ABCC11
Das Protein ABCC11 (ATP-binding cassette sub-family C member 11, dt.: ‚ATP-bindende Kassette, Unterfamilie C (CFTR/MRP), Mitglied 11‘), auch MRP8 (Multidrug Resistance-Related Protein 8, dt.: ‚Resistenz gegen mehrere Medikamente, Protein 8‘) genannt, ist ein Membrantransporter, der bestimmte Moleküle aus dem Inneren einer Zelle transportiert. ABCC11 findet sich vor allem in apokrinen Drüsen, also Duftdrüsen, die zum Beispiel in den Haarfollikeln enden. Das korrespondierende ABCC11-Gen wurde bisher nur bei einigen Säugetierarten – einschließlich des Menschen – gefunden. Beim Menschen liegt es auf Chromosom 16. Eine Vielzahl von Menschen, speziell im nordostasiatischen Raum, verfügt über kein funktionsfähiges ABCC11. Die Ursache hierfür ist eine Punktmutation im ABCC11-Gen. Die beiden Genotypen führen zu sichtbar unterschiedlichen Merkmalen (phänotypische Variation). So haben Menschen, bei denen beide ABCC11-Gene auf dem entsprechenden Chromosomenpaar die Mutation aufweisen, beispielsweise weißes, trockenes Ohrenschmalz und einen nur schwachen Körpergeruch. Der Wildtyp mit zwei „normalen“ ABCC11-Genen und der heterozygote Typ mit einem defekten ABCC11-Gen haben dagegen gelbliches, feuchtes Ohrenschmalz und einen deutlicher ausgeprägten Körpergeruch. Der Gendefekt wird rezessiv vererbt. Die regionale Häufigkeit der Punktmutation liefert wichtige Indizien zur Ausbreitung des Menschen über die Erde und die Durchmischung verschiedener Populationen.
| ATP-bindende Kassette, Unterfamilie C (CFTR/MRP), Mitglied 11 | ||
|---|---|---|
| Eigenschaften des menschlichen Proteins | ||
| Masse/Länge Primärstruktur | 1382 Aminosäuren (Isoform: 1344 AS) | |
| Kofaktor | ATP | |
| Bezeichner | ||
| Gen-Namen | ABCC11 MRP8 | |
| Externe IDs | ||
| Enzymklassifikation | ||
| Substrat | lipophile Anionen | |
| Orthologe (Mensch) | ||
| Entrez | 85320 | |
| Ensembl | ENSG00000121270 | |
| UniProt | Q96J66 | |
| Refseq (mRNA) | NM_032583 | |
| Refseq (Protein) | NP_115972 | |
| Genlocus | Chr 16: 48.17 – 48.25 Mb | |
| PubMed-Suche | 85320 | |
Genetik
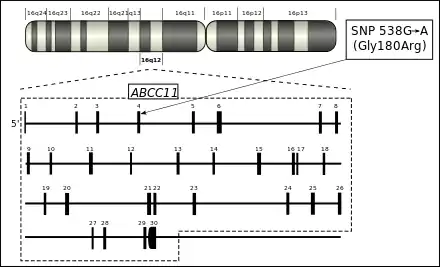
Das ABCC11-Gen befindet sich beim Menschen auf Chromosom 16, Genlocus q12.1. Die genomischen Koordinaten (nach GRCh37[# 1]) sind 16:48.200.781–48.281.478.[2] Das Gen besteht somit aus 80.697 Basenpaaren (bp). Diese untergliedern sich in 30 Exons.[1][3]
Im Jahr 2001 wurde ABCC11 von drei Arbeitsgruppen[4][5][3] unabhängig voneinander erstmals aus der cDNA-Bibliothek von einer humanen Leber isoliert.[1] In unmittelbarer Nähe, auf demselben Genlocus und nur etwa 20 kbp voneinander getrennt, liegt – in Schwanz-an-Kopf-Orientierung zu ABCC11 – das ABCC12-Gen. Beide Gene weisen untereinander und zu ABCC3 und ABCC5 eine hohe Ähnlichkeit auf. Auch sie besitzen zwei ATP-bindende Kassetten und zwölf transmembrane Helices.[1]
Das ABCC11-Gen wurde bisher nur bei Säugetieren gefunden. Durch Klonierung konnte es bisher bei folgenden Arten nachgewiesen werden (Stand: September 2014): Mensch (Homo sapiens),[6] Gemeiner Schimpanse (Pan troglodytes),[7] Westlicher Flachlandgorilla (Gorilla gorilla gorilla),[8] Sumatra-Orang-Utan (Pongo abelii),[9] Nördlicher Weißwangen-Schopfgibbon (Nomascus leucogenys),[10] Rhesusaffe (Macaca mulatta),[11] Weißbüschelaffe (Callithrix jacchus),[12] Philippinen-Koboldmaki (Tarsius syrichta), Kleinohr-Riesengalago (Otolemur garnettii),[13] Wildkaninchen (Oryctolagus cuniculus), Nördliches Spitzhörnchen (Tupaia belangeri), Afrikanischer Elefant (Loxodonta africana),[14] Kleiner Igeltenrek (Echinops telfairi), Haushund (Canis familiaris),[15] Frettchen (Mustela putorius furo),[16] Großer Panda (Ailuropoda melanoleuca),[17] Hauskatze (Felis catus),[18] Großer Tümmler (Tursiops truncatus), Hausrind (Bos taurus),[19] Hausschaf (Ovis aries),[20] Alpaka (Vicugna pacos), Braunbrustigel (Erinaceus europaeus), Waldspitzmaus (Sorex araneus), Hauspferd (Equus caballus),[21] Beutelteufel (Sarcophilus harrisii),[22] Schnabeltier (Ornithorhynchus anatinus) und Derbywallaby (Macropus eugenii).[23]
Bei den Modellorganismen Ratte und Maus gibt es kein dem humanen ABCC11 entsprechendes Gen (orthologes Gen).[24] ABCC11 ist ein paraloges Gen, das im Laufe der Evolution durch Genverdopplung aus ABCC12 entstanden ist. Zum humanen ABCC12 gibt es bei Nagetieren und vielen anderen Tieren die entsprechenden orthologen Gene.[24][25][1]
Mittels Transkriptionsanalyse wurde festgestellt, dass die mRNA von ABCC11 in allen adulten und fetalen Geweben exprimiert wird.[4][3] Besonders große Mengen an ABCC11-mRNA finden sich in Brustkrebs-Gewebe.[5][3][1]
Proteomik
Das ABCC11-Gen kodiert für ein aus 1382 Aminosäuren bestehendes Protein, dessen Sequenz zu etwa 47 % identisch mit MRP9, dem Genprodukt von ABCC12, ist. Eine Spleißvariante von ABCC11 (Variante A) führt zu einem Genprodukt mit 1344 Aminosäuren. Ursache hierfür ist der vollständige Ausfall von Exon 28. Variante A hat ebenfalls zwölf transmembrane Helices, allerdings fehlen an der zweiten ATP-bindenden Kassette 38 Aminosäuren in den Positionen 1261 bis 1298. Diese Variante wird zu etwa 25 % von den Zellen exprimiert.[26]
Funktion
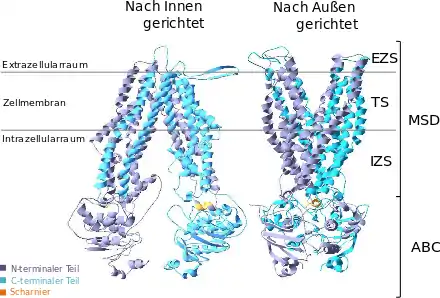
Links die nach innen gerichtete Konformation zur Aufnahme eines Substrates aus dem Intrazellularraum. Rechts die in den Extrazellularraum gerichtet Konformation zur Abgabe des Substrates. EZS = extrazelluläre Schleife, TS = Transmembran-Segment, MSD = membranspannende Domäne, IZS = intrazelluläre Schleife, ABC = ATP-bindende Kassette. Das Scharnier bildet den Drehpunkt, der dem Protein seine Beweglichkeit zur Konformationsänderung gibt. Für die Kristallstruktur des ABCC11-Proteins gibt es noch keine Daten, da eine Kristallisation bisher (Stand September 2014) noch nicht gelang.[29]
ABCC11 ist ein Membrantransporter (veraltet: Effluxpumpe). Dies sind membranständige Transportproteine, die bestimmte, für den Transporter spezifische Moleküle (Substrate) aus der Zelle oder einem Zellkompartiment hinaus befördern. ABCC11 gehört zur Familie der ABC-Transporter (ABC = ATP binding cassette). Diese Membranproteine haben als gemeinsames Strukturelement mindestens eine ATP-bindende Kassette (ATP = Adenosintriphosphat). Ist die Konzentration eines Substrates außerhalb der Zelle höher als innerhalb, so muss für den Transportvorgang gegen diesen Konzentrationsgradienten Energie aufgewendet werden. Im Fall der ABC-Transporter geschieht dies durch Bindung und Hydrolyse des zellulären Energieträgers ATP, in der ATP-bindenden Kassette.
Das menschliche Genom umfasst 48 ABC-Transporter,[30] die in drei Unterfamilien (ABCA, ABCB und ABCC) unterteilt sind. Die ABCC-Unterfamilie hat zehn Mitglieder, die Multidrug Resistance-Related Proteine (MRP). Ein wichtiger ABCC-Transporter ist beispielsweise Cystic Fibrosis Transmembrane Conductance Regulator (CFTR), das Schlüsselprotein bei der Erbkrankheit Mukoviszidose. Substrate für ABCC11 sind lipophile Anionen. Dazu gehören unter anderem cyclische Nukleotid-Monophosphate (cNMP) wie cyclisches Adenosinmonophosphat (cAMP) und cyclisches Guanosinmonophosphat (cGMP),[31] Glutathion-Konjugate wie beispielsweise Leukotrien C4 und 2,4-Dinitrophenyl-S-Glutathion (DNP-SG), Steroid-Sulfate wie beispielsweise Estron-3-Sulfat (E13S) und Dehydroepiandrosteron-3-Sulfat (DHEAS), Glucuronide wie beispielsweise Estradiol-17-β-D-Glucuronid, die Gallensäuren Glyco- und Taurocholsäure sowie Folsäure.[32][33][1] Die verwandten Transporter ABCC4 und ABCC5 weisen eine hohe strukturelle Ähnlichkeit zu ABCC11 auf. Folglich haben alle drei sehr ähnliche Zielsubstrate.[1]
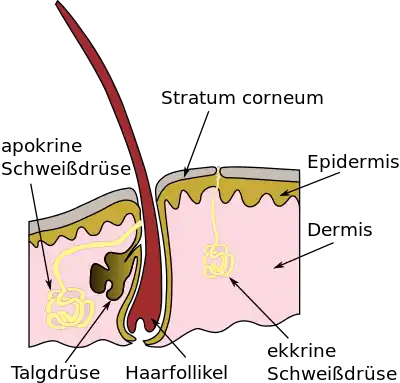
Das ABCC11-Protein wird vor allem in den Epithelzellen der apokrinen Schweißdrüsen (Glandulae sudoriferae apocrinae) exprimiert.[34] Die apokrinen Schweißdrüsen, die auch als ‚Duftdrüsen‘ bezeichnet werden, befinden sich beim Menschen in Bereichen behaarter Haut, vor allem in den Achselhöhlen, im Genital- und Analbereich, den Brustwarzen und dem Warzenhof sowie dem Nasenvorhof (Vestibulum nasi). Eine Duftdrüse ist ein röhrenartiges (tubuläres) Knäuel. Im Inneren des Tubulus befinden sich in einer einschichtigen Lage Epithelzellen. Von diesen Zellen ragen Cytoplasmakuppen in den Tubulus hinein. Diese Kuppen werden bei der Sekretion abgeschnürt und in das Innere des Tubulus (das Lumen) abgegeben. Die Epithelzellen verlieren dabei einen Teil ihres Cytoplasmas und ihrer Zellmembran. Diese apokrine Sekretion wird auch Apozytose genannt. Modifizierte apokrine Schweißdrüsen befinden sich am Lidrand der Wimpern, die Moll-Drüsen (Glandulae ciliares conjunctivales), und in den äußeren Gehörgängen, die Ohrenschmalzdrüsen (Glandulae ceruminosae).[35]
Außer im Epithel der apokrinen Schweißdrüsen wird ABCC11 noch auf Leberzellen, weißen Blutkörperchen und Blasten im Knochenmark exprimiert.[36][30]
Die Expression von ABCC11 wird über den Estrogenrezeptor-α reguliert. Durch Estradiol wird die Expression reduziert.[37] Entsprechend wird die Expression durch Tamoxifen, ein Estradiol-Antagonist, der über kompetitive Hemmung an den Estrogenrezeptor-α bindet, hochreguliert.[38]
Multidrug Resistance
Krebszellen haben eine hohe Mutations- und Zellteilungsrate. Sie werden ständig auf Überleben und Proliferation selektiert. Während dieses evolutionären Prozesses nutzen die entarteten Zellen grundlegende physiologische Mechanismen, um sich vor der toxischen Wirkung der Chemotherapie zu schützen. Einer dieser Mechanismen ist der Transport (Efflux) von Chemotherapeutika aus der Zelle. In einer Vielzahl von Studien wurde gezeigt, dass der Efflux über die Behandlungsdauer, also über die Anzahl an Therapiezyklen, mit einem Chemotherapeutikum zunimmt. Die Krebszellen exprimieren mehr und mehr Transporter, um die für die Zelle toxischen Substanzen wieder „herauspumpen“ zu können.[39] Für den behandelten Patienten bedeutet dies, dass sein Tumor, oder dessen Metastasen, nicht mehr auf den Wirkstoff ansprechen, weil sich eine Wirkstoffresistenz ausgebildet hat. In der Regel wird dann – wenn vorhanden – ein strukturell anderes Chemotherapeutikum verwendet, das idealerweise für die überexprimierten Transporter kein geeignetes Substrat ist. Grundsätzlich ist die Resistenzbildung bei Krebszellen, aber auch bei mit Viren infizierten Zellen, für den Therapieerfolg ausgesprochen ungünstig.
Verantwortlich für den Wirkstofftransport aus der Zelle heraus sind die ABC-Transporter, zu denen auch ABCC11 gehört. Daher auch der Name Multidrug Resistance-Related Protein 8 für ABCC11. Zu diesen Arzneimitteln, die ABCC11 aus der Zelle heraustransportieren können, gehören unter anderem Nukleosid- und Folsäure-Analoga. Nukleosidanaloga sind beispielsweise das wichtige Chemotherapeutikum 5-Fluoruracil (5-FU) und seine Abkömmlinge Floxuridin und Doxifluridin, sowie Cytarabin,[29] das HIV-Therapeutikum Zalcitabin und der Hepatitis-B-Wirkstoff Adefovir. Zellen, die ABCC11 überexprimieren, können im Vergleich zu normalen Zellen eine etwa dreifach höhere Dosis an 5-FU und Doxifluridin überleben, im Fall von Floxuridin und Adefovir die 5-fache, und bei Zalcitabin gar die 6-fache Dosis.[31] Das Folsäure-Analogon Methotrexat (MTX) ist ebenfalls ein Substrat für ABCC11.[1] Die Verabreichung von 5-Fluoruracil beeinflusst direkt die Expression von ABCC11.[40]
Genetischer Polymorphismus
Bisher (Stand 2013) sind über zehn nicht-synonyme Einzelnukleotid-Polymorphismen (engl. single nucleotide polymorphism, SNP) beim humanen ABCC11-Gen bekannt. Nicht-synonyme SNPs sind Variationen einzelner Basenpaare in einem Gen, die dazu führen, dass im Genprodukt – dem Protein – eine einzelne Aminosäure ausgetauscht wird. Nicht-synonyme SNPs sind eine spezielle Form einer Punktmutation, die gehäuft bei mindestens 1 % der jeweiligen Population auftritt und sich im Genpool dieser Population erfolgreich durchgesetzt hat. Als Polymorphismus bezeichnet man das Auftreten mehrerer Genvarianten innerhalb einer Population.
| Wildtyp (Basensequenz des Sense-Stranges) |
▾530 ▾538 ▾550 ATT GCC AGT GTA CTC GGG CCA ATA TTG ATT ATA CCA |
|---|---|
| Wildtyp (Aminosäurensequenz) |
175 176 177 178 179 180 181 182 183 184 185 186 Ile Ala Ser Val Leu Gly Pro Ile Leu Ile Ile Pro |
| 538G→A / Gly180Arg (Basensequenz des Sense-Stranges) |
▾530 ▾538 ▾550 ATT GCC AGT GTA CTC AGG CCA ATA TTG ATT ATA CCA |
| 538G→A / Gly180Arg (Aminosäuresequenz) |
175 176 177 178 179 180 181 182 183 184 185 186 Ile Ala Ser Val Leu Arg Pro Ile Leu Ile Ile Pro |
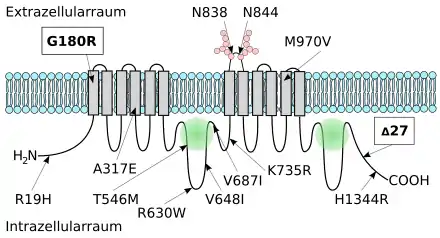
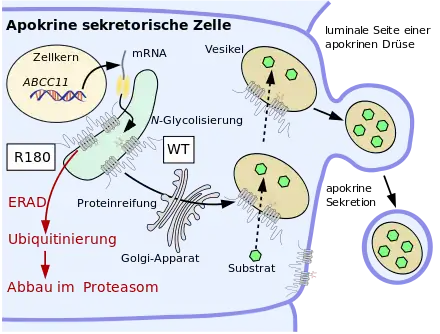
Eine besondere Bedeutung hat der SNP rs17822931.[41][42] Bei dieser Mutation ist in Position 538 die Nukleinbase Guanin (G) durch Adenin ersetzt (538G→A). Aus dem ursprünglichen Basentriplett GGG wird dadurch AGG. Während GGG für die Aminosäure Glycin kodiert, kodiert AGG für Arginin. Deshalb wird in Position 180 bei der Proteinbiosynthese im ABCC11-Protein nach der Translation statt Glycin Arginin eingebaut (Gly180Arg). Dieser Aminosäurenaustausch bewirkt, dass eine korrekte posttranslationale N-Glykolisierung von Asparagin in Position 838 und 844 des Proteins im endoplasmatischen Retikulum nicht möglich ist. Dies hat wiederum zur Folge, dass das Protein ‚falsch‘ faltet. Diese Fehlfaltung wird von der Proteinqualitätskontrolle erkannt, das fehlerhafte ABCC11 ubiquitinyliert, das heißt als fehlerhaft markiert, und anschließend im Proteasom abgebaut.[43] Der Einzelnukleotid-Polymorphism 538G→A führt folglich im betroffenen Gen zu einem Funktionsverlust. Das durch die Proteinqualitätskontrolle ausgesonderte Protein wird nicht an der Zellmembran exprimiert.[# 2] Man spricht in solchen Fällen von einer Loss-of-Function-Mutation (Funktionsverlustmutation). Wie die meisten Loss-of-Function-Mutationen wird auch die Mutation 538G→A rezessiv vererbt.
Nachfolgend werden die bisher bekannten Phänotypen des Einzelnukleotid-Polymorphism 538G→A gezeigt. Die anderen nicht-synonymen Einzelnukleotid-Polymorphismen sind wesentlich seltener und vergleichsweise bedeutungslos. Neben den Einzelnukleotid-Polymorphismen ist noch eine Deletion von 27 Basenpaaren in Exon 29 in der Literatur beschrieben. Diese mit Δ27 beziehungsweise 3939–3965del27 bezeichnete Mutation ist vergleichsweise selten. Bei einem 2007 durchgeführten Screening von 722 Probanden in sieben japanischen und je einer koreanischen und deutschen Population, hatten lediglich zwei Probanden, einer aus der Präfektur Yamagata und einer aus Korea, diese Mutation. Ein Proband, bei dem die Mutation gefunden wurde, hat trotz heterozygotem Genotyp (G/A-Typ) weißes Ohrenschmalz. Die Mutation befindet sich daher auf dem „normalen“ G-Allel und bewirkt dort einen Funktionsverlust aus dem daraus kodierten ABCC11-Protein, wie dies auch beim A-Allel des Probanden der Fall ist.[28][44]
Ohrenschmalz-Phänotyp

Es gibt beim Menschen zwei Arten von Ohrenschmalz (Cerumen): eine trockene, helle Form mit einem hohen Anteil gesättigter Fettsäuren und eine feuchte, gelb-braune Form, mit einem hohen Anteil ungesättigter Fettsäuren. Die Art des Cerumen ist genetisch bedingt. Dabei ist die feuchte Form gegenüber der trockenen Form dominant, beziehungsweise ist die trockene Form gegenüber der feuchten Form rezessiv. Während die trockene Variante in der Bevölkerung Nordamerikas, Europas und Afrikas mit unter 3 Prozent sehr selten ist, liegt ihr Anteil in der Bevölkerung Ostasiens bei über 80 %. Die Ursache für den Dimorphismus beim menschlichen Ohrenschmalz ist die Einzelnukleotidmutation 538G→A im ABCC11-Gen. Sie entscheidet über Aussehen und Zusammensetzung des Cerumen. Dem dominanten Erbgang des Merkmals entsprechend haben der reinerbige (homozygote) G/G- und der mischerbige (heterozygote) G/A-Typ die feuchte Form von Ohrenschmalz. Die trockene Form weist nur der homozygote A/A-Typ auf, bei dem beide Allele die mutierte Form des ABCC11-Gens aufweisen.[43] Beim heterozygoten Typ wird der Ausfall eines Allels durch das andere weitgehend kompensiert, da die Gendosis des intakten Allels ausreicht, um den Funktionsverlust zu kompensieren. Mit dem Einzelnukleotid-Polymorphismus 538G→A in ABCC11 wurde 2005[44] erstmals ein DNA-Polymorphismus entdeckt, der ein sichtbares genetisches Merkmal – Farbe und Konsistenz von Ohrenschmalz – zur Folge hat.[43] Die antibakteriellen Eigenschaften der beiden Cerumentypen sind weitgehend gleich.[45]
Körpergeruch
Der Mensch hat zwei Arten von Schweißdrüsen: ekkrine und apokrine. Während die über den ganzen Körper verteilten ekkrinen Schweißdrüsen in die Hautoberfläche münden, enden die apokrinen Drüsen – auch Duftdrüsen genannt – in den Haarfollikeln. Folglich finden sich die apokrinen Schweißdrüsen nur in bestimmten behaarten Hautgebieten wie den Achselhöhlen, den Brustwarzen, sowie im Genital- und Perianalbereich. Der von den zwei Schweißdrüsenarten produzierte Schweiß unterscheidet sich in seiner Zusammensetzung deutlich, ist aber in beiden Fällen weitgehend geruchslos. Allerdings können die Ausscheidungen der apokrinen Schweißdrüsen durch Bakterien, vor allem der Gattungen Corynebacterium und Staphylococcus, in Riechstoffe umgewandelt werden.[46][43] Dies gibt jedem Menschen einen einzigartigen und charakteristischen Körpergeruch, der unterschiedliche, hochkomplexe Funktionen erfüllt und so beispielsweise Physiologie und Verhaltensweise beeinflusst.[47][48] Die apokrinen Schweißdrüsen sind zwar bereits bei der Geburt angelegt, nehmen aber erst mit dem Beginn der Pubertät ihre sekretorische Aktivität auf.[49][27] Auch aus diesem Grund nimmt man an, dass die apokrinen Schweißdrüsen einen wichtigen Beitrag zum Sozial- und Sexualverhalten des Menschen leisten.[50][51] Dies wird auch bei der nonverbalen Kommunikation, beispielsweise durch Angstschweiß, angenommen.[52][53][54][55] Im Gegensatz zum farblosen, wässrigen ekkrinen Schweiß ist der apokrine Schweiß von milchiger, viskoser Beschaffenheit. Der apokrine Schweiß enthält unter anderem Dehydroepiandrosteron-3-sulfat (DHEAS),[56][34] Androsteronsulfat,[57] Cysteinyl-Glycyl-3-Methylhexanol[58][59] und kurzkettige, verzweigte Fettsäuren, die mit Glutamin konjugiert sind.[60][27] Diese Verbindungen sind nicht flüchtig und folglich geruchsfrei. Sie sind aber Vorläufermoleküle (Präkursoren), die durch die Bakterien der Hautflora in flüchtige Verbindungen, im Beispiel Dehydroepiandrosteron, Androsteron (beide mit Moschus- bis Urin-artigem Geruch[61][62]), 3-Methyl-3-sulfanylhexanol (zwiebelartiger Geruch[27]) und die freien kurzkettigen, verzweigten Fettsäuren (säuerlich-ranziger Geruch) zerlegt werden. Sie sind – zusammen mit weiteren Stoffwechselprodukten der Bakterien – für den als schlecht empfundenen Geruch des Schweißes verantwortlich.[63] Allgemein wird in der entwickelten Welt Körpergeruch negativ bewertet. Außer den individuellen Unterschieden gibt es auch erhebliche ethnische Unterschiede beim Körpergeruch. In vielen asiatischen Ethnien ist der charakteristische und intensive Körpergeruch von Menschen mit europäischer oder afrikanischer Herkunft kaum vertreten. Dort überwiegt ein schwach saurer Körpergeruch.[64][65][34] Beispielsweise empfinden Japaner das europäisch-afrikanische Geruchsmuster als unangenehm, weshalb sie früher – ab etwa dem 19. Jahrhundert – Europäer und Amerikaner als bata-kusai (バタ臭い, dt.: ‚Butterstinker‘) bezeichneten.[66] Sie vermuteten den in Japan zu dieser Zeit unüblichen Konsum von Butter durch die Gaijins (‚Mensch von draußen‘) als Ursache für den „Gestank“.[67] Durch die verbesserte Körperhygiene und die Verbreitung von Deodorants hat der Begriff bata-kusai in Japan mittlerweile eine Wandlung erfahren. Er steht heute abwertend für Dinge, die unangenehm oder anstößig westlich sind.[68]
Bereits 1937 wurde ein Zusammenhang zwischen dem Phänotyp des Ohrenschmalzes (trocken oder feucht) und dem Körpergeruch (schwach oder intensiv) bei unterschiedlichen Ethnien beschrieben.[65] Tatsächlich wurde 2009 mittels Gen-Analyse festgestellt, dass es sich um denselben Genotyp handelt. Ursache ist in beiden Fällen der Einzelnukleotid-Polymorphismus 538G→A im ABCC11-Gen. Der Ausfall des ABCC11-Transporters ist die Ursache dafür, dass die für den Körpergeruch verantwortlichen Präkursoren nicht mehr aus dem Cytoplasma der Epithelzellen der apokrinen Schweißdrüsen in die Vesikel für die apokrine Sekretion transportiert werden können und die Bakterien der Hautflora nicht die Duftstoffe aus den Präkursoren bilden können. Experimentell zeigt sich dies deutlich in der Zusammensetzung des Schweißes der drei Genotypen. In einer Studie mit drei Genotyp-Populationen wurden folgende Werte erhalten:[34]
| Molekül | Genotyp A/A | Genotyp G/A | Genotyp G/G |
|---|---|---|---|
| Fettsäuremethylester | |||
| 3-Hydroxy-3-methylhexansäuremethylester | 0,14 µg[*] | 44,7 µg | 45,8 µg |
| 3-Hydroxy-4-methyloctansäuremethylester | 0,00 µg | 1,84 µg | 0,69 µg |
| (E)-3-Methyl-2-hexensäuremethylester | 0,02 µg | >2,8 µg | >2,8 µg |
| 3-Hydroxyhexansäuremethylester | 0,03 µg | 0,08 µg | 0,13 µg |
| 3-Hydroxyoctansäuremethylester | 0,09 µg | 1,29 µg | 1,98 µg |
| freie Fettsäuren | |||
| Buttersäure | 0,61 µg | 0,99 µg | 1,27 µg |
| Isovaleriansäure | 0,08 µg | 0,23 µg | 0,76 µg |
| 2-Methylbuttersäure | 0,23 µg | 2,60 µg | 12,9 µg |
| Capronsäure | 1,37 µg | 0,85 µg | 1,07 µg |
| Caprylsäure | 1,38 µg | 0,77 µg | 1,42 µg |
| Steroide | |||
| Dehydroepiandrosteron | 2,04 ng/ml | 94,3 ng/ml | 131,7 ng/ml |
| Dehydroepiandrosteronsulfat | 1,85 ng/ml | 41,4 ng/ml | 15,0 ng/ml |
| Androstenon | 302,16 ng/ml | 1070 ng/ml | 1370 ng/ml |
| Testosteron | 0,24 ng/ml | 0,33 ng/ml | 0,46 ng/ml |
| Gesamt-Protein | 277 µg/ml | 826 µg/ml | 1041 µg/ml |
[*] die Massenwerte beziehen sich auf die Menge aus zwei Wattepads
Im Schweiß der Probanden mit dem A/A-Genotyp waren – im Gegensatz zu den beiden anderen Genotypen – keine Präkursoren auf Aminosäurebasis, wie 3-Hydroxy-3-methyl-hexansäure-Gln, (E)-3-Methyl-2-hexensäure-Gln und 3-Methyl-3-sulfanylhexan-1-ol-Cys-Gly per LC/MS nachweisbar.[34] Aus den Daten der Tabelle ist erkennbar, dass der heterozygote G/A-Genotyp in den meisten Fällen zwar weniger Duftstoffe als der G/G-Genotyp produziert, die Gendosis aber ausreichend ist, um typische ABCC11-Substrate in erheblich größeren Mengen in den Schweiß zu transportieren, als beim A/A-Genotyp, bei dem der ABCC11-Transporter nicht vorhanden ist.
In einer detaillierten Analyse einer groß angelegten Studie mit etwa 17.000 Personen im County of Avon, England (Avon Longitudinal Study of Parents and Children, ALSPAC) wurde die Häufigkeit der Nutzung von Deodorants mit dem ABCC11-Genotyp in Korrelation gebracht. Dabei stellte es sich heraus, dass Personen mit dem A/A-Genotyp in der Kategorie „benutze kein Deodorant“ und „benutze Deodorant selten“ mit 22,2 % gegenüber dem G/A- und G/G-Genotyp (4,7 %) fast fünffach überrepräsentiert waren. Dennoch nutzten 77,8 % der Menschen vom A/A-Genotyp Deodorants, obwohl es für sie eigentlich weitgehend sinnlos ist. Der Gebrauch von Deodorants ist bei diesen Personen vermutlich durch soziokulturelle Faktoren bestimmt. Beim heterozygoten G/A-Genotyp war der Deodorantgebrauch signifikant geringer als beim G/G-Genotyp.[69]
Mit den zu Beginn des 21. Jahrhunderts gewonnenen Erkenntnissen über die Transport- und Stoffwechselprozesse der für den Schweißgeruch relevanten ABCC11-Substrate und der Funktion von ABCC11 ergeben sich mögliche neue Ansatzpunkte für die Entwicklung von neuartigen Deodorantien.[27][70] Die Beiersdorf AG meldete 2008 beim Deutschen Patent- und Markenamt kosmetische Zubereitungen mit einem Wirkstoff „aus der Gruppe der ABCC-Inhibitoren“ zum Patent an. Zu den in der Patentschrift aufgeführten Inhibitoren gehören beispielsweise das Antidiabetikum Glibenclamid und das Chemotherapeutikum Lonidamin.[71] Diese Wirkstoffe haben ganz erhebliche Nebenwirkungen, die eine Anwendung in Deodorants als sehr unwahrscheinlich erscheinen lassen. Bisher ist kein Deodorant mit einem ABCC11-Inhibitor auf dem Markt (Stand 2014). Beiersdorf hat im Dezember 2013 die Patentanmeldung zurückgezogen.[72]
Kolostrum und Muttermilch
Apokrine Drüsen finden sich nicht nur in der Nähe von Haarfollikeln, sondern auch in der weiblichen Brust. Diese besteht aus der Brustdrüse (Milchdrüse, Glandula mammaria) und einem bindegewebeartigen Stroma. Die Brustdrüse entwickelt sich aus den Anlagen apokriner Drüsen.[73] Unmittelbar nach dem Gebären produzieren die Milchdrüsen einer Frau die Vormilch, Kolostrum genannt. Im Kolostrum konnten Inhaltsstoffe nachgewiesen werden, die denen des apokrinen Schweißes entsprechen. Sie finden sich zwar auch in der späteren Muttermilch, allerdings in Konzentrationen, die um den Faktor zehn geringer sind. Außerdem konnten sie auch im Fruchtwasser nachgewiesen werden. Insgesamt sind die Konzentrationen aber deutlich niedriger als im apokrinen Schweiß. Diese Inhaltsstoffe gelangen ebenfalls über den ABCC11-Transporter in diese Körperflüssigkeiten. In mehreren Studien wurde nachgewiesen, dass der Geruch von Muttermilch den Saugreflex eines Säuglings stimulieren kann.[74] Die apokrin sezernierten Inhaltsstoffe von Kolostrum, Muttermilch und Fruchtwasser unterliegen einer sehr individuellen Zusammensetzung und Konzentration. Entsprechend individuell ist auch der Geruch dieser drei Körperflüssigkeiten. Es besteht daher die Hypothese, dass diese individuellen Unterschiede möglicherweise schon beim Embryo, oder später beim Säugling, eine Prägung auf die Mutter bewirken können,[27] beziehungsweise, dass Neugeborene zwischen den Körperflüssigkeiten ihrer Mutter und einer anderen Mutter unterscheiden können.[75]
Mütter, die den A/A-Genotyp aufweisen, können deutlich seltener und weniger Vormilch (Kolostrum) über ihre Milchdrüsen produzieren. In einer japanischen Studie wurden 225 Frauen genotypisiert. Dabei waren 155 A/A-homozygot und 70 G/A-heterozygot oder G/G-homozygot. 67,7 % der Frauen (=105 Personen) vom A/A-Genotyp hatten keine Kolostrumbildung, während dies in der anderen Gruppe nur bei 40 % der Fall war. Noch deutlicher war der Unterschied in der Menge an gebildetem Kolostrum. Bei den 50 Frauen vom A/A-Genotyp, die Kolostrum bilden konnten, lag das durchschnittliche Volumen bei 1,6 mL, während in der Gruppe mit mindestens einem G-Allel 4,0 mL gebildet wurden.[76] Welche Auswirkungen dies auf die Ernährung von Säuglingen oder gar evolutionsgeschichtlich hat, beziehungsweise haben konnte, ist noch weitgehend unklar.
Brustkrebsrisiko
Brustkrebs ist mit einem Anteil von etwa 22 % die häufigste Krebserkrankung bei Frauen in den entwickelten Ländern. Dabei gibt es erhebliche regionale Unterschiede. Bei westlichen Frauen ist die Rate an Brustkrebserkrankungen deutlich höher als bei japanischen und taiwanesischen Frauen. Bereits 1971 wurde in der angesehenen Fachzeitschrift Science über einen möglichen Zusammenhang zwischen einem höheren Risiko einer Brustkrebserkrankung und einem feuchten Ohrenschmalz-Phänotyp berichtet.[77] Zu diesem Zeitpunkt war die Wahrscheinlichkeit, an Brustkrebs zu erkranken, für US-amerikanische Frauen um den Faktor vier höher als für japanische Frauen.[78] In den nachfolgenden Jahren wurde über die Korrelation zwischen Cerumen-Phänotyp und Brustkrebsrisiko sehr kontrovers diskutiert.[79][80] Auch neuere Studien liefern widersprüchliche Ergebnisse, die von einem „signifikant erhöhtem Brustkrebsrisiko“ für Trägerinnen von mindestens einem G-Allel ausgehen,[80] und bis zu „kein erhöhtes Risiko“ reichen.[81][82]
Gegenwärtig ist noch unklar, ob der ABCC11-Genotyp wirklich einen Einfluss auf das Risiko einer Brustkrebserkrankung hat. Das Risiko wird durch die G-Allele – wenn überhaupt – vermutlich nur mäßig erhöht.[30] In einer im Jahr 2010 veröffentlichten Studie wurden 270 japanische Brustkrebspatientinnen und 273 gesunde Probandinnen genotypisiert. Dabei waren homozygote G/G-Patientinnen um den Faktor 1,77 häufiger in der Gruppe der Brustkrebspatientinnen vertreten. Bei heterozygoten G/A-Patientinnen lag der Wert bei 1,41.[1]
Über die letzten Jahrzehnte ist die Zahl der Brustkrebserkrankungen in Japan deutlich gestiegen[83] und hat sich seit den 1970er Jahren etwa verdoppelt.[78] Hierfür werden im Wesentlichen geänderte Verhaltensweisen, beispielsweise bei der Ernährung, verantwortlich gemacht.[78]
Populationsgenetik
| Die A-Allelfrequenz in verschiedenen Populationen | ||||
|---|---|---|---|---|
| Population | Häufigkeit | |||
| Afroamerikaner | 0 % | |||
| Afrikaner | 2 % | |||
| Lateinamerikaner | 4 % | |||
| Iberer | 10 % | |||
| Deutsche | 16 % | |||
| Westeuropäer | 17 % | |||
| Osteuropäer | 20 % | |||
| Pazifische Insulaner | 21 % | |||
| Kasachen | 38 % | |||
| Philippiner | 47 % | |||
| Indianer | 50 % | |||
| Vietnamesen | 73 % | |||
| Japaner | 76 % | |||
| Südostasiaten | 78 % | |||
| Mongolen | 87 % | |||
| Chinesen | 94 % | |||
| Koreaner | 100 % | |||
| Quellen:[44] und[84] | ||||
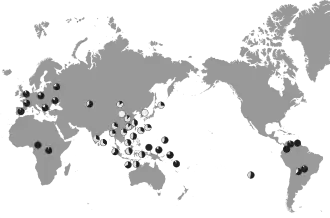
Die Häufigkeit der Einzelnukleotid-Polymorphism 538G→A unterliegt einer geografischen Abhängigkeit. In der Populationsgenetik betrachtet man dabei die Allelfrequenz, die ein Maß für die relative Häufigkeit eines Allels in einer Population ist. Ein Wert von 0,20 bedeutet, dass 20 % der Bevölkerung das entsprechende Allel im Genom hat. In Deutschland haben nach einer Studie mit 132 Probanden etwa 16 % der Bevölkerung mindestens ein A-Allel.[84] Diese 16 % sind allerdings im Wesentlichen der heterozygote G/A-Typ. Der weist – gemäß dem rezessiven Erbgang – den gleichen Phänotyp wie der G/G-Typ auf, das heißt gelb-brauner, feuchter Ohrenschmalz und Körpergeruch. Der homozygote A/A-Typ, mit weißem, trockenem Ohrenschmalz und sehr schwachem Körpergeruch, ist deutlich seltener. Sein Anteil errechnet sich aus dem Quadrat der Allelfrequenz. Im Beispiel Deutschland sind dies 0,16² = 0,0256, das heißt nur etwa 2,6 % der Bevölkerung Deutschlands hat den A/A-Genotyp. Wesentlich höher ist die A-Allelfrequenz von ABCC11 beispielsweise in Südkorea. Dort liegt sie je nach Studie zwischen 0,98 und 1,0.[84] Entsprechend haben über 96 % aller Koreaner den A/A-Genotyp. Kloniert man Mitglieder weiterer Ethnien, so bekommt man eine geografische Verteilung der Allelfrequenz von ABCC11. Hierbei zeigt sich eine deutliche Zunahme in östlicher, sowie nördlicher Richtung. In Afrika, der „Wiege der Menschheit“, ist die A-Allelfrequenz extrem niedrig. In neun Studien mit insgesamt 403 Probanden war kein einziger Träger eines A-Allels von ABCC11. Nur in einer Studie die mit 45 Probanden aus dem Volk der Chagga durchgeführt wurde, hatte einer ein A-Allel.[84]
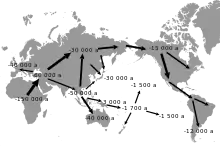
Die Out-of-Africa-Theorie, also die Annahme, dass sich die Gattung Homo vom Heimatkontinent Afrika aus über die ganze Welt verbreitete, gilt durch mehrere, unterschiedliche Indizien, wie Fossilienfunde, Populationsgenetik und Sprachgeographie für die Art Homo sapiens, als wissenschaftlich allgemein akzeptiert (siehe dazu Ausbreitung des Menschen).[85] Die Wege der urzeitlichen Bevölkerungsströme nach Ost-, Zentral- und Südostasien, sowie in die Arktis, nach Amerika und auf pazifische Inseln sind allerdings noch nicht vollständig erforscht. Es wurden dafür zwei Routen vorgeschlagen: Nach der Abtrennung vom mit den Bewohnern Europas, Nordafrikas und Vorderasiens gebildeten gemeinsamen Hauptstrom vor 150.000 bis 60.000 Jahren[86] migrierten sie nach Südasien. Man nimmt dabei an, dass ein Zweig dieses Bevölkerungsstroms dort ansässig wurde beziehungsweise weiter in süd-östliche Richtung durch Sundaland wanderte und schließlich vor 50.000 bis 46.000 Jahren Sahul – den heutigen Kontinent Australien – erreichte. In einer letzten Migrationswelle vor 3000 bis 1500 Jahren wurde schließlich Ozeanien besiedelt. Ein anderer Zweig der Bevölkerung Ost-, Zentral- und Südostasiens migrierte in nördliche Richtung und erreichte dabei das Gebiet um den Baikalsee, entlang dem Altai. Alternativ zu diesem Weg ist ein Teil möglicherweise direkt vom gemeinsamen Hauptstrom nach Sibirien migriert oder ist von Südostasien nach Nordostasien eingewandert.[87] Es wird ferner vermutet, dass durch die Vergletscherung während des letzten glacialen Maximums vor 30.000 bis 15.000 Jahren kleinere Stämme im Norden weitgehend isoliert lebten.[1]
Man geht heute davon aus, dass die Einzelnukleotid-Polymorphismus-induzierte Mutation 538G→A vermutlich vor etwa 40.000 Jahren in einem Stamm der urzeitlichen nördlichen Mongolei auftrat.[88] Betrachtet man den Nord-Süd-Gradienten in der Verteilung des 538A-Allels mit seinem Maximum in Nordchina und Korea über Japan nach Südostasien, so spiegelt dies wahrscheinlich die Migration urzeitlicher nordmongolischer Bevölkerungsteile wider. Der Ost-West-Gradient von Sibirien nach Europa[89] ist dagegen deutlich späteren Migrations- beziehungsweise Invasionswellen, wie beispielsweise dem Mongolensturm, geschuldet.[1] Eine ähnliche geografische Verteilung liegt bei einer Mutation im mitochondrialen ALDH2-Gen vor, das für das Enzym Aldehyd-Dehydrogenase 2 kodiert. Der Gendefekt ist für eine erhöhte Alkoholempfindlichkeit bei den Betroffenen verantwortlich.[90]
Besiedlung Japans
Japan wurde nach dem gegenwärtigen Kenntnisstand durch zwei Migrationswellen bevölkert. Die erste Welle fand nach der letzten Eiszeit, in der Jōmon-Zeit, vor etwa 13 000 bis 3000 Jahren statt. Die zweite Migrationswelle kam während der Yayoi-Zeit, vor etwa 2500 bis 1700 Jahren. Die Jōmon wurden dabei von den Yayoi entweder verdrängt oder assimiliert.[91][92][93] Eine Hypothese besagt, dass der trockene Ohrschmalz-Genotyp von den Yayoi nach Japan kam, während bei den Jōmon der feuchte Typ dominierte. Für diese Hypothese spricht, dass der 538G-Genotyp in entlegenen Gebieten – den Rückzugsgebieten der Jōmon – deutlich verbreiteter als in den Yoyoi-Gebieten ist. Daraus lässt sich auch schließen, dass die genetische Durchmischung von Jōmon und Yayoi in Japan noch nicht abgeschlossen ist.[1]
Aus der Häufigkeit des 538G-Allels und des damit assoziierten feuchten Ohrenschmalz-Genotyps lässt sich bei den Ainu, den Ureinwohner Japans, und der Bevölkerung der Ryūkyū-Inseln („Okinawa“) schließen, dass sie keine direkten Nachfahren der Yayoi sind, die aus der nördlichen Mongolei und Sibirien nach Japan kamen. Auch andere Studien kommen zu dem Ergebnis, dass die Ainu genetisch erhebliche Unterschiede zu anderen ostasiatischen Populationen aufweisen.[94][95]
Selektionsvorteil oder Gendrift

Weitgehend unklar ist, warum sich der Einzelnukleotid-Polymorphismus 538G→A so erfolgreich im asiatischen Raum ausbreiten konnte.[96] Zwei unterschiedliche Erklärungsansätze gibt es derzeit dafür. Der eine Ansatz geht dabei von einem Selektionsvorteil für den A/A-Genotyp aus, der andere von einer Gendrift.
Bei einer großen Population muss ein ausgesprochen hoher Selektionsdruck vorliegen, damit sich eine Mutation in vergleichsweise kurzer Zeit erfolgreich durchsetzen kann. Im Fall des A/A-Genotyps ist ein Selektionsvorteil über die Zusammensetzung des Ohrenschmalzes schwer vorstellbar. Wahrscheinlicher ist, dass ein anderer Einzelnukleotid-Polymorphismus rs6500380 im LONP2-Gen, der ein starkes Kopplungsungleichgewicht zu rs17822931 aufweist (r²=0,91) und folglich sehr häufig gemeinsam in den entsprechenden Populationen auftritt, den Selektionsvorteil bietet. LONP2 kodiert für das Enzym peroxisomale LON-Protease Homolog 2. Nach dem gegenwärtigen Kenntnisstand hat rs6500380 allerdings eine deutlich geringere funktionelle Bedeutung als rs17822931, so dass ein möglicher Selektionsvorteil sehr wahrscheinlich doch von rs17822931 kommt.[88] Der könnte sich unter anderem daraus ergeben, dass sich der A/A-Genotyp der urzeitlichen Bevölkerung Nordostasiens durch die geringere Schweißabsonderung besser an das erheblich kältere Klima der Region anpassen konnte.[44] Möglich ist auch, dass die Veränderung der Zusammensetzung des Colostrums oder eine noch unbekannte Dysfunktion von ABCC11 in anderen Geweben, wie beispielsweise Hoden, Leber, Placenta, Lunge oder Gehirn, in denen ebenfalls ABCC11 exprimiert wird, einen Selektionsvorteil bot. Am derzeit plausibelsten scheint die Hypothese eines Vorteils bei der sexuellen Selektion. Vergangene östliche Kulturen hatten – im Vergleich zu denen im Westen – eine deutlich ausgeprägtere Tradition der Sauberkeit und Hygiene.[97] Der körpergeruchsarme A/A-Genotyp hatte möglicherweise bei der Partnerwahl einen signifikanten Vorteil gegenüber dem ‚stärker riechenden‘ G/G- beziehungsweise G/A-Genotyp. Eine Hypothese, die zumindest aus heutiger Sicht, in der intensiver Körpergeruch auch in westlichen Kulturen als sozial anstößig wahrgenommen wird, durchaus plausibel erscheint.[34] Simulationsrechnungen aus dem Jahr 2011 kommen zu dem Ergebnis, dass die Mutation von rs17822931-A vor etwa 2006 Generationen stattgefunden hat, bei einem Konfidenzintervall von 95 % im Bereich von 1023 bis 3901 Generationen. Dazu wurde ein Selektionskoeffizient von 0,01 angenommen.[88]
Geht man von eher kleinen Populationen aus, können völlig andere evolutionäre Effekte als der Selektionsdruck zu plötzlichen und massiven Änderungen bei der Häufigkeit eines Genotyps in den nachfolgenden Generationen sorgen. Die vererbten Mutationen unterliegen dabei nicht der Selektion. Sie sind neutral, das heißt, sie haben keinen Einfluss auf die Fitness des Phänotyps (Survival of the Fittest). Diese Form der Verschiebung der Allelfrequenz, das heißt der Häufigkeit eines Allels in einer Population, nennt man Gendrift. Der ‚Erfolg‘ einer Mutation hängt von der Größe der Population und dem Selektionsdruck ab.[98] Gendrift und Selektion sind Evolutionsfaktoren, die gleichzeitig wirken. In großen Populationen dominiert die Selektion, in kleinen die Gendrift.[99] Bei der Gendrift handelt es sich um einen statistischen, zufälligen Effekt, der durch kleine Populationen begünstigt wird. Kleine, abgeschlossene Populationen stellen einen genetischen Flaschenhals dar. Dieser kann beispielsweise durch starke klimatische Änderungen, Seuchen oder Hungersnöte hervorgerufen werden. Aber auch weniger dramatische Änderungen, wie beispielsweise das Abwandern von wenigen Individuen aus einer großen Population, der sogenannte Gründereffekt, können zu einem genetischen Flaschenhals führen. Dabei können sich Allelfrequenzen einstellen, die im Widerspruch zur natürlichen Selektion stehen. Auch für die erfolgreiche Verbreitung des A/A-Genotyps wird ein genetischer Flaschenhals und eine damit verbundene Gendrift als mögliche Ursache diskutiert.[100][101][1]
Fußnoten
- Genome Reference Consortium human genome (build 37)
- Es gibt auch Studienergebnisse, die zu dem Ergebnis kommen, dass die funktionslose ABCC11-Variante Gly180Arg dennoch an der Zellmembran exprimiert wird (siehe Annette Martin, Matthias Saathoff, Fabian Kuhn, Heiner Max, Lara Terstegen, Andreas Natsch: A functional ABCC11 allele is essential in the biochemical formation of human axillary odor. In: Journal of Investigative Dermatology. Band 130, Nr. 2, 2010, S. 529–540, doi:10.1038/jid.2009.254, PMID 19710689.).
Einzelnachweise
- T. Ishikawa, Y. Toyoda u. a.: Pharmacogenetics of human ABC transporter ABCC11: new insights into apocrine gland growth and metabolite secretion. In: Frontiers in genetics. Band 3, 2012, ISSN 1664-8021, S. 306, doi:10.3389/fgene.2012.00306, PMID 23316210, PMC 3539816 (freier Volltext). (Open Access)
- UCSC Genome Browser on Human Feb. 2009 (GRCh37/hg19) Assembly. Abgerufen am 1. September 2014
- H. Yabuuchi, H. Shimizu u. a.: Multiple splicing variants of two new human ATP-binding cassette transporters, ABCC11 and ABCC12. In: Biochemical and biophysical research communications. Band 288, Nummer 4, November 2001, ISSN 0006-291X, S. 933–939, doi:10.1006/bbrc.2001.5865, PMID 11688999.
- J. Tammur, C. Prades u. a.: Two new genes from the human ATP-binding cassette transporter superfamily, ABCC11 and ABCC12, tandemly duplicated on chromosome 16q12. In: Gene. Band 273, Nummer 1, Juli 2001, ISSN 0378-1119, S. 89–96, PMID 11483364.
- T. K. Bera, S. Lee u. a.: MRP8, a new member of ABC transporter superfamily, identified by EST database mining and gene prediction program, is highly expressed in breast cancer. In: Molecular medicine. Band 7, Nummer 8, August 2001, ISSN 1076-1551, S. 509–516, PMID 11591886, PMC 1950066 (freier Volltext).
- uniprot.org: Homo sapiens (Human) abgerufen am 1. September 2014
- uniprot.org: Pan troglodytes (Chimpanzee) abgerufen am 1. September 2014
- uniprot.org: Gorilla gorilla gorilla (Lowland gorilla) abgerufen am 1. September 2014
- uniprot.org: Pongo abelii (Sumatran orangutan) (Pongo pygmaeus abelii) abgerufen am 1. September 2014
- uniprot.org: Nomascus leucogenys (Northern white-cheeked gibbon) (Hylobates leucogenys) abgerufen am 1. September 2014
- uniprot.org: Macaca mulatta (Rhesus macaque) abgerufen am 1. September 2014
- uniprot.org: Callithrix jacchus (White-tufted-ear marmoset) abgerufen am 1. September 2014
- uniprot.org: Otolemur garnettii (Small-eared galago) (Garnett's greater bushbaby) abgerufen am 1. September 2014
- uniprot.org: Loxodonta africana (African elephant) abgerufen am 1. September 2014
- uniprot.org: Canis familiaris (Dog) (Canis lupus familiaris) abgerufen am 1. September 2014
- uniprot.org: Mustela putorius furo (European domestic ferret) (Mustela furo) abgerufen am 1. September 2014
- uniprot.org: Ailuropoda melanoleuca (Giant panda) abgerufen am 1. September 2014
- uniprot.org: Felis catus (Cat) (Felis silvestris catus) abgerufen am 1. September 2014
- uniprot.org: Bos taurus (Bovine) abgerufen am 1. September 2014
- uniprot.org: Ovis aries (Sheep) abgerufen am 1. September 2014
- uniprot.org: Equus caballus (Horse) abgerufen am 1. September 2014
- uniprot.org: Sarcophilus harrisii (Tasmanian devil) (Sarcophilus laniarius) abgerufen am 1. September 2014
- omabrowser.org: Entry HUMAN10683 abgerufen am 1. September 2014
- H. Shimizu, H. Taniguchi u. a.: Characterization of the mouse Abcc12 gene and its transcript encoding an ATP-binding cassette transporter, an orthologue of human ABCC12. In: Gene. Band 310, Mai 2003, ISSN 0378-1119, S. 17–28, PMID 12801629.
- N. Ono, I. Van der Heijden u. a.: Multidrug resistance-associated protein 9 (ABCC12) is present in mouse and boar sperm. In: The Biochemical journal. Band 406, Nummer 1, August 2007, ISSN 1470-8728, S. 31–40, doi:10.1042/BJ20070292, PMID 17472575, PMC 1948986 (freier Volltext).
- ABCC11. In: Online Mendelian Inheritance in Man. (englisch)
- Tim Baumann: Molekulare Charakterisierung von Stoffwechsel- und Transportprozessen in der apokrinen Schweißdrüse für die Bildung von Schweißgeruchsvorstufen. Dissertation, Ernst-Moritz-Arndt-Universität Greifswald, 2012, S. 1–16.
- T. Kitano, I. Yuasa u. a.: Allele frequencies of a SNP and a 27-bp deletion that are the determinant of earwax type in the ABCC11 gene. In: Legal medicine. Band 10, Nummer 2, März 2008, S. 113–114, ISSN 1344-6223. doi:10.1016/j.legalmed.2007.08.003. PMID 18037328.
- M. Honorat, R. Terreux u. a.: Localization of putative binding sites for cyclic guanosine monophosphate and the anti-cancer drug 5-fluoro-2'-deoxyuridine-5'-monophosphate on ABCC11 in silico models. In: BMC structural biology. Band 13, 2013, S. 7, ISSN 1472-6807. doi:10.1186/1472-6807-13-7. PMID 23641929. PMC 3668285 (freier Volltext). (Open Access)
- Y. Toyoda, T. Ishikawa: Pharmacogenomics of human ABC transporter ABCC11 (MRP8): potential risk of breast cancer and chemotherapy failure. In: Anti-Cancer Agents in Medicinal Chemistry. Band 10, Nummer 8, Oktober 2010, ISSN 1875-5992, S. 617–624, PMID 21182469, PMC 3319924 (freier Volltext). (Review-Artikel im Open Access, CC BY 2.5)
- Y. Guo, E. Kotova u. a.: MRP8, ATP-binding cassette C11 (ABCC11), is a cyclic nucleotide efflux pump and a resistance factor for fluoropyrimidines 2',3'-dideoxycytidine and 9'-(2'-phosphonylmethoxyethyl)adenine. In: The Journal of biological chemistry. Band 278, Nummer 32, August 2003, ISSN 0021-9258, S. 29509–29514, doi:10.1074/jbc.M304059200, PMID 12764137.
- Z. S. Chen, Y. Guo u. a.: Transport of bile acids, sulfated steroids, estradiol 17-beta-D-glucuronide, and leukotriene C4 by human multidrug resistance protein 8 (ABCC11). In: Molecular pharmacology. Band 67, Nummer 2, Februar 2005, ISSN 0026-895X, S. 545–557, doi:10.1124/mol.104.007138, PMID 15537867.
- M. Bortfeld, M. Rius u. a.: Human multidrug resistance protein 8 (MRP8/ABCC11), an apical efflux pump for steroid sulfates, is an axonal protein of the CNS and peripheral nervous system. In: Neuroscience. Band 137, Nummer 4, 2006, ISSN 0306-4522, S. 1247–1257, doi:10.1016/j.neuroscience.2005.10.025, PMID 16359813.
- A. Martin, M. Saathoff u. a.: A functional ABCC11 allele is essential in the biochemical formation of human axillary odor. In: The Journal of investigative dermatology. Band 130, Nummer 2, Februar 2010, ISSN 1523-1747, S. 529–540, doi:10.1038/jid.2009.254, PMID 19710689.
- Helga Fritsch, Wolfgang Kühnel: Taschenatlas der Anatomie. Band 2: Innere Organe. 11. Auflage, Georg Thieme Verlag, 2013, ISBN 3-13-150921-X, S. 430. eingeschränkte Vorschau in der Google-Buchsuche
- T. Magdy, R. Arlanov u. a.: ABCC11/MRP8 polymorphisms affect 5-fluorouracil-induced severe toxicity and hepatic expression. In: Pharmacogenomics. Band 14, Nummer 12, September 2013, ISSN 1744-8042, S. 1433–1448, doi:10.2217/pgs.13.139, PMID 24024896.
- I. Bièche, I. Girault u. a.: Relationship between intratumoral expression of genes coding for xenobiotic-metabolizing enzymes and benefit from adjuvant tamoxifen in estrogen receptor alpha-positive postmenopausal breast carcinoma. In: Breast cancer research. Band 6, Nummer 3, 2004, S. R252–R263, ISSN 1465-542X, doi:10.1186/bcr784, PMID 15084249, PMC 400681 (freier Volltext).
- M. Honorat, A. Mesnier u. a.: ABCC11 expression is regulated by estrogen in MCF7 cells, correlated with estrogen receptor alpha expression in postmenopausal breast tumors and overexpressed in tamoxifen-resistant breast cancer cells. In: Endocrine-related cancer. Band 15, Nummer 1, März 2008, ISSN 1351-0088, S. 125–138, doi:10.1677/ERC-07-0189, PMID 18310281.
- Gergely Szakács, Kenneth Kin Wah To u. a.: Multidrug Resistance Mediated by MDR-ABC Transporters. In: Kapil Metha, Zahid H. Siddik (Hrsg.): Drug Resistance in Cancer Cells. Springer Science & Business Media, 2009, ISBN 0-387-89445-4, S. 1–20. eingeschränkte Vorschau in der Google-Buchsuche
- T. Oguri, Y. Bessho u. a.: MRP8/ABCC11 directly confers resistance to 5-fluorouracil. In: Molecular cancer therapeutics. Band 6, Nummer 1, Januar 2007, ISSN 1535-7163, S. 122–127, doi:10.1158/1535-7163.MCT-06-0529, PMID 17237272.
- ensembl.org: rs17822931 SNP abgerufen am 1. September 2014
- H. Nakagawa, Y. Toyoda u. a.: Ubiquitin-mediated proteasomal degradation of ABC transporters: a new aspect of genetic polymorphisms and clinical impacts. In: Journal of pharmaceutical sciences. Band 100, Nummer 9, September 2011, S. 3602–3619, ISSN 1520-6017. doi:10.1002/jps.22615. PMID 21567408. (Review).
- Y. Toyoda, A. Sakurai u. a.: Earwax, osmidrosis, and breast cancer: why does one SNP (538G>A) in the human ABC transporter ABCC11 gene determine earwax type? In: FASEB journal. Band 23, Nummer 6, Juni 2009, ISSN 1530-6860, S. 2001–2013, doi:10.1096/fj.09-129098, PMID 19383836.
- K. Yoshiura, A. Kinoshita u. a.: A SNP in the ABCC11 gene is the determinant of human earwax type. In: Nature genetics. Band 38, Nummer 3, März 2006, ISSN 1061-4036, S. 324–330, doi:10.1038/ng1733, PMID 16444273.
- M. Stone, R. S. Fulghum: Bactericidal activity of wet cerumen. In: The Annals of otology, rhinology, and laryngology. Band 93, Nummer 2 Pt 1, 1984 Mar-Apr, S. 183–186, ISSN 0003-4894. PMID 6370076.
- N. H. Shehadeh, A. M. Kligman: The effect of topical antibacterial agents on the bacterial flora of the axilla. In: The Journal of investigative dermatology. Band 40, Januar 1963, ISSN 0022-202X, S. 61–71, PMID 13977054.
- M. J. Sergeant: Female perception of male body odor. In: Vitamins and hormones. Band 83, 2010, ISSN 0083-6729, S. 25–45, doi:10.1016/S0083-6729(10)83002-X, PMID 20831941. (Review).
- M. F. Bhutta: Sex and the nose: human pheromonal responses. In: Journal of the Royal Society of Medicine. Band 100, Nummer 6, Juni 2007, ISSN 0141-0768, S. 268–274, doi:10.1258/jrsm.100.6.268, PMID 17541097, PMC 1885393 (freier Volltext). (Review).
- K. Saga: Structure and function of human sweat glands studied with histochemistry and cytochemistry. In: Progress in histochemistry and cytochemistry. Band 37, Nummer 4, 2002, ISSN 0079-6336, S. 323–386, PMID 12365351. (Review).
- W. B. Cutler, G. Preti u. a.: Human axillary secretions influence women's menstrual cycles: the role of donor extract from men. In: Hormones and behavior. Band 20, Nummer 4, Dezember 1986, ISSN 0018-506X, S. 463–473, PMID 3793027.
- T. K. Saxton, A. Lyndon u. a.: Evidence that androstadienone, a putative human chemosignal, modulates women's attributions of men's attractiveness. In: Hormones and behavior. Band 54, Nummer 5, November 2008, ISSN 1095-6867, S. 597–601, doi:10.1016/j.yhbeh.2008.06.001, PMID 18601928.
- K. Ackerl, M. Atzmueller, K. Grammer: The scent of fear. In: Neuro endocrinology letters. Band 23, Nummer 2, April 2002, ISSN 0172-780X, S. 79–84, PMID 12011790.
- B. M. Pause, A. Ohrt u. a.: Positive emotional priming of facial affect perception in females is diminished by chemosensory anxiety signals. In: Chemical senses. Band 29, Nummer 9, November 2004, ISSN 0379-864X, S. 797–805, doi:10.1093/chemse/bjh245, PMID 15574815.
- A. Prehn, A. Ohrt u. a.: Chemosensory anxiety signals augment the startle reflex in humans. In: Neuroscience letters. Band 394, Nummer 2, Februar 2006, ISSN 0304-3940, S. 127–130, doi:10.1016/j.neulet.2005.10.012, PMID 16257486.
- L. R. Mujica-Parodi, H. H. Strey u. a.: Chemosensory cues to conspecific emotional stress activate amygdala in humans. In: PloS one. Band 4, Nummer 7, 2009, S. e6415, ISSN 1932-6203, doi:10.1371/journal.pone.0006415, PMID 19641623, PMC 2713432 (freier Volltext).
- J. N. Labows, G. Preti u. a.: Steroid analysis of human apocrine secretion. In: Steroids. Band 34, Nummer 3, September 1979, ISSN 0039-128X, S. 249–258, PMID 158859.
- M. Julesz: New advances in the field of androgenic steroidogenesis of the human skin. In: Acta medica Academiae Scientiarum Hungaricae. Band 25, Nummer 3, 1968, ISSN 0001-5989, S. 273–285, PMID 4238397.
- R. Emter, A. Natsch: The sequential action of a dipeptidase and a beta-lyase is required for the release of the human body odorant 3-methyl-3-sulfanylhexan-1-ol from a secreted Cys-Gly-(S) conjugate by Corynebacteria. In: The Journal of biological chemistry. Band 283, Nummer 30, Juli 2008, ISSN 0021-9258, S. 20645–20652, doi:10.1074/jbc.M800730200, PMID 18515361, PMC 3258934 (freier Volltext).
- C. Starkenmann, Y. Niclass u. a.: Identification of the precursor of (S)-3-methyl-3-sulfanylhexan-1-ol, the sulfury malodour of human axilla sweat. In: Chemistry & biodiversity. Band 2, Nummer 6, Juni 2005, ISSN 1612-1880, S. 705–716, doi:10.1002/cbdv.200590048, PMID 17192014.
- X. N. Zeng, J. J. Leyden u. a.: Analysis of characteristic odors from human male axillae. In: Journal of chemical ecology. Band 17, Nummer 7, Juli 1991, ISSN 0098-0331, S. 1469–1492, doi:10.1007/BF00983777, PMID 24257805.
- R. A. Decréau, C. M. Marson u. a.: Production of malodorous steroids from androsta-5,16-dienes and androsta-4,16-dienes by Corynebacteria and other human axillary bacteria. In: The Journal of steroid biochemistry and molecular biology. Band 87, Nummer 4–5, Dezember 2003, ISSN 0960-0760, S. 327–336, PMID 14698214.
- D. B. Gower, S. Bird u. a.: Axillary 5 alpha-androst-16-en-3-one in men and women: relationships with olfactory acuity to odorous 16-androstenes. In: Experientia. Band 41, Nummer 9, September 1985, ISSN 0014-4754, S. 1134–1136, PMID 4043321.
- A. Natsch, S. Derrer u. a.: A broad diversity of volatile carboxylic acids, released by a bacterial aminoacylase from axilla secretions, as candidate molecules for the determination of human-body odor type. In: Chemistry & biodiversity. Band 3, Nummer 1, Januar 2006, ISSN 1612-1880, S. 1–20, doi:10.1002/cbdv.200690015, PMID 17193210.
- P. Schiefferdecker: Die Hautdrüsen des Menschen und der Säugetiere, ihre biologische und rassenanatomische Bedeutung, sowie die Muscularis sexualis. In: Zoologica. Nummer 72, 1922, S. 1–154.
- B. Adachi: Das Ohrenschmalz als Rassenmerkmal und der Rassengeruch ('Achselgeruch') nebst dem Rassenunterschied der Schweissdrüsen. In: Z Rassenheilkunde. Band 6, 1937, S. 273–307.
- Dieter E. Zimmer: Riechen. Ein Wissenschaftsreport. In: Zeitmagazin Nummer 44, vom 23. Oktober 1987
- Dayle Hayes, Rachel Laudan: Food and Nutrition. Marshall Cavendish, 2009, ISBN 0-7614-7820-5, S. 141. eingeschränkte Vorschau in der Google-Buchsuche
- Andrew J. Eckstein: Japan’s National Identity: Nationalists or Not? Lehigh University, International Relations-163, vom 21. März 1999
- S. Rodriguez, C. D. Steer u. a.: Dependence of deodorant usage on ABCC11 genotype: scope for personalized genetics in personal hygiene. In: The Journal of investigative dermatology. Band 133, Nummer 7, Juli 2013, S. 1760–1767, ISSN 1523-1747. doi:10.1038/jid.2012.480. PMID 23325016. PMC 3674910 (freier Volltext).
- T. Baumann, Z. Jovanovic u. a.: ABCC11 – The Key Anti-Odor Target. In: Flavour. April 2014, 3:P16. doi:10.1186/2044-7248-3-S1-P16 (Open Access)
- Patentanmeldung DE102008001811A1: Kosmetische Zubereitungen zur Verminderung von Schweißgeruch mit ABCC-Modulatoren. Angemeldet am 15. Mai 2008, veröffentlicht am 19. November 2009, Anmelder: Beiersdorf AG, Erfinder: A. Martin, M. Saathoff, L. Terstegen.
- Kosmetische Zubereitungen zur Verminderung von Schweißgeruch mit ABCC-Modulatoren. (Application deemed withdrawn, or ip right lapsed, due to non-payment of renewal fee)
- Helga Fritsch, Wolfgang Kühnel: Taschenatlas der Anatomie. Band 2: Innere Organe. 11. Auflage, Georg Thieme Verlag, 2013, ISBN 3-13-150921-X, S. 436. eingeschränkte Vorschau in der Google-Buchsuche
- L. Marlier, B. Schaal: Human newborns prefer human milk: conspecific milk odor is attractive without postnatal exposure. In: Child development. Band 76, Nummer 1, 2005 Jan-Feb, S. 155–168, ISSN 0009-3920. doi:10.1111/j.1467-8624.2005.00836.x. PMID 15693764.
- C. Hartmann, S. Doucet u. a.: Human sweat odour conjugates in human milk, colostrum and amniotic fluid. In: Food Chemistry. Band 135, Nummer 1, 2012, S. 228–233. doi:10.1016/j.foodchem.2012.04.077
- K. Miura, K. Yoshiura u. a.: A strong association between human earwax-type and apocrine colostrum secretion from the mammary gland. In: Human genetics. Band 121, Nummer 5, Juni 2007, S. 631–633, ISSN 0340-6717. doi:10.1007/s00439-007-0356-9. PMID 17394018.
- N. L. Petrakis: Cerumen genetics and human breast cancer. In: Science. Band 173, Nummer 3994, Juli 1971, S. 347–349, ISSN 0036-8075. PMID 5559907.
- K. Saika, T. Sobue: Epidemiology of Breast Cancer in Japan and the US In: JMAJ. Band 52, Nummer 1, 2009, S. 39–44.
- R. Ing, L. Petrakis, H. C. Ho: Evidence against association between wet cerumen and breast cancer. In: Lancet. Band 1, Nummer 7793, Januar 1973, S. 41, ISSN 0140-6736. PMID 4118561.
- I. Ota, A. Sakurai u. a.: Association between breast cancer risk and the wild-type allele of human ABC transporter ABCC11. In: Anticancer Research. Band 30, Nummer 12, Dezember 2010, S. 5189–5194, ISSN 1791-7530. PMID 21187511.
- J. Beesley, S. E. Johnatty u. a.: No evidence for an association between the earwax-associated polymorphism in ABCC11 and breast cancer risk in Caucasian women. In: Breast Cancer Research and Treatment. Band 126, Nummer 1, Februar 2011, S. 235–239, ISSN 1573-7217. doi:10.1007/s10549-010-1292-2. PMID 21165769.
- T. Lang, C. Justenhoven u. a.: The earwax-associated SNP c.538G>A (G180R) in ABCC11 is not associated with breast cancer risk in Europeans. In: Breast Cancer Research and Treatment. Band 129, Nummer 3, Oktober 2011, S. 993–999, ISSN 1573-7217. doi:10.1007/s10549-011-1613-0. PMID 21655989.
- K. Wakai, S. Suzuki u. a.: Epidemiology of breast cancer in Japan. In: International journal of epidemiology. Band 24, Nummer 2, April 1995, S. 285–291, ISSN 0300-5771. PMID 7635587.
- ALFRED: Allele Frequency For Polymorphic Site: G/A SNP (Gly180Arg). The ALlele FREquency Database, abgerufen am 13. September 2014
- L. L. Cavalli-Sforza: The Human Genome Diversity Project: past, present and future. In: Nature reviews. Genetics. Band 6, Nummer 4, April 2005, ISSN 1471-0056, S. 333–340, doi:10.1038/nrg1596, PMID 15803201. (Review).
- M. Nei: Evolution of human races at the gene level. In: Progress in clinical and biological research. Band 103 Pt A, 1982, ISSN 0361-7742, S. 167–181, PMID 7163193.
- The HUGO Pan-Asian SNP Consortium: Mapping human genetic diversity in Asia. In: Science. Band 326, Nummer 5959, Dezember 2009, ISSN 1095-9203, S. 1541–1545, doi:10.1126/science.1177074, PMID 20007900.
- J. Ohashi, I. Naka, N. Tsuchiya: The impact of natural selection on an ABCC11 SNP determining earwax type. In: Molecular biology and evolution. Band 28, Nummer 1, Januar 2011, ISSN 1537-1719, S. 849–857, doi:10.1093/molbev/msq264, PMID 20937735.
- V. A. Spitsyn, I. Afanasèva: [Genetic geography of inherited dimorphism of ear wax by its consistency]. In: Genetika. Band 25, Nummer 10, Oktober 1989, ISSN 0016-6758, S. 1854–1860, PMID 2620812. (in russischer Sprache)
- Q. Xiao, H. Weiner, D. W. Crabb: The mutation in the mitochondrial aldehyde dehydrogenase (ALDH2) gene responsible for alcohol-induced flushing increases turnover of the enzyme tetramers in a dominant fashion. In: The Journal of clinical investigation. Band 98, Nummer 9, November 1996, ISSN 0021-9738, S. 2027–2032, doi:10.1172/JCI119007, PMID 8903321, PMC 507646 (freier Volltext).
- M. Iizuka, T. Nakahashi: A population genetic study on the transition from Jomon people to Yayoi people. In: Genes & genetic systems. Band 77, Nummer 5, Oktober 2002, S. 287–300, ISSN 1341-7568. PMID 12441640.
- D. H. Temple, B. M. Auerbach u. a.: Variation in limb proportions between Jomon foragers and Yayoi agriculturalists from prehistoric Japan. In: American journal of physical anthropology. Band 137, Nummer 2, Oktober 2008, S. 164–174, ISSN 1096-8644. doi:10.1002/ajpa.20853. PMID 18484628.
- D. H. Temple: Patterns of systemic stress during the agricultural transition in prehistoric Japan. In: American journal of physical anthropology. Band 142, Nummer 1, Mai 2010, S. 112–124, ISSN 1096-8644. doi:10.1002/ajpa.21208. PMID 19953616.
- K. Tokunaga, J. Ohashi u. a.: Genetic link between Asians and native Americans: evidence from HLA genes and haplotypes. In: Human immunology. Band 62, Nummer 9, September 2001, S. 1001–1008, ISSN 0198-8859. PMID 11543902.
- A. Tajima, M. Hayami u. a.: Genetic origins of the Ainu inferred from combined DNA analyses of maternal and paternal lineages. In: Journal of human genetics. Band 49, Nummer 4, 2004, S. 187–193, ISSN 1434-5161. doi:10.1007/s10038-004-0131-x. PMID 14997363.
- Mark A. Jobling, Edward Hollox u. a.: Human Evolutionary Genetics. 2. Auflage, Garland Science, 2013, ISBN 1-317-95226-X, S. 336. eingeschränkte Vorschau in der Google-Buchsuche
- Charles D. Benn: China’s Golden Age: Everyday Life During the Tang Dynasty. Oxford University Press, 2004, ISBN 0-19-517665-0, S. 344.
- M. C. Whitlock: Fixation probability and time in subdivided populations. In: Genetics. Band 164, Nummer 2, Juni 2003, ISSN 0016-6731, S. 767–779, PMID 12807795, PMC 1462574 (freier Volltext).
- S. P. Otto, M. C. Whitlock: The probability of fixation in populations of changing size. In: Genetics. Band 146, Nummer 2, Juni 1997, ISSN 0016-6731, S. 723–733, PMID 9178020, PMC 1208011 (freier Volltext).
- Razib Khan: Body odor, Asians, and earwax. Bei: discovermagazine.com vom 25. Oktober 2010
- Doug Van Hoewyk: Using a Case Study Article on Earwax to Enhance Understanding and Interest in Genetics. In: Journal of College Science Teaching. Band 41, Nummer 6, 2012, S. 60–62.
Weblinks
- twn: Menschen ohne Achselduft: Die Geruchlosen In: Spiegel Online vom 18. Januar 2013
- ATP-Binding Cassette, Sub-Family C (CFTR/MRP), Member 11 bei GeneCards (englisch)
- ABCC11 ATP-binding cassette, sub-family C (CFTR/MRP), member 11, Homo sapiens (human) beim NCBI (englisch)
- Homo sapiens ATP-binding cassette, sub-family C (CFTR/MRP), member 11 (ABCC11), RefSeqGene on chromosome 16 die gesamte Nukleotid- und Protein-Sequenz von ABCC11 beim NCBI (englisch)