Magen
Der Magen (von mittelhochdeutsch mage; altgriechisch γαστῆρ gaster, lateinisch ventriculus; auch griechisch-lateinisch stomachus) ist ein Verdauungsorgan fast aller Tiere. Die primitivste Form ist der Gastralraum der Nesseltiere, der zugleich Ausscheidungsorgan ist. Der Magen ist ein Hohlorgan aus Muskelgewebe, das innen mit einer Schleimhaut ausgekleidet ist. Im Gegensatz zum einhöhligen Magen des Menschen und der meisten Tierarten besitzt der Magen bei Wiederkäuern und Vögeln mehrere abgegrenzte Hohlraumsysteme (mehrhöhliger Magen) sowie hochspezialisierte Organe bei Insekten (Honigmagen, Saugmagen). Magenlos sind z. B. die Karpfenfische (man deutet dies als ursprüngliche Anpassung an Schnecken- und Muschel-Nahrung, deren Kalkschalen die Magensäure nutzlos machten).[1]
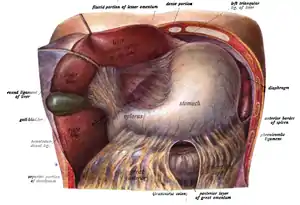
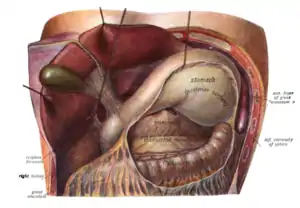
Das Fassungsvermögen des menschlichen Magens ist individuell unterschiedlich und beträgt etwa 1,5 l.[2] In diesem Hohlraum wird der Nahrungsbrei mit dem Magensaft vermengt, der im Wesentlichen aus dem eiweißspaltenden Enzym Pepsin und Salzsäure besteht. Der Magen bringt auch den Nahrungsbrei auf die gleiche Temperatur, außerdem schichtet und speichert er ihn. Im Ruhezustand sondern die Drüsen etwa 10 ml Magensaft pro Stunde ab. Bei Nahrungsaufnahme kann die Bildung von Magensaft auf bis zu 1000 ml pro Stunde erhöht werden. Die Bildung wird sowohl durch Nerven-Impulse (vor allem vor der Nahrungsaufnahme) als auch durch Hormone gesteuert. Durch Muskelkontraktionen (Peristaltik) wird der Nahrungsbrei weiter zum Pförtner transportiert. Dieser bildet als Sphinkter (Schließmuskel) einen Verschluss zwischen Magen und Darm und kann sich für den Durchtritt des angedauten Speisebreis (Chymus) etwa 13 mm weit öffnen. Dieses geschieht regelmäßig, um den Nahrungsbrei gleichmäßig in den dem Magenausgang folgenden Zwölffingerdarm weiterzuleiten.
Biologische Bedeutung
Vorverdauung
Die eigentliche Verdauung und Resorption findet bei den meisten Tieren und beim Menschen im Darm oder spezieller im Dünndarm statt. Im Magen werden vor allem die Proteine (Eiweiße) in sogenannte Polypeptidketten vorverdaut. Die notwendigen Enzyme zur Eiweißverdauung, Pepsin und Kathepsin, haben durch den niedrigen pH-Wert höchste Aktivität. Sie werden von den Zellen des Magens als inaktive Vorstufen (Pepsinogen) abgesondert und erst durch Kontakt mit der Magensäure aktiviert. Fette passieren den Magen größtenteils ungehindert, sie werden durch die Peristaltik jedoch weicher gemacht (verflüssigt). Kohlenhydrate passieren den Magen genauso ungehindert, da die enzymatische Spaltung der α(1-4)-Glykosidbindung von Amylose durch α-Amylase (aus dem Speichel) durch den sauren pH-Wert im Magen wieder gehemmt wird.
Abtöten von Bakterien
Der saure Magensaft verhindert effektiv das Überleben der meisten Bakterien und schützt so vor Infektionen. Lange Zeit galt die Lehrmeinung, dass im extrem sauren Milieu des menschlichen Magens ein Wachstum von Mikroorganismen prinzipiell nicht möglich sei. Mit der Entdeckung von Helicobacter pylori in der Magenschleimhaut im Jahr 1983 musste diese Ansicht jedoch revidiert werden. Fortan galt H. pylori als die einzige Ausnahme – ein Bakterium, das in der Lage ist, sich unterhalb der schützenden Schleimschicht anzusiedeln. Wissenschaftler der Stanford University konnten 2005/2006 jedoch nachweisen, dass die Flora (vgl. Darmflora) deutlich komplexer ist als bisher angenommen: Sie entdeckten insgesamt 128 weitere den Magen besiedelnde Bakterienarten, darunter auch einen Verwandten der extremophilen Gattung Deinococcus. Insgesamt etwa zehn Prozent der identifizierten Mikroorganismen unterschieden sich genetisch erheblich von allen bisher bekannten Bakterien.
Fälschlicherweise wird der im Magen vorherrschende pH-Wert oft mit konzentrierter Säure verglichen. Der Magensaft hat zwar auch einen sehr niedrigen pH-Wert von 0,8 bis 1,5,[3] die Konzentration der Salzsäure ist jedoch geringer. Dieser niedrige pH-Wert gilt allerdings nur bei leerem Magen. Bei Nahrungszufuhr steigt der Wert und kann abhängig von der Pufferkapazität des Speisebreis auf 4,5 bis 6,5 ansteigen.[4]
Regelmäßige weitere Verdauung
Der Magen ermöglicht es, dass Lebewesen mit wenigen größeren Mahlzeiten pro Tag auskommen. Er ist in der Lage, seine Wandspannung an die jeweilige Inhaltsmenge anzupassen (Akkommodation). Der Pförtner (Pylorus) sorgt dafür, dass alle Stoffe genügend lange im Magen verbleiben und ausreichend mit den Verdauungssäften versetzt werden. Der Speisebrei wird dann langsam und gleichmäßig dem Darm zugeführt.
Aufbau beim Menschen und Tieren mit einhöhligem Magen
Anatomie
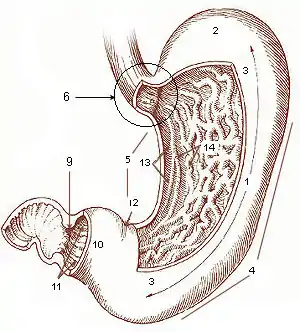
Von außen gesehen unterscheidet man am Magen die vordere obere Fläche (Paries anterior) und die hintere untere Fläche (Paries posterior), die große Krümmung (Curvatura major), an der das Omentum majus zum Teil befestigt ist, sowie die kleine Krümmung (Curvatura minor), an der das Omentum minus befestigt ist.
Makroskopisch wird er in folgende Bereiche untergliedert:
- Die Pars cardiaca (auch Cardia oder Kardia, „Mageneingang“) – mit dem Ostium cardiacum („Magenmund“) – stellt den Übergang zwischen Speiseröhre (Oesophagus) und dem Magen dar.
- Der Fundus ventriculi (oder Fundus gastricus, Fornix ventriculi, Fornix gastricus, „Magengrund“, „Magenkuppel“, „Magengewölbe“) liegt unterhalb des Zwerchfells links und kranial vom Mageneingang und ist mit bei der Nahrungsaufnahme verschluckter Luft gefüllt. Auf dem Röntgenbild erscheint er als Magenblase.
- Das Corpus ventriculi (oder Corpus gastricum, „Magenkorpus“, „Magenkörper“) macht den größten Anteil des Magens aus.
- Die Pars pylorica ist enger als das Korpus, wobei der dem Duodenum nähere Teil, der Canalis pyloricus („Pförtnerkanal“), noch enger ist als der entferntere Teil, das Antrum pyloricum („Pförtnerhöhle“), genannt auch Vestibulum pyloricum.
- Der Pylorus („Pförtner“) ist über das Ostium pyloricum („Pförtnermund“) die Verbindung zum Duodenum. Er erscheint von außen als harte Einschnürung und bildet gegen innen den Schließmuskel (Musculus sphincter pylori).
Die Form und Lage des Magens ist nicht konstant, sondern vielmehr abhängig von dessen Füllung und der Stellung des Menschen. Die Pars cardiaca ist aber durch Bindegewebe relativ stark befestigt, während der nur am Ligamentum hepatoduodenale befestigte Pylorus sich bei größerem Mageninhalt absenkt, wodurch der Magen die typische Hakenform bildet. Bei Frauen ist der Magen meist tiefer und steiler gelegen als bei Männern.
Histologie
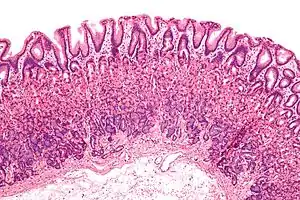
Mikroskopisch gesehen besteht der Magen aus:
- der Magenschleimhaut (Tunica mucosa gastrica), die aus den Laminae epithelialis, propria und muscularis mucosae besteht. Sie ist durch Einsenkungen (Foveolae gastricae) in zahlreiche Felder (Areae gastricae) gegliedert, mit einer Vielzahl von Drüsen (Glandulae gastricae).
- einer Bindegewebsschicht (Tela submucosa) mit Blutgefäßen
- einer Muskelschicht (Tunica muscularis gastrica) aus glatter Muskulatur, die aus den Fibrae obliquae, dem Stratum circulare und dem Stratum longitudinale besteht
- einem Überzug der Tunica serosa (Peritoneum)
Der Magen besitzt für die Sekretion von Hormonen und anderen Sekreten in seiner Schleimhaut verschiedene Zellarten, die an typischen Stellen des Magens lokalisiert sind, insbesondere
- in Korpus und Fundus: Nebenzellen (bilden Schleim), Hauptzellen (bilden Pepsinogen), Parietal- oder Belegzellen (bilden Salzsäure und Intrinsic Factor),
- im Antrum: G-Zellen (bilden Gastrin).
Embryologie
Der Magen entsteht als spindelförmige Erweiterung des Vorderdarms (der vordere Abschnitt des primitiven Darmrohrs des Embryos). Diese ist über zwei Gekröse (Mesogastrium dorsale und ventrale) an der rückenseitigen beziehungsweise bauchseitigen Körperwand befestigt. Die zunächst in der Längsachse des Embryos stehende Magenanlage erweitert sich rückenseitig zur Curvatura major und wölbt sich bauchseitig zur flach konkaven Curvatura minor ein.
Die endgültige Form und Lage des Magens bildet sich durch unterschiedliches Wachstum der Magenwandabschnitte heraus. Die Lageveränderungen werden auch als „Magendrehungen“ (nicht mit der Erkrankung Magendrehung zu verwechseln) bezeichnet, obwohl dabei nicht passive Drehungen, sondern komplizierte Umbildungsprozesse der Magenanlage stattfinden. Die sogenannte „1. Magendrehung“ kann man sich als eine Verdrehung um die Längsachse nach links um 140 Grad vorstellen. Dadurch gelangt die große Magenkrümmung von der Rückenseite nach ventrolateral (links-bauchwärts), die kleine Magenkrümmung entsprechend nach rechts dorsolateral (seitlich-rückenwärts). Die „2. Magendrehung“ lässt sich als Drehung um die senkrechte Achse um 90 Grad nach links umschreiben. Dadurch gelangt der Mageneingang nach links und der Pylorus nach rechts. Die „3. Magendrehung“ erfolgt wiederum um die Längsachse nach rechts um etwa 45 Grad. Die Curvatura major zeigt nun nach links und kaudoventral, die Curvatura minor nach rechts und kraniodorsal.
Blutversorgung des Magens
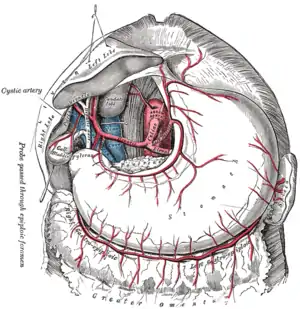
Der Magen wird arteriell über den Truncus coeliacus versorgt. Dieser kurze Gefäßstamm geht direkt von der Aorta ab und zweigt sich in drei Hauptäste auf: die Arteria hepatica communis, die Arteria gastrica sinistra und die Arteria splenica. Der Magen wird dabei direkt über die Arteria gastrica sinistra oder indirekt über weitere Abgänge der beiden anderen Äste versorgt. Der obere Teil der kleinen Kurvatur erhält sein sauerstoffreiches Blut direkt aus der Arteria gastrica sinistra. Sie läuft mit der Arteria gastrica dextra zusammen, die ursprünglich der Arteria hepatica communis entspringend den unteren Teil der kleinen Kurvatur versorgt. Beide verlaufen am Magen im kleinen Netz (Omentum minus), wo sie sich an der rechten Magenseite entlang schlängeln. Ebenfalls ursprünglich der Arteria hepatica communis entsprungen, versorgt die Arteria gastroomentalis dextra die untere Hälfte der großen Kurvatur. Sie läuft mit der Arteria gastroomentalis sinistra zusammen, die wiederum aus der Arteria splenica entstammt. Beide verlaufen am Magen im großen Netz (Omentum majus). Zusätzlich gibt die Arteria splenica noch einige Arteriae gastricae breves ab, die für die Versorgung des Fundus zuständig sind, sowie eine Arteria gastrica posterior, welche für die Durchblutung der Magenrückwand sorgt.
Vergleichende Anatomie
Magen der Wiederkäuer
Bei Wiederkäuern sind dem eigentlichen, mit einer drüsenhaltigen Schleimhaut ausgekleideten Magen, hier als Labmagen bezeichnet, noch drei Vormagenabschnitte vorgeschaltet. Dies sind Pansen, Netzmagen und Blättermagen. Sie besitzen eine drüsenlose Schleimhaut. In diesen Vormägen finden der mikrobiologische Aufschluss von Zellulose sowie erste Resorptionsvorgänge statt.
Magen der Vögel
Der Magen der Vögel ist in zwei Abschnitte unterteilt. Im eigentlichen Drüsenmagen (Ventriculus glandularis oder Proventriculus) werden ebenfalls Enzyme und Salzsäure abgegeben. Diesem Drüsenmagen ist der Muskelmagen (Ventriculus muscularis) nachgeschaltet. Er besteht aus kräftiger Muskulatur und dient der mechanischen Zerkleinerung der Nahrung und ersetzt so die Funktion der Zähne und des Kauens. Die Drüsen des Muskelmagens sondern ein Sekret ab, das durch die Salzsäure des Proventriculus zu einer Reibeplatte aushärtet (Koilinschicht). Zudem nehmen viele Vögel Steine (oder andere harte Partikel wie Muscheln bei Seevögeln) auf, die zusammen mit dieser Reibeplatte die Nahrung zermahlen. Diese Magensteine werden als Gastrolithen oder als Grit bezeichnet. Bei Vögeln, die sich von leichtverdaulicher oder weicher Nahrung ernähren, ist der Muskelmagen nur gering entwickelt.
Verlust des Magens im Laufe der Evolution
Der Magen ist vor etwa 450 Millionen Jahren erstmals bei den Wirbeltieren (Vertebrata) aufgetreten. Es wird vermutet, dass viele Wirbeltiere den Magen und vielfach auch die zugehörigen Gene (z. B. für Pepsinogen und die Protonen-Kalium-Pumpe) danach wieder verloren hätten, dazu gehören u. a. urtümliche Säugetiere wie das Schnabeltier und der Ameisenigel sowie etwa ein Viertel aller Fischarten, i. e. S. die echten Knochenfische (Teleostei).[5] Der Verlust des Magens bei zahlreichen Abstammungslinien könnte mit dem Fehlen bestimmter Protonenpumpen sowie pepsinogener Enzyme zusammenhängen.[6]
Physiologie
- Magenentleerung
Geschluckte Speisen werden vorübergehend gespeichert und dabei zerkleinert. Diese Homogenisierung geschieht während einer Verweildauer von 1–6 Stunden. Durch eine Dehnung des Magens im oberen Korpusbereich durch den Chymus, werden peristaltische Wellen ausgelöst. Der Entleerungsvorgang des Magens hängt von zahlreichen Faktoren ab. Die Magenentleerung erfolgt reflektorisch und portionsweise durch Erschlaffung des Magenpförtners (Pylorus). Sie wird, neben dem vegetativen Nervensystem, zudem durch gastrointestinale Hormone und Peptide (deren genaue Funktion noch nicht geklärt ist) sowie durch die Nahrungszusammensetzung (z. B. Menge, Temperatur) gesteuert.[7] Ein weiterer Mechanismus zur Steuerung der Entleerungsrate sind Chemosensoren im Dünndarm.
Die Flüssigkeitsentleerung ist, durch den niedrigen Magenpförtnerdruck, vor allem vom Druckgradienten zwischen Magen und Zwölffingerdarm abhängig. Flüssigkeiten verlassen den Magen relativ schnell (Halbwertszeit von Wasser auf nüchternen Magen: 10–20 Minuten). Die Entleerung fester Bestandteile ist in erster Linie vom Magenpförtnerwiderstand und der Partikelgröße abhängig. Partikel verlassen den Magen ab einer Zerkleinerung auf zwei Millimeter.[7]
Große oder unverdauliche Nahrungsmittel verlassen den Magen in der Verdauungsruhe (interdigestive Motoraktivität).[7][8]
Fehlbildungen des Magens
Erkrankungen des Magens
Siehe auch
- Magenknurren
- Magenband – eine medizinische Behandlungsmethode bei krankhafter Adipositas
- Menenius: Parabel vom Magen und den Gliedern
Literatur
- Franz X. Sailer: Chirurgie der Bauchorgane und der Bauchwand: Magen. In: Chirurgie historisch gesehen: Anfang – Entwicklung – Differenzierung. Hrsg. von Franz X. Sailer und F. W. Gierhake, Dustri-Verlag, Deisenhofen bei München 1973, ISBN 3-87185-021-7, S. 42–71.
- Franz-Viktor Salomon: Magen, Ventriculus (Gaster). In: Salomon u. a. (Hrsg.): Anatomie für die Tiermedizin. Enke-Verlag Stuttgart, 2., erw. Auflage 2008, ISBN 978-3-8304-1075-1, S. 272–293.
- Hans Adolf Kühn: Krankheiten des Magens und Zwölffingerdarmes. In: Ludwig Heilmeyer (Hrsg.): Lehrbuch der Inneren Medizin. Springer-Verlag, Berlin/Göttingen/Heidelberg 1955; 2. Auflage ebenda 1961, S. 767–804.
Weblinks
- Albertinen-Krankenhaus Hamburg: Endoskopieatlas mit Bildern und Videos des Magens
- Lehrstuhl für Humanbiologie der Technischen Universität München: Magen-Darmmotorik mit Hilfe der Videofluoroskopie sichtbar gemacht: Der bewegte Darm von Hans Jörg Ehrlein und Michael Schemann
Einzelnachweise
- Henryk Szarski (1965), Bull. Acad. Pol. Sci., Biol., 4: 155 f.
- Benninghoff, Drenckhahn: Anatomie. 16. Auflage, Elsevier Verlag (2002), S. 655.
- Mutschler: Arzneimittelwirkungen. 9. Auflage, Wissenschaftliche Verlagsgesellschaft, Stuttgart 2008, ISBN 978-3-8047-1952-1.
- Rehner, Daniel: Biochemie der Ernährung. 3. Auflage. Spektrum Akademischer Verlag, 2010, ISBN 978-3-8274-2041-1, S. 320.
- How The Platypus And A Quarter Of Fishes Lost Their Stomachs. Abgerufen am 1. April 2014.
- L. Filipe C. Castro, Odete Gonçalves, Sylvie Mazan, Boon-Hui Tay, Byrappa Venkatesh und Jonathan M. Wilson: Recurrent gene loss correlates with the evolution of stomach phenotypes in gnathostome history. Proceedings of the Royal Society B, Vol. 281, No. 1775; 2014. doi:10.1098/rspb.2013.2669
- Robert F. Schmidt, Florian Lang, Manfred Heckmann: Physiologie des Menschen. mit Pathophysiologie. 31. Auflage. SpringerMedizin Verlag, Heidelberg 2010, ISBN 978-3-642-01650-9, S. 803–809.
- Robert F. Schmidt, Florian Lang, Manfred Heckmann: Physiologie des Menschen. mit Pathophysiologie. 31. Auflage. SpringerMedizin Verlag, Heidelberg 2010, ISBN 978-3-642-01650-9, S. 797.