Luftsack (Vogel)
Die Luftsäcke (Sacci pneumatici) der Vögel sind dünnwandige Anhänge der Lunge, die wie Blasebälge die Luft durch die Lunge führen. In ihnen findet jedoch kein Gasaustausch statt. Es handelt sich um hauchdünne Säcke mit einer durchsichtigen Wand. Neben ihrer Funktion als „Motor der Atmung“ sind sie auch an der Stimmbildung beteiligt. Hochfrequente Exspirationen (Ausatmungen) werden im Stimmkopf (Syrinx) zum Vogelgesang moduliert. Die dritte wichtige Funktion der Luftsäcke ist die Beteiligung an der Thermoregulation durch die Wärmeabgabe über Verdunstung.
Anatomie
Gewebe
Die Luftsäcke sind größtenteils von einem einschichtigen Plattenepithel ausgekleidet. Teilweise treten auch kubische und hochprismatische Flimmerepithelzellen auf. In ihre Wand sind elastische Fasern und glatte Muskulatur eingelagert. Darin verlaufen auch einzelne Nervenfasern, bei denen bei einigen Vögeln der Ursprung aus dem Nervus vagus beschrieben ist, über deren Funktion bislang aber keine Kenntnisse vorliegen.
Einteilung der Luftsäcke
Die Luftsäcke werden während der Entwicklung bereits sehr früh im Embryo angelegt. Sie entstehen beim Huhn am 5. und 6. Embryonaltag, gleichzeitig mit der Entwicklung der Lungen, aber vor der Bildung der Sekundärbronchien. Am 10. Tag sind die Luftsäcke bereits vollständig entwickelt und zeigen danach nur noch ein Größenwachstum.
Embryonal werden sechs Luftsackpaare angelegt, von denen zwei Paare bei fast allen Vögeln (Ausnahme Haubentaucher) in der weiteren Entwicklung zum einheitlichen Schlüsselbeinluftsack verschmelzen. Beim Truthuhn werden nur fünf Luftsackpaare angelegt.
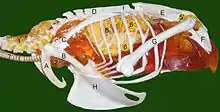
1 Halsluftsack, 2 Schlüsselbeinluftsack,
3 vorderer und 4 hinterer Brustluftsack,
5 Bauchluftsack (5' Divertikel in den Beckengürtel), 6 Lunge, 7 Luftröhre
Knochen:
A Schlüsselbein, B Rabenbein,
C Schulterblatt, D Notarium, E Synsacrum,
F Becken, G Oberschenkelknochen,
H Brustbein
- Der linke und rechte Halsluftsack (Saccus cervicalis) liegt entlang der Halswirbelsäule und besitzt Aussackungen (Divertikel) inner- und außerhalb der Halswirbel. Bei Hühnervögeln verschmelzen beide Halsluftsäcke zu einer medianen Hauptkammer. Der Haubentaucher hat keine Halsluftsäcke.
- Der Schlüsselbeinluftsack (Saccus clavicularis) besteht ursprünglich aus zwei Luftsackpaaren, die bei den Vögeln (mit Ausnahme des Haubentauchers) median zu einem unpaaren Sack verschmelzen. Vom Schlüsselbeinluftsack gehen Divertikel innerhalb und außerhalb des Brustkorbs aus. Letztere erstrecken sich auch in die Knochen des Schultergürtels und den Oberarmknochen. Diese Knochen sind somit bei Vögeln luftgefüllt (pneumatisiert). Beim Strauß ist der Oberarmknochen nicht pneumatisiert. Die funktionelle Bedeutung dieser Pneumatisation ist noch nicht endgültig geklärt.
- Die vorderen Brustluftsäcke (Sacci thoracici craniales) liegen zwischen zwei Bindegewebsmembranen (Septum horizontale und obliquum) innerhalb des Brustkorbs und umfassen das Herz und den Drüsenmagen (Proventriculus). Bei Singvögeln verschmelzen sie mit dem einheitlichen Schlüsselbeinluftsack.
- Die hinteren Brustluftsäcke (Sacci thoracici caudales) liegen direkt der Körperwand an und befinden sich hinter den vorderen Brustluftsäcken. Truthühner haben keine (werden embryonal auch nicht angelegt), Hühner nur kleine hintere Brustluftsäcke. Bei Störchen sind sie geteilt, so dass sie 4 hintere Brustluftsäcke haben.
- Die Bauchluftsäcke (Sacci abdominales) liegen als dünne Ballons zwischen den Darmschlingen und pneumatisieren auch den Beckengürtel. Bei Papageien, Straußen und Sperlingsvögeln sind die Bauchluftsäcke relativ klein.
Zahl der Luftsäcke
Die Vögel besitzen maximal 12 Luftsäcke, aufgrund der Verschmelzung der vier Schlüsselbeinluftsäcke aber im Regelfall nur 9. Die artspezifische Zahl schwankt zwischen 7 und 11.
| Anzahl der Luftsäcke bei einigen Vögeln | ||
|---|---|---|
| 11 | Störche | hintere Brustluftsäcke nochmals unterteilt (also 5 Paare + Schlüsselbeinluftsack) |
| 10 | Haubentaucher | 4 Schlüsselbeinluftsäcke, aber keine Halsluftsäcke (also 5 Paare) |
| 9 | Regelfall | 4 Paare + einheitlicher Schlüsselbeinluftsack |
| 8 | Haushuhn | Hals- und Schlüsselbeinluftsack unpaar, 3 paarige Luftsäcke |
| 7 | Truthuhn | Halsluftsack unpaar, hintere Brustluftsäcke fehlen |
| Singvögel | unpaarer Schlüsselbeinluftsack mit vorderen Brustluftsäcken verschmolzen | |
Luftsacksysteme
Man unterscheidet funktionell zwei Luftsacksysteme:
- Das kraniale Luftsacksystem besteht aus den Hals-, Schlüsselbein- und den beiden vorderen Brustluftsäcken. Sie werden über die so genannten medioventralen Sekundärbronchien (zur Mitte und nach unten gerichtete Abgänge der Hauptbronchien) belüftet und sind meist nur bei verstärkter Atmung aktiv.
- Das kaudale Luftsacksystem besteht aus den hinteren Brust- sowie den Bauchluftsäcken. Sie sind über einen lateroventralen Sekundärbronchus bzw. direkt an den Hauptbronchus angeschlossen.
Atemmechanik bei Vögeln
Prinzip
Die Atmung der Vögel unterscheidet sich grundsätzlich von der der Säugetiere. Bei Säugetieren ändern die Muskeln für Inspiration (Einatmung) und Exspiration (Ausatmung) das Volumen des Brustkorbs und mit diesem das Volumen der Lungen, wodurch Luft in die Lungen ein- und ausströmt. Auch bei Reptilien ändern Muskeln das Volumen des Brustkorbs – und sofern vorhanden, das ihrer Luftsäcke – zur Atmung, während die Lunge von Vögeln keine Volumenänderungen ausführen kann.
Die wichtigsten Inspirationsmuskeln sind die Rippenanhangsmuskeln (Musculi appendicocostales). Ihre Kontraktion führt zur Erweiterung des Brustkorbs. Auch die Abwärtsbewegung des Brustbeins trägt maßgeblich zu diesem Prozess bei. Dies führt zu einem Unterdruck in der Leibeshöhle und damit zu einer Erweiterung der Luftsäcke, wodurch Luft durch die Lunge hindurch angesaugt wird. Die wichtigsten Exspirationsmuskeln sind die Bauchmuskeln, die die Leibeshöhle verengen und damit die Luft aus den Luftsäcken verdrängen. Sowohl bei der Ein- als auch bei der Ausatmung strömt die Luft von hinten nach vorn durch die Parabronchien der Lunge.
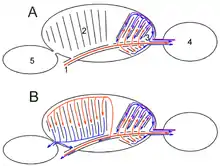
Rot: Frischluft, blau: sauerstoffarme Luft, violett: Mischluft
1 Trachea, 2 Palaeopulmo, 3 Neopulmo,
4 hinteres Luftsacksystem,
5 vorderes Luftsacksystem
Ruheatmung
In der Ruheatmung (Abb. A) ist zumeist nur das hintere Luftsacksystem aktiv. Von den beiden hinteren Luftsackpaaren wird Luft durch einen Teil der Lunge (so genannte Neopulmo) gesaugt, zum Teil gelangt auch Frischluft in diese Luftsäcke. Damit befindet sich am Ende der Einatmung Mischluft mit einem noch nutzbaren Sauerstoffanteil in den hinteren Luftsäcken. Bei der Ausatmung wird diese Mischluft nun noch einmal durch die Lunge geleitet und somit viel effektiver ausgenutzt als beim Säugetier mit seinen blind endenden Luftwegen und dem dementsprechend hohen Totraum.
Dieses effiziente System kompensiert auch das große Volumen der Luftröhre bei Vögeln, die im Vergleich zu Säugetieren länger und dicker ist und ein etwa 4,5faches Totraumvolumen hat.
Forcierte Atmung
Bei vermehrter Atmung (Abb. B) kommt zu der normalen Atmung noch das vordere Luftsacksystem dazu. Es saugt Frischluft durch den übrigen Teil der Lunge (so genannte Palaeopulmo). Die verbrauchte Luft dieses Luftsacksystems gelangt bei der Ausatmung direkt in die Luftröhre, wird also nicht noch einmal durch die Lunge geleitet.
Tierartliche Besonderheiten
Bei Vogelarten ohne Neopulmo (z. B. Pinguine) gelangt nur Frischluft in die hinteren Luftsäcke, sie nutzen bei der Einatmung das vordere, bei der Ausatmung das hintere System. Bei Störchen ist die Neopulmo schwach entwickelt.
Konsequenzen aus der Atemmechanik
Beim Festhalten und bei der Manipulation von Vögeln ist die komplexe Atemmechanik zu beachten. Beim In-die-Hand-nehmen ist darauf zu achten, dass der Brustkorb nicht zu stark in seiner Bewegung eingeschränkt wird. Legt man Vögel auf den Rücken, so behindert die Last der übrigen Organe die Entfaltung der Bauchluftsäcke und die Schwerkraft des Brustmuskels die des Brustbeins, so dass es schnell zu Atemnot oder gar zu einer Erstickung kommen kann.
Evolutionärer Ursprung des Luftsacksystems
.jpg.webp)
Bei zahlreichen Vertretern der Theropoden, der Klade der Dinosaurier, aus denen die modernen Vögel hervorgingen, sind Wirbel und Rippen mit pneumatischen Foramina vergleichbar denen rezenter Vögel nachgewiesen, ebenso beim Urvogel Archaeopteryx. Sie weisen darauf hin, dass bereits bei den Vorfahren der Vögel bestimmte Knochen mit Aussackungen (Divertikeln) eines Luftsacksystems gefüllt (das heißt pneumatisiert) waren. Innerhalb der Archosaurier, deren rezente Vertreter die Krokodile und die Vögel sind, besaßen neben den Theropoden, die als Stammgruppe der Vögel gelten, weitere Gruppen (die Flugsaurier und die Sauropoden) pneumatisierte Knochen, so dass von manchen Paläontologen angenommen wird, ein Luftsacksystem sei nicht nur ein urtümliches Merkmal der Vögel, sondern das einer weitaus größeren und älteren Gruppe, der so genannten Ornithodiren, gewesen.
Inwiefern die Funktion des Atmungssystems bei rezenten Vögeln und Nicht-Vogel-Theropoden ähnlich war, wird kontrovers diskutiert: O’Connor und Claessens (2005) schlagen für Nicht-Vogel-Theropoden eine den Vögeln ähnliche Form der Durchströmungsatmung vor. Sie wiesen in einem gut erhaltenen fossilen Exemplar der urtümlichen Theropoden-Gattung Majungatholus pneumatische Öffnungen in Hals-, Brust- und Beckenwirbeln nach.
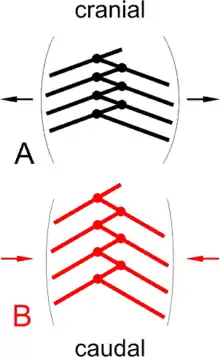
Dass die Neuralbögen der Brustwirbel 12 und 13 verglichen mit den davor und dahinter liegenden besonders kleine Öffnungen zeigen, interpretieren sie als Trennung zwischen einem vorderen (kranialen) und einem hinteren (kaudalen) Luftsacksystem, denen jeweils die Divertikel in den vorderen bzw. hinteren Wirbelkörpern und Neuralbögen zuzuordnen sind. Das Vorhandensein eines hinteren Luftsacksystems, das mechanisch einfacher zu „belüften“ sei als ein vorderes, halten O’Connor und Claessens für eine entscheidende Voraussetzung für eine vogelähnliche Durchströmung der Lunge (siehe Ruheatmung bei rezenten Vögeln, Abb. A).
Theropoden einschließlich früher Vögel besaßen im Gegensatz zu rezenten flugfähigen Vögeln nur ein kleines, kaum den vorderen Brustbereich einnehmendes Brustbein und ein dreistrahliges Becken – das Schambein war bauchwärts gerichtet und nicht wie bei späteren Vögeln parallel zu Darmbein und Sitzbein rotiert. Offenbar spielten Auf- und Abwärtsbewegungen des Brustbeins sowie Rotationen des Beckens („Beckenatmung“) beim Ein- und Ausatmen zu Beginn der Vogelevolution noch keine Rolle. Andererseits waren Theropoden einschließlich urtümlicher mesozoische Vögel wie Archaeopteryx, Confuciusornis und Sinornis im Besitz miteinander verzahnter Bauchrippen (Gastralia). Durch die Kontraktion der Bauchmuskulatur (besonders des Musculus rectus abdominis) wurden laut Claessens (2004) die Bauchrippen gegeneinander geschoben und so der hintere Brustraum seitlich erweitert. Somit ermöglichte die Kontraktion derselben Muskeln, die bei rezenten Vögeln zur Verringerung des Brustraumvolumens beim Ausatmen beitragen, bei frühen Vögeln und Nicht-Vogel-Theropoden paradoxerweise das Einatmen. Dieser Atemmechanismus, der auf Verschiebung eines Systems von Bauchrippen beruht, wird nach Carrier und Farmer (2000) auch als „Kürassatmung“ (cuirassal breathing) bezeichnet und als eine notwendige Voraussetzung für eine urtümliche Durchströmungsatmung angesehen. Bei späteren Vögeln wurde mit dem Verlust der Bauchrippen diese Form der Brustraumkontraktion und -expansion durch andere Formen abgelöst.
In welchem phylogenetischen Stadium zuerst ein Atemmechanismus ähnlich dem der heutigen Vögel vorlag, ist noch weitgehend offen. Bevor es der Atmung diente, könnte das Luftsacksystem bei Dinosauriern und Flugsauriern thermoregulatorische Funktion gehabt und zur Gewichtsreduktion beigetragen haben. Ruben u. a. (1997, 1999) glauben, bei Fossilien der kleinen theropoden Dinosaurier Sinosauropteryx und Scipionyx versteinertes Weichgewebe des Brust- und Bauchraums vorliegen zu haben, das zeige, dass die Atmung der Theropoden der der Vögel nicht ähnlich sei: Angeblich sei der Brustraum wie bei Krokodilen durch ein Zwerchfell unterteilt gewesen, dessen kolbenartige Bewegung das Ein- und Ausatmen steuerte. Diese Theorie lässt jedoch den Nachweis pneumatischer Foramina in einer großen Zahl von Theropoden unbeachtet. Und dass die Versteinerungen bzw. Hautschatten im Bereich des Bauchraums der genannten Exemplare tatsächlich oben genannte Aussagen zur Lage und Anatomie der inneren Organe zulassen, gilt unter Paläontologen als zweifelhaft.
Klinische Bedeutung
Das Luftsacksystem ist häufig bei Erkrankungen der Atemwege mit betroffen. Der Bau der Luftsäcke führt dazu, dass Vögel, anders als Säugetiere, Fremdkörper nicht abhusten können. Erkrankungen des Luftsacks gehen im Allgemeinen mit schwerer Atemnot einher.
- Die häufigsten bakteriellen Erreger einer Luftsackentzündung (Aerosacculitis) sind Escherichia coli, Mykoplasmen (Mycoplasma gallisepticum u. a.) und Chlamydien (Ornithose, Chlamydophila psittaci), seltener Pasteurellen (Pasteurella gallinarum), Ornithobacterium rhinotracheale und Bordetellen (Bordetella avium). Zum Nachweis von bakteriellen Erregern kann eine Spülung analog der bronchoalveolären Lavage durchgeführt werden.
- An durch Schimmelpilze hervorgerufenen Luftsackmykosen ist vor allem Aspergillus fumigatus beteiligt. Die Aspergillose ist die häufigste Luftsackinfektion.
- Auch bei Viruserkrankungen wie Geflügelpest („Vogelgrippe“), Newcastle-Krankheit (atypische Geflügelpest), Infektiöse Bronchitis (IB) und Infektiöser Laryngotracheitis (ILT) kann eine Aerosacculitis auftreten.
- Schließlich können auch Parasiten wie Würmer (Serratospiculum spp.) oder Milben (Luftsackmilbe Cytodites nudus) den Luftsack befallen.
Die Luftsäcke sind limitierender Faktor für die Ultraschalluntersuchung bei Vögeln, da die Grenzfläche Gewebe/Luft eine Totalreflexion der Schallwellen verursacht. Eine direkte Applikation von Isofluran in die Luftsäcke ist als Narkosemöglichkeit beschrieben.
Literatur
- B. B. Britt, P. J. Makovicky, J. Gauthier, N. Bonde: Postcranial pneumatization in Archaeopteryx. In: Nature. 395/1998, S. 374–376, ISSN 0028-0836.
- D. R. Carrier, C. G. Farmer: The evolution of pelic aspiration in archosaurs. In: Paleobiology. 26(2)/2000, S. 271–293, ISSN 0094-8373.
- L. P. A. M. Claessens: Dinosaur gastralia: origin, morphology, and function. In: Journal of Vertebrate Paleontology. 24(1)/2004, S. 89–106.
- J. R. Codd, D. F. Boggs, S. F. Perry, D. R. Carrier: Activity of three muscles associated with the uncinate processes of the giant Canada Goose Branta canadensis maximus. In: Journal of Experimental Biology. 208/2005, S. 849–857. (Volltext)
- H.-R. Duncker: The lung air sac system of birds. In: Ergebnisse der Anatomie und Entwicklungsgeschichte. 45/1971, S. 1–171, ISSN 0301-5556.
- P. M. O’Connor, L. P. A. M. Claessens: Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs. In: Nature. 436/2005, S. 253–256, ISSN 0028-0836.
- J. A. Ruben, T. D. Jones, N. R. Geist, W. J. Hillenius: Lung Structure and Ventilation in Theropod Dinosaurs and Early Birds. In: Science. 278/1997, S. 1267–1270.
- J. A. Ruben, C. D. Sasso, N. R. Geist, W. J. Hillenius, T. D. Jones, M. Signore: Pulmonary Function ans Metabolic Physiology of Theropod Dinosaurs. In: Science. 283/1999, S. 514–516.
- F.-V. Salomon (Hrsg.): Lehrbuch der Geflügelanatomie. Fischer-Verlag, Jena/Stuttgart 1993, ISBN 3-334-60403-9.
- J. H. Smith, J. L. Meier, C. Lamke, P. J. Neill, E. D. Box: Microscopic and submicroscopic anatomy of the parabronchi, air sacs, and respiratory space of the budgerigar (Melopsittacus undulatus). In: American Journal of Anatomy. 177/1986, S. 221–242, ISSN 0002-9106.
- M. J. Wedel: Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs. In: Paleobiology. 29(2)/2003, S. 243–255, ISSN 0094-8373.
Weblinks
- birdrespiration (englisch)