Diastema (Zoologie)
Diastema (von altgriechisch διάστημα diástēma „Zwischenraum“)[1] oder Diastem nennt man in der Zoologie eine evolutionär erworbene, folglich nicht-pathologische, mehr oder weniger breite Lücke in der Zahnreihe von Säugetieren und anderen Amnioten mit stark differenziertem Gebiss. Diastemata kommen bei zahlreichen rezenten und ausgestorbenen Amniotenarten an verschiedenen Positionen im Gebiss vor. In der Regel treten Diastemata im Bereich des Eckzahnes bzw. im Bereich zwischen dem hintersten Schneidezahn und dem vordersten Backenzahn auf. Besonders ausgeprägte Diastemata finden sich bei Vertretern von Amniotengruppen, die sich auf hartfaserige pflanzliche Nahrung spezialisiert haben, unter anderem bei Huftieren.
Primaten
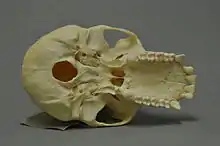
Das Diastem der Primaten ist der relativ kleine Zwischenraum zwischen Eck- und Schneidezahn im Oberkiefer und der noch kleinere Zwischenraum zwischen Eckzahn und erstem Prämolar im Unterkiefer. Dieses Diastem wird auch als Affenlücke oder Primatenlücke bezeichnet. Sie tritt sowohl bei Hunds- als auch bei Menschenaffen auf und hat die Funktion, bei geschlossenem Maul Platz für die Spitzen der relativ großen, insbesondere bei Männchen bisweilen sehr kräftig entwickelten Eckzähne der gegenüberliegenden Zahnreihe zu bieten. Beim modernen Menschen (Homo sapiens) gibt es keine Affenlücke, die Zahnreihe ist geschlossen und die Eckzähne sind nicht größer als die Schneidezähne. Bei fossilen Kiefern von Frühmenschen gilt das Vorhandensein oder Fehlen der Affenlücke als Hinweis darauf, ob das entsprechende Exemplar einer eher affenähnlichen, „primitiven“ bzw. einer eher menschenähnlichen, „fortschrittlichen“ Form zuzuordnen ist. So ist schon bei einigen Australopithecinen die Affenlücke komplett reduziert.[2]
Huftiere

Bei Huftieren besteht zwischen den äußeren (hinteren) Schneidezähnen und den vorderen Backenzähnen (meist die Prämolaren) ein relativ weites Diastem. Die Eckzähne sind bei vielen Huftieren komplett reduziert. Bei zahlreichen Paarhuferlinien, vor allem aus der Gruppe der Wiederkäuer, sind zudem auch die oberen Schneidezähne komplett reduziert, sodass im Oberkiefer kein Diastem im eigentlichen Sinn vorliegt, weil die Bezahnung erst mit den Prämolaren beginnt. Beim sehr stark modifizierten Schädel und Gebiss der Elefanten tritt ein echtes Diastem nur im Oberkiefer auf, weil der Unterkiefer keine Schneidezähne besitzt.
Die Funktion des Diastems der Huftiere ist nicht genau geklärt. Unter anderem wird eine größere Bewegungsfreiheit der Zunge beim Weiterreichen der Nahrung von den Schneidezähnen zu den Backenzähnen bzw. generell bei der Manipulation der Nahrung im Mundraum angeführt.[3] Ein anderer Erklärungsansatz besagt, dass das Diastem eher ein Ausdruck der nachlassenden Hebelwirkung mit zunehmender Länge des Hebelarms ist: Weil Mahlzähne relativ hohe metabolische Kosten verursachen, wäre bei langen Schädeln bzw. Kiefern eine Mahlfläche, die bis zu den Schneidezähnen reicht, unökonomisch. Ein Diastem wäre somit schlicht das Nebenprodukt eines langen Schädels.[4]
Nage- und Hasentiere

Auch Nage- und Hasentiere besitzen ein deutlich ausgeprägtes Diastem zwischen den Prämolaren und den Schneidezähnen bei reduziertem Eckzahn, wobei die Nagetiere nur einen Schneidezahn pro Kieferhälfte besitzen, der aber kräftiger ausgebildet ist als die beiden Schneidezähne der Hasentiere.
Die Präsenz des Diastems bei den Nagetieren steht in engem Zusammenhang mit der Evolvierung der großen „wurzellosen“ Schneidezähne („Nagezähne“), die tief in den Knochen hineinreichen, wodurch im vorderen Teil der Kiefer kein Platz mehr für die Wurzeln anderer Zähne ist. Zudem sorgt das Diastem für eine funktionelle Entkopplung der Nagezähne von den Backenzähnen: Beim Gebrauch der Nagezähne wird der Unterkiefer nach vorn geschoben, und die Backenzähne des Unterkiefers befinden sich dann gegenüber dem Diastem im Oberkiefer. So sind sie vor unnötiger Abnutzung geschützt, während das Tier die Kiefer bewegt. Zudem können Nager mit ihrer tief gespaltenen Oberlippe während des Einsatzes der Nagezähne als reine Gebrauchswerkzeuge (z. B. beim Biber während des Fällens von Bäumen für den Dammbau) den dahinterliegenden Mundraum verschließen, sodass sich die Nagezähne dann faktisch außerhalb des funktionellen Mundraumes befinden.[5]
Beuteltiere

Auch einige auf hartfaserige Pflanzennahrung spezialisierte Vertreter der Beuteltiergruppe Diprotodontia, unter anderem die Kängurus, besitzen ein teils sehr breites Diastem zwischen den Schneidezähnen und den Backenzähnen mit Reduktion der Eckzähne. Ursache und Funktion des Diastems dürften denen bei den plazentalen Säugetieren ähnlich sein. Da Beuteltiere mit Diastem und plazentale Säuger mit Diastem verschiedene gemeinsame Vorfahren ohne Diastem haben, handelt es sich jeweils um konvergente Entwicklungen.
Dinosaurier

Eine ähnliche Entwicklung in der Gebissmorphologie fand außerhalb der Säugetiere bei den Vogelbeckendinosauriern statt, insbesondere bei den Ceratopsiern und den Ornithopoden. Die Vertreter dieser Gruppen dürften sich ähnlich wie Huftiere ernährt und ihre Nahrung wahrscheinlich ausgiebig gekaut haben. Sowohl bei Ceratopsiern als auch bei Ornithopoden wurden die vorderen Zähne komplett durch einen scharfen Hornschnabel ersetzt, und die individuell eher kleinen Zähne im hinteren Bereich der Kiefer bildeten sogenannte Zahnbatterien, die in der Funktion den Mahlzähnen der Säugetiere sehr ähnlich waren. Als Diastem wird bei diesen Vertretern der zahnlose Bereich zwischen Hornschnabel und Zahnbatterie bezeichnet.[6]
Siehe auch
Einzelnachweise
- Wilhelm Gemoll: Griechisch-Deutsches Schul- und Handwörterbuch. München·Wien 1965.
- Erich Thenius: Evolution des Lebens – und der Mensch. Die erdgeschichtliche Dokumentation. Vortrag, Wien 1980 (PDF).
- Cristine M. Janis: Correlations between craniodental morphology and feeding behaviour in ungulates: reciprocal illumination between living and fossil taxa. In: Jeff J. Thomason (Hrsg.): Functional Morphology in Vertebrate Paleontology. Cambridge University Press, Cambridge UK·New York·Melbourne 1995, ISBN 0-521-44095-5, S. 76–98
- Walter S. Greaves: Functional predictions from theoretical models of the skull and jaws in reptiles and mammals. In: Jeff J. Thomason (Hrsg.): Functional Morphology in Vertebrate Paleontology. Cambridge University Press, Cambridge UK·New York·Melbourne 1995, ISBN 0-521-44095-5, S. 99–135
- Stuart O. Landry, Jr.: The Rodentia as Omnivores. The Quarterly Review of Biology. Bd. 45, Nr. 4, 1970, S. 351–372 (JSTOR 2821009)
- David E. Fastovsky, Joshua B. Smith: Dinosaur Paleoecology. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. Zweite Auflage. University of California Press, Berkeley 2004, S. 614–626, ISBN 0-520-24209-2