Endoplasmatisches Retikulum
Das endoplasmatische Retikulum (ER, endoplasmatisch „im Cytoplasma“, lat. reticulum „Wurfnetz“) ist ein verzweigtes Kanalsystem flächiger Hohlräume, das von Membranen umschlossen ist. Man findet das endoplasmatische Retikulum mit Ausnahme von ausgereiften Erythrozyten in allen eukaryotischen Zellen; je nach Zelltyp ist es unterschiedlich stark entwickelt.
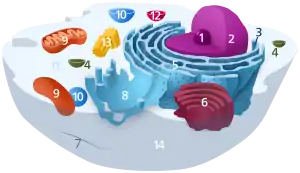
1. Nucleolus (Kernkörperchen)
2. Zellkern (Nukleus)
3. Ribosomen
4. Vesikel
5. Raues (Granuläres) ER (Ergastoplasma)
6. Golgi-Apparat
7. Cytoskelett
8. Glattes (Agranuläres) ER
9. Mitochondrien
10. Lysosom
11. Cytoplasma (mit Cytosol und Cytoskelett)
12. Peroxisomen
13. Zentriolen
14. Zellmembran
| Übergeordnet |
| Organell |
| Untergeordnet |
| ER-Membran ER-Lumen Ribosomen Proteinkomplexe |
| Gene Ontology |
|---|
| QuickGO |
Aufbau
Das endoplasmatische Retikulum besteht aus einem weit verzweigten Membrannetzwerk aus Röhren, Bläschen und Zisternen (sackähnlichen Strukturen), die von der ER-Membran umgeben werden. Die ER-Membran schließt das Innere des endoplasmatischen Retikulums, das ER-Lumen, vom Zytosol ab. Das Membranlabyrinth des endoplasmatischen Retikulums macht über die Hälfte der gesamten Membranmenge in einer Eukaryotenzelle aus.
Die ER-Membran geht direkt in die Kernhülle des Zellkerns über, das heißt Kernhülle und ER stellen ein morphologisches Kontinuum dar. Das ER-Lumen steht mit dem Membranzwischenraum der Kernhülle, dem perinukleären Raum, in Verbindung.
Teile des endoplasmatischen Retikulums, raues ER genannt, sind auf ihren Membranflächen mit Ribosomen besetzt; andere Bereiche sind glatt und ribosomenfrei und heißen daher glattes ER. Raues und glattes ER unterscheiden sich in ihrer Funktion.
Die Struktur des endoplasmatischen Retikulums ist dynamisch und einer stetigen Reorganisation unterworfen. Dazu gehören die Verlängerung oder auch Retraktion von Membrantubuli, ihre Verzweigung, Verschmelzung oder Aufspaltung. Diese Motilität des ER ist abhängig vom Cytoskelett. In Pflanzenzellen und Hefe spielt vor allem F-Aktin dabei eine wichtige Rolle. In tierischen Zellen dagegen erfolgt der Auf- und Umbau des ER unter dem dominierenden Einfluss der Mikrotubuli. Es wurde gezeigt, dass ein Vertreter der Aktin-assoziierten Motorproteine der Myosinfamilie Myosin V bei der Zellteilung für die Weitergabe des peripheren ER an die Tochterzellen verantwortlich ist.
Aufgaben und Typen
Am und im ER finden Translation, Proteinfaltung, Proteinqualitätskontrolle, posttranslationale Modifikationen von Proteinen und Proteintransport von Transmembranproteinen und sekretorischen Proteinen (siehe Exozytose) statt. Außerdem ist das ER der Ort, an dem (z. B. nach der Mitose) neue Kernmembranen gebildet und abgeschnürt werden. Auch dient das ER als intrazellulärer Calcium-Speicher, womit ihm eine Schlüsselrolle in der Signaltransduktion zukommt. In Muskelzellen (dort nennt man das ER Sarkoplasmatisches Retikulum, kurz SR) ist die Freisetzung von Calcium der Mediator einer Kontraktion. Die Aufgaben von rauem und glattem ER sowie SR sind unterschiedlich.
Glattes ER (agranuläres ER)
Das glatte ER (englisch smooth endoplasmic reticulum, sER) spielt eine wichtige Rolle bei mehreren Stoffwechselprozessen (= metabolischen Prozessen). Enzyme des glatten ERs sind für die Synthese verschiedener Lipide (vor allem Phospholipide, Fettsäuren und Steroide) von Bedeutung. Weiterhin spielt das glatte ER eine wichtige Rolle beim Kohlenhydratstoffwechsel, der Entgiftung der Zelle und der Speicherung von Calcium. Dementsprechend ist das in Parenchymzellen der Niere und der Leber aufgefundene ER vorwiegend glattes ER.
Hormonsynthese
Zu den im glatten ER gebildeten Steroiden gehören Steroidhormone, wie etwa diejenigen der Nebennierenrinde oder die Geschlechtshormone der Wirbeltiere. So enthalten in Hoden und Eierstöcken diejenigen Zellen, die für die Hormonproduktion zuständig sind, besonders viel glattes ER.
Kohlenhydratspeicherung
In den Parenchymzellen der Leber (= Hepatozyten) wird Glucose als ein Polymer namens Glykogen gespeichert. Die Freisetzung von Einzelmolekülen (= Monomeren) aus Glykogen heißt Glykogenolyse. Die Glykogenolyse in Leberzellen ist ein wichtiger Vorgang zur Steuerung der Glucosekonzentration im Blut (= des Blutzuckerspiegels). Im Wege der Glykogenolyse entsteht aber nicht Glucose selbst, sondern Glucose-6-phosphat. Das Enzym Glucose-6-Phosphatase auf der Membran des glatten ERs spaltet die Phosphatgruppe des Glucose-6-phosphats ab; (erst) die dadurch entstehende Glucose kann die Leberzelle verlassen und so den Blutzuckerspiegel erhöhen.
Entgiftung
Das glatte ER der Hepatozyten enthält auch membrangebundene Enzyme, die an der Biotransformation körperfremder Stoffe (z. B. Medikamente) beteiligt sind. Diese Enzyme (sog. CYPs) gehören der Cytochrom-P450-Klasse an und werden auch in einigen anderen Geweben außerhalb der Leber exprimiert. In der Leber haben CYPs eine Entgiftungsfunktion; sie können eine Vielzahl körperfremder Stoffe als Substrate erkennen. CYPs oxidieren meist ihre Substrate und fügen dadurch polare Gruppen in sie ein. Die entstehenden Stoffe sind besser wasserlöslich als die körperfremden Ausgangsstoffe und können daher von den Nieren besser ausgeschieden werden (= sind nierengängig).
Calcium-Speicher
Im Lumen des ER erreicht die Calcium2+-Konzentration millimolare Werte (ca. 10−3 M). Im Cytosol beträgt die Konzentration freier Calcium-Ionen in Ruhe dagegen nur etwa 100–150 nM (also etwa 10−7 M). Damit besteht über die Membran des ER ein Konzentrationsgradient von vier Größenordnungen. Sowohl die Aufnahme von Calcium in das ER als auch die Freisetzung von Calcium-Ionen aus dem ER unterliegt unter physiologischen Bedingungen einer feinen Regulation, die für die Aufrechterhaltung der Calcium-Homöostase außerordentlich wichtig ist.
Da Calcium-Ionen im Cytosol ein wichtiger „second messenger“ sind, spielt die regulierte Freisetzung von Calcium aus dem ER eine Schlüsselrolle für die intrazelluläre Signalgebung. Die Wirkungen einer durch Freisetzung aus dem ER erfolgten Erhöhung der intrazellulären Calciumkonzentration sind vielfältig:
- Enzyme werden aktiviert oder gehemmt,
- die Genexpression wird reguliert,
- in Neuronen wird die synaptische Plastizität beeinflusst,
- in der Muskulatur kontrahieren die Muskelfasern (Calcium-Ionen werden aus dem ER freigesetzt, das in Muskelzellen als sarkoplasmatisches Retikulum (SR) bezeichnet wird),
- Zellen des Immunsystems setzen Antikörper frei usw.
Calcium-Ionen verlassen das ER durch zwei Arten von Calciumkanälen: die IP3-Rezeptoren und die Ryanodin-Rezeptoren. Die Abkürzung IP3 steht für Inositoltrisphosphat, welches ebenfalls ein second messenger ist. IP3 entsteht als Spaltprodukt des Phosphatidylinositol-4,5-bisphosphats unter Vermittlung der Phospholipase C, die ihrerseits durch bestimmte G-Proteine (Gq) aktiviert wird. Das geschieht, wenn ein mit diesem G-Protein gekoppelter metabotroper Rezeptor in der Plasmamembran angeregt wird. – Das so erzeugte IP3 bindet an seinen spezifischen Rezeptor in der Membran des ER, woraufhin sich die Calcium-Konzentration im Zytoplasma durch den Ausstrom aus dem ER durch die Kanäle der IP3-Rezeptoren erhöht. Als Teil dieser Signalkette (metabotroper Rezeptor – G-Protein – Phospholipase C – IP3 – IP3-Rezeptor – Calcium-Freisetzung) kann Calcium auch als tertiärer Bote angesehen werden.
Die Ryanodin-Rezeptoren sind Calcium-sensitive Calciumkanäle. Sie sind also einerseits permeabel für Calcium und werden andererseits durch Calcium-Ionen aktiviert. Das geschieht, wenn im Zytoplasma die Calciumkonzentration ansteigt. Calcium-Ionen binden an die Ryanodin-Rezeptoren, diese öffnen sich, und Calcium-Ionen strömen durch sie aus dem ER in das Cytosol. Diesen Prozess nennt man „Calcium-induzierte Calciumfreisetzung“ (engl. „CICR“ – calcium-induced calcium release). Am bekanntesten ist die Rolle von CICR bei der Kontraktion der Herzmuskelzellen.
In der Membran des ER befinden sich Calcium-ATPasen vom SERCA-Typ. SERCA steht für sarkoendoplasmatisches-Retikulum-ATPase. Diese Proteine transportieren Calcium-Ionen aus dem Zytoplasma in das ER zurück. Da hierbei der oben erwähnte steile Konzentrationsgradient überwunden werden muss, kann dieser Transportvorgang nur unter ATP-Verbrauch stattfinden. Dieser Transportvorgang gehört somit zu den primär aktiven Transportvorgängen.
Membranproduktion
Das glatte ER lässt seine eigene Membran wachsen und dirigiert Membranbestandteile über Transportvesikeln zu anderen Teilen des inneren Membransystems, also dem Golgi-Apparat, Endosomen, Lysosomen und auch der Plasmamembran. Auch die Phospholipide werden durch das glatte ER hergestellt, indem Enzyme der ER-Membran sie aus Vorläufermolekülen, die sich im Zytosol und in der ER-Membran befinden, zusammensetzen.
Transitorische Funktion
Von einigen Abschnitten des glatten Endoplasmatischen Retikulums knospen sich Vesikel ab, die dann mit der cis-Seite des Golgi-Apparats fusionieren und so Proteine zur weiteren Bearbeitung (z. B. Anfügen von Zuckerseitenketten) überbringen. Dies ist ein Teil des so genannten "Sekretorischen Weges", da ihn vor allem Proteine betreffen, die später von der Zelle durch Exocytose in den Extrazellulären Raum ausgeschieden werden.
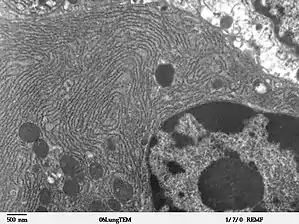
Raues ER (granuläres ER)
Das raue ER (rER), auch granuläres ER oder Ergastoplasma genannt, hat zwei Funktionen: die Proteinbiosynthese und die Membranproduktion. Seinen Namen hat es von den Ribosomen, die auf seinen Membranoberflächen sitzen. Es findet sich vorwiegend in den Zellen exokriner Drüsen und der Leber sowie in Nerven- (Nissl-Schollen) und Embryonalzellen. Das raue ER lässt sich mit basischen Farbstoffen wie Hämatoxylin, Kresylviolett oder Toluidinblau sichtbar machen (Nissl-Färbung).[1][2]
Proteinbiosynthese
Proteine werden häufig von spezialisierten Zellen ausgeschieden (Sekretion). Diese Proteine werden von den Ribosomen produziert, die dem rauen ER anhaften. Eines dieser Proteine ist zum Beispiel das Insulin aus Zellen der Bauchspeicheldrüse.
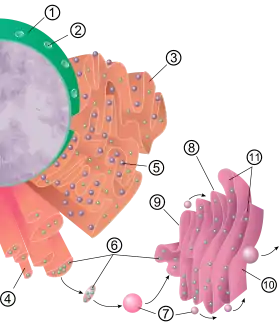
Alle in membranengebundenen Ribosomen entstehenden Polypeptidketten werden zunächst in das Lumen des ER geschleust. Dies geschieht durch porenbildende Proteine (Kotranslation). Auch im Zytosol synthetisierte Proteine werden in das Lumen des ER befördert (Posttranslation). Im Lumen des ER werden die Polypeptidketten zurechtgeschnitten und gefaltet.
Die linearen Aminosäureketten werden nach der Translokation in das ER gefaltet, erhalten also ihre dreidimensionale Struktur. Dieser Prozess wird von anderen Proteinen im ER unterstützt (Chaperone) und kontrolliert. Fehlgefaltete Proteine werden umgehend retranslokiert, das heißt zurück ins Zytosol transportiert und dort durch das Proteasom degradiert. Das Cholera-Bakterium nutzt diesen Mechanismus, um sein Toxin über diesen Prozess in das Zytosol zu bringen, wo es aber der Degradation durch das Proteasom entkommt und seine toxische Wirkung entfalten kann.
Die meisten Sekretionsproteine sind Glycoproteine, welche kovalent gebundene Kohlenhydrate tragen. Diese Kohlenhydrate, es handelt sich um Oligosaccharide, werden im Lumen des ER durch die Enzyme des ER angeheftet. Die fertigen sekretorischen Proteine verbleiben im Lumen des ER und werden somit von Proteinen im Zytosol, welche von freien Ribosomen erstellt wurden, ferngehalten. Die sekretorischen Proteine werden in Form kleiner Membranbläschen abgeschnürt und verlassen so das Lumen des ER als Transportvesikel in Richtung Golgi-Apparat.
1999 erhielt Günter Blobel den Nobelpreis für Physiologie oder Medizin für seine 1975 gemachte Entdeckung, dass Proteine durch endogene Protein-Signale (Signalsequenzen) vom ER aus in verschiedene Zellkompartimente zielgerichtet weitergeleitet werden. Als Signalsequenz in diesem Sinne wird eine bestimmte N-terminale Peptidsequenz bezeichnet, die nach dem Transport durch die Membran des ER durch die Signalpeptidase abgespalten wird. Proteine, die für Ziele außerhalb des ERs bestimmt sind, werden anschließend in Transportvesikel verpackt und entlang des Zytoskeletts zu ihrem Bestimmungsort weitergeleitet.
Sarkoplasmatisches Retikulum (Glattes Retikulum, SR)
Das glatte ER in Muskelzellen wird als sarkoplasmatisches Retikulum bezeichnet (SR). Das SR ist ein spezialisiertes ER der Muskelzellen. Es speichert Calciumionen. Diese werden beim Eintreffen eines elektrischen Impulses (Aktionspotential) in das Sarkoplasma (Cytoplasma der Muskelzellen) ausgeschüttet, diffundieren zwischen die Aktin- und Myosinfilamente der Muskelfibrillen und lösen das Ineinandergleiten der Filamente aus. Dadurch kommt es zur Kontraktion der Muskelfaser. Treffen keine weiteren Erregungen mehr an der Muskelfaser ein, werden die Calciumionen aktiv in das SR zurückgepumpt. Dadurch wird eine erneute Kontraktion verhindert. Das sarkoplasmatische Retikulum dient so der Regulation der Muskelkontraktion. Das Sarkoplasmatische Retikulum ist zudem Teil des longitudinal tubulären Systems.
Literatur
- Bruce Alberts u. a.: Molecular Biology of the Cell. 4. Auflage. Garland Science, New York 2002, ISBN 0-8153-4072-9.
- Neil A. Campbell u. a.: Biologie. 1. Aufl., 1. korrigierter Nachdr. Spektrum, Heidelberg 1997, ISBN 3-8274-0032-5.
Weblinks
- Endoplasmic Reticulum (Animation über die Strukturveränderung des ER)
- Raues ER: elektronenmikroskopische Bilder (Dr. Jastrows EM-Atlas)
- Glattes ER: elektronenmikroskopische Bilder (Dr. Jastrows EM-Atlas)
Einzelnachweise
- Wissenschaft-Online-Lexika: Eintrag zu Ergastoplasma im Lexikon der Biologie, abgerufen am 26. November 2011.
- Ergastoplasma im Roche-Medizinlexikon, 5. Auflage.