Nashörner
Die Nashörner (Rhinocerotidae) oder auch Rhinozerosse bilden eine Familie der Unpaarhufer (Perissodactyla) mit heute noch fünf lebenden Arten. Sie sind charakterisiert durch einen kräftigen Körper und kurze Gliedmaßen mit drei Zehen sowie einen großen Kopf, der bei allen heute lebenden Vertretern eine markante Bildung, bestehend aus einem oder zwei – für die Familie namensgebenden – Hörnern, trägt. Die Familie stellt eine der vielfältigsten und erfolgreichsten in der Geschichte der Säugetiere dar und war während ihrer vor nahezu 50 Millionen Jahren beginnenden Entwicklungsgeschichte über weite Teile Eurasiens, Afrikas und Nordamerikas verbreitet. Ihr Niedergang begann Ende des Miozäns vor rund 6 bis 5 Millionen Jahren in Verbindung mit klimatischen und damit einhergehenden Landschaftsveränderungen, die zum Aussterben der nordamerikanischen sowie zahlreicher weiterer Nashornvertreter im ursprünglichen Verbreitungsgebiet führten. Gegen Ende des Pleistozäns gab es eine erneute Aussterbephase, während der alle nordeurasischen Vertreter verschwanden. Überlebt haben die heute noch bestehenden Nashornarten Breitmaul- und Spitzmaulnashorn in Afrika südlich der Sahara sowie Panzer-, Java- und Sumatra-Nashorn im süd- bis südöstlichen Asien, die teilweise in ihrem Bestand durch Vernichtung ihres Lebensraumes und Wilderei abermals stark geschrumpft sind.
| Nashörner | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Breitmaulnashörner in Namibia | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Rhinocerotidae | ||||||||||||
| Owen, 1845 |
Merkmale



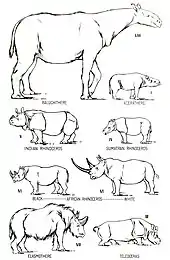
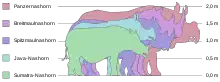
Nashörner sind große bis sehr große Säugetiere. Sie weisen bei den heute lebenden Vertretern eine Kopf-Rumpf-Länge zwischen 2,5 und 3,8 m auf (zuzüglich eines 40 bis 60 cm langen Schwanzes) bei Schulterhöhen variierend von 1,2 bis 1,8 m und einem Körpergewicht schwankend zwischen 500 und 3.600 kg. Die größte heute lebende Nashornart ist das Breitmaulnashorn (Ceratotherium simum). Fossil traten aber noch deutlich größere Arten auf. So erreichten einige Vertreter von Elasmotherium und Brachypotherium jeweils mehr als 5.000 kg Körpergewicht.[1]
Allgemein haben Nashörner einen massigen Körper mit großem Kopf und kurzen, kräftigen Beinen. Jeder Fuß hat drei Zehen, die jeweils in breiten Hufen enden. Die Haut ist dick und grau oder braun gefärbt. Bei den asiatischen Arten ist die Haut am Ansatz des Halses und der Beine stark gefaltet, so dass es aussieht, als seien die Tiere gepanzert. Die meisten heutigen Nashörner sind unbehaart mit Ausnahmen der Ohrspitzen und Schwanzenden. Allerdings weist das Sumatra-Nashorn (Dicerorhinus sumatrensis) noch teilweise ein recht dichtes Haarkleid auf. Beim fossilen Wollnashorn (Coelodonta antiquitatis) ist ein sehr dichtes Fell durch im Permafrost Sibiriens erhaltene Kadaver nachgewiesen,[2] für andere ausgestorbene Nashornarten wird es teils angenommen. Darüber hinaus haben Nashörner ein geringes Sehvermögen, doch wird dieser Nachteil durch einen ausgeprägten Geruchssinn und ein sehr gutes Gehör ausgeglichen.[3]
Schädel- und Gebissmerkmale
Nashörner haben einen sehr großen Schädel, der gewöhnlich recht lang gestreckt und am Hinterhauptsbein aufgerichtet ist. Das Nasenbein ist häufig massiv und deutlich nach vorn gewölbt und ragt über den Zwischenkieferknochen. An den Stellen, wo die Hörner ansetzen, befinden sich auf den Knochenoberflächen deutliche, meist perl- oder blumenkohlförmig aufgeraute Strukturen. Die Gehirnkapsel ist verhältnismäßig klein.[4]
Das Gebiss ist sehr unterschiedlich aufgebaut, bei den heutigen Nashörnern aber generell reduziert. Bei den afrikanischen Nashörnern fehlt das gesamte vordere Gebiss bestehend aus Schneidezähnen und Eckzahn. Die asiatischen Nashornarten hingegen besitzen noch ein oder zwei Paare an Schneidezähnen je untere und obere Gebisshälfte. Hierbei haben die innersten oberen Schneidezähne (I1) eine meißelartige Gestalt, die jeweils zweiten unteren (i2) erinnern an Hauer oder kleine Stoßzähne. Dies wird als „Meißel-Stoßzahn“-Komplex bezeichnet. Er findet sich auch bei zahlreichen ausgestorbenen Vertretern und dient zur Unterscheidung der Nashörner von ihren stammesgeschichtlich nächsten Verwandten. Die Molaren weisen wie bei allen Unpaarhufern zwei aus Zahnschmelz gebildete Querleisten auf der Kaufläche (bilophodont) auf. Als generelles Kennzeichen verläuft der Zahnschmelz auf den Kauflächen der vorderen beiden Oberkiefermolare charakteristisch „π“-förmig, während dieser auf den Unterkiefermolaren „L“-förmig gestaltet ist. Dieses Merkmal gilt auch für alle fossilen Nashörner und für deren unmittelbar nächsten Verwandten, so dass dieses sich schon vor etwa 50 Millionen Jahren herausgebildet hatte.[5] Abhängig von der Ernährungsweise können die hinteren Backenzähne nieder- oder hochkronig (brachyodont oder hypsodont) ausgebildet sein. Die Prämolaren sind bei den rezenten Arten weitgehend molarisiert, das heißt, sie unterscheiden sich nur wenig von den Molaren. Je nach stammesgeschichtlicher Entwicklung gibt es jedoch unterschiedliche Grade der Molarisierung der Prämolaren bei den einzelnen fossilen Arten.[4][6] Urtümliche Nashörner besaßen noch die vollständige Gebissformel bestehend aus drei Schneidezähnen, einem Eckzahn, vier Prämolaren und drei Molaren je Kieferast; das am stärksten reduzierte Gebiss wurde bei Elasmotherium mit nur zwei Prämolaren und drei Molaren je Kieferast nachgewiesen. Zusätzlich wiesen die Backenzähne dieser Nashorngattung die höchsten Zahnkronen innerhalb der gesamten Ordnung der Unpaarhufer auf, und sie werden in diesem Merkmal nur von einigen Vertretern der Nagetiere übertroffen.[7] Die Reduktion der Zahnanzahl begann sehr früh in der Stammesgeschichte der Nashörner. In der Regel bildete sich zuerst der untere Eckzahn zurück, gefolgt vom äußersten unteren und oberen Schneidezahn sowie vom oberen Eckzahn.[8]
Hörner


Ein wesentliches, optisch markantes und namensgebendes Merkmal der Nashörner sind die Hörner. Bei den rezenten Vertretern gibt es je nach Art ein Horn oder zwei Hörner. So besitzen das asiatische Panzer- (Rhinoceros unicornis) und das Java-Nashorn (Rhinoceros sondaicus) nur ein vorderes Horn (Nasalhorn), welches dem Nasenbein entwächst. Die afrikanischen Nashornarten Breitmaul- und Spitzmaulnashorn (Diceros bicornis) und das asiatische Sumatra-Nashorn haben dagegen zwei Hörner, wobei das Nasalhorn ebenfalls auf der Nase sitzt, das hintere (Frontalhorn) aber auf dem Stirnbein wächst. Bei ausgestorbenen Nashornarten können die Hörner meist nur anhand der Ansatzstellen am Schädel nachgewiesen werden. So trugen zum Beispiel Vertreter der Gattung Elasmotherium nur ein Horn auf der Stirn, während jene von Diceratherium zwei Nasalhörner hatten und ebensolche von Aceratherium wohl gar keine besaßen.[9]
Das Horn besteht aus agglutiniertem Keratin, einem fibrillären Protein, das auch in Haaren vorkommt, und enthält trotz seiner Festigkeit weder Knochensubstanz noch, wie stellenweise irrtümlich behauptet wird, Elfenbein. Es setzt sich zusammen aus zahlreichen langen fadenförmigen Strängen, Hornsäulchen oder Filamente genannt, deren Zwischenräume mit Hornsubstanz verfestigt sind. Diese Fäden verlaufen durch die gesamte Hornlänge, verjüngen sich aber nach oben hin deutlich. Der Kern des Hornes ist deutlich fester ausgebildet und meist schwarz gefärbt, nach außen hin wird es deutlich faseriger und nimmt eine hellgraue Farbe an.[10][11] Fossiles Horn ist bisher nur vom Wollnashorn überliefert,[12] weist aber prinzipiell den gleichen Aufbau auf wie rezentes.[13] Das Horn nutzt sich mit der Zeit durch Reiben am Untergrund oder an Steinen kontinuierlich ab, beim Kampf mit Artgenossen oder infolge traumatischer Erlebnisse kann es auch abbrechen, wächst aber das gesamte Leben lang nach. Das bisher größte bekannte Horn hat 1,58 m über die vordere Krümmung gemessen.[14]
Innere Organe
Wie alle Unpaarhufer sind Nashörner gute Enddarmfermentierer, so dass die Verdauung weitgehend im Darm erfolgt. Der Magen erreicht eine Länge von 120 cm. Der Blinddarm ist zwischen 60 und 90 cm lang, der Grimmdarm 5 bis 8 m, der gesamte Darmtrakt kann bis über 20 m lang werden. Zum Abbau unverdaulicher Pflanzenteile dienen im hintersten Teil des Darms Mikroorganismen. Das Herz wiegt bis zu 5 kg.[3][15][16] Die männlichen Tiere besitzen keinen Hodensack; die Hoden liegen im Leibesinneren.[17]
Verbreitung und Lebensraum
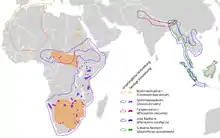
Nashörner leben heute in Afrika südlich der Sahara und in Süd- bzw. Südostasien sowohl in Savannenlandschaften als auch in Tropischen Regenwäldern in Hoch- oder Tiefländern. Die ursprüngliche Verbreitung war aber wesentlich weiter. Die stammesgeschichtlich ältesten Arten lassen sich im Mittleren Eozän vor rund 50 Millionen Jahren in Eurasien und Nordamerika nachweisen. Afrika erreichten die Nashörner erstmals mit der Schließung des Tethys-Ozeans und der Schaffung einer Landbrücke im frühen Miozän vor etwa 20 Millionen Jahren.[1][9] Am Ende des Miozäns und zum Übergang zum Pliozän vor 4 Millionen Jahren starben die Nashörner aufgrund von Klimaänderungen in Nordamerika aus.[18]
Während des Pleistozäns mit seinem stark schwankenden Klima waren Nashörner auch im hohen Norden Eurasiens verbreitet und lebten in subarktischen Tundrenlandschaften. Am Ende der letzten Eiszeit verschwanden die Nashörner aus Nordasien und Europa.[9][19] Im Laufe ihrer Stammesgeschichte hatten die unterschiedlichen Nashornarten nahezu alle für große, terrestrisch lebende Säugetiere erreichbaren ökologischen Nischen besetzt.[1]
Lebensweise
Sozialverhalten, Fortpflanzung und Entwicklung

Nashörner leben häufig als Einzelgänger, die aber in Savannen auch in kleinen, matriarchalisch organisierten Herden auftreten können. Bullen sind meistens Einzelgänger und leben territorial. Die einzelnen Tiere bewohnen dabei eng definierte Reviere, die mit Urin und Kot markiert werden, ebenso wie die häufig begangenen Wege. Am Tage schlafen Nashörner oder halten sich an Suhlplätzen auf, aktiv fressend sieht man sie in der Dämmerung und nachts. Sie sind scheue Tiere, die menschliche Nähe meiden. Berichte über die Angriffslust der Tiere sind in der Regel stark übertrieben. Die Angriffe sind, wenn es dazu kommt, kaum zielgerichtet, können aber durch die Hörner, die vorderen Zähne sowie die Kraft und Masse des Tieres mit tödlichen Verletzungen enden. Ein Nashorn läuft überdies bis zu 45 km/h (12,5 m/s) schnell und übertrifft damit knapp menschliche Spitzensportler. Dabei kann es abrupt die Richtung wechseln. Zudem sind bei Kämpfen zwei unterschiedliche Strategien zu beobachten. Die asiatischen Nashornarten, die alle über eine vordere Bezahnung verfügen, nutzen ihre meist kleinen Hörner nur selten bei Kämpfen, die dann hochritualisiert in Horngefechten ausgetragen werden. Als eigentliche Waffen dienen die dolchartigen Schneidezähne des Unterkiefers, mit denen sie gefährliche und tiefe Wunden reißen. Die afrikanischen Nashornarten, die kein Vordergebiss aufweisen, setzen ihre häufig wesentlich längeren Hörner – vor allem das Nasalhorn – neben Drohgebärden auch aktiv als Waffe zur Selbst-, Revier-, aber auch zur Futterverteidigung ein, um damit den Gegner mittels Aufspießens zu schwächen.[3][20]
Während der Brunftzeit einer Kuh kann es zu Kämpfen unter den Bullen kommen, wobei der Sieger in auffallender Weise um das weibliche Tier wirbt. Dies erfolgt durch gegenseitiges Jagen oder Scheingefechte, anschließend kommt es zur Kopulation. Nach einer Tragzeit von 15 bis 18 Monaten wird ein Junges geboren, das zweieinhalb bis drei Jahre bei der Mutter bleiben kann. Kommt ein zweites Jungtier zur Welt, so wird das ältere zumindest für die Zeit des Säugens von der Mutter verjagt. Die Lebenserwartung heutiger Nashörner liegt zwischen 30 und maximal 50 Jahren.[3][20] Die Altersbestimmung fossiler Arten erfolgt überwiegend im Vergleich zum Breitmaulnashorn und beruht auf anatomischen Merkmalen wie Zahndurchbruch, Abkauungsgrad der einzelnen Zähne oder Verwachsungsstadien bestimmter Knochennähte. Allgemein hängt die Lebenserwartung bei Säugetieren eng mit dem Körpergewicht der erwachsenen Tiere zusammen,[21] bei ausgestorbenen Nashörnern, die ähnlich groß waren wie die heutigen, lag sie wahrscheinlich in einem gleichwertigen Rahmen. Genauere Analysen zu einzelnen fossilen Populationen liegen aber nur selten vor, für pleistozäne Vertreter betrifft dies weitgehend das Wollnashorn (Coelodonta antiquitatis),[22][23] bei älteren Formen sind solche für die eher häufig auftretenden Gattungen wie Teleoceras[24] oder Chilotherium[25] aus dem Miozän bekannt.
Die heute lebenden, weitgehend offene Landschaften bewohnenden Nashornarten, wie die afrikanischen Nashörner und das Panzernashorn, werden oft von Vögeln wie Madenhackern oder Kuhreihern begleitet, die auf ihrer Haut sitzen und sie von Parasiten reinigen. Die Alarmrufe der Madenhacker warnen die Tiere außerdem vor Gefahren oder Störungen, einschließlich des Menschen, in Distanzen von 27 bis 61 m. Dabei vermag ein Tier bei höherer Anzahl an Madenhackern die potentielle Störung in größerer Entfernung zu orten, die Rufe geben aber keine Auskunft über die Richtung. Dies ermitteln die Nashörner über den Geruchssinn.[26] Bei den überwiegend im tropischen Regenwald lebenden Java- und Sumatra-Nashorn wurde dies bisher nicht beobachtet, was mit ihrer Lebensweise in geschlossenen Landschaftsgebieten zusammenhängt. Jungtiere können in einzelnen Fällen von Großkatzen und Hyänen, gelegentlich auch von Wildhunden erbeutet werden, ausgewachsene Nashörner haben außer dem Menschen keine natürlichen Feinde.[20] Auch ein Großteil der fossilen Vertreter hatte eher selten Konfrontationen mit Beutegreifern zu fürchten, gelegentlich treten aber Bisswunden bei heute ausgestorbenen Vertretern auf, die teilweise von sehr großen Raubtieren verursacht wurden.[27][28]
Ernährung

Alle Nashörner ernähren sich ausschließlich von Pflanzenkost und sind an diese Ernährungsweise mit breiten Backenzähnen angepasst. Dabei haben sich die Arten aber auf unterschiedliche Pflanzennahrung spezialisiert. Vier der fünf heute lebenden Nashornarten bevorzugen weiche Pflanzenkost, wie Blätter, Äste, Zweige, Knospen und Früchte (browsing). Die Backenzähne dieser Arten weisen meist niedrige Zahnkronen und weniger Zahnzement auf. Außerdem besitzen diese Nashornarten aufgrund der hohen Kopfhaltung ein eher rechtwinkliges kurzes Hinterhauptsbein. Das Panzernashorn (Rhinoceros unicornis) ernährt sich aber auch zum Teil von Gräsern, besitzt aber wie die anderen, Blattnahrung zu sich nehmenden Nashornarten eine spitze, bewegliche Oberlippe. Das Breitmaulnashorn ist die einzige rezente, vollständig an Grasnahrung angepasste Nashornart (grazing). Da Gräser Kieselsäure enthalten, die sehr hart ist, haben sich bei ihm aufgrund des hohen Abriebs beim Kauen Zähne mit hohen Zahnkronen und einem hohen Zahnzementanteil ausgebildet. Die dauerhafte niedrige Kopfhaltung während der Nahrungsaufnahme führte evolutiv zur Verlängerung des Hinterhauptes und damit zu einer tiefen Kopfhaltung und Ausbildung eines Nackenbuckels. Des Weiteren besitzt das Breitmaulnashorn namengebende breite wulstige Lippen.[3][29][30]
Ein Großteil der fossilen Nashörner war aufgrund ihrer Anatomie auf weiche Pflanzennahrung spezialisiert. Dennoch kam es immer wieder während der Stammesgeschichte infolge von Klima- und damit verbundenen Landschaftsveränderungen innerhalb der verschiedenen Nashornlinien unabhängig voneinander (konvergente Evolution) zur Spezialisierung auf Grasnahrung. Bei den moderneren Nashörnern der Gruppe der Rhinocerotinae war dies mit Veränderungen der Schädelanatomie, unter anderem mit der Verlängerung des Hinterhauptsbeines und einem daraus resultierenden, tief hängenden Kopf, verbunden. Dies geschah vor allem gegen Ende des Miozäns und während des Plio- und des Pleistozäns. Neben dem heute lebenden Breitmaulnashorn sind die bekanntesten fossilen grasfressenden Vertreter das Wollnashorn, das Steppennashorn (Stephanorhinus hemitoechus) und die verschiedenen Mitglieder der Gattung Elasmotherium. Zumindest beim Wollnashorn ist durch die fossil erhaltenen Eismumien eine dem Breitmaulnashorn ähnlich geformte Maulpartie nachgewiesen.[2][4][9] Ein weiteres Anpassungsmodell an Grasnahrung ist bei stammesgeschichtlich älteren Linien, wie den Aceratheriinae feststellbar. Hier erfolgte eine Kürzung der Gliedmaßen, die eine niedrige Körper- und Kopflage verursachte. Diese Nashornvertreter konnten mit nur einer leichten Senkung des Kopfes den Boden erreichen, eine Verlängerung des Hinterhauptsbeines fand nicht statt, so dass der Schädel überwiegend horizontal getragen wurde. Bekannte Gattungen hierbei sind Chilotherium und Teleoceras aus dem späten Miozän. Beide Entwicklungsmodelle zeichnen sich aber durch die Erhöhung der Zahnkronen aus, um einem verstärkten Abrieb durch die harte Grasnahrung entgegenzuwirken.[28][31]
Systematik
Äußere Systematik
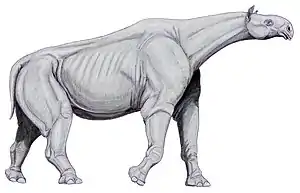
Kladogramm der Nashornverwandtschaft nach Bai et al. 2020[32]
|
Die nächsten heute lebenden Verwandten der Nashörner sind die Pferde und Tapire. Die Trennung von den Tapiren erfolgte dabei vor etwa 47 bis 57 Millionen Jahren, die Pferde hatten sich bereits vor 56 bis 60 Millionen Jahren abgespalten.[33][34][35] Dabei bilden die Nashörner einen Teil der Überfamilie Rhinocerotoidea (Nashornartige). Zu dieser gehören in einer klassischen Auffassung zwei weitere, ausgestorbene Familien: die Amynodontidae und die Hyracodontidae, wobei letztere zusammen mit den Nashörnern eine enger verwandte Gruppe bilden, die sich durch eine im Vergleich zum Schädel längere Backenzahnreihe und in einzelnen Zahnmerkmalen von den Amynodontidae abheben.[36] Unterschiede finden sich vor allem im vorderen Gebissaufbau. So weisen die Nashörner eine typische „Meißel-Stoßzahn“-Bildung der oberen und unteren Schneidezähne auf, die Amynodontidae haben vergrößerte Eckzähne und kleine Schneidezähne, während die Hyracodontidae ein vielfältigeres Muster zeigen.[37] Die Amynodontidae bestanden vom Mittleren Eozän bis zum Mittleren Miozän und traten in Eurasien und Nordamerika auf. Vertreter dieser Nashornartigen waren teilweise so groß wie heutige Nashörner, trugen aber kein Horn. Sie lebten halb-aquatisch und ernährten sich wahrscheinlich von Wasserpflanzen. Die Hyracodontidae, welche ebenfalls vom Mittleren Eozän bis zum Unteren Miozän vorkamen, zeigen in dieser Definition eine hohe Variabilität, die sich in drei bestehenden Unterfamilien ausdrückt. Dazu gehören die relativ kleinen Hyracodontinae, die Allaceropinae und die großen Indricotheriinae. Letztere stellten mit Paraceratherium (auch unter den Synonymen Baluchitherium und Indricotheriumbekannt) das größte bekannte Landsäugetier der Erdgeschichte. Es war langhalsig und hornlos und kam hauptsächlich im Oligozän in Asien vor.[38] Neuere Untersuchungen fassen die Hyracodontidae als nicht einheitliche Gruppe auf. Sie werden daher in die Hyracodontidae im engeren Sinne, in die Eggysodontidae (etwa den Allaceropinae entsprechend) und in die Indricotheriidae (= Paraceratheriidae) aufgespalten. Letztere beiden stehen den Nashörnern mit ihrem spezialisierteren Vordergebiss deutlich näher als die klassischen Hyracodontidae mit ihren eher generalisierten Schneide- und Eckzähnen.[39][40] Allerdings bilden vermutlich auch die Indricotheriidae keine in sich geschlossene Gruppe, so dass in einer Auffassung aus dem Jahr 2020 diese in die riesenhaften eigentlichen Indricotheriidae und die urtümlicheren und kleineren Forsterccoperiidae aufgeteilt werden müssen. Letztere sind sehr basal einzuordnen, erstere formen die Schwestergruppe der eigentlichen Nashörner.[32]
Innere Systematik
|
Die eigentlichen Nashörner (Rhinocerotidae) entwickelten mehrere Linien: Die Unterfamilie der Diceratheriinae umfasst mit den Diceratheriini und den Trigoniadini zwei Triben. Sie waren charakterisiert durch paarige Hörner auf der Nase und stellten die ersten Nashörner mit derartigen Bildungen dar. Sie lebten weitgehend im Oligozän und Miozän hauptsächlich in Nordamerika, wo sie bis vor 21 Millionen Jahren die größte endemische Säugetiergruppe darstellten.[18] In Eurasien sind Fossilien dieser Nashorngruppe eher selten überliefert. Eine weitere Unterfamilie bilden die Aceratheriinae, die nur selten Hörner besaßen und in die Triben Aceratheriini und Teleoceratini aufgegliedert werden. Ihre Entwicklung begann ebenfalls bereits im Oligozän, sie überlebten aber teilweise bis zum Ende des Pliozäns und besiedelten sowohl Eurasien und Nordamerika als auch Afrika.[36][41]
Die Unterfamilie der Rhinocerotinae splitterte sich ebenfalls in mehrere Linien auf. Die Tribus der Menoceratini umfasst dabei die Stammgruppe, wurde aber ursprünglich aufgrund der paarigen Nasenhörner in die Nähe der Diceratheriinae gestellt.[38] Sie weisen aber wesentlich modernere Entwicklungsmerkmale als diese auf.[41] Die Elasmotheriini entwickelten sich im Unteren Miozän,[43] wobei die bekannteste Gattung, das elefantengroße Elasmotherium, noch in der letzten Kaltzeit (Weichsel-Kaltzeit) lebte und sich durch ein überdimensionales, möglicherweise bis zu 2 m langes Horn auszeichnete.[7]
Verwandtschaftsverhältnis der Nashörner basierend auf genetischen Daten nach Liu et al. 2021[44]
|
Die fünf heute lebenden Arten gehören alle zur Tribus der Rhinocerotini. Ihre nächsten Verwandten bilden wahrscheinlich die Elasmotheriini. Zwischen den beiden Linien liegt laut molekulargenetischen Untersuchungen eine tiefe zeitliche Kluft, da ihre Trennung bereits im Paläogen begann, die Angaben reichen je nach Studie vom Mittleren Eozän vor rund 47 Millionen Jahren bis zum Unteren Oligozän vor etwa 31 Millionen Jahren.[35][45][44] Die Rhinocerotini kann man in drei Gruppen einteilen, welche die zweihörnigen (Dicerorhinina) und einhörnigen Formen (Rhinocerotina) Asiens und die (generell) zweihörnigen (Dicerotina) Afrikas umfassen. Ihre Verwandtschaftsverhältnisse zueinander werden vielfältig diskutiert. Als zwei gegenständige Ansichten haben sich die „Horn-Hypothese“ (die zweihörnigen Nashörner sind gegenüber den einhörnigen enger miteinander verwandt) und die „geographische Hypothese“ (die asiatischen Nashörner und die afrikanischen bilden jeweils eigene Verwandtschaftsgemeinschaften) herausgestellt. Als dritte Möglichkeit kommt eine Schwestergruppenbeziehung des zweihörnigen Sumatra-Nashorns (Dicerorhinus sumatrensis) zu allen anderen rezenten Nashörnern in Betracht.[44]
Eine Studie aus dem Jahr 2021 sieht die geographische Variante bestätigt. Ihren Ergebnissen zufolge spalteten sich die heutigen zwei asiatischen und eine afrikanische Linie im Mittleren Miozän vor rund 15,6 Millionen Jahren voneinander ab. Als Ursache hierfür wird die Ausbildung von Landverbindungen zwischen den afrikanischen und der eurasischen Kontinentalmassen im Unteren Miozän diskutiert. Die asiatische Linie wiederum teilte sich nur wenig später, vor etwa 14,8 Millionen Jahren, in die Dicerorhinina und die Rhinocerotina auf. Das hochgradig gefährdete Sumatra-Nashorn stellt hierbei die einzige überlebende Art der Dicerorhinina dar. Diesen werden allerdings auch die während des Pleistozäns über weite Teile des nördlichen Eurasiens verbreiteten Nashorngattungen Coelodonta mit dem bekannten Wollnashorn und Stephanorhinus, zu der das weniger bekannte Waldnashorn gehört, zugeordnet.[46] Ihre Diversifizierung setzte im Oberen Miozän vor rund 9,4 Millionen Jahren ein. Die Rhinocerotina, die mit dem gefährdeten Panzernashorn (Rhinoceros unicornis) und dem hochgradig gefährdeten Java-Nashorn (Rhinoceros sondaicus) zwei Arten einschließen, vollzogen ihre Aufspaltung erst im Unteren Pliozän vor gut 4,3 Millionen Jahren. Auf dem afrikanischen Kontinent hatten sich derweil bereits im ausgehenden Miozän vor etwa 6,8 Millionen Jahren die Linien des Breitmaulnashorns (Ceratotherium simum) und des Spitzmaulnashorns (Diceros bicornis) als Angehörige der Dicerotina genetisch voneinander entfernt.[44] Andere molekulargenetische Untersuchungen erbrachten dagegen weitaus ältere Radiationsdaten. Demnach begann die Aufspaltung der rezenten Nashörner bereits im Unteren Oligozän vor etwa 29 bis 30 Millionen Jahren. Hierbei formten sich zuerst die Dicerorhinina heraus mit einer Abspaltung der Coelodonta-Stephanorhinus-Linie vor gut 21 Millionen Jahren im Unteren Miozän.[47] Die afrikanischen Dicerotina wiederum bildeten vor rund 17 Millionen Jahren die heutigen Linien aus, während sich jene der asiatischen Rhinocerotina vor etwa 11,7 Millionen Jahren herausdifferenzierten.[33][48]
Alternative innere Systematik der Nashörner nach Cerdeño 1995[49]
|
Alternative innere Systematik der Nashörner nach Antoine 2003[50]
|
Die hier dargestellte Nashorn-Systematik basiert auf den Ausarbeitungen von Kurt Heissig,[4][41] Donald R. Prothero[38] und Colin Peter Groves.[51] Dabei ist die systematische Stellung der lebenden Nashornarten als Tribus oder Untertribus in Diskussion,[4] allerdings scheint die Stellung als Untertribus gerechtfertigt zu sein.[42] Problematisch ist des Weiteren die Stellung von Dicerorhinus. das manchmal auch der Untertribus Rhinocerotina[4] oder keiner speziellen Untertribus zugewiesen wird.[33]
Daneben gibt es noch weitere Ordnungsschemata der Nashörner wie jenes von Pierre-Olivier Antoine, in dem die zwei Unterfamilien Elasmotheriinae und Rhinocerotinae unterschieden werden. Dabei umfasst erstere die Triben Elasmotheriini und Diceratheriini, während Rhinocerotini und Aceratheriini zu den Rhinocerothinae gestellt werden.[50] Ebenfalls ein zusätzliches Ordnungsschema stammt von Esperanza Cerdeño, welches die Unterfamilien Acertheriinae mit den Triben Alicornopini bzw. Teleoceratini und die Unterfamilie Rhinocerothinae mit der alleinigen Tribus Rhinocerotini herausstellt.[49]
Übersicht über die rezenten und fossilen Nashorn-Taxa
Einschließlich der rezenten Vertreter sind bis heute über 70 Gattungen mit mehreren hundert Arten beschrieben worden.[52] Das hier verwendete Ordnungsschema folgt weitgehend Prothero und Schoch 1989,[42] wobei Änderungen der Gliederung von Heissig 2007[41] und weitere Überarbeitungen und Ergänzungen aus neuerer Zeit berücksichtigt wurden:[50][53][54][55][56][57][58][59][60][61][62]
- Familie: Rhinocerotidae Owen, 1845.
- Meschotherium Gabunia, 1964
- Woodoceras Prothero, 2005
- Gulfoceras Albright, 1999
- Molassitherium Becker & Antoine, 2013
- Unterfamilie: Diceratheriinae Dollo, 1885
- Tribus: Trigoniadini Heissig, 1989
- Ronzotherium Aymard, 1854
- Guixia You, 1977
- Epiaceratherium Abel, 1910
- Penetrigonias Tanner & Martin, 1976
- Trigonias Lucas, 1900
- Amphicaenopus Wood, 1927
- Tribus: Diceratheriini Dollo, 1885
- Subhyracodon (= Caenopus, Leptaceratherium) Brandt, 1878
- Diceratherium Marsh, 1875
- Skinneroceras Prothero, 2005
- Unterfamilie: Aceratheriinae Dollo, 1885
- Persiatherium Pandolfi, 2016
- Tribus: Aceratheriini Dollo, 1885
- Mesaceratherium Heissig, 1969
- Proaceratherium Ginsburg & Hugueney, 1980
- Floridaceras Wood, 1964
- Galushaceras Prothero, 2005
- Aphelops Cope, 1873
- Peraceras Cope, 1880
- Aceratherium Kaup, 1832
- Hoploaceratherium Ginsburg & Heissig, 1989
- Dromoaceratherium Crusafont & Villalta, 1955
- Plesiaceratherium Young, 1937
- Alicornops Ginsburg & Guérin, 1979
- Chilotherium (= Subchilotherium) Ringström, 1924
- Chilotheridium Hooijer, 1971
- Shansirhinus Kretzoi, 1942
- Acerorhinus Kretzoi, 1942
- Sinorhinus Schlosser, 1903
- Tribus: Teleoceratini Hay, 1902
- Diaceratherium Dietrich, 1931
- Brachydiceratherium Lavocat, 1951
- Prosantorhinus Heissig, 1973
- Teleoceras Hatcher, 1894
- Brachypotherium Roger, 1904
- Turkanatherium Dereniyagala, 1951
- Aprotodon Forster-Cooper, 1915
- Symphyssorrachis Beliajeva, 1954
- Brachypodella Heissig, 1973
- Unterfamilie: Rhinocerotinae Owen, 1845
- Tribus: Menoceratini Prothero, Manning & Hanson, 1986
- Menoceras (= Moschoedestes) Troxell, 1921
- Pleuroceros Roger, 1898
- Protaceratherium Abel, 1910
- Tribus: Elasmotheriini Bonaparte, 1845
- Bugtirhinus Antoine & Welcomme, 2000
- Kenyatherium Aguirre und Guérin, 1974
- Hispanotherium Crusafont & Villalta, 1947
- Victoriaceros Geraads, MacCrossin & Benefit, 2012
- Samburuceros Handa, Nakatsukasa, Kunimatsu & Nakaya, 2017
- Kenyatherium Aguirre & Guérin, 1974
- Ougandatherium Guérin & Pickford, 2003
- Caementodon Heissig, 1972
- Procoelodonta Matthew, 1931
- Gobitherium Kretzoi, 1943
- Beliajevina Heissig, 1974
- Bergertherium Beliajeva, 1971
- Huaqingtherium Huang & Yan, 1983
- Iranotherium Ringström, 1924
- Eoazara Geraads & Zouhri, 2021
- Parelasmotherium Killgus, 1923
- Ningxiatherium Chen, 1977
- Sinotherium Ringström, 1922
- Elasmotherium Fischer, 1808
- Tribus: Rhinocerotini Owen, 1845
- Untertribus: Dicerorhinina Ringström, 1924
- Dicerorhinus (= Didermocerus, Ceratorhinus) Gloger, 1841 (einschließlich Sumatra-Nashorn)
- Rusingaceros Geraads, 2010
- Pliorhinus Pandolfi, Pierre-Olivier, Bukhsianidze, Lordkipanidze & Rook, 2021
- Stephanorhinus (= Brandtorhinus) Kretzoi, 1942
- Lartetotherium Ginsburg, 1974
- Dihoplus Brandt, 1878
- Coelodonta (= Tichorhinus) Bronn, 1831
- Untertribus: Rhinocerotina Ringström, 1924
- Iberotherium Antunes & Ginsburg, 2000
- Gaindatherium Colbert, 1934
- Punjabitherium Khan, 1971
- Nesorhinus Antoine, Reyes, Amano, Bautista, Chang, Claude, Vos & Ingicco, 2021
- Rhinoceros Linnaeus, 1758 (einschließlich Panzernashorn und Java-Nashorn)
- Untertribus: Dicerotina Ringström, 1924
- Paradiceros Hooijer, 1968
- Diceros Gray, 1821 (einschließlich Spitzmaulnashorn)
- Ceratotherium Gray, 1867 (einschließlich Breitmaulnashorn)
Formen mit unklarerer Zugehörigkeit zu den Nashörnern:[32]
- Uintaceras Hollbrook & Lucas, 1997
- Teletaceras Hanson, 1989
Stammesgeschichte
Adaptive Radiation
Die Familie der Nashörner gehört zu den erfolgreichsten und vielfältigsten Säugetiergruppen der jüngeren Erdgeschichte und lässt sich rund 50 Millionen Jahre zurückverfolgen. Während dieser Zeit vollzog sich eine breit angelegte evolutionäre Entwicklung und Anpassung an die jeweiligen Lebensräume (Adaptive Radiation). Vertreter der Nashörner besetzten nahezu jedes terrestrische Biotop.[18] Innerhalb der Stammesgeschichte gab es zahlreiche anatomische Veränderungen. Allgemeine Evolutionstrends bei den Nashörnern, die weitgehend alle Linien durchliefen und jeweils Anpassungen an bestimmte Biotope darstellen, sind die Kürzung und Verbreiterung des Schädels, vor allem im vorderen Gesichtsbereich, Längenreduktion der Gliedmaßen, Reduktion des Gebisses, Molarisierung der Prämolaren und Vergrößerung der Höhe der Zahnkronen bei den Backenzähne. Weiterhin können zwei generelle Entwicklungsmodelle festgestellt werden: hornlose Nashörner mit einem weitgehend intakten oder umfangreichen vorderen Gebiss mit vergrößerten Zähnen und horntragende Nashörner mit einer stark reduzierten oder nicht vorhandenen vorderen Bezahnung.[4]
Eozän

Der früheste bekannte Vorläufer der Nashörner war Hyrachyus aus dem Mittleren Eozän Nordamerikas und Europas. Er war etwa so groß wie ein heutiger Schäferhund und besaß die vollständige Säugetierbezahnung mit kaum molarisierten Prämolaren. Aufgrund der sehr basalen Stellung wird diese Gattung je nach Auffassung zu den Tapiroiden oder zu den Rhinocerotoiden gestellt.[63] Mögliche Frühformen der Nashörner sind mit Uintaceras und Teletaceras ebenfalls im Mittleren oder Oberen Eozän nachgewiesen, besaßen aber wie alle urtümlichen Vertreter keine Hornbildung. Beide Formen waren ebenfalls relativ klein. Uintaceras ist weitgehend nur aus Nordamerika belegt, Teletaceras hingegen kam weitverbreitet vor und ist unter anderem aus der Clarno-Formation in Nordamerika sowie aus der Pondaung-Formation in Südostasien überliefert.[64][65] Im Oberen Eozän traten dann auch die ersten echten und schon größeren Nashörner auf, wie etwa das rindergroße Trigonias.[1][18]
Oligozän
Die ersten Nashörner Europas erschienen mit dem großen und hornlosen Ronzotherium erstmals im Unteren Oligozän, das von zahlreichen Fundstellen belegt ist.[66] Ebenso trat Epiaceratherium auf, möglicherweise kam die Form aber schon im Oberen Eozän vor. Es hatte vergleichbar zu Ronzotherium eine weitgehend westeurasische Verbreitung. Einzelne Funde wie ein Schädel aus der Fossillagerstätte von Na Dương in Vietnam zeigen jedoch ein weites Vorkommen bis in den östlichen Teil des Großkontinentes an.[67] Wie Ronzotherium gehört Epiaceratherium noch der Stammgruppe der Nashörner an. In Asien breiteten sich des Weiteren Vertreter von Guixia aus. Im Oberen Oligozän teilten sich Europa und Asien zahlreiche gemeinsame Nashornlinien. So entwickelten sich mit Protaceratherium ein Mitglied der Aceratheriini, die charakterisiert waren durch eine lange, rüsselartige Schnauze und eine zurückgebildete Nasenregion und auf pflanzliche Weichkost spezialisiert waren. Ebenso entstanden mit Diaceratherium (nicht zu verwechseln mit Diceratherium) und Brachydiceratherium erste Vertreter der flusspferdartigen kurzbeinigen und teils in offenen Waldlandschaften, später auch in Savannen lebenden Teleoceratini, die teilweise spezialisierte Grasfresser waren und sehr hochkronige Backenzähne entwickelten.[1][18]
In Nordamerika bildeten sich eigene Nashornlinien aus, insgesamt war aber die Diversität der dortigen Nashörner verglichen mit Eurasien wesentlich geringer. Subhyracodon, das ebenfalls schon im Oberen Eozän auftrat, entwickelte sich zur Diceratherium-Linie weiter. Diese stellten die ersten Nashörner mit Hornbildungen dar, die bei dieser Gruppe aber paarig auf der Nase saßen.[18] Mit dem Auftreten von Subhyracodon und Diceratherium wurden zudem in Nordamerika zum Ende des Oligozäns alle urtümlichen Nashörner mit vierzehigen durch modernere mit dreizehigen Vorderfüßen ersetzt, ein Prozess der in Eurasien und Afrika noch bis weit in das Miozän hinein andauerte.[68]
Miozän

Im Miozän kam es zu einer starken Radiation der Nashörner mit einem großen Formenreichtum.[9][19] Im Unteren Miozän betraten die Nashörner erstmals afrikanischen Boden. Frühe Funde sind mit Aceratherium und Brachypotherium in Ägypten und Libyen nachgewiesen.[1] Vertreter der Aceratheriini erreichten außerdem Nordamerika und bildeten unter anderem mit Floridaceras und Aphelops eigene Gattungen aus.[36] Auch die Teleoceratini wanderten nach Nordamerika aus. Hier findet sich in Nebraska am Verdigre Creek in der Ash-Hollow-Formation, wo eine komplette Herde mit intakten Mageninhalten unter Vulkanasche begraben wurde, einer der besten Nachweise der für diese Nashorngruppe namengebenden Gattung Teleoceras, der zudem einer der dominantesten Pflanzenfresser jener Zeit in Nordamerika war.[69] Weiterhin erschien das aus Eurasien stammende Menoceras, das wie Diceratherium paarige Nasalhörner besaß aber ein basaler Vertreter der Rhinocerotinae ist.
Aber auch nach Eurasien wanderten verschiedene, ursprünglich nordamerikanische Nashörner ein, wie einzelne Vertreter der Diceratheriini. Bedeutend ist hier jedoch die Ausbreitung der hornlosen Nashörner, der Aceratheriini, die zahlreiche Formen hervorbrachten und die neben dem bereits erwähnten Aceratherium auch Alicornops und Chilotherium einschließen. Vor allem Letzteres war sehr häufig. Weiterhin können schon im Unteren Miozän erstmals Angehörige modernerer Nashornformen, wie Bugtirhinus als Basalform der Elasmotheriini, nachgewiesen werden.[70] Bedeutend ist darüber hinaus das erste Auftreten der heute noch bestehenden Gattungen, wie der afrikanischen Formen Ceratotherium und Diceros im Oberen Miozän.[71] Dabei muss vor allem das Verbreitungsgebiet von Diceros. ursprünglich wesentlich größer gewesen sein, da die stammesgeschichtlich alte Art Diceros gansuensis. vor 5 bis 7 Millionen auch in Ostasien auftrat.[72] Weiterhin sind in Eurasien mit Dicerorhinus frühe Vertreter der Dicerorhinina nachgewiesen,[1] während Sinotherium und Ningxiatherium deutlich entwickeltere Angehörige der Elasmotheriini darstellen.[43] Die Rhinocerotina sind in dieser Zeitphase mit Gaindatherium und Punjabitherium nachweisbar.[1]
Am Ende des Miozäns kam es aufgrund klimatischer Abkühlung verbunden mit der Ausbreitung von offenen Steppenlandschaften zu einem Aussterben zahlreicher Nashornarten. Dies betraf vor allem die Aceratheriini und einen Teil der Teleoceratini. In Nordamerika verschwand die gesamte Nashornfauna, in Eurasien überlebten nur Elasmotheriini, Dicerorhinina und Rinocerotina. In Afrika hielten sich des Weiteren die Dicerotina und einige Arten von Brachypotherium aus der Gruppe der Teleoceratini.[9]
Plio- und Pleistozän
_-_Mauricio_Ant%C3%B3n.jpg.webp)
Die Zeit des Plio- und Pleistozäns war vor allem geprägt durch die Weiterentwicklung der modernen Nashorngruppen, wie Elasmotheriini und Rhinocerotini. Letztere schließen die noch heute bestehenden Arten ein. Vor allem die urtümlichen Dicerorhinina Eurasiens zeigten eine große Diversität. Allerdings ist von den direkten Vorfahren des Sumatra-Nashornes wenig bekannt. Bedeutend ist der Aufstieg des seit dem mittleren Pliozän bekannten Coelodonta zum am Ende des Pleistozäns lebenden Wollnashorn.[73] Die Schwestergruppe Stephanorhinus spaltete sich in mehrere, an unterschiedliche Biotope angepasste Arten auf, wie es die mittelpleistozänen Formen des Wald- und Steppennashorns zeigen. Beide Nashornlinien enden jedoch im späten Pleistozän.[74] Die Gattung Rhinoceros ist seit dem Pliozän nachweisbar und spaltete sich in mehrere Formen auf. Auf den Philippinen kam im Mittleren Pleistozän das nahe verwandte Nesorhinus vor, eine der wenigen Formen der Nashörner mit Inselverzwergung.[60] Im nördlichen Eurasien, vor allem in Zentralasien sind die verschiedenen Vertreter von Elasmotherium überliefert. Diese Nashorngattung stirbt allerdings ebenfalls im Jungpleistozän aus.[1] In Afrika entwickeln sich Diceros und Ceratotherium zu den heute bekannten Arten.[71][55]
Am Ende des Pleistozäns kam es zu einer erneuten Aussterbewelle, in deren Zuge die Nashornlinien des nördlichen Eurasiens verschwanden (siehe Quartäre Aussterbewelle). Nur die heute noch existierenden Nashornvertreter überlebten.
Taxonomie

Der Name Rhinoceros wurde 1758 von Linnaeus als wissenschaftliche Bezeichnung für das Nashorn eingeführt, genutzt wurde der Name aber schon mindestens seit dem Beginn des 16. Jahrhunderts, so unter anderem von Albrecht Dürer bei seinem berühmten Holzschnitt Rhinocerus, den er 1515 anfertigte.[75] Einer der frühesten Hinweise auf die Namensverwendung stammt jedoch mit rhinókerôn aus der Bibliothéke historiké des antiken Historikers Diodor von Agyrion aus dem 1. Jahrhundert v. Chr.[76] Die Bezeichnung setzt sich aus den griechischen Wörtern ῥίς (rhīs „Nase“; Genitiv rhinos) und κέρας (kéras „Horn“) zusammen, bezieht sich also eindeutig auf die Hornbildung auf der Nase.[77] Linnaeus unterschied die damals bekannten zwei Arten: R. unicornis, das Panzernashorn, welches er in Indien und Afrika verortete, und R. bicornis, das Spitzmaulnashorn, welches seiner Meinung nach aber in Indien lebte.[78] Linnaeus' Irrtümer bezüglich der Verbreitung beruhen wahrscheinlich auf der Verwendung häufig älterer Quellen bei der Erstellung seines Werkes Systema Naturae. Zahlreiche Gelehrte seiner Zeit lehnten eine Einteilung der Nashörner in zwei Arten ab und bevorzugten stattdessen nur eine, die bestimmte Variationen aufwies. Der Widerspruch löste sich erst auf, als der niederländische Anatom Petrus Camper 1771 ein zweihörniges Nashorn aus Südafrika studierte und neben dem zweiten Horn auch Unterschiede im Gebissaufbau bemerkte; die vollständige Publikation seiner Ergebnisse erfolgte aber erst 1782.[75] Ungefähr im gleichen Zeitraum, 1769, entdeckte der deutsche Naturforscher Peter Simon Pallas ein ebenfalls zweihörniges Nashorn in Sibirien, das Wollnashorn.[79]
Der britische Naturforscher John Edward Gray führte 1821 die Familienbezeichnung Rhynocerotidae ein. Seine kurze Beschreibung lautete: „Nose short, rounded, bones very thick, bearing a horn formed of agglutinated hair; toes three to each foot; stomach simple; intestine and caecum large“ („Nase kurz, gerundet, Knochen sehr dick, horntragend, bestehend aus zusammengewachsenem Haar; drei Zehen an jedem Fuß; Magen einfach; Darm und Blinddarm groß“).[80] Aufgrund der falschen Schreibweise wird diese Bezeichnung aber offiziell nicht anerkannt, allerdings unterschied Gray in seiner Beschreibung formal die beiden heute noch bestehenden Gattungen Rhinoceros (Panzernashorn) und Diceros (Spitzmaulnashorn). Bereits 1811 hatte der deutsche Zoologe Johann Karl Wilhelm Illiger den Namen Nasicornia für die Nashörner vorgeschlagen, der, da er nicht auf einem vergebenen Gattungsnamen beruhte, ebenfalls ungültig ist.[81] Im Jahr 1845 legte Richard Owen, neben Charles Darwin einer der bedeutendsten Naturhistoriker des Viktorianischen Zeitalters, eine Studie vor, in der er die Familie der Nashörner mit einbezog und auch die für die Säugetiertaxonomie wichtigen Zähne berücksichtigte. Hier bezeichnete er die Nashörner korrekt als Rhinocerotidae, weswegen viele Experten die Gruppenbenennung „Rhinocerotidae Owen, 1845“ gegenüber der eigentlich nicht korrekten Benennung „Rhinocerotidae Gray, 1821“ bevorzugen.[82]
Nashörner und der Mensch
Nashörner in Kunst und Kultur
.jpg.webp)
Nashörner gehören, bezogen auf ihre Körpergröße und ihren Habitus, zu den beeindruckendsten landlebenden Säugetieren und fanden aufgrund dessen auch Einzug in die Kunst und Kultur des Menschen, besonders häufig trifft dies dabei auf Jäger-und-Sammler-Populationen zu. Die frühesten bekannten Darstellungen von Nashörnern finden sich im westeurasischen Jungpaläolithikum (vor 40.000 bis 10.000 Jahren) und sind wenigstens 31.000 Jahre alt. Hervorzuheben sind hier die Malereien der frankokantabrischen Höhlenkunst, wo Nashörner in mehr als einem halben Dutzend Höhlen mit über 80 Darstellungen abgebildet wurden. Am häufigsten sind sie in der Grotte Chauvet (Frankreich) mit 65, teils in roten oder schwarzen Farbpigmenten gehaltenen Zeichnungen überliefert, allein ein Panneau enthält 17 Darstellungen eines wohl in Bewegung befindlichen Tieres. Des Weiteren finden sich in Europa auch außerhalb dieses Kulturkreises in der mobilen jungpaläolithischen Kleinkunst Abbildungen und Darstellungen von Nashörnern in Form von Knochen- oder Steinritzungen, aber auch als kleine Statuetten modellierten Tonfiguren, die zu den ältesten Keramikgegenständen der Welt gehören. Alle diese Kunsterzeugnisse werden heute als Darstellungen des Wollnashorns interpretiert, welches damals in den nordeurasischen Kältesteppen lebte, einige ältere auch manchmal als Steppennashorn.[83][84] Einige Forscher vertreten die Meinung, einzelne charakteristische Nashorndarstellungen, wie zum Beispiel in der Höhle Rouffignac, könnten als Abbildungen von Elasmotherium angesehen werden,[85] doch war diese Gattung weder räumlich noch zeitlich so weit verbreitet.[86]
Auch vorgeschichtliche Jäger- und Sammlergemeinschaften in anderen Erdteilen bildeten Nashörner ab. In Südasien existieren zahlreiche Malereien in Höhlen und Abris, die in diesem Falle das Panzernashorn wiedergeben. Diese werden dem dortigen Mesolithikum zugewiesen und sind zwischen 12.000 und 7.000 Jahre alt. Bedeutend und zu den ältesten in dieser Region gehören jene vom Marodeo-Felsen nahe Pachmarhi im indischen Bundesstaat Madhya Pradesh oder eine Jagdszene, dargestellt bei Mirzapur im indischen Bundesstaat Uttar Pradesh.[87][88] Aus weiten Bereichen des südlichen Afrika sind zahlreiche Felszeichnungen und -gravierungen von sowohl Breitmaul- als auch Spitzmaulnashorn bekannt, die überwiegend den Khoisan-Bevölkerungsgruppen zugewiesen werden und bis zu 14.000 Jahre alt sein können, teilweise aber auch bedeutend jünger sind. Diese sind mitunter sehr zahlreich, wobei gebietsweise mehr Gravierungen als Zeichnungen zu beobachten sind. Allein in den südafrikanischen Provinzen Nordkap, Nordwest und Freistaat sind mehr als 500 Darstellungen überliefert.[89][90]

Mit dem Sesshaftwerden des Menschen gehen die Abbildungen von Nashörnern rapide zurück. Aus der Kupferzeit sind Darstellungen der Tiere auf Siegeln der Indus-Kultur bekannt,[91] vereinzelt finden sie sich auch als Reliefdarstellungen im Alten Ägypten.[92] Bemerkenswert sind dabei auch Relieffiguren einhörniger Nashörner an Tempelfriesen in Angkor Vat (Kambodscha), die aus dem 12. Jahrhundert stammen und aufgrund des historischen Verbreitungsgebietes zu den wenigen Abbildungen des Java-Nashornes gehören.[93] Eine hohe kulturelle Bedeutung für das südliche Afrika hat das „Goldene Nashorn“ aus dem „Originalen Goldgrab“ vom Mapungubwe-Hügel in der südafrikanischen Provinz Limpopo. Der archäologische Fundstätten-Komplex wurde von lokalen Stämmen im 11. bis 13. Jahrhundert angelegt und in den 1920er bis 1930er Jahren ausgegraben. Das kleine Nashornfigürchen von 5,5 cm Höhe bildete möglicherweise gemeinsam mit anderen zoomorphen Statuetten wie einem Büffel, einer Raubkatze, einem Elefanten und einem Krokodil ein Zierelement eines größeren Gefäßes, das vermutlich beim Wahrsagen Verwendung fand.[94][95][96] Für die europäische Kunstgeschichte herausragend sind der weit verbreitete Holzschnitt Rhinocerus von Albrecht Dürer aus dem Jahr 1515, entstanden nach nur indirekten Berichten über ein indisches Panzernashorn in Portugal, und das Ölgemälde Rhinoceros Clara von Jean-Baptiste Oudry aus dem Jahr 1749.[97]
Bedrohung und Schutz
| Art | Bestandsgröße 2007[98] | Bestandsgröße 2012[99] | Bestandsgröße 2020[100] |
|---|---|---|---|
| Breitmaulnashorn | 14.500 | 20.400 | 17.212–18.915 |
| Spitzmaulnashorn | 3.725 | 5.050 | 5.366–5.627 |
| Panzernashorn | 2.619 | 3.300 | >3.600 |
| Sumatra-Nashorn | 275 | <100 | <80 |
| Java-Nashorn | 55 | 35–44 | 70–74 |

Die heute lebenden Nashörner gehören aufgrund mehrerer Faktoren – enorme Größe, langsames Wachstum, territoriale Einzelgänger, wenige Nachkommen in menschlicher Obhut – zu den nicht domestizierbaren Wildtieren.[101] Dadurch ist die Bedeutung der Tiergruppe für den heutigen Menschen als Nahrungs- und Rohstofflieferant relativ gering, in Asien besteht jedoch eine große Nachfrage nach Hörnern der Nashörner. Diese werden im Nahen Osten, vor allem im nördlichen Jemen, traditionell für die Griffe des Jambia-Dolches verwendet, der als Statussymbol dient.[102] In Ostasien dagegen sind sie Bestandteil kunsthandwerklicher Schnitzereien und der Traditionellen Chinesischen Medizin.[103] Bei Letzterer dienen die Hörner vor allem in pulverisierter Form als Medikament gegen Fieber und Schmerzen, ein häufig angenommener Einsatz als Aphrodisiakum ist historisch nicht verbürgt. Vor allem aufgrund dieses Marktes sind Nashörner durch die damit verbundene Wilderei vom Aussterben bedroht. So wurden auf dem Schwarzmarkt im Jahr 2012 zwischen 30.000 und 65.000 US-Dollar für jedes Kilogramm Horn gehandelt, im Zeitraum von 1993 bis 2009 betrug der Wert noch rund 4700 bis 5000 US-Dollar je Kilogramm.[104] Durch die hohe Nachfrage aus Ostasien an Horn stieg in den letzten Jahren nicht nur die Anzahl der durch Wilderei getöteten freilebenden Nashörner vor allem im südlichen Afrika,[105] sondern es kam auch verstärkt zu Diebstählen in Museen, Sammlungen und Auktionshäusern und damit verbundenem Schmuggel.[106][107] Mehrere durchgeführte Tests seitens der Pharmaindustrie und biowissenschaftlicher Forschungsinstitute ergaben jedoch keinerlei medizinische Wirkung.[108][109] Jedoch gab es immer wieder Gerüchte über Wunderheilungen, so dass zuletzt 2009 die Nachfrage stark angestiegen war.[110] Weitere Ursachen für die Bedrohung der heutigen Nashornarten sind darüber hinaus die Zerstörung der Lebensräume durch Landwirtschaft oder Bau von Verkehrswegen aber auch die Ausdehnung der menschlichen Siedlungen bis an die Grenzen der Schutzgebiete.[3]
In neuerer Zeit gibt es allerdings wieder eine leichte Bestandserholung einiger Arten. In Afrika lebten Ende 2010 wieder rund 20.000 Breitmaul- und über 5.000 Spitzmaulnashörner. Beide Bestände haben sich seit 1995 fast verdoppelt.[111][112] Ein großer Anteil der Nashörner lebt davon in Südafrika zumeist in eingezäunten und bewachten Reservaten. In Kenia finden sich ebenfalls verschiedene private und öffentliche Schutzgebiete, z. B. die private Farm Ol ari Nyiro von Kuki Gallmann oder der Lake-Nakuru-Nationalpark. Zwar hat sich der Gesamtbestand der afrikanischen Nashörner insgesamt laut IUCN stabilisiert, doch sind einzelne Unterarten teils stark bedroht. Besonders kritisch steht es dabei um das Nördliche Breitmaulnashorn (C. s. cottoni), welches nur noch zwei Exemplare umfasst und durch ein Zuchtprogramm im Ol Pejeta Reservat in Kenia vor dem Aussterben bewahrt werden soll.[113] Durch die steigende Anzahl an durch Wilderer getöteten Nashörnern im südlichen Afrika (2013 fast 800 Nashörner) wurden 2013 neben bewaffneten Wildhütern, Enthornungen von wildlebenden Tieren oder Umsiedlungen einzelner Individuen beziehungsweise kleinerer Populationen neue Gegenmaßnahmen eingeleitet. Diese umfassen unter anderem das Einspritzen von Antiparasitika in die Hörner, hauptsächlich Arzneistoffe gegen äußere Parasiten, die im normalen Einsatz für die Nashörner unschädlich sind, beim Menschen aber Übelkeit oder Konvulsion erzeugen und die Hörner so unbrauchbar für den Markt der Traditionellen Chinesischen Medizin machen. Eine weitere Methode ist die Markierung der Hörner mit Farbstoffen, die deren Inneres rot oder pink färben. Dies soll helfen, den internationalen Handel mit illegal erbeuteten Hörnern zu verfolgen, da diese, vergleichbar zu durch ein ähnliches Verfahren markierten Banknoten, auch durch Scanner an Flughäfen aufgespürt werden können.[114][115] Umweltschützer sehen diese Vorgehensweisen bisher eher skeptisch, einerseits weil vereinzelt Nashörner bei der Durchführung der Prozedur starben, andererseits auch weil kaum wissenschaftliche Untersuchungen zu den neuen Maßnahmen und der damit verbundenen potentiellen gesundheitlichen Risiken für die behandelten Tiere durchgeführt wurden. Weiterhin können sie auch eine höhere Sicherheit der Populationen an Nashörnern lediglich vortäuschen, da zwar in einzelnen Schutzgebieten mit derartig behandelten Tieren die Wilderei zurückging, ein Großteil der Tiere aber diese aufgrund des Druckes durch Wilderei verlassen hatte. Eine Verschiebung des Jagddrucks ist dadurch nicht auszuschließen.[116] Der WWF Deutschland informierte im März 2014 über ein Projekt, bei dem 1000 zu schützenden Nashörnern in Kenia ein Chip ins Horn eingepflanzt werden soll, um Wilderer besser überführen zu können.[117]

Auch Schutzbemühungen Indiens und Nepals waren erfolgreich, so dass der Bestand an Panzernashörnern auf 2.850 Tiere Ende 2010 und damit im Vergleich zu 1995 deutlich gestiegen ist; gegenwärtig wird von einem Bestand von mehr als 3.500 Tieren ausgegangen.[112] Der Bestand der Sumatra-Nashörner ist dagegen im selben Zeitraum von etwas mehr als 300 auf etwa 220 bis 280 Tiere zurückgegangen, teilweise werden auch weniger als 100 Tiere angenommen[112] – Ursache ist offensichtlich, dass die Regierungen von Indonesien und Malaysia nur wenige finanzielle Mittel für den Schutz dieser Tiere bereitstellen. Das Java-Nashorn ist mittlerweile das am stärksten gefährdete Großsäugetier der Erde. Schätzungsweise 60 Exemplare bewohnen Reste des ehemaligen Verbreitungsgebiets der Art im Westen von Java.[118] Die letzte verbliebene kleine Population des Java-Nashorns in Vietnam[119] wurde im Oktober 2011 vom WWF für erloschen erklärt.[120] Damit gilt die vietnamesische Unterart R. s. annamiticus des Java-Nashorns offiziell als ausgestorben. Aufgrund des hohen Risikos, die letzten zur Fortpflanzung fähigen Kühe zu isolieren, und der Tatsache, dass sich Nashörner allgemein in menschlicher Obhut teilweise nur selten fortpflanzen,[3] sind Nachzuchtprogramme schwer umzusetzen, so dass sich die früheren Schutzbemühungen auf das Ausrufen neuer Schutzgebiete und die Rettung der Restpopulationen beschränken. Zwei bedeutende neuere Projekte in diesem Zusammenhang sind das zum Schutz des Sumatra-Nashorns im Jahr 1997 gestartete Nachzuchtprogramm im Sumatran Rhino Sanctuary auf Sumatra und das 2011 begonnene Projekt Javan Rhino Study and Conservation Area für die Erhaltung des Java-Nashorns.[121][122]
Literatur
- Anonymous (Hrsg.): Die Nashörner: Begegnung mit urzeitlichen Kolossen. Fürth, Filander Verlag, 1997, ISBN 3-930831-06-6.
- Esperanza Cerdeño: Diversity and evolutionary trends of the the family Rhinocerotidae (Perissodactyla). In: Palaeo. 141, 1998, S. 13–34.
- Claude Guérin: La famille des Rhinocerotidae (Mammalia, Perissodactyla): systématique, histoire, évolution, paléoécologie. In: Cranium. 6, 1989, S. 3–14.
- Kurt Heissig: The Rhinocerotidae. In: Donald R. Prothero, R. M. Schoch (Hrsg.): The evolution of the Perissodactyls. New York 1989, S. 399–417.
- Donald R. Prothero, Claude Guérin, Earl Manning: The history of Rhinocerotoidea. In: Donald R. Prothero, R. M. Schoch (Hrsg.): The evolution of the Perissodactyls. New York 1989, S. 321–340.
- Donald R. Prothero: Fifty million years of rhinoceros evolution. In: O. A. Ryder (Hrsg.): Rhinoceros biology and conservation: Proceedings of an international conference, San Diego, U.S.A. Zoological Society, San Diego 1993, S. 82–91.
- Rudolf Schenkel, Ernst M. Lang: Das Verhalten der Nashörner (= Handbuch der Zoologie. 8 (46)). 1969, S. 1–56, ISBN 3-11-000664-2.
Einzelnachweise
- Esperanza Cerdeño: Diversity and evolutionary trends of the the family Rhinocerotidae (Perissodactyla). In: Palaeo. 141, 1998, S. 13–34.
- Ralf-Dietrich Kahlke, Frédéric Lacombat: The earliest immigration of woolly rhinoceros (Coelodonta tologoijensis, Rhinocerotidae, Mammalia) into Europe and its adaptive evolution in Palaearctic cold stage mammal faunas. In: Quaternary Science Reviews. 27, 2008, S. 1951–1961.
- R. Fulconis: Save the rhinos: EAZA Rhino Campaign 2005/6. Info Pack, London 2005.
- Kurt Heissig: The Rhinocerotidae. In: Donald R. Prothero, R. M. Schoch (Hrsg.): The evolution of the Perissodactyls. New York 1989, S. 399–417.
- Donald R. Prothero: Rhino giants. The paleobiology of Indricotheres. Indiana University Press, 2013, S. 54–55, ISBN 978-0-253-00819-0.
- Frédéric Lacombat: The Evolution of the rhinoceros. In: R. Fulconis (Hrsg.): Save the rhinos: EAZA Rhino Campaign 2005/6. Info Pack, London 2005, S. 46–49.
- Vladimir Zhegallo, Nikolay Kalandatze, Andrey Shapovalov, Zoya Bessudnova, Natalia Noskova, Ekaterina Tesakova: On the fossile rhinoceros Elasmotherium (Including the collections of the Russian Academy of sciences). In: Cranium. 22 (1), 2005, S. 17–40.
- Jérémy Tissier, Pierre-Olivier Antoine, Damien Becker: New material of Epiaceratherium and a new species of Mesaceratherium clear up the phylogeny of early Rhinocerotidae (Perissodactyla). In: Royal Society Open Science. 7, 2020, S. 200633, doi:10.1098/rsos.200633.
- Donald R. Prothero, Claude Guérin, Earl Manning: The history of Rhinocerotoidea. In: Donald R. Prothero, R. M. Schoch (Hrsg.): The evolution of the Perissodactyls. New York 1989, S. 321–340.
- Peter Kemnitz, Wolfgang Puschmann, Michael Schröpel, Dieter Krause, Rüdiger Schöning: Feingewebliche Untersuchungen zur Struktur und Ontogenese des Hornes von Nashörnern, Rhinocerotidae: ein Atlas mit neuen Ansichten auf und über ein altes Problem. In: Zoologischer Garten. 61 (3), 1991, S. 177–199.
- M. L. Ryder: Structure of rhinoceros horn. In: Nature. 193, 1962, S. 1199–1201.
- Fedor K. Shidlovskiy, Irina V. Kirillova, John Wood: Horns of the woolly rhinoceros Coelodonta antiquitatis (Blumenbach, 1799) in the Ice Age Museum collection (Moscow, Russia). In: Quaternary International. 2011, S. 1–5.
- О. Ф. Чернова, И. В. Кириллова: Новые данные о морфологии рога шерстистого носорога (Coelodonta antiquitatis BLUMENBACH, 1799). In: Труды Зоологического института РАН. 314 (3), 2010, S. 333–342.
- Colin P. Groves: Species characters in rhinoceros horns. In: Zeitschrift für Säugetierkunde. 36 (4), 1971, S. 238–252.
- Colin P. Groves, Fred Kurt: Dicerorhinus sumatrensis. In: Mammalian Species. 21, 1972, S. 1–6.
- W. A. Laurie, E. M. Lang, Colin P. Groves: Rhinoceros unicornis. In: Mammalian Species. 211, 1983, S. 1–6.
- Nan E. Schaffer, George L. Foley, Sophia Gill, C. Earl Pope: Clinical implications of rhinoceros reproductive tractanatomy and histology. In: Journal of Zoo and Wildlife Medicine. 32(1), 2001, S. 31–46.
- Donald R. Prothero: Fifty million years of rhinoceros evolution. In: O. A. Ryder (Hrsg.): Rhinoceros biology and conservation: Proceedings of an international conference, San Diego, U.S.A. Zoological Society, San Diego 1993, S. 82–91.
- Toni Bürgin, Urs Oberli: Ein Zufallsfund aus dem Steinbruch. In: Fossilien. Zeitschrift für Sammler und Hobbypaläontologen. 27 (4), 2010, S. 238 f.
- Rudolf Schenkel, Ernst M. Lang: Das Verhalten der Nashörner. (= Handbuch für Zoologie. 8 (46)). 1969, S. 1–56, ISBN 3-11-000664-2.
- Adrian Lister und Paul Bahn: Mammuts – Die Riesen der Eiszeit. Sigmaringen, 1997 (S. 89).
- Nina V. Garutt: Dental ontogeny of the woolly rhinoceros Coelodonta antiquitatis (Blumenbach, 1799). In: Cranium. 11, 1994, S. 37–48.
- Irina V. Kirillova, Fedor K. Shidlovskiy: Estimation of individual age and season of death in woolly rhinoceros, Coelodonta antiquitatis (Blumenbach, 1799), from Sakha-Yakutia, Russia. In: Quaternary Science Reviews. 29, 2010, S. 3106–3114.
- Matthew C. Mihlbachler: Demography of late Miocene rhinoceroses (Teleoceras proterum and Aphelops malacorhinus) from Florida: linking mortality and sociality in fossil assemblages. In: Paleobiology. 29 (3), 2003, S. 412–428.
- Liang Zhong, Deng Tao: Age structure and habitat of the Rhinoceros Chilotherium during the Late Miocene in the Linxia Basin, Gansu, China. In: Vertebrata Palasiatica. 43 (3), 2005, S. 319–230.
- Roan D. Plotz, Wayne L. Linklater: Oxpeckers Help Rhinos Evade Humans. In: Current Biology. 30 (10), 2020, S. 1965–1969.e2, doi:10.1016/j.cub.2020.03.015.
- Nina Garutt: Traumatic skull damages in the woolly rhinoceros, Coelodonta antiquitatis BLUMENBACH, 1799. In: Cranium. 14, 1997, S. 37–46.
- Deng Tao, Tseng Zhijie J: Osteological evidence for predatory behavior of the giant percrocutid (Dinocrocuta gigantea) as an active hunter. In: Chinese Science Bulletin. 55 (17), 2010, S. 1790–1794.
- Jan van der Made, René Grube: The rhinoceroses from Neumark-Nord and their nutrition. In: Harald Meller (Hrsg.): Elefantenreich – Eine Fossilwelt in Europa. Halle/Saale 2010, S. 382–394.
- Friedrich E. Zeuner: Die Beziehungen zwischen Schädelform und Lebensweise bei den rezenten und fossilen Nashörnern. In: Berichte der Naturforschenden Gesellschaft in Freiburg. 34, 1934, S. 21–80.
- Kurt Heissig: Family Rhinocerotidae. In: Gertrud E. Rössner, Kurt Heissig: The Miocene land mammals of Europe. München 1999, S. 175–188.
- Bin Bai, Jin Meng, Chi Zhang, Yan-Zin Gong, Yuan-Qing Wang: The origin of Rhinocerotoidea and the phylogeny of Ceratomorpha (Mammalia, Perissodactyla). In: Communications Biology. 3, 2020, S. 509, doi:10.1038/s42003-020-01205-8.
- Christelle Tougard, Thomas Delefosse, Catherine Hänni und Claudine Montgelard: Phylogenetic Relationships of the Five Extant Rhinoceros Species (Rhinocerotidae, Perissodactyla) Based on Mitochondrial Cytochrome b and 12S rRNA Genes. In: Molecular Phylogenetics and Evolution. 19, 2001, S. 34–44.
- Cynthia C. Steiner, Oliver A. Ryder: Molecular phylogeny and evolution of the Perissodactyla. In: Zoological Journal of the Linnean Society. 163, 2011, S. 1289–1303.
- Pavel Kosintsev, Kieren J. Mitchell, Thibaut Devièse, Johannes van der Plicht, Margot Kuitems, Ekaterina Petrova, Alexei Tikhonov, Thomas Higham, Daniel Comeskey, Chris Turney, Alan Cooper, Thijs van Kolfschoten, Anthony J. Stuart, Adrian M. Lister: Evolution and extinction of the giant rhinoceros Elasmotherium sibiricum sheds light on late Quaternary megafaunal extinctions. In: Nature Ecology & Evolution. 3, 2019, S. 31–38, doi:10.1038/s41559-018-0722-0.
- Donald R. Prothero: Rhinocerotidae. In: Christine M Janus, Kathleen M Scott, Louis L Jacobs (Hrsg.): Evolution of Tertiary mammals from North America. Vol. 1, Cambridge 1998, S. 595–605.
- Leonard B. Radinsky: The families of the Rhinocerotoidea (Mammalia, Perissodactyla). In: Journal of Mammalogy. 47 (4), 1966, S. 631–639.
- Donald R. Prothero, Earl Manning, C. Bruce Hanson: The phylogeny of the rhinocerotoidea (Mammalia, Perissodactyla). In: Zoological Journal of the Linnean Society. 87, 1986, S. 341–366.
- Haibing Wang, Bin Bai, Jin Meng, Yuanqing Wang: Earliest known unequivocal rhinocerotoid sheds new light on the origin of Giant Rhinos and phylogeny of early rhinocerotoids. In: Scientific Reports. 6 (39607), 2016, S. 1–9, doi:10.1038/srep39607.
- Bin Bai, Jin Meng, Yuan Qing Wang, Hai Bing Wang, Luke T. Holbrook: Osteology of the Middle Eocene ceratomorph Hyrachyus modestus (Mammalia, Perissodactyla). In: Bulletin of the American Museum of Natural History. 413, 2017, S. 1–68.
- Kurt Heissig, Oldřich Fejfar: Die fossilen Nashörner (Mammalia, Rhinocerotidae) aus dem Untermiozän von Tuchorice in Nordwestböhmen. Sborník Národního Muzea v Praze. In: Acta Musei Nationalis Pragae (series B, Natural History). 63 (1), 2007, S. 19–64.
- Donald R. Prothero, Robert M. Schoch: Classification of the Perissodactyla. In: Donald R. Prothero, R. M. Schoch (Hrsg.): The evolution of the Perissodactyls. New York 1989, S. 530–537.
- Pierre-Olivier Antoine, Francisco Alférez, Carlos Iñigo: A new elasmotheriine (Mammalia, Rhinocerotidae) from the early Miocene of Spain. In: Comptes Rendus Pale. vol 1, 2002, S. 19–26.
- Shanlin Liu, Michael V. Westbury, Nicolas Dussex, Kieren J. Mitchell, Mikkel-Holger S. Sinding, Peter D. Heintzman, David A. Duchêne, Joshua D. Kapp, Johanna von Seth, Holly Heiniger, Fátima Sánchez-Barreiro, Ashot Margaryan, Remi André-Olsen, Binia De Cahsan, Guanliang Meng, Chentao Yang, Lei Chen, Tom van der Valk, Yoshan Moodley, Kees Rookmaaker, Michael W. Bruford, Oliver Ryder, Cynthia Steiner, Linda G.R. Bruins-van Sonsbeek, Sergey Vartanyan, Chunxue Guo, Alan Cooper, Pavel Kosintsev, Irina Kirillova, Adrian M. Lister, Tomas Marques-Bonet, Shyam Gopalakrishnan, Robert R. Dunn, Eline D. Lorenzen, Beth Shapiro, Guojie Zhang, Pierre-Olivier Antoine, Love Dalén, M. Thomas P. Gilbert: Ancient and modern genomes unravel the evolutionary history of the rhinoceros family. In: Cell., 2021, doi:10.1016/j.cell.2021.07.032.
- Ashot Margaryan, Mikkel-Holger S. Sinding, Shanlin Liu, Filipe Garrett Vieira, Yvonne L. Chan, Senthilvel K. S. S. Nathan, Yoshan Moodley, Michael W. Bruford, M. Thomas P. Gilbert: Recent mitochondrial lineage extinction in the critically endangered Javan rhinoceros. In: Zoological Journal of the Linnean Society., 2020, doi:10.1093/zoolinnean/zlaa004/5802322.
- Claude Guérin: La famille des Rhinocerotidae (Mammalia, Perissodactyla): systématique, histoire, évolution, paléoécologie. In: Cranium. 6, 1989, S. 3–14.
- Ludovic Orlando, Jennifer A. Leonard, Aurélie Thenot, Vincent Laudet, Claude Guerin, Catherine Hänni: Ancient DNA analysis reveals woolly rhino evolutionary relationships. In: Molecular Phylogenetics and Evolution. 28, 2003, S. 485–499.
- Xiufeng Xu, Axel Janke, Ulfur Arnason: The Complete Mitochondrial DNA Sequence of the Greater Indian Rhinoceros, Rhinoceros unicornis, and the Phylogenetic Relationship Among Carnivora, Perissodactyla, and Artiodactyla (+ Cetacea). In: Molecular Biology and Evolution. 13 (9), 1996, S. 1167–1173.
- Esperanza Cerdeño: Cladistic analysis of the family Rhinocerotidae (Perissodactyla). In: American Museum Novitates. 3143, 1995, S. 1–25.
- Pierre-Olivier Antoine: Middle Miocene elasmotheriine Rhinocerotidae from China and Mongolia: taxonomic revision and phylogenetic relationships. In: The Norwegian Academy of Science and Letters - Zoologica Scripta. 32, 2003, S. 5–118.
- Colin P. Groves: Die Nashörner – Stammesgeschichte und Verwandtschaft. In: Die Nashörner: Begegnung mit urzeitlichen Kolossen. Fürth 1997, S. 14–32, ISBN 3-930831-06-6.
- Colin P. Groves: Rhinoceros sondaicus. In: Mammalian Species. 43 (887), 2011, S. 190–208.
- Deng Tao: A primitive species of Chilotherium (Perissodactyla, Rhinocerotidae) from the Late Miocene of the Linxia Basin (Gansu, China). In: Cainozoic Research. 5 (1-2), 2006, S. 93–102.
- Donald R. Prothero: The evolution of North American rhinoceroses. Cambridge University Press, 2005.
- Denis Geraads: Rhinocerotidae. In: L. Werdelin, D. J. Sanders (Hrsg.): Cenozoic Mammals of Africa. Berkeley 2010, S. 669–683.
- Denis Geraads, Monte MacCrossin, Brenda Benefit: A New Rhinoceros, Victoriaceros kenyensis gen. et sp. nov., and other Perissodactyla from the Middle Miocene of Maboko, Kenya. In: Journal of Mammalian Evolution. 19 (1), 2012, S. 57–75.
- Damien Becker, Pierre-Olivier Antoine, Olivier Maridet: A new genus of Rhinocerotidae (Mammalia, Perissodactyla) from the Oligocene of Europe. In: Journal of Systematic Palaeontology. 11 (8), 2013, S. 947–972, doi:10.1080/14772019.2012.699007.
- Luca Pandolfi: Persiatherium rodleri, gen. et sp. nov. (Mammalia, Rhinocerotidae) fro, the Upper Miocene of Maragheh (Northwestern Iran). In: Journal of Vertebrate Paleontology. 36 (1), 2016, S. e1040118, doi:10.1080/02724634.2015.1040118.
- Naoto Handa, Masato Nakatsukasa, Yutaka Kunimatsu, Hideo Nakaya: A new Elasmotheriini (Perissodactyla, Rhinocerotidae) from the upper Miocene of Samburu Hills and Nakali, northern Kenya. In: Geobios. 50 (3), 2017, S. 197–209, doi:10.1016/j.geobios.2017.04.002.
- Pierre-Olivier Antoine, Marian C. Reyes, Noel Amano, Angel P. Bautista, Chun-Hsiang Chang, Julien Claude, John De Vos, Thomas Ingicco: A new rhinoceros clade from the Pleistocene of Asia sheds light on mammal dispersals to the Philippines. In: Zoological Journal of the Linnean Society., 2021, doi:10.1093/zoolinnean/zlab009.
- Denis Geraads, Samir Zouhri: A new late Miocene elasmotheriine rhinoceros from Morocco. In: Acta Palaeontologica Polonica. 66, 2021, doi:10.4202/app.00904.2021.
- Luca Pandolfi, Antoine Pierre-Olivier, Maria Bukhsianidze, Daid Lordkipanidze, Lorenzo Rook: Northern Eurasian rhinocerotines (Mammalia, Perissodactyla) by the Pliocene–Pleistocene transition: phylogeny and historical biogeography. In: Journal of Systematic Palaeontology., 2021, doi:10.1080/14772019.2021.1995907.
- Kerstin Hlawatsch, Jörg Erfurt: Zahnmorphologie und stratigraphische Verteilung von Hyrachyus minimus (Perissodactyla, Mammalia) in den eozänen Geiseltalschichten. In: Jörg Erfurt, Lutz Christian Maul (Hrsg.): 34. Tagung des Arbeitskreises für Wirbeltierpaläontologie der Paläontologischen Gesellschaft 16.3 bis 18.3.2007 in Freyburg/Unstrut. (= Hallesches Jahrbuch für Geowissenschaften. 23). 2007, S. 161–173.
- C. Bruce Hanson: Teletaceras radinskyi, a new primitive rhinocerotid from the late Eocene Clarno formation of Oregon. In: Donald R. Prothero, Robert M. Schoch (Hrsg.): The evolution of perissodactyls. New York und London, 1989, S. 379–398.
- Takehisa Tsubamoto, Naoko Egi, Masanaru Takai, Chit Sein, Maung Maung: Middle Eocene ungulate mammals from Myanmar: A review with description of new specimens. In: Acta Palaeontologica Polonica. 50 (1), 2005, S. 117–138.
- Jérémy Tissier, Pierre-Olivier Antoine, Damien Becker: New species, revision, and phylogeny of Ronzotherium Aymard, 1854 (Perissodactyla, Rhinocerotidae). In: European Journal of Taxonomy. 753, 2021, S. 1–80, doi:10.5852/ejt.2021.753.1389.
- Madelaine Böhme, Manuela Aiglstorfer, Pierre-Olivier Antoine, Erwin Appel, Philipe Havlik, Grégoire Métais, Laq The Phuc, Simon Schneider, Fabian Setzer, Ralf Tappert, Dang Ngoc Tran, Dieter Uhl, Jérôme Prieto: Na Duong (northern Vietnam) – an exceptional window into Eocene ecosystems from Southeast Asia. In: Zitteliana. A 53, 2013, S. 120–167.
- Kurt Heissig: The American genus Penetrigonias Tanner & Martin, 1976 (Mammalia: Rhinocerotidae) as a stem group elasmothere and ancestor of Menoceras Troxell, 1921. In: Zitteliana. A 52, 2012, S. 79–95.
- Michael R. Voorhies, Joseph R. Thomasson: Fossil grass anthoecia within miocene rhinoceros skeletons: diet in an extinct species. In: Science. 206 (Oct 19), 1979, S. 331–333.
- Pierre-Olivier Antoine, Jean Loup Welcomme: A new rhinoceros from the lower miocene of the Bugti Hills, Baluchistan, Pakistan: the earliest elasmotheriine. In: Palaeontology. 43 (5), 2000, S. 795–816.
- Denis Geraads: Pliocene Rhinocerotidae (Mammalia) from Hadar and Dikika (Lower Awash, Ethiopia) and a revision of the origin of modern African rhinos. In: Journal of Vertebrate Paleontology. 25 (2), 2005, S. 451–461.
- Deng Tao: Linxia Basin: An Ancient Paradise for Late Cenozoic Rhinoceroses in North China. In: Paleomammalogy. 24 (2), 2010, S. 103–106.
- Tao Deng, Xiaoming Wang, Mikael Fortelius, Qiang Li, Yang Wang, Zhijie J. Tseng, Gary T. Takeuchi, Joel E. Saylor, Laura K. Säilä, Guangpu Xie: Out of Tibet: Pliocene Woolly Rhino Suggests High-Plateau Origin of Ice Age Megaherbivores. In: Science. 333, September 2011, S. 1285–1288.
- Akira Fukuchi, Hideo Nakaya, Masanaru Takai, Shintaro Ogino: A preliminary report on the Pliocene rhinoceros from Udunga, Transbaikalia, Russia. In: Asian Paleoprimatology. 5, 2009, S. 61–98.
- L. C. Rookmaaker: Early rhinoceros systematics. In: A. Wheeler u. a. (Hrsg.): History in the service of systematics. Society for the Bibliography of Natural History, London 1981, S. 111–118.
- Ragnar K. Kinzelbach: An Indian Rhino, Rhinoceros unicornis, (V34) on the Artemidorus Papyros. Its position in the antique cultural tradition. In: Jochen Althöff, Sabine Föllinger, Georg Wöhrle (Hrsg.): Antike Naturwissenschaft und ihre Rezeption. Wissenschaftlicher Verlag Trier, 2012, S. 93–131, ISBN 978-3-86821-389-8.
- Wilhelm Pape und Max Sengebusch: Handwörterbuch der griechischen Sprache. 3. Auflage, 6. Abdruck, Vieweg & Sohn, Braunschweig, 1914 ()
- Linnaeus: Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis Editio decima, reformata. Holmiae, Laurentii Salvii, 1758, S. 56.
- Mikael Fortelius: The morphology and paleobiological significance of the horns of Coelodonta antiquitatis (Mammalia: Rhinocerotidae). In: Journal of Vertebrate Paleontology. 3 (2), 1983, S. 125–135.
- John Edward Gray: On the natural arrangement of vertebrose animals. In: London Medical Repository. 15, 1821, S. 297–310 (306) (PDF)
- M. C. MacKenna, S. K. Bell: Classification of mammals above the species level. Columbia University Press, New York 1997, S. 483–485.
- Richard Owen: Odontography; or, A treatise on the comparative anatomy of the teeth; their physiological relations, mode of development, and microscopic structure, in the vertebrate animals. Hippolyte Bailliere, London 1840–1845, S. 587–599 ()
- Ingmar Braun, Wolfgang Zessin: Paläolithische Nashorndarstellungen und der Versuch ihrer zoologisch-ethologischen Interpretation. In: Ursus, Mitteilungsblatt des Zoovereins und des Zoos Schwerin. 15 (1), 2009, S. 3–19.
- Michel Lorblanchet: Höhlenmalerei. Ein Handbuch. Sigmaringen 1997.
- W. T. Schaurte: Darstellung eines Elasmotherium in der Felsmalerei von Rouffignac. In: Natur und Museum. 94 (9), 1964, S. 354–356.
- Baldyrgan S. Kožamkulova: Elasmotherium sibiricum und sein Verbreitungsgebiet auf dem Territorium der UdSSR. In: Quartärpaläontologie. 4, 1981, S. 85–91.
- Meenakshi Dubey: Rock paintings of Pachmarhi Hills. In: Michel Lorblanchet (Hrsg.): Rock art in the old world. Indira Gandhi National Centre for the Arts, New Delhi 2001, S. 131–145.
- G. Kreuzer: Tierdarstellungen in der indischen Felsbildkunst. In: Die Blauen Hefte. 58, 1978, S. 405–407.
- Sven Ouzman: Seeing is deceiving: rock art and the non-visual. In: World Archaeology. 33 (2), 2001, S. 237–256.
- Sven Ouzman, Jim M. Feely: Black or white? The identification and significance of rhinoceroses in South African rock art. In: Digging Stick. (SA Archaeological Society) 19 (2), 2010, S. 9–12.
- A. van der Geer: Rhinoceros unicornis: The Indian Rhinoceros. In: A. van der Geer: Animals in stone: Indian mammals sculptured through time. (= Handbook of Oriental Studies. section 2: India 21). Leiden/ Boston 2008, S. 380–386.
- W. F. Gowers: The classical rhinoceros. In: Antiquity. 24, 1950, S. 61–71.
- H. H. de Iongh, H. H. T. Prins, N. van Strien, L. C. Rookmaaker: Some observations on the presence of one-horned rhinos in the bas reliefs of the Angkor Wat temple complex, Cambodia. In: Pachyderm. 38 (Januay-June), 2005, S. 98–100.
- Maryna Steyn: The Mapungubwe Gold Graves Revisited. In: The South African Archaeological Bulletin. 62 (186), 2007, S. 140–146.
- Stephan Woodborne, Marc Pienaar, Sian Tiley-Nel: Dating the Mapungubwe Hill Gold. In: Journal of African Archaeology. 7 (1), 2009, S. 99–105.
- Alex Duffey: Mapungubwe: Interpretation of the Gold Content of the "Original Gold Burial M1, A620". In: Journal of African Archaeology. 10 (2), 2012, S. 175–187.
- L. C. Rookmaaker: Captive rhinoceroses in Europe from 1500 until 1810. In: Bijdragen tot de Dierkunde. 43, 1973, S. 39–63.
- International Rhino Foundation: 2007 Annual Report. Yulee, 2007.
- International Rhino Foundation: Annual Report 2013. ().
- International Rhino Foundation: Annual Report 2020. (), abrufbar auf Rhinos.org.
- Jared Diamond: Guns, Germs, and Steel: The Fates of Human Societies. New York/ London 1997, S. 168–174, ISBN 0-393-31755-2.
- E. B. Martin: Tackling the rhino horn trade - a new approach. In: Species. 7, 1986, S. 7–8.
- U. A. Casal: Carved Rhinoceros Horns of China. In: Cultureel Indie. 2 (Aug./Sept.), 1940, S. 212–216.
- V. Dao Truong, Madelon Willemsen, Nam V. H. Dang, Trinh Nguyen, C. Michael Hall: The marketplace management of illegal elixirs: illicit consumptionof rhino horn. Consumption Markets & Culture 19, 2016 (4), S. 353–369, doi:10.1080/10253866.2015.1108915.
- Mike Knight: African Rhino Specialist Group report. In: Pachyderm. 50 (Juli bis Dezember), 2011, S. 7–14 (online)
- RP: Die erste Spur zur Nashorn-Mafia. () rp-online.de vom 3. März 2012:
- Kate Katharina Ferguson: Rhino Horn Thefts a Growing Problem in Europe. In: Spiegel Online. 13. März 2012 (PDF).
- L. C. Rookmaaker: New literature in the Rhino Resource Center. In: Electronic Newsletter of the Rhino Resource Center. no. 25 (November), 2011, S. 1–16 (PDF).
- Felix Patton: The medicinal value of rhino horn – a quest for the truth. Unpublished report (first available on RRC), 2011, S. 1–8 (PDF).
- Lothar Frenz: Nashörner. Naturkunden No. 26, Matthes & Seitz, Berlin 2017, S. 86.
- Mike H. Knight: African Rhino Specialist Group Chair report. In: Pachyderm. 49, 2011, S. 6–15 ().
- International Rhino Foundation: State of the Rhino. ().
- OlPejetaConservancy: Rhinos. () auf: olpejetaconservancy.org.
- International Rhino Foundation: Poaching Deterrents. (10. September 2014)
- Rhino Rescue Project: Proactive Poaching Prevention. rhinorescueproject.com vom 12. September 2014.
- Sam Ferreira, Markus Hofmeyr, Danie Pienaar, Dave Cooper: Chemical horn infusions: a poaching deterrent or an unnecessary deception? In: Pachyderm. 55, 2014, S. 54–61.
- WWF: Nashörner in Kenia: Big Brother is watching you. () wwf.de vom 2. März 2014.
- Bibhab Kumar Talukdar: Asian Rhino Specialist Group report. In: Pachyderm. 49, 2011, S. 16–19 ((online)).
- WWF: Vietnams letzte Nashörner sollen Straße weichen. () wwf.de vom 26. Mai 2009.
- WWF: Schwarzer Tag für den Artenschutz. () wwf.de vom 25. Oktober 2011.
- Abdul Wahab Ahmad Zafir, Junai di Payne, Azlan Mohamed, Ching Fong Lau, Dionys i us Shankar Kumar Sharma, Raymond Alfred Amirtharaj Christy Williams, Senthival Nathan, Widodo S . Ramono, Gopalasamy Reuben Clements: Now or never: what will it take to save the Sumatran rhinoceros Dicerorhinus sumatrensis from extinction? In: Fauna & Flora International, Oryx. 45 (2), 2011, S. 225–233.
- Susie Ellis, Maggi Moore: Conserving the Javan rhinos. In: International Zoo News. 58 (6), 2011, S. 403–405.
{kind=link}
Weblinks
- Nashörner Spiegel online, Aktuelle Artikel und Hintergründe
- World Wildlife Fund (WWF): Rhino: Species, Places, Threats, Our Work
- Themen & Projekte: Nashorn WWF Deutschland