Adenosintriphosphat
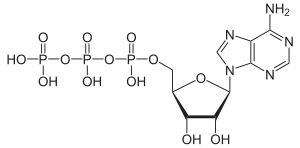
Adenosintriphosphat, kurz ATP, ist ein Nukleotid, nämlich das Triphosphat des Nucleosids Adenosin.
| Strukturformel | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||||||||
| Allgemeines | ||||||||||||||||||||||
| Name | Adenosintriphosphat | |||||||||||||||||||||
| Andere Namen | ||||||||||||||||||||||
| Summenformel | C10H16N5O13P3 | |||||||||||||||||||||
| Kurzbeschreibung |
farbloser Feststoff[2] | |||||||||||||||||||||
| Externe Identifikatoren/Datenbanken | ||||||||||||||||||||||
| ||||||||||||||||||||||
| Eigenschaften | ||||||||||||||||||||||
| Molare Masse | 507,18 g·mol−1 | |||||||||||||||||||||
| Aggregatzustand |
fest | |||||||||||||||||||||
| Sicherheitshinweise | ||||||||||||||||||||||
| ||||||||||||||||||||||
| Soweit möglich und gebräuchlich, werden SI-Einheiten verwendet. Wenn nicht anders vermerkt, gelten die angegebenen Daten bei Standardbedingungen. | ||||||||||||||||||||||
Adenosintriphosphat ist der universelle und unmittelbar verfügbare Energieträger in Zellen und wichtiger Regulator energieliefernder Prozesse. Das Molekül des Adenosintriphosphats besteht aus einem Adeninrest, dem Zucker Ribose und drei Phosphaten (α bis γ) in Ester- (α) bzw. Anhydridbindung (β und γ).[4]
Geschichte
Adenosintriphosphat wurde 1929 von dem deutschen Biochemiker Karl Lohmann entdeckt.[5] Eine chemische Synthese von ATP wurde erstmals 1949 von James Baddiley und Alexander Robertus Todd veröffentlicht. Die Rolle als Hauptenergiequelle in Zellen wurde 1939 bis 1941 von Fritz Lipmann aufgeklärt[6], nachdem schon Wladimir Alexandrowitsch Engelhardt 1935 gezeigt hatte, dass ATP für Muskelkontraktionen notwendig ist, und Herman Moritz Kalckar 1937 den Zusammenhang der Zellatmung mit der Biosynthese von ATP nachgewiesen hatte.[7] Untereinheiten der dafür zuständigen ATP-Synthase wurden erstmals von Efraim Racker ab 1960 isoliert.[8]
Energieträger
Auch Prozesse in Zellen benötigen Energie, um chemische Arbeit wie Synthese organischer Moleküle, osmotische Arbeit wie aktiven Stofftransport durch Biomembranen und mechanische Arbeit wie bei der Muskelkontraktion zu leisten. Als Überträger von Energie wird vornehmlich ATP genutzt. Die Phosphatreste dieses Nukleosidtriphosphats sind über Phosphoranhydrid-Bindungen (Säureanhydrid-Bindungen) miteinander verbunden. Durch enzymkatalysierte Hydrolyse können eine oder zwei Phosphatgruppen abgespalten werden und es entsteht Adenosindiphosphat (ADP) und Monophosphat oder Adenosinmonophosphat (AMP) und Pyrophosphat. Bei der Spaltung der Phosphatbindungen wird unter Standardbedingungen jeweils 32,3 kJ/mol bei Spaltung einer Bindung oder 64,6 kJ/mol bei Spaltung beider Bindungen für Arbeitsleistungen nutzbar.
Signalmolekül
Intrazellulär
ATP ist ein Cosubstrat der Kinasen, einer Gruppe von phosphatübertragenden Enzymen, die im Metabolismus und bei der Stoffwechselregulation eine Schlüsselrolle spielen. Bedeutende Mitglieder der letzteren Gruppe sind die Proteinkinasen, die je nach ihrem Aktivierungsmechanismus als Proteinkinase A (PKA, cAMP-abhängig), Proteinkinase C (PKC, Calcium-abhängig), Calmodulin-abhängige Kinase, oder Insulin-stimulierte Proteinkinase (ISPK) bezeichnet werden, um nur einige Beispiele zu nennen. Unter Blutzucker werden einige Grundprinzipien angesprochen, nach denen eine Serie von Kinasen zu einer Enzymkaskade zusammengeschaltet sein kann.
Extrazellulär
ATP (wie auch ADP und Adenosin) ist Agonist purinerger Rezeptoren, die sowohl im zentralen als auch im peripheren Nervensystem eine Rolle spielen. Somit ist es beteiligt an Prozessen wie der Durchblutungsregulation oder der Vermittlung von Entzündungsreaktionen. Es wird nach neuronalen Verletzungen ausgeschüttet und kann die Proliferation von Astrozyten und Neuronen stimulieren.
Regeneration
Aus dem bei der Energieabgabe aus ATP entstandenen AMP bzw. ADP regeneriert die Zelle das ATP. Dafür gibt es zwei verschiedene Wege, die als Substratkettenphosphorylierung und Elektronentransportphosphorylierung (Atmungskette) bezeichnet werden.
Bei der Substratkettenphosphorylierung wird ein Phosphatrest an ein Zwischenprodukt des Abbaus von stofflichen Energiequellen gebunden und nach weiterem Umbau des Zwischenprodukts auf ADP übertragen.
Bei der Elektronentransportphosphorylierung werden durch einen Transport von Elektronen entlang eines Redoxgradienten über verschiedene Elektronen- und Wasserstoff-Überträger in einer Membran Protonen von einem durch die Membran umschlossenen Raum der Zelle in einen anderen transportiert. In Bakterien werden so Protonen nach außen gepumpt. In Eukaryoten finden diese Prozesse in den Mitochondrien statt. Dort werden aus der Matrix des Mitochondriums Protonen in den Intermembranraum exportiert. In beiden Fällen wird ein Protonengradient erzeugt und als chemiosmotisches Potenzial ΔP genutzt, das sich aus einem Protonenkonzentrationsunterschied ΔpH und einer elektrischen Potentialdifferenz ΔΨ zusammensetzt. Der Rückfluss der Protonen durch das ebenfalls in der Membran lokalisierte Enzym ATP-Synthase treibt die von diesem Enzym katalysierte energieverbrauchende Bindung anorganischer Phosphatreste an das ADP an. In manchen Organismen werden anstatt Protonen Natriumionen verwendet, sie verfügen analog über eine Na+-abhängige ATP-Synthase.
Bei chemotrophen Organismen werden die Elektronen in Form der Reduktionsmittel NADH, NADPH, FADH2 oder reduziertes Ferredoxin in die Atmungskette eingespeist. Diese stammen aus dem oxidativen Abbau energiereicher Verbindungen, wie beispielsweise Kohlenhydraten oder Fettsäuren. Die Elektronen werden bei aeroben Organismen auf Sauerstoff übertragen, dabei entsteht Wasser. In der anaeroben Atmung werden andere Elektronenakzeptoren verwendet, beispielsweise Schwefel oder Eisen(II). In beiden Fällen entsteht eine elektrochemische Differenz, die zur ATP-Bildung genutzt wird. Bei Eukaryoten findet der Vorgang in den Mitochondrien, bei Prokaryoten im Cytoplasma statt.
Bei phototrophen Organismen werden nach Absorption von Licht durch Chlorophylle von diesen Elektronen auf einem hohen Energieniveau abgegeben. Die Lichtenergie wird damit genutzt, um eine elektrochemische Differenz zu erzeugen. Bei grünen Pflanzen findet dies in den Chloroplasten, bei Bakterien im Cytoplasma statt. Wegen der Nutzung des Lichts spricht man in diesem Fall von Photophosphorylierung.
Kurzzeitregeneration in Muskelzellen
Da die oxidative Phosphorylierung in der Atmungskette ein relativ langsamer Prozess ist, muss der ATP-Vorrat in stark beanspruchten Zellen (Muskelzellen) auch kurzfristig wieder aufgefüllt werden. Der ATP-Vorrat (in der Muskelzelle ca. 6 mmol/kg Muskel) reicht bei maximaler Kontraktion nur etwa 2–3 Sekunden. Eine Reserve stellen hier Moleküle mit höherem Gruppenübertragungspotenzial als ATP dar. Säugetiermuskelzellen halten einen Vorrat an Kreatinphosphat (21 mmol/kg Muskel; 0,08 % pro Körpergewicht[9]) bereit. Die Creatin-Kinase katalysiert die Übertragung der Phosphorylgruppe vom Kreatinphosphat an das ADP. Ist dieser Vorrat nach 6–10 Sekunden verbraucht, müssen die oben genannten Mechanismen die ATP-Regeneration allein tragen.
Energiebereitstellung in Muskelzellen
Während starker Muskelbeanspruchung bauen Muskelzellen Glucose zu Lactat in der Milchsäuregärung ab, um schnell ATP zu erzeugen. Lactat selbst wird in der Leber wieder zu Pyruvat und dann zu Glucose unter ATP-Verbrauch aufgebaut (Gluconeogenese). Diese Glucose wird dann wieder dem Muskel als Energiequelle zur Verfügung gestellt. Dieser Kreislauf wird auch als Cori-Zyklus bezeichnet.
Im Notfall werden zur Energieerzeugung auch körpereigene Proteine abgebaut. Proteine werden in Aminosäuren zerlegt, und diese meistens zu Pyruvat abgebaut. In einem dem Cori-Zyklus ähnlichen Weg wird Pyruvat zunächst zu Alanin transaminiert und zur Leber transportiert. Dort kehren sich diese Schritte um und die Leber erzeugt aus Pyruvat wieder Glucose, die dem Muskel bereitgestellt wird. Dieser Zyklus wird auch als Glucose-Alanin-Zyklus bezeichnet.
Energiebereitstellung im Herzmuskel
Der Herzmuskel nutzt Fettsäuren als Brennstoff, diese werden in der β-Oxidation in den zahlreichen Mitochondrien abgebaut. Des Weiteren können auch Glucose, Lactat (über Reoxidation zu Pyruvat), Ketonkörper und Glykogen abgebaut werden. Bei hoher Belastung können bis zu 60 % der Energie aus der Oxidation von Lactat gewonnen werden.
Konzentrationen
In der Zelle ist die ATP-Konzentration eine Regelgröße: Das Absinken unter 4–5 mmol/l aktiviert energieliefernde Reaktionen (siehe Phosphofructokinase); das Übersteigen des Schwellenwertes bewirkt Energiespeicherung, z. B. durch Bildung von Kreatinphosphat als schnell verfügbaren (ATP-liefernden) Speicher im Muskel oder Aufbau von Glykogen als „Energiepolster“ in der Leber. Kohlenhydrat- und Proteinspeicher sind allerdings limitiert. Weiterer Energieüberschuss führt (über Acetyl-CoA) zur Speicherung von Fett.
Umsatz
Bei einem durchschnittlichen Erwachsenen entspricht die Menge ATP, die täglich in seinem Körper auf- und abgebaut wird, etwa seiner halben Körpermasse. So setzt ein 80 kg schwerer Mann etwa 40 kg ATP am Tag um, was etwa 78,8 mol oder 1025 Molekülen entspricht, die wieder neu gebildet werden. Bei intensiver körperlicher Arbeit kann der ATP-Umsatz auf 0,5 kg pro Minute ansteigen.
Siehe auch
Literatur
- Reginald H. Garrett, Charles M. Grisham: Biochemistry. 4th edition, international edition. Brooks/Cole, Cengage Learning Services, Boston MA u. a. 2009, ISBN 978-0-495-11464-2.
Einzelnachweise
- Eintrag zu ADENOSINE TRIPHOSPHATE in der CosIng-Datenbank der EU-Kommission, abgerufen am 27. März 2020.
- Eintrag zu Adenosin-5′-triphosphat. In: Römpp Online. Georg Thieme Verlag, abgerufen am 30. Mai 2014.
- Datenblatt Adenosintriphosphat bei Sigma-Aldrich, abgerufen am 12. Juni 2011 (PDF).
- J. R. Knowles: Enzyme-catalyzed phosphoryl transfer reactions. In: Annual Review of Biochemistry. Band 49, 1980, ISSN 0066-4154, S. 877–919, doi:10.1146/annurev.bi.49.070180.004305, PMID 6250450.
- Über die Pyrophosphatfraktion im Muskel. In: Naturwissenschaften. Band 17, Nr. 31, 1. August 1929, ISSN 0028-1042, S. 624–625, doi:10.1007/BF01506215 (springer.com [abgerufen am 25. April 2018]).
- Geschichte des ATP auf der Nobel-Webseite.
- H. M. Kalckar: Phosphorylation in Kidney Tissue. In: Enzymologia. 2, 1937, S. 47–53.
- M. E. Pullman, H. S. Penefsky, A. Datta, E. Racker: Partial resolution of the enzymes catalyzing oxidative phosphorylation. I. Purification and properties of soluble dinitrophenol-stimulated adenosine triphosphatase. In: The Journal of Biological Chemistry. Band 235, November 1960, ISSN 0021-9258, S. 3322–3329, PMID 13738472.
- Reginald H. Garrett, Charles M. Grisham: Biochemistry. 4th edition, international edition. Brooks/Cole, Cengage Learning Services, Boston MA u. a. 2009, ISBN 978-0-495-11464-2, S. 849.