediglich 0,48. Ostrom schlussfolgerte, “the only reasonable conclusion is that Deinonychus was not the most fleet-footed of theropods“ („die einzige akzeptable Erklärung ist, dass Deinonychus nicht der schnellste unter den Theropoden war“).[27]
Ostrom vermutete, dass das Hinterbein mit der Sichelkralle als Waffe benutzt wurde, um Beutetiere aufzuschlitzen. Der kurze Mittelfuß könnte laut Ostrom dabei eine Anpassung gewesen sein, um die Belastung der Beinknochen während einer derartigen Attacke zu reduzieren. Die Anordnung von Muskel-Ansatzstellen in den Beinknochen unterschied sich ebenfalls von der anderer Theropoden – als Erklärung gab Ostrom an, dass beim Angriff auf Beutetiere andere Muskeln benutzt worden seien als beim normalen Laufen. Ostrom schlussfolgerte daher, dass die Beine von Deinonychus einen Kompromiss repräsentieren, da sie einerseits ein möglichst schnelles Laufen erlauben, andererseits Belastungen während einer Attacke minimieren sollen.[27]
Das 1974 entdeckte Exemplar (MCZ 4371) war größer als das sehr vollständige, 1931 entdeckte Exemplar (AMNH 3015). Wie Ostrom (1976) bemerkte, war die Größenzunahme nicht einheitlich: So war der Arm bei MCZ 4371 um 20 %, das Schienbein um 18 %, der Mittelfuß jedoch nur um 12 % länger als bei AMNH 3015. Dies zeigt, dass der Mittelfuß mit zunehmendem Alter proportional kürzer wurde, was andeutet, dass die Fähigkeit zum schnellen Laufen mit fortschreitendem Alter der Tiere abnahm.[27]
Richard Kool untersuchte fossile Fährtenfolgen aus der Gething-Formation in der kanadischen Provinz British Columbia und schätzte die Geschwindigkeit ab, mit der verschiedene Tiere gelaufen waren, als sie die Fußspuren hinterließen. Kool schätzte für eine Fährtenfolge der Ichnogattung Irenichnites gracilis, die evtl. von Deinonychus stammen könnte, dass das Tier mit 10,1 km/h gegangen war.[54] Eine biomechanische Studie von Sellers und Manning (2007) schätzt die Höchstgeschwindigkeit des verwandten Velociraptor auf 10,8 m/s (38,9 km/h).[55]
Eier

Grellet-Tinner und Makovicky (2000) entdeckten Eierschalen sowie ein fragmentarisches Ei in Gesteinsblöcken, die zu dem bereits 1931 von Barnum Brown entdeckten Deinonychus-Exemplar AMNH 3015 gehören, aber nicht präpariert wurden. Das Ei befindet sich im Kontakt mit Bauchrippen des Deinonychus-Exemplars und wurde wahrscheinlich von diesem gelegt. Die Forscher schließen aus, dass das Ei lediglich eine Mahlzeit des Deinonychus war, weil die Eifragmente zwischen den Bauchrippen und den Armknochen lagen und somit kein Mageninhalt sein konnten. Des Weiteren zeigt die Art und Weise, wie das Ei zerdrückt und fragmentiert wurde, dass es noch intakt war, als es von Sedimenten begraben wurde, und erst durch den Fossilationsprozess verformt wurde. Auch die Möglichkeit, dass das Ei zufällig mit dem Skelett in Verbindung kam, wird als unwahrscheinlich eingeschätzt. So wurden die umgebenden Knochen nicht zerstreut, sondern verblieben artikuliert in ihren ursprünglichen Positionen, was zeigt, dass der Fundort nicht nachträglich gestört wurde. Die Bauchrippen, die nur sehr selten artikuliert gefunden werden, unterstützen diese Interpretation. Grellet-Tinner und Makovicky geben an, dass alle Hinweise darauf schließen lassen, dass das Ei unter dem Körper des Deinonychus lag, als es begraben wurde. Der Fund könnte einen brütenden Deinonychus über seinem Ei zeigen; auch möglich ist, dass sich das Ei noch im Legedarm (Oviduct) befand.[23]
Eine Untersuchung der Mikrostruktur des Eis bestätigt, dass es von einem Theropoden stammt. Verglichen mit anderen maniraptoren Theropoden ist das Ei denen der Oviraptoriden ähnlicher als denen der Troodontiden, obwohl Troodontiden enger mit Dromaeosauriden verwandt sind. Die Größe des Eis konnte wegen der starken Verformung nicht ermittelt werden. Die Forscher untersuchten jedoch die Breite des Beckenkanals, den das Ei passieren musste, und schätzten daraus den Durchmesser auf etwa 7 Zentimeter. Die Schätzung wird dadurch unterstützt, dass die größten Eier des Oviraptoriden Citipati mit 7,2 Zentimeter Durchmesser ähnlich groß waren, und dass erwachsene Citipati und Deinonychus etwa gleich groß wurden. Des Weiteren ist die Dicke der Eierschalen von Citipati fast identisch mit der von Deinonychus, und da die Dicke der Eierschale mit dem Eivolumen korreliert, ist dies ein weiteres Anzeichen dafür, dass die Eier beider Spezies etwa die gleiche Größe hatten.[23]
In der Populärkultur
Deinonychus gehört zusammen mit Utahraptor und Velociraptor zu den populärsten Dromaeosauriden und ist Darsteller in Filmen, Fernsehshows, Kinderbüchern, Comics und Videospielen. Beispielsweise taucht Deinonychus in der britischen Science-Fiction-Fernsehserie Primeval – Rückkehr der Urzeitmonster (ausgestrahlt seit 2007)[56] sowie als Actionfigur und als Darsteller in den Comics und Zeichentrickserien der Dino-Riders-Serie aus den späten 1980er-Jahren auf[57] und lieferte auch die Vorlage für die dort so genannten Velociraptoren in Jurassic Park.[58] Des Weiteren ist der Deinonychus einer der Hauptsaurier in den Carnosaurus-Filmen. Die niederländische Doom-Metal-Band Deinonychus (1992–2008) benannte sich nach diesem Dinosaurier.[59]
Montierte Skelettabgüsse von Deinonychus finden sich in Naturkundemuseen auf der ganzen Welt. Im American Museum of Natural History in New York ist das einzige montierte Skelett zu besichtigen, das zum Teil aus echten Knochen aufgebaut ist.[60] Diese originalen Knochen gehören zu dem Exemplar AMNH 3015, das Barnum Brown bereits 1931 entdeckte und das bis heute eines der vollständigsten Deinonychus-Skelette ist. Selbst diesem montierten Skelett fehlen Elemente wie Brustbein, Brustbein-Rippen, Gabelbein und Bauchrippen.
Literatur
- John H. Ostrom: A new theropod dinosaur from the Lower Cretaceous of Montana. In: Peabody Museum of Natural History, Yale University. Postilla. Bd. 128, 1969, ISSN 0079-4295, S. 1–17.
- John H. Ostrom: Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana (= Peabody Museum of Natural History. Bulletin. Bd. 30, ISSN 0079-032X). Peabody Museum of Natural History – Yale University, New Haven CT 1969.
- John H. Ostrom: Stratigraphy and paleontology of the Cloverly Formation (Lower Cretaceous) of the Bighorn Basin area, Wyoming and Montana (= Peabody Museum of Natural History. Bulletin. Bd. 35). Peabody Museum of Natural History – Yale University, New Haven CT 1970.
- John H. Ostrom: On a new specimen of the Lower Cretaceous theropod dinosaur Deinonychus antirrhopus (= Breviora. Nr. 439, ISSN 0006-9698). Museum of Comparative Zoology, Cambridge MA 1976.
- W. Desmond Maxwell, John H. Ostrom: Taphonomy and paleobiological implications of Tenontosaurus-Deinonychus associations. In: Journal of Vertebrate Paleontology. Bd. 15, Nr. 4, 1995, ISSN 0272-4634, S. 707–712, doi:10.1080/02724634.1995.10011256.
- Gerald Grellet-Tinner, Peter Makovicky: A possible egg of the dromaeosaur Deinonychus antirrhopus: phylogenetic and biological implications. In: Canadian Journal of Earth Sciences. Bd. 43, Nr. 6, 2006, ISSN 0008-4077, S. 705–719, doi:10.1139/e06-033.
Einzelnachweise
- John H. Ostrom: Stratigraphy and paleontology of the Cloverly Formation (Lower Cretaceous) of the Bighorn Basin area, Wyoming and Montana (= Peabody Museum of Natural History. Bulletin. Bd. 35). Peabody Museum of of Natural History – Yale University, New Haven CT 1970.
- Gregory S. Paul: Predatory Dinosaurs of the World. A Complete Illustrated Guide. Simon & Schuster, New York NY 1988, ISBN 0-671-61946-2, S. 366–369.
- John H. Ostrom: Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana (= Peabody Museum of Natural History. Bulletin. Bd. 30, ISSN 0079-032X). Peabody Museum of of Natural History – Yale University, New Haven CT 1969.
- William D. Maxwell, Lawrence M. Witmer: New Material of Deinonychus (Dinosauria, Theropoda). In: Journal of Vertebrate Paleontology. Bd. 16, Supplement to Nr. 3 = Abstracts of Papers Fifty-Sixth Annual Meeting. Society of Vertebrate Paleontology. American Museum of Natural History, New York, New York October 16–19, 1996, 1996, ISSN 0272-4634, S. 51A.
- Mark A. Norell, Peter J. Makovicky: Dromaeosauridae. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 196–209.
- Lawrence M. Witmer, William D. Maxwell: The skull of Deinonychus (Dinosauria: Theropoda): New insights and implications. In: Journal of Vertebrate Paleontology. Bd. 16, Supplement to Nr. 3 = Abstracts of Papers Fifty-Sixth Annual Meeting. Society of Vertebrate Paleontology. American Museum of Natural History, New York, New York October 16–19, 1996, 1996, S. 73A.
- Rihui Li, Martin G. Lockley, Peter J. Makovicky, Masaki Matsukawa, Mark A. Norell, Jerald D. Harris, Mingwei Liu: Behavioral and faunal implications of Early Cretaceous deinonychosaur trackways from China. In: Die Naturwissenschaften. Bd. 95, Nr. 3, 2008, S. 185–191, doi:10.1007/s00114-007-0310-7.
- Mark A. Norell, Peter J. Makovicky: Important features of the dromaeosaurid skeleton. 2, Information from newly collected specimens of Velociraptor mongoliensis (= American Museum Novitates. Nr. 3282, ISSN 0003-0082). American Museum of Natural History, New York NY 1999, online.
- Xing Xu, Zhonghe Zhou, Xiaolin Wang, Xuewen Kuang, Fucheng Zhang, Xiangke Du: Four-winged dinosaurs from China. In: Nature. Bd. 421, Nr. 6921, 2003, S. 335–340, doi:10.1038/nature01342.
- Sunny H. Hwang, Mark A. Norell, Ji Qiang, Gao Keqin: New specimens of Microraptor zhaoianus (Theropoda: Dromaeosauridae) from northeastern China (= American Museum Novitates. Nr. 3381). American Museum of Natural History, New York NY 2002, online.
- Alan H. Turner, Peter J. Makovicky, Mark A. Norell: Feather Quill Knobs in the Dinosaur Velociraptor. In: Science. Bd. 317, Nr. 5845, 2007, S. 1721, doi:10.1126/science.1145076.
- Mark A. Norell, James M. Clark, Alan H. Turner, Peter J. Makovicky, Rinchen Barsbold, Timothy Rowe: A new dromaeosaurid theropod from Ukhaa Tolgod (Ömnögov, Mongolia) (= American Museum Novitates. Nr. 3545). American Museum of Natural History, New York NY 2006, online.
- Alan H. Turner, Sunny H. Hwang, Mark A. Norell: A small derived theropod from Öösh, Early Cretaceous, Baykhangor Mongolia (= American Museum Novitates. Nr. 3557). American Museum of Natural History, New York NY 2007, online.
- Ринченгийн Барсболд: Хищные динозавры мела Монголии. Совместная Советско-Монгольская палеонтологическая экспедиция (= Труды. 19). Наука, Москва 1983 (In englischer Sprache: Rinchen Barsbold: Carnivorous dinosaurs from the Cretaceous of Mongolia. The Joint Soviet-Mongolian Paleontological Expedition (= Transaction. Bd. 19). Selbstverlag des Verfassers, Berkeley CA 1983, online).
- Gregory S. Paul: Predatory Dinosaurs of the World. A Complete Illustrated Guide. Simon & Schuster, New York NY 1988, ISBN 0-671-61946-2, S. 358.
- Philip J. Currie: New information on the anatomy and relationships of Dromaeosaurus albertensis (Dinosauria: Theropoda). In: Journal of Vertebrate Paleontology. Bd. 15, Nr. 3, 1995, S. 576–591, doi:10.1080/02724634.1995.10011250.
- Nicholas R. Longrich, Philip J. Currie: A microraptorine (Dinosauria–Dromaeosauridae) from the Late Cretaceous of North America. In: Proceedings of the National Academy of Sciences of the United States of America. Bd. 106, Nr. 13, 2009, doi:10.1073/pnas.0811664106, Digitalisat (PDF;763,02 kB).
- Michael J. Benton: Vertebrate Palaeontology. 3. Auflage. Blackwell, Malden MA 2005, ISBN 0-632-05637-1, S. 472, Kladogramm (Memento vom 19. Oktober 2008 im Internet Archive).
- Daniel L. Brinkman, Richard L. Cifelli, Nicholas J. Czaplewski: First occurrence of Deinonychus antirrhopus (Dinosauria: Theropoda) from the Antlers Formation (Lower Cretaceous: Aptian-Albian) of Oklahoma (= Oklahoma Geological Survey Bulletin. Bd. 146, ISSN 0078-4389). University of Oklahoma, Norman OK 1998, Digitalisat (PDF; 2,13 MB).
- Zi-Qiang Chen, Susan Lubin: A Fission-Track Study of the Terrigenous Sedimentary Sequences of the Morrison and Cloverly Formations in the Northeastern Bighorn Basin, Wyoming. In: The Mountain Geologist. Bd. 34, Nr. 2, 1997, ISSN 0027-254X, S. 51–62.
- Darrin Burton, Brent W. Greenhalgh, Brooks B. Britt, Bart J. Kowallis, William S. Elliott, Reese Barrick: New radiometric ages from the Cedar Mountain Formation, Utah and the Cloverly Formation, Wyoming: implications for contained dinosaur faunas. In: Geological Society of America Abstracts with Programs. Bd. 38, Nr. 7, 2006, ISSN 0016-7592, S. 52 (Zusammenfassung).
- Thomas R. Lipka: Thomas R. Lipka. In: The Affinities of the Enigmatic Theropods of the Arundel Clay Facies (Aptian), Potomac Formation, Atlantic Coastal Plain of Maryland. In: Spencer G. Lucas, James I. Kirkland, John W. Estep (Hrsg.): Lower and Middle Cretaceous terrestrial ecosystems (= New Mexico Museum of Natural History and Science. Bulletin. Bd. 14, ISSN 1524-4156). New Mexico Museum of Natural History and Science, Albuquerque NM 1998, S. 229–234, online.
- Gerald Grellet-Tinner, Peter Makovicky: A possible egg of the dromaeosaur Deinonychus antirrhopus: phylogenetic and biological implications. In: Canadian Journal of Earth Sciences. Bd. 43, Nr. 6, 2006, ISSN 0008-4077, S. 705–719, doi:10.1139/e06-033.
- Mark A. Norell, Eugene S. Gaffney, Lowell Dingus: Discovering dinosaurs in the American Museum of Natural History. Knopf, New York NY 1995, ISBN 0-679-43386-4, S. 126–130.
- John H. Ostrom: A new theropod dinosaur from the Lower Cretaceous of Montana. In: Peabody Museum of Natural History, Yale University. Postilla. Bd. 128, 1969, ISSN 0079-4295, S. 1–17.
- John H. Ostrom: The Pectoral Girdle and Forelimb Function of Deinonychus (Reptilia: Saurischia): A Correction. In: Peabody Museum of Natural History, Yale University. Postilla. Bd. 165, 1974, S. 1–11.
- John H. Ostrom: On a new specimen of the Lower Cretaceous theropod dinosaur Deinonychus antirrhopus (= Breviora. Nr. 439, ISSN 0006-9698). Museum of Comparative Zoology, Cambridge MA 1976.
- Peter J. Makovicky, Gerald Grellet-Tinner: Association between a specimen of Deinonychus antirrhopus and theropod eggshell. In: Ana M. Bravo, Teresa Reyes (Hrsg.): Extended abstracts. 1r Congrés Internacional sobre Ous i Cries de Dinosaures. = Primer Congreso Internacional sobre Huevos y Crías de Dinosaurios. = First International Symposium on Dinosaur Eggs and Babies. Isona i Conca Dellà, Catalonia, Spain, 23–26 setembre 1999. s. n., Isona i Conca Dellà 2000, S. 123–128.
- Gerald Grellet-Tinner: Oology and the evolution of thermophysiology in saurischian dinosaurs: homeotherm and endotherm deinonychosaurians? In: Papeis Avulsos de Zoologia. Bd. 46, Nr. 1, 2006, ISSN 0031-1049, S. 1–10, doi:10.1590/S0031-10492006000100001.
- Gregory M. Erickson, Kristina Curry Rogers, David J. Varricchio, A. Mark Norell, Xing Xu: Growth patterns in brooding dinosaurs reveals the timing of sexual maturity in non-avian dinosaurs and genesis of the avian condition. In: Biology Letters. Bd. 3, Nr. 5, 2007, ISSN 1744-9561, S. 558–561, doi:10.1098/rsbl.2007.0254, (PDF; 213 kB).
- David E. Fastovsky, David B. Weishampel: Theropoda I: Nature Red in Tooth and Claw. In: David E. Fastovsky, David B. Weishampel: The Evolution and Extinction of the Dinosaurs. 2nd edition. Cambridge University Press, Cambridge u. a. 2005, ISBN 0-521-01046-2, S. 265–299.
- John H. Ostrom: Archaeopteryx and the origin of birds. In: Biological Journal of the Linnean Society. Bd. 8, Nr. 2, 1976, ISSN 0024-4066, S. 91–182, doi:10.1111/j.1095-8312.1976.tb00244.x.
- Catherine A. Forster: The paleoecology of the ornithopod dinosaur Tenontosaurus tilletti from the Cloverly Formation, Big Horn Basin of Wyoming and Montana. In: The Mosasaur. Bd. 2, 1984, ISSN 0736-3907, S. 151–163.
- Mathew J. Wedel, Richard L. Cifelli: Sauroposeidon: Oklahoma's Native Giant. In: Oklahoma Geology Notes. Bd. 65, Nr. 2, 2005, ISSN 0030-1736, S. 40–57, Digitalisat (PDF; 2,15 MB).
- David B. Weishampel, Paul M. Barrett, Rodolfo A. Coria, Jean Le Loeuff, Xing Xu, Xi-Jin Zhao, Ashok Sahni, Elizabeth M. P. Gomani, Christopher R. Noto: Dinosaur Distribution. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd Edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 517–606, hier S. 556.
- W. Desmond Maxwell, John H. Ostrom: Taphonomy and paleobiological implications of Tenontosaurus-Deinonychus associations. In: Journal of Vertebrate Paleontology. Bd. 15, Nr. 4, 1995, S. 707–712, doi:10.1080/02724634.1995.10011256.
- Kenneth Carpenter: Evidence of predatory behavior by carnivorous dinosaurs. In: Gaia. Revista de Geociências. Nr. 15, 1998, ISSN 0871-5424, S. 135–144, Digitalisat (PDF; 3,1 MB).
- Paul M. Gignac, Peter J. Makovicky, Gregory M. Erickson, Robert P. Walsh: A description of Deinonychus antirrhopus bite marks and estimates of bite force using tooth indentation simulations. In: Journal of Vertebrate Palaeontology. Bd. 30, Nr. 4, 2010, S. 1169–1177, doi:10.1080/02724634.2010.483535.
- Daniel L. Brinkman, Richard L. Cifelli, Nicholas J. Czaplewski: First occurrence of Deinonychus antirrhopus (Dinosauria: Theropoda) from the Antlers Formation (Lower Cretaceous: Aptian-Albian) of Oklahoma (= Oklahoma Geological Survey Bulletin. Bd. 146). University of Oklahoma, Norman OK 1998, S. 27, Digitalisat (PDF; 2,13 MB).
- Brian T. Roach, Daniel L. Brinkman: A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs. In: Bulletin of the Peabody Museum of Natural History. Bd. 48, Nr. 1, 2007, S. 103–138, doi:10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2.
- Phil Senter: Comparison of Forelimb Function Between Deinonychus and Bambiraptor (Theropoda: Dromaeosauridae). In: Journal of Vertebrate Paleontology. Bd. 26, Nr. 4, 2006, S. 897–906, doi:10.1671/0272-4634(2006)26[897:COFFBD]2.0.CO;2.
- William L. Parsons, Kristen M. Parsons: Gut contents of a lower cretaceous dromaeosaurid dinosaur: new data on the feeding behavior of Deinonychus antirrhopus (Saurischia, Theropoda). In: Society of Vertebrate Paleontology Annual Conference Abstract. 2007, Zusammenfassung (Memento vom 3. Juli 2010 im Internet Archive).
- Dawn Adams: The bigger they are, the harder they fall: Implications of ischial curvature in ceratopsian dinosaurs. In: Philip J. Currie, Emlyn H. Koster (Hrsg.): Fourth symposium on mesozoic terrestrial ecosystems, Drumheller, August 10–14, 1987. Short Papers (= Occasional Paper of the Tyrrell Museum of Palaeontology. Nr. 3, ZDB-ID 1217807-x). Tyrrell Museum of Palaeontology, Drumheller 1987, S. 1–6.
- Phillip L. Manning, David Payne, John Pennicott, Paul M. Barrett, Roland A. Ennos: Dinosaur killer claws or climbing crampons? In: Biology Letters. Bd. 2, Nr. 1, 2006, S. 110–112, doi:10.1098/rsbl.2005.0395.
- Stephen J. J. F. Davies: Ratites and Tinamous. Tinamidae, Rheidae, Dromaiidae, Casuariidae, Apterygidae, Struthionidae (= Bird Families of the World. Bd. 9). Oxford University Press, Oxford u. a. 2002, ISBN 0-19-854996-2.
- E. Thomas Gilliard: Living Birds of the World. Doubleday, Garden City NY 1958.
- Christopher P. Kofron: Attacks to humans and domestic animals by the southern cassowary (Casuarius casuarius johnsonii) in Queensland, Australia. In: Journal of Zoology. Bd. 249, Nr. 4, 1999, ISSN 0022-5460, S. 375–381, doi:10.1111/j.1469-7998.1999.tb01206.x.
- Christopher P. Kofron: Case histories of attacks by the southern cassowary in Queensland. In: Memoirs of the Queensland Museum. Bd. 49, Nr. 1, 2003, ISSN 0079-8835, S. 335–338.
- Phillip L. Manning, Lee Margetts, Mark R. Johnson, Philip J. Withers, William I. Sellers, Peter L. Falkingham, Paul M. Mummery, Paul M. Barrett, David R. Raymont: Biomechanics of Dromaeosaurid Dinosaur Claws: Application of X-Ray Microtomography, Nanoindentation, and Finite Element Analysis. In: The Anatomical Record. AR. Advances in integrative Anatomy and evolutionary Biology. Bd. 292, Nr. 9, 2009, ISSN 1932-8486, S. 1397–1405, doi:10.1002/ar.20986.
- William L. Parsons, Kristen M. Parsons: Further descriptions of the osteology of Deinonychus antirrhopus (Saurischia, Theropoda) (Memento vom 3. Juli 2010 im Internet Archive). In: Bulletin of the Buffalo Society of Natural Sciences. Bd. 38, 2009, ISSN 0096-4131, S. 43–54, (PDF; 449 kB).
- Kenneth Carpenter: Forelimb biomechanics of nonavian theropod dinosaurs in predation. In: Senckenbergiana Lethaea. Bd. 82, Nr. 1, 2002, ISSN 0037-2110, S. 59–76, doi:10.1007/BF03043773.
- Jaques Gauthier, Kevin Padian: Phylogenetic, Functional, and Aerodynamic Analyses of the Origin of Birds and Their Flight. In: Max K. Hecht, John H. Ostrom, Günter Viohl, Peter Wellnhofer: The Beginnings of Birds. Proceedings of the International Archaeopteryx Conference, Eichstätt 1984. Freunde des Jura-Museums Eichstätt, Eichstätt 1985, ISBN 3-9801178-0-4, S. 185–197.
- William L. Parsons, Kristen M. Parsons: Morphology and Size of an Adult Specimen of Deinonychus antirrhopus, (Saurischia, Theropoda). In: Journal of Vertebrate Paleontology. Bd. 26, Supplement to Nr. 3 = Abstracts of Papers. Sixty-Sixth Annual Meeting Society of Vertebrate Paleontology. Canadian Museum of Nature. Marriott Ottawa Crowne Plaza Ottawa, Ottawa, Ontario, Canada October 18–21, 2006, 2006, S. 109A.
- Richard Kool: The walking speed of dinosaurs from the Peace River Canyon, British Columbia, Canada. In: Canadian Journal of Earth Sciences. Bd. 18, Nr. 4, 1981, S. 823–825, doi:10.1139/e81-077.
- William Irvin Sellers, Phillip Lars Manning: Estimating dinosaur maximum running speeds using evolutionary robotics. In: Proceedings of the Royal Society. Series B: Biological Sciences. Bd. 274, Nr. 1626, 2007, ISSN 0962-8452, S. 2711–2716, doi:10.1098/rspb.2007.0846, online.
- Deinonychus. In: Primeval-Wiki. Abgerufen am 3. August 2014 (englisch).
- DinoRiders Rulon Deinonychus. In: toyarchive.com. Abgerufen am 3. August 2014 (englisch).
- Brian Switek: You say „Velociraptor,“ I say „Deinonychus“. Abgerufen am 3. August 2014 (englisch).
- Deinonychus bei BNR Metal (englisch), abgerufen am 31. Mai 2021
- Deinonychus. In: American Museum of Natural History. Archiviert vom Original am 27. April 2009; abgerufen am 3. August 2014 (englisch).
Weblinks
- Ein Zwischenspiel: Deinonychus - Streit um die Vorfahren des Archaeopteryx. Von Nadja Podbregar, Stand 13. August 2010