Unpaarhufer
Die Unpaarhufer oder Unpaarzeher (Perissodactyla, früher auch Mesaxonia) sind eine Ordnung der Säugetiere (Mammalia). Im Gegensatz zu den Paarhufern sind sie durch eine meist ungerade Anzahl von Zehen charakterisiert. Die Ordnung umfasst drei rezente Familien, die Pferde (Equidae), Nashörner (Rhinocerotidae) und Tapire (Tapiridae) mit insgesamt rund 17 Arten. Dass diese drei sehr unterschiedlich aussehenden Familien miteinander verwandt sind, erkannte als erster der Zoologe Richard Owen im 19. Jahrhundert, der auch den Begriff Unpaarhufer (engl. odd-toed ungulates) prägte.
| Unpaarhufer | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Afrikanischer Esel (Equus asinus) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Perissodactyla | ||||||||||||
| Owen, 1848 |

Merkmale
Allgemeines
Als Anpassung an verschiedene Habitate und Lebensweisen haben die Unpaarhufer deutliche Unterschiede im Körperbau entwickelt. Gemeinsame Merkmale gibt es im Bau der Gliedmaßen und der Zähne. Bei allen lebenden und der überwiegenden Mehrzahl der ausgestorbenen Arten handelt es sich um recht große Tiere. Mit den Nashörnern gehören die nach den Elefanten zweitgrößten landlebenden Säugetiere zu dieser Gruppe. Das ausgestorbene Paraceratherium, ein hornloses, nashornverwandtes Tier aus dem Oligozän, gilt sogar als größtes Landsäugetier aller Zeiten. Einige ursprüngliche Vertreter der Ordnung wie das Urpferdchen Hyracotherium waren mit nur 20 cm Schulterhöhe ziemlich klein. Abgesehen von Zwergzüchtungen des Hauspferdes und des Hausesels erreichen die heutigen Unpaarhufer eine Kopfrumpflänge von 180 bis 420 Zentimeter und ein Gewicht von 150 bis 3500 Kilogramm. Während Nashörner nur spärlich behaart sind und eine dicke Epidermis aufweisen, sind Tapire und Pferde mit einem dichten, kurzen Haarkleid versehen. Die meisten Arten sind grau oder braun gefärbt, Zebras allerdings tragen ein typisches Streifenkleid, und junge Tapire weisen weiße Längsstreifen auf.
Gliedmaßen

Die Hauptachse sowohl der vorderen als auch der hinteren Füße verläuft durch den Mittelstrahl, die dritte Zehe ist dementsprechend bei allen Arten die größte. Die übrigen Strahlen sind in unterschiedlichem Ausmaß reduziert worden, am wenigsten bei den Tapiren. Diese Tiere besitzen an den Vorderfüßen noch vier Zehen, eine Anpassung an den weichen Untergrund ihres Lebensraumes, und an den Hinterfüßen drei. Heutige Nashörner haben an Vorder- und Hinterfüßen drei Zehen. Bei den Pferden ist die Reduktion der Seitenstrahlen am weitesten fortgeschritten, diese Tiere besitzen nur mehr eine einzige Zehe (Monodaktylie). Die Füße sind mit Hufen versehen, die allerdings nur bei Pferden die Zehe fast vollständig bedecken; bei Nashörnern und Tapiren ist nur der Vorderrand mit einem Huf bedeckt, die Unterseite ist weich – Nashörner haben zusätzlich ein weiches Sohlenkissen.
Innerhalb der Beine sind die Elle und das Wadenbein verkleinert, bei den Pferden sind diese Knochen in der unteren Hälfte sogar mit Speiche beziehungsweise Schienbein verwachsen. Eine Autapomorphie (ein gemeinsames Merkmal, das diese Gruppe eindeutig von anderen Gruppen unterscheidet) ist das sattelförmige Talonaviculargelenk – das Sprunggelenk zwischen Sprungbein (Talus) und Kahnbein (Naviculare) –, das die Beweglichkeit stark einschränkt. Der Oberschenkel ist relativ kurz, das Schlüsselbein fehlt.
Schädel und Zähne

Unpaarhufer haben einen langgestreckten Kopf, wofür in erster Linie ein langer Oberkiefer (Maxillare) verantwortlich ist. Die verschiedenen Schnauzenformen der einzelnen Familien gehen auf Unterschiede im Bau des Zwischenkieferbeines (Prämaxillare) zurück. Das Tränenbein (Lacrimale) weist einen in die Augenhöhle vorspringenden Höcker auf; eine Autapomorphie ist der breite Kontakt zwischen Tränenbein und Nasenbein (Nasale). Kennzeichnend ist weiterhin ein insbesondere bei den grasfressenden Arten massiver Kiefer. Das Kiefergelenk liegt hoch und der Unterkieferast ist vergrößert.
Nashörner haben ein oder zwei Hörner, die im Gegensatz zu den Hörnern der Paarhufer nicht aus Knochensubstanz, sondern aus agglutiniertem Keratin bestehen.
Anzahl und Bau der Zähne sind je nach Nahrung unterschiedlich. Schneide- und Eckzähne können sehr klein sein oder komplett fehlen (etwa bei den beiden afrikanischen Nashornarten; bei den Pferden besitzen meist nur männliche Tiere Eckzähne – „Hakenzahn“). Aufgrund des langgestreckten Oberkiefers klafft zwischen den vorderen Zähnen und den Backenzähnen eine als Diastema bezeichnete Lücke. Die Prämolaren (Vorbackenzähne) sind meist molarartig entwickelt; die Oberflächenform und Höhe der Molaren (Hinterbackenzähne) ist stark davon abhängig, ob eher weiches Laub oder hartes Gras den Hauptbestandteil der Nahrung ausmacht. Pro Kieferhälfte sind drei oder vier Prämolaren und stets drei Molaren vorhanden, so dass die Zahnformel der Unpaarhufer lautet: I 0–3/0–3 · C 0–1/0–1 · P 2–4/2–4 · M 3/3.
Innere Anatomie
Im Bau des Verdauungstraktes weisen die Unpaarhufer große Unterschiede zu den ebenfalls meist pflanzenfressenden Paarhufern auf. Unpaarhufer sind – ähnlich den Nagetieren – Enddarmfermentierer, das heißt, dass die Verdauung größtenteils erst im Dickdarm stattfindet. Der Magen ist im Gegensatz zu dem der Paarhufer stets einfach gebaut und einkammerig; die Fermentation findet im sehr großen Blinddarm (der etwa bei Pferden bis zu 90 Liter fasst) und im doppelschlingigen Grimmdarm (Colon) statt. Der Darm ist sehr lang (beim Pferd etwa bis zu 26 Meter). Die Nahrungsausnutzung ist relativ gering, das hat vermutlich dazu geführt, dass es heute keine kleinen Unpaarhufer mehr gibt, da bei großen Tieren der Nahrungsbedarf pro Kilogramm Körpergewicht geringer und das Oberfläche-Volumen-Verhältnis kleiner ist (was besser für den Wärmehaushalt ist).
Im Bereich des Urogenitaltraktes sind die Weibchen ursprünglich durch eine „zweihörnige Gebärmutter“ (Uterus bicornis) gekennzeichnet. Die Eierstöcke (Ovarien) liegen bei Nashörnern und Tapiren in einer Tasche des Bauchfells (Eierstocktasche, Bursa ovarica), bei Pferden bedeckt die Eierstocktasche das Ovar nur teilweise. Pferde unterscheiden sich im Bau des Eierstocks von allen anderen Säugetieren: Das gewöhnlich als „Rinde“ bezeichnete Eierstockgewebe mit den Follikeln liegt bei Pferden im Inneren des Organs, das gefäßführende Eierstockmark dagegen außen. Die Eierstockrinde reicht nur an einer Stelle an die Oberfläche. Diese Stelle ist als Einziehung auch von außen sichtbar und wird als „Ovulationsgrube“ (Fossa ovarii) bezeichnet, nur an dieser Stelle kann der Eisprung (Ovulation) erfolgen. Bei den männlichen Unpaarhufern liegen die Hoden bei Nashörnern und Tapiren inguinal (in der Leistenregion), lediglich Pferde haben ein Skrotum.
Verbreitungsgebiet

Das heutige Verbreitungsgebiet der Unpaarhufer besteht nur noch aus einem kleinen Teil eines einst größeren, nahezu die ganze Erde umfassenden Vorkommens. Wildlebende Vertreter dieser Gruppe finden sich heute in Mittel- und Südamerika, im östlichen und südlichen Afrika sowie im mittleren, südlichen und südöstlichen Asien. Während der Blütezeit der Unpaarhufer vom Eozän bis in das Oligozän erstreckte sich das Verbreitungsgebiet mit großem Formenreichtum über einen Großteil des Globus mit Ausnahme Australiens und Antarktikas, den südamerikanischen Kontinent erreichten sie mit Pferden und Tapiren nach der Bildung des Isthmus von Panama im Pliozän vor rund 3 Millionen Jahren. In Nordamerika starben sie vor rund 10.000 Jahren aus, in Europa mit dem Verschwinden des Tarpans im 19. Jahrhundert. Bejagung und Einschränkung des Lebensraums haben dazu geführt, dass auch die heutigen freilebenden Arten oft nur mehr in zersplitterten Reliktpopulationen vorkommen. Im Gegensatz dazu haben Hauspferde und Hausesel als Nutztiere eine weltweite Verbreitung erlangt, verwilderte Tiere beider Arten kommen mittlerweile auch in Regionen vor, in denen ursprünglich keine Unpaarhufer beheimatet waren, etwa in Australien.
Lebensweise und Ernährung
Je nach Lebensraum führen die verschiedenen Arten der Unpaarhufer eine unterschiedliche Lebensweise. Es sind eher dämmerungs- oder nachtaktive Tiere. Tapire leben einzelgängerisch und bewohnen vorwiegend tropische Regenwälder und andere Wälder. Nashörner leben ebenfalls eher als Einzelgänger und kommen in Afrika eher in trockenen Savannen und in Asien in feuchten Sumpf- oder Waldgebieten vor. Pferde schließlich bewohnen offene Gebiete wie Grasländer, Steppen oder Halbwüsten und leben in Gruppen zusammen. Unpaarhufer sind ausschließlich Pflanzenfresser, die sich in unterschiedlichem Ausmaß von Gräsern, Blättern und anderen Pflanzenteilen ernähren. Man unterscheidet meist zwischen vorwiegend grasfressenden Formen (Breitmaulnashorn, Einhufer) und laubfressenden (Tapire, andere Nashörner).

Fortpflanzung und Entwicklung
Unpaarhufer sind durch eine lange Tragzeit und eine kleine Wurfgröße gekennzeichnet, in der Regel kommt ein einzelnes Jungtier zur Welt. Die Trächtigkeitsdauer liegt zwischen 330 und 500 Tagen, am längsten ist sie bei den Nashörnern. Neugeborene Unpaarhufer sind Nestflüchter, junge Pferde und Nashörner können der Mutter schon nach wenigen Stunden folgen; lediglich Tapirbabys verbringen ihre ersten Lebenstage in einem geschützten Lager.
Die Jungtiere werden relativ lang gesäugt, oft bis ins zweite Lebensjahr hinein und erreichen die Geschlechtsreife mit rund zwei bis acht Jahren. Es sind eher langlebige Tiere, für mehrere Arten ist in menschlicher Obhut ein Alter von knapp 50 Jahren belegt.
Systematik
Äußere Systematik
Innere Systematik der Laurasiatheria und phylogenetische Stellung der Unpaarhufer nach Maureen A. O’Leary et al. 2013[1]
|
Traditionell wurden die Unpaarhufer mit anderen huftragenden Säugetieren wie Paarhufern, Schliefern, Rüsseltieren und anderen als „Huftiere“ (Ungulata) zusammengefasst. Ein nahes Verwandtschaftsverhältnis wurde insbesondere zu den Schliefern vermutet und durch Gemeinsamkeiten im Bau des Ohres, des Verlaufes der Halsschlagader und der Zehen begründet.[2][3][4]
Aufgrund molekulargenetischer Untersuchungen wurden in jüngerer Zeit jedoch erhebliche Zweifel an der Verwandtschaft der Huftiere deutlich, vermutlich stellen diese eine polyphyletische Gruppe dar, das heißt, dass die Ähnlichkeiten nur auf konvergenter Evolution und nicht auf einer gemeinsamen Abstammung beruhen. Elefanten und Schliefer werden heute zumeist in die Überordnung der Afrotheria eingeordnet, sind also nicht näher mit den Unpaarhufern verwandt. Diese wiederum werden in den Laurasiatheria geführt, einer Überordnung, die ihrem Ursprung im Kontinent Laurasia hatte. Der molekulargenetische Befund deutet an, dass das Schwestertaxon der Unpaarhufer die Cetartiodactyla bilden, in denen die Paarhufer (Artiodactyla) und die Wale (Cetacea) enthalten sind; beide Gruppen formen zusammen die Euungulata.[5][1] Weiter außerhalb stehen die Fledertiere (Chiroptera) und die Ferae (ein gemeinsames Taxon aus Raubtieren (Carnivora) und Schuppentieren (Pholidota)).[6] In einem alternativen Szenario besteht auch eine engere Verbindung zwischen den Unpaarhufern, den Raubtieren und den Fledertieren, wobei diese gemeinsame Gruppe dann Pegasoferae bezeichnet wird.[7]
Innere Systematik der Euungulata nach Welker et al. 2015[8]
|
Nach Untersuchungen, die im März 2015 veröffentlicht wurden, stehen die Unpaarhufer in einem nahen Verwandtschaftsverhältnis mit zumindest einigen der sogenannten Meridiungulata („Südamerikanische Huftiere“), einer vom Paläozän bis zum Pleistozän in Südamerika vorkommenden, sehr vielfältigen Gruppe von huftragenden Säugetieren, deren systematische Einheitlichkeit weitgehend ungeklärt[9] ist. Teilweise wurden diese aufgrund ihrer paläogeographischen Verbreitung mit den Afrotheria in Verbindung gebracht, wofür auch einige anatomische Merkmale wie der Bau der Wirbelsäule oder des Sprungbeins sprachen.[10] Allerdings konnte mit Hilfe von Proteinsequenzierungen und dem Vergleich mit fossilem Kollagen, gewonnen an Überresten einiger stammesgeschichtlich junger Vertreter der „Meridiungulata“ (speziell Macrauchenia aus der Gruppe der Litopterna und Toxodon aus der Gruppe der Notoungulata), eine enge Beziehung zu den Unpaarhufern herausgearbeitet werden. Beide Verwandtschaftsgruppen, die Unpaarhufer und die Litopterna-Notoungulata, werden nun in dem übergeordneten Taxon der Panperissodactyla zusammengefasst. Diese Verwandtschaftsgruppe steht dabei innerhalb der Euungulata den Paarhufern und Walen gegenüber (Cetartiodactyla).[8][11] Zwei Jahre später konnte das Ergebnis mit Hilfe von molekulargenetischen Untersuchungen zumindest für die Litopterna-Linie bestätigt werden. Die Trennung dieser von der Linie, die zu den Unpaarhufern führte, erfolgte demnach vor rund 66,15 Millionen Jahren, was in etwa der Kreide-Tertiär-Grenze entspricht.[12] Als Ausgangspunkt für die Entwicklung der beiden Gruppen können vermutlich die „Condylarthra“ in Betracht gezogen werden, welche eine heterogene Gruppe urtümlicher Huftiere darstellen, die im Paläogen hauptsächlich die nördliche Hemisphäre bewohnten.[13] Studien aus dem Jahr 2020 lassen auch auf anatomischer Basis zumindest für die Litopterna ein Schwestergruppenverhältnis zu den Unpaarhufern annehmen.[14] Ebenso führten anatomische Erwägungen dazu, dass im Jahr 2021 in einer Studie die Südamerikanischen Huftiere in zwei Großgruppen, die Panameriungulata und die Sudamericungulata, aufgeteilt wurden. Die Panameriungulata schließen dabei die Litopterna und die Didolodontidae ein und sollen laut den Autoren der Studie, Leonardo S. Avilla und Dimila Mothé, den Unpaarhufern nahestehen. Dem gegenüber vereinen die Sudamericungulata die restlichen Südamerikanischen Huftiere (Notoungulata, Astrapotheria, Pyrotheria und Xenungulata) und werden hierbei als Teilgruppe der Afrotherien angesehen. Die Auffassung steht im Widerspruch zu den erwähnten biochemischen Analysen, worauf Avilla und Mothé hinweisen.[15]
Innere Systematik der Euungulata nach Rose et al. 2020[16]
|
Als weitere übergeordnete Gruppe wurden im Jahr 2020 die Perissodactylamorpha eingeführt. Diese schließen neben den Unpaarhufern auch die Anthracobunia ein, welche wiederum die Anthracobunidae und die Cambaytheriidae beinhalten. Beide Gruppen kamen während des Paläogens in Asien vor. Momentan ist dabei das Verhältnis der Perissodactylamorpha zu den Panperissodactyla unklar, letztere beruhen auf genetischen, erstere auf anatomischen Befunden. Die Autoren vermuten aber, dass die Perissodactylamorpha eine Untergruppe der Panperissodactyla darstellen. In einer eher traditionellen Auffassung werden zumindest die Anthracobunidae als Basalform der Tethytheria eingestuft und damit den Afrotheria zugewiesen.[16]
Rezente Vertreter
Innere Systematik der rezenten Unpaarhufer nach Tougard et al. 2001[17], Steiner und Ryder 2012[18] und Cozzuol et al. 2013[19]
|
Man unterscheidet in der Ordnung der Unpaarhufer (Perissodactyla) drei rezente Familien mit rund 17 Arten – innerhalb der Pferde ist die genaue Artanzahl noch umstritten. Nashörner und Tapire sind dabei näher miteinander verwandt und stehen den Pferden gegenüber. Die Trennung der Pferde von den übrigen Unpaarhufern erfolgte laut molekulargenetischen Analysen im Paläozän vor rund 56 Millionen Jahren, während sich die Nashörner und Tapire im Unteren Mittleren Eozän vor etwa 47 Millionen Jahren aufspalteten.[17][18]
- Ordnung: Unpaarhufer (Perissodactyla Owen, 1848)
- Familie: Pferde (Equidae Linnaeus, 1758)
- Tarpan (Equus ferus Boddaert, 1785)
- Przewalski-Pferd (Equus przewalskii Poljakov, 1881)
- Afrikanischer Esel, auch Wildesel oder Echter Esel (Equus asinus Linnaeus, 1758)
- Asiatischer Esel, auch Halbesel oder Pferdeesel (Equus hemionus Pallas, 1775)
- Kiang (Equus kiang Moorcroft, 1841)
- Steppenzebra (Equus quagga Boddaert, 1785)
- Bergzebra (Equus zebra Linnaeus, 1758)
- Grevyzebra (Equus grevyi Oustalet, 1882)
- Familie: Tapire (Tapiridae Gray, 1821)
- Flachlandtapir (Tapirus terrestris (Linnaeus, 1758))
- Bergtapir (Tapirus pinchaque (Roulin, 1829))
- Kabomani-Tapir (Tapirus kabomani Cozzuol, Clozato, Holanda, Rodrigues, Nienow, de Thoisy, Redondo & Santos, 2013)
- Mittelamerikanischer Tapir (Tapirus bairdii (Gill, 1865))
- Schabrackentapir (Tapirus indicus Desmarest, 1819)
- Familie: Nashörner (Rhinocerotidae Owen, 1845)
- Breitmaulnashorn (Ceratotherium simum (Burchell, 1817))
- Spitzmaulnashorn (Diceros bicornis (Linnaeus, 1758))
- Sumatra-Nashorn (Dicerorhinus sumatrensis (Fischer, 1814))
- Panzernashorn (Rhinoceros unicornis Linnaeus, 1758)
- Java-Nashorn (Rhinoceros sondaicus Desmarest, 1822)
- Familie: Pferde (Equidae Linnaeus, 1758)
Das Hauspferd wurde aus einem wildlebenden Vorläufer domestiziert, der Hausesel stammt vom Afrikanischen Esel ab. In manchen Systematiken gelten der Khur (Equus khur) und der Achdari (Equus hemippus) sowie das Hartmann-Bergzebra (Equus hartmannae) und das Nördliche Breitmaulnashorn (Ceratotherium cottoni) als eigenständige Arten.[20]
Fossile Vertreter
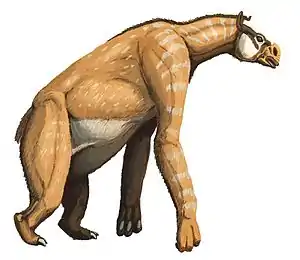
Fossil traten Unpaarhufer in einer hohen Zahl mit variantenreichen Formen auf, zu den wichtigsten Entwicklungslinien zählen folgende Gruppen:[21]
- Die Brontotherioidea gehörten zu den frühesten bekannten Großsäugern, bestehend aus den Familien der Brontotheriidae (synonym Titanotheriidae), deren bekanntester Vertreter Megacerops ist, und der eher basalen Gruppe der Lambdotheriidae. Sie waren vor allem in ihrer Spätphase durch ein knöchernes Horn am Übergang vom Nasen- zum Stirnbein und generell durch flache, für weiche Pflanzennahrung geeignete Backenzähne charakterisiert. Am Beginn des Oberen Eozäns starben die Brontotheroidea, die fast ausschließlich auf Nordamerika und Asien beschränkt waren, aus.
- Die Equoidea (Pferdeartige) entwickelten sich ebenfalls im Eozän. Die Palaeotheriidae, die vor allem aus Europa bekannt sind und deren bekanntester Vertreter Hyracotherium war, sind im Oligozän ausgestorben. Die (eigentlichen) Pferde (Equidae) hingegen florierten und breiteten sich aus. Bei der Entwicklung dieser Gruppe ist die Reduktion der Zehenanzahl, die Verlängerung der Gliedmaßen und die fortschreitende Anpassung der Zähne an harte Grasnahrung anhand von Fossilienfunden gut zu beobachten.
- Die Chalicotherioidea stellten eine weitere charakteristische Gruppe dar, die aus den Familien der Chalicotheriidae und der Lophiodontidae bestand. Innerhalb der Chalicotheriidae kam es zur Entwicklung von Klauen statt Hufen und zu einer drastischen Verlängerung der Vorderbeine. Die bekanntesten Gattungen sind unter anderem Chalicotherium und Moropus. Die Chalicotherioidea starben erst im Pleistozän aus.
- Die Rhinocerotoidea (Nashornverwandte) kamen vom Eozän bis in das Oligozän mit einem großen Formenreichtum vor, es gab hundsgroße Blätterfresser, semiaquatische (teilweise im Wasser lebende) Tiere und auch riesige, langhalsige Tiere – Hörner auf der Nase hatten die wenigsten davon. Die eigentlichen Nashörner (Rhinocerotidae) entstanden im Mittleren Eozän, fünf Arten überleben bis auf den heutigen Tag. Daneben sind noch mehrere fossile Familien belegt, die weitgehend im Eozän und Oligozän präsent waren. In einer eher klassischen Sichtweise werden die Amynodontidae, flusspferdähnliche, im Wasser lebende Tiere, und die eher langbeinigen Hyracodontidae unterschieden. Letztere entwickelten extrem große, langhälsige Formen wie etwa Paraceratherium (früher auch als Baluchitherium oder Indricotherium bekannt), das größte bekannte Landsäugetier. Da sich die Hyracodontidae aber als „Abfalleimertaxon“ erwiesen, gibt es in jüngerer Zeit verschiedene Versuche einer neuen Gliederung. Teilweise werden die Indricotheriidae (auch Paraceratheriidae) und die Eggysodontidae aus den Hyracodontidae ausgegliedert und von ersteren wiederum die Forstercooperiidae abgespalten. Dadurch stehen die nun eigentlichen Hyracodontidae eher basal innerhalb der Rhinocerotoidea.[22][23]
- Die Tapiroidea (Tapirartige) erreichten ihre größte Vielfalt im Eozän, als mehrere Gattungen in Eurasien und Nordamerika beheimatet waren. Sie behielten am ehesten einen urtümlichen Körperbau bei, bemerkenswert ist lediglich die Entwicklung eines Rüssels. Zu den ausgestorbenen Familien zählen unter anderem die Helaletidae, die recht formenreich in Nordamerika und Eurasien auftraten, bekannte Gattungen sind hier Helaletes, Heptodon, Colodon oder Desmatotherium. Neben dieser basalen Gruppe waren die Deperetellidae eher auf Eurasien beschränkt. Sie umfassen Formen wie Deperetella, Teleolophus und Irenolophus. Möglicherweise sind auch die Lophialetidae in die Tapiroidea einzuordnen, bei denen es sich um eine asiatisch verbreitete Gruppe urtümlicher Unpaarhufer handelt. Andererseits können sie auch als eher basale Formengruppe der Nashorn-Tapir-Verwandtschaftsgemeinschaft aufgefasst werden.[24][25][26][23]
Höhere Systematik der Unpaarhufer
Innere Systematik der Perissodactyla nach Holbrook und Lapergola 2011[27]
|
Innere Systematik der Perissodactyla nach Hooker und Dashzeveg 2004[28]
|
Die Beziehungen der Großgruppen der Unpaarhufer untereinander sind noch nicht vollständig geklärt. Anfänglich nach der Etablierung des Begriffes Perissodactyla durch Richard Owen im Jahr 1848 wurden die heutigen Vertreter als gleichrangig eingestuft. In der ersten Hälfte des 20. Jahrhunderts begann unter Einbeziehung fossiler Formen eine stärkere systematische Differenzierung der Unpaarhufer, wodurch diese in die beiden großen Unterordnungen der Hippomorpha und der Ceratomorpha unterschieden wurden. Dabei umfassen die Hippomorpha nach gegenwärtiger Auffassung die heutigen Pferde und deren fossilen Vertreter (Equoidea), die Ceratomorpha dagegen die Tapire und Nashörner zuzüglich ihrer ausgestorbenen, weiterläufigen Verwandtschaft (Tapiroidea und Rhinocerotoidea).[29] Die Bezeichnungen Hippomorpha und Ceratomorpha wurden 1937 von Horace Elmer Wood eingeführt, wobei er damit auf Kritik reagierte, die nach seinen drei Jahre zuvor vorgeschlagenen Namen Solidungula für die Verwandtschaftsgruppe der Pferde und Tridactyla für den Nashorn-Tapir-Komplex aufgekommen war.[30][31] Die ausgestorbenen Brontotheriidae wurden von Wood ebenfalls zu den Hippomorpha gestellt und besäßen demnach eine nahe Verwandtschaft zu den Pferden. Einige Forscher sehen diese Zuweisung aufgrund ähnlicher Zahnmerkmale heute noch gerechtfertigt, es gibt aber auch die Ansicht einer sehr basalen Stellung innerhalb der Unpaarhufer, wobei sie dann der Gruppe der Titanotheriomorpha angehören.[28][27][3]
Ursprünglich wurden von Wood auch die Chalicotheriidae als Mitglieder der Hippomorpha gesehen, im Jahr 1941 stellte William Berryman Scott sie aber als klauentragende Unpaarhufer den huftragenden gegenüber und verwies sie in die neue Unterordnung Ancylopoda (wobei er die Ceratomorpha und Hippomorpha als huftragende Unpaarhufer in der Gruppe der Chelopoda vereinte).[32] Der Begriff Ancylopoda war bereits 1889 von Edward Drinker Cope für die Chalicotherien etabliert worden. Weitere morphologische Untersuchungen aus den 1960er Jahren ergaben jedoch eine Mittelstellung der Ancylopoda zwischen den Hippomorpha und Ceratomorpha. Leonard Burton Radinsky sah alle drei Großgruppen der Unpaarhufer als gleichrangig an, was er unter anderem mit der extrem langen und unabhängigen stammesgeschichtlichen Entwicklung der drei Linien begründete.[33] In den 1980er Jahren erkannte Jeremy J. Hooker im Zahnbau eine generelle Ähnlichkeit der Ancylopoda zu den Ceratomorpha, vor allem bei den frühesten Vertretern, was 1984 zur Vereinigung der beiden Unterordnungen in der Zwischenordnung Tapiromorpha führte (gleichzeitig erweiterte er die Ancylopoda um die Lophiodontidae). Der Name Tapiromorpha wiederum geht auf Ernst Haeckel zurück, der ihn 1873 prägte; er galt aber lange Zeit als synonym zu Ceratomorpha, da Wood ihn 1937 bei der Etablierung der Ceratomorpha aufgrund seiner stark unterschiedlichen Verwendung in der Vergangenheit nicht beachtet hatte.[34][31] Ebenfalls im Jahr 1984 benutzte Robert M. Schoch den konzeptionell ähnlichen Begriff Moropomorpha, der heute als synonym zu Tapiromorpha gilt.[35] Innerhalb der Tapiromorpha stehen die heute ausgestorbenen Isectolophidae als Schwestergruppe der Ancylopoda-Ceratomorpha-Gruppe gegenüber und sind somit als die ursprünglichsten Vertreter dieses Verwandtschaftskomplexes anzusehen.[36][3]
Neben den hier weitgehend anerkannten Großgruppen der Unpaarhufer bestehen noch weitere höhere Taxa, die je nach verwendeter systematischer Aufteilungen der Ordnung Bedeutung haben, in der Regel aber nur selten gebraucht werden. Jerry J. Hooker und Demberelyin Dashzeveg führten im Jahr 2004 die Lophodontomorpha ein, die alle Unpaarhufergruppen mit Ausnahme der Brontotherien und ihre unmittelbare Nahverwandtschaft einschließen. Auf beide Autoren gehen auch die Euperissodactyla zurück, die wiederum die Ceratomorpha und die Hippomorpha vereinen.[28] Dagegen propagierten Malcolm C. McKenna und Susan K. Bell im Jahr 1997 in ihrem Übersichtswerk zur Gliederung der Säugetiere die Selenida, in denen sie die Brontotherien und die Chalicotherien nebst ihrer Verwandtschaft aufnahmen.[37]
Entwicklungsgeschichte
Ursprünge
Die Entwicklungsgeschichte der Unpaarhufer ist vergleichsweise gut durch Fossilien überliefert; zahlreiche Funde lassen die Radiation dieser früher viel formenreicheren und weiter verbreiteten Gruppe erkennen. Die Ursprünge der Unpaarhufer sind ungeklärt, teilweise wird eine Entstehung in Nordamerika angenommen, was mit der rapiden Entwicklung der Phenacodontidae einhergehen würde.[38] Andererseits könnte auch Asien als eigentliches Evolutionszentrum angesehen werden. Als einer der ältesten nahen Verwandten der Unpaarhufer wird teilweise Radinskya aus dem ausgehenden Paläozän Ostasiens angesehen.[39][40] Der nur etwas mehr als 8 cm messende Schädel gehört einem sehr kleinen und ursprünglichen Tier an, das mit der leicht π-förmigen Gestaltung des Zahnschmelzes auf den hinteren Backenzähnen Ähnlichkeiten zu Unpaarhufern, vor allem den Nashörnern und ihren Verwandten zeigt. Andere Merkmale wie die Ausbildung der Zahnhöckerchen könnten aber auch für eine engere Beziehung mit den Tethytheria sprechen, zu denen etwa die Rüsseltiere zählen.[41] Funde aus dem westlichen Indien unterstützen die Ansicht eines asiatischen Ursprungs und grenzen das Entstehungszentrum auf Südasien ein. Aus der Cambay-Shale-Formation, die ins Untere Eozän vor rund 54,5 Millionen Jahre datiert, stammen Überreste von Cambaytherium, welches zur Familie der Cambaytheriidae gehört.[42][43] Deren Zähne zeigen vergleichbar zu Radinskya Ähnlichkeiten zu den frühen Unpaarhufern und Tethytherien.[44][45] Von Cambaytherium liegt auch umfangreiches Material des Körperskeletts vor. Hier sprechen unter anderem der sattelförmige Einzug des Naviculargelenks am unteren Ende des Sprungbeins und die mesaxonische Gestaltung der Vorder- und Hinterfüße – die Hauptachse des Fußes ging durch den Mittelstrahl (Strahl III) – für eine nahe Verwandtschaft mit den Unpaarhufern. Da die Füße aber abweichend von den frühesten Unpaarhufern noch fünfstrahlig waren, werden die Cambaytherien heute als deren Schwestergruppe betrachtet. Möglicherweise gelangten Vorfahren der Unpaarhufer über eine Inselbrücke von der Afroarabischen Landmasse auf den Indischen Subkontinent, der damals eine Insel war und nach Norden Richtung Asien driftete.[46] Aus der gleichen Formation wurde mit Vastanolophus einer der urtümlichsten bekannten Vertreter der Unpaarhufer beschrieben, der allerdings bisher nur mit zwei Zähnen belegt ist.[47]
Stammesgeschichte

Die Unpaarhufer erscheinen relativ unvermittelt zu Beginn des Unteren Eozäns vor rund 56 Millionen Jahren sowohl in Nordamerika als auch in Asien. Die ältesten nordamerikanischen Funde stammen unter anderem von Sifrhippus, einem Vertreter der Pferde aus der Willwood-Formation im nordwestlichen Wyoming.[48][49] Etwa zeitgleich ist in Asien aus der Lingcha-Formation im nordwestlichen China mit Erihippus ebenfalls ein Verwandter der Pferde belegt. Zusätzlich kommen hier mit Orientolophus ein Vertreter der Stammgruppe der Nashorn-Tapir-Linie, mit Protomoropus eine frühe Form der Ancylopoda und mit Danjiangia ein erster Angehöriger der Brontotheriidae vor.[50] Nur wenig später treten in Asien weitere entfernte Vorfahren der Tapire und Nashörner auf wie beispielsweise Ganderalophus oder Meridiolophus, daneben auch Litolophus, das in der Entwicklungslinie der Chalicotheriidae steht. In Nordamerika ist wiederum Eotitanops aus der Gruppe der Brontotheriidae belegt.[51][52][53] Somit sind im Verlauf des Unteren Eozän die wichtigsten Großgruppen durch urtümliche Vertreter nachweisbar. Anfangs sahen sich die Angehörigen der verschiedenen Linien noch recht ähnlich mit einem aufgewölbten Rücken und generell vier Zehen an den Vorder- und drei an den Hinterfüßen. Hyracotherium, das als Vertreter der Pferdeverwandten gilt, ähnelte äußerlich beispielsweise sehr Hyrachyus, dem ersten Vertreter der Linie, die zu den heutigen Nashörnern und Tapiren führte.[54] Alle waren im Vergleich zu späteren Formen relativ klein und lebten als Frucht- und Laubfresser in Wäldern. Erste Riesenformen entstanden mit den Brontotherien bereits im Mittleren und Oberen Eozän, wobei das bekannte Megacerops aus Nordamerika bis 2,5 m Schulterhöhe erreichte und gut 3 t gewogen haben könnte. Der Niedergang der Brontotherien am Ausgang des Eozäns steht dabei im Zusammenhang mit dem Aufkommen konkurrenzfähigerer Pflanzenfresser.[21][55]
Vor allem am Ende des Eozäns entstanden weitere erfolgreiche Linien der Unpaarhufer, etwa die Chalicotheriidae und die Nashörner und ihre näheren Verwandten, deren Entwicklung ebenfalls mit sehr kleinen Formen begann. Die Hyracodontidae, die nächsten fossilen Verwandten der Nashörner, stellten dabei mit Paraceratherium das größte landlebende Säugetier aller Zeiten. Dieses wog bis zu 20 t und lebte im Oligozän in Eurasien.[56] Andere florierende Gruppen jener Zeit waren die Amynodontidae mit Amynodon, Cadurcodon oder Sellamynodon, darüber hinaus auch die Eggysodontidae.[57][58][22] Mit dem Beginn des Miozäns erreichten die Unpaarhufer vor etwa 20 Millionen Jahren erstmals Afrika, das durch die Schließung der Tethys nun mit Eurasien verbunden war. Allerdings gelangten durch den nun folgenden Faunenaustausch neue Tiergruppen in die alten Besiedlungsgebiete der Unpaarhufer, so etwa die Rüsseltiere, deren Konkurrenz auch zum Aussterben einiger Unpaarhufer-Linien führte. Auch der Aufstieg der Wiederkäuer, die ähnliche ökologische Nischen besetzten und deren Verdauungssystem als effizienter gilt, wird mit dem Rückgang der Diversität der Unpaarhufer in Verbindung gebracht. Einen erheblichen Anteil am Niedergang der Unpaarhufer hatten aber klimatische Veränderungen im Verlauf des Miozäns hin zu kühlerem und trockenem Klima, das mit der Ausbreitung offener Landschaften einherging. Allerdings florierten auch einige Linien, etwa jene der Pferde und Nashörner, indem sich zahlreiche Vertreter durch anatomische Modifikationen an die härtere Grasnahrung anpassten. Dadurch entstanden Offenlandformen, die die neu entstandenen Landschaftstypen besiedelten. Mit der Entstehung des Isthmus von Panama im Pliozän und dem anschließenden Großen Amerikanischen Faunenaustausch erreichten die Unpaarhufer, hier die Pferde und Tapire, vor rund 3 Millionen Jahren mit Südamerika einen der letzten für sie zugänglichen Lebensräume.[59][60] Allerdings waren auch die Unpaarhufer vom Aussterben der Großsäuger am Ende des Pleistozäns betroffen, so verschwanden zu dieser Zeit unter anderem die Pferde Nord- und Südamerikas sowie das Wollnashorn. Ob die Bejagung durch den Menschen (Overkill-Hypothese), klimatische Veränderungen, die mit dem Ende der Eiszeit einhergingen, oder eine Kombination beider Faktoren für dieses Aussterben verantwortlich war, ist umstritten.[21]
Forschungsgeschichte

Die Erkenntnis, dass die Unpaarhufer eine systematische Einheit bilden, reicht bis in die Mitte des 19. Jahrhunderts zurück. Davor wurden die heutigen Vertreter der Unpaarhufer meist anderen Gruppen zugeordnet. Linnaeus (1707–1778) stellte 1758 in seinem wegweisenden Werk Systema naturae die Pferde (Equus) an die Seite der Flusspferde (Hippopotamus). Diese enthielten damals auch die Tapire (Tapirus), genauer den Flachlandtapir, der zu jener Zeit in Europa die einzige bekannte Tapirart war und den Linnaeus als Hippopotamus terrestris klassifizierte. Beide Gattungen verwies Linnaeus zur Gruppe der Belluae. Die Nashörner (Rhinoceros) vereinte er dagegen mit den Glires, einer Gruppe heute bestehend aus den Hasenartigen und den Nagetieren. Erst Mathurin-Jacques Brisson (1723–1806) trennte 1762 mit der Einführung des Begriffes le tapir die Tapire von den Flusspferden, zudem gliederte er die Nashörner aus den Nagern aus, vereinte aber die drei heutigen Familien der Unpaarhufer nicht. Im Übergang zum 19. Jahrhundert wurden die einzelnen Unpaarhufer-Gattungen mit verschiedenen anderen Gruppen, so den Rüsseltieren oder heutigen Paarhufern assoziiert, in jene Zeit fällt auch die Etablierung des Begriffes „Dickhäuter“ (Pachydermata), den Étienne Geoffroy Saint-Hilaire (1772–1844) und Georges Cuvier (1769–1832) im Jahr 1795 als Pachydermes einführten[61] (und der neben den Nashörnern und Elefanten auch die Flusspferde, Schweine, Pekaris, Tapire und Schliefer einbezog).[62] Die Pferde galten aber weitgehend als von den anderen Säugetieren abgetrennte Gruppe und wurden häufig unter der Bezeichnung Solidungula[63] oder Solipèdes[61][64] geführt, was beides so viel wie „Einhufer“ bedeutet.[29][3]
Im Jahr 1816 war es Henri Marie Ducrotay de Blainville (1777–1850), der die Huftiere nach dem Aufbau der Füße gliederte und so Tiere mit einer geraden von solchen mit einer ungeraden Anzahl von Zehen unterschied. Er schob die Pferde als solidungulate in die Nähe der Tapire und Nashörner als multungulate Tiere und bezeichnete alle zusammen als onguligrades à doigts impairs, was dem Konzept der Unpaarhufer als systematische Einheit sehr nahekam.[65] Richard Owen (1804–1892) berief sich in seiner Studie zu fossilen Säugetieren der Isle of Wight auf Blainville und führte den Namen Perissodactyla ein. Wie Blainville schloss er in diese aber neben den Pferden, Nashörnern und Tapiren auch die Schliefer mit ein.[66] In den folgenden Jahren verwies Owen noch weitere, heute ausgestorbene Formen den Unpaarhufern zu, so etwa Coryphodon, Toxodon oder Nesodon, die heute aber allesamt anderen Säugetierordnungen angehören.[29][3]
Othniel Charles Marsh (1831–1899) führte im Jahr 1884 den Begriff Mesaxonia ein. Diesen verwendete er für die heutigen Angehörigen der Unpaarhufer einschließlich deren ausgestorbenen Verwandten, schloss die Schliefer aber ausdrücklich aus.[67] Mesaxonia gilt heute als Synonym zu Perissodactyla. Teilweise wurde er aber für die eigentlichen Unpaarhufer als Unterordnungsbegriff verwendet (Nashörner, Pferde, Tapire), während Perissodactyla als Bezeichnung für die gesamte Ordnung einschließlich der Schliefer stand. Die Annahme, die Schliefer seien ein Teil der Unpaarhufer hielt sich noch bis weit in das 20. Jahrhundert.[68] Erst mit dem Aufkommen molekulargenetischer Untersuchungsmethoden wurde erkannt, dass die Schliefer nicht mit den Unpaarhufern näher verwandt sind, sondern mit den Rüsseltieren und Seekühen in Beziehung stehen.[5][69]
Unpaarhufer und Menschen
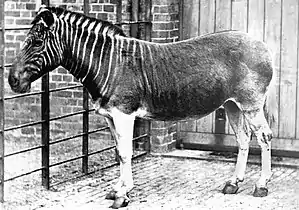
Das Hauspferd und der Hausesel spielen insbesondere als Reit-, Arbeits- und Lasttiere eine wichtige Rolle in der menschlichen Geschichte, die Domestikation beider Arten begann bereits mehrere Jahrtausende vor Christi Geburt. Aufgrund der Motorisierung der Landwirtschaft und der Verbreitung des Automobilverkehrs ist dieser Verwendungszweck in den westlichen Industrieländern stark zurückgegangen, das Reiten wird meist nur mehr als Hobby oder Sport betrieben. In den wenig entwickelten Regionen der Erde ist der Einsatz dieser Tiere aber immer noch weit verbreitet. In geringerem Ausmaß werden Pferde und Esel auch wegen ihres Fleisches und ihrer Milch gehalten.
Im Gegensatz dazu sind die Bestände fast aller übrigen Arten der Unpaarhufer durch Bejagung und Zerstörung des Lebensraumes drastisch zurückgegangen. Das Quagga ist ausgestorben, das Przewalski-Pferd gilt als in freier Wildbahn ausgerottet.
Gegenwärtige Gefährdungsstufen laut IUCN (Stand 2012):[70]
- Vier Arten sind vom Aussterben bedroht (critically endangered): das Java-Nashorn, das Sumatra-Nashorn, das Spitzmaulnashorn und der Afrikanische Esel.
- Sechs Arten sind stark gefährdet (endangered): der Bergtapir, der Mittelamerikanische Tapir, der Schabrackentapir, das Wildpferd, der Asiatische Esel und das Grevyzebra.
- Drei Arten sind gefährdet (vulnerable): das Panzernashorn, der Flachlandtapir und das Bergzebra.
- Potenziell gefährdet (near threatened) ist gegenwärtig das Breitmaulnashorn, allerdings steht die nördliche Unterart Ceratotherium simum cottoni (Nördliches Breitmaulnashorn) kurz vor der Ausrottung.
- Nicht gefährdet (least concern) sind das Steppenzebra und der Kiang.
Literatur
- Martin S. Fischer: Mesaxonia (Perissodactyla), Unpaarhufer. In: Wilfried Westheide, Reinhard Rieger (Hrsg.): Spezielle Zoologie. Teil 2: Wirbel- oder Schädeltiere. Spektrum Akademischer Verlag, Heidelberg und Berlin 2004, S. 646–655, ISBN 3-8274-0307-3.
- Ronald M. Nowak: Walker’s mammals of the world. 6. Auflage. Johns Hopkins University Press, Baltimore 1999, ISBN 0-8018-5789-9 (englisch).
- Thomas S. Kemp: The Origin & Evolution of Mammals. Oxford University Press, Oxford 2005. ISBN 0-19-850761-5.
- A. H. Müller: Lehrbuch der Paläozoologie, Band III: Vertebraten, Teil 3: Mammalia. 2. Auflage. Gustav Fischer Verlag, Jena und Stuttgart 1989. ISBN 3-334-00223-3.
- Don E. Wilson, DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. 3. Ausgabe. The Johns Hopkins University Press, Baltimore 2005, ISBN 0-8018-8221-4.
Einzelnachweise
- Maureen A. O’Leary, Jonathan I. Bloch, John J. Flynn, Timothy J. Gaudin, Andres Giallombardo, Norberto P. Giannini, Suzann L. Goldberg, Brian P. Kraatz, Zhe-Xi Luo, Jin Meng, Xijun Ni, Michael J. Novacek, Fernando A. Perini, Zachary S. Randall, Guillermo W. Rougier, Eric J. Sargis, Mary T. Silcox, Nancy B. Simmons, Michelle Spaulding, Paúl M. Velazco, Marcelo Weksler, John R. Wible und Andrea L. Cirranello: The Placental Mammal Ancestor and the Post–K-Pg Radiation of Placentals. Science 339 (6120), 2013, S. 662–667, doi:10.1126/science.1229237
- D. Tab Rasmussen, Mario Gagnon und Elwyn L. Simons: Taxeopody in the carpus and tarsus of Oligocene Pliohyracidae (Mammalia: Hyracoidea) and the phyletic position of Hyraxes. PNAS 87, 1990, S. 4688–4691
- Robert M. Schoch: A brief historical review of Perissodactyl classification. In: Donald R. Prothero und Robert M. Schoch (Hrsg.): The evolution of perissodactyls. New York und London 1989, S. 13–23
- Martin S. Fischer: Hyracoids, the sister-group of perissodactyls. In: Donald R. Prothero und Robert M. Schoch (Hrsg.): The evolution of perissodactyls. New York und London 1989, S. 37–56
- Dan Graur, Manolo Gouy und Laurent Duret: Evolutionary Affinities of the Order Perissodactyla and the Phylogenetic Status of the Superordinal Taxa Ungulata and Altungulata. Molecular Phylogenetics and Evolution 7 (2), 1997, S. 195–200
- Jingyang Hu, Yaping Zhang und Li Yu: Summary of Laurasiatheria (Mammalia) Phylogeny. Zoological Research 33 (E5–6), 2012, S. E65–E74
- Hidenori Nishihara, Masami Hasegawa und Norihiro Okada: Pegasoferae, an unexpected mammalian clade revealed by tracking ancient retroposon insertions. PNAS 103, 2006; S. 9929–9934 (Volltext, PDF verfügbar)
- Frido Welker, Matthew J. Collins, Jessica A. Thomas, Marc Wadsley, Selina Brace, Enrico Cappellini, Samuel T. Turvey, Marcelo Reguero, Javier N. Gelfo, Alejandro Kramarz, Joachim Burger, Jane Thomas-Oates, David A. Ashford, Peter D. Ashton, Keri Rowsell, Duncan M. Porter, Benedikt Kessler, Roman Fischer, Carsten Baessmann, Stephanie Kaspar, Jesper V. Olsen, Patrick Kiley, James A. Elliott, Christian D. Kelstrup, Victoria Mullin, Michael Hofreiter, Eske Willerslev, Jean-Jacques Hublin, Ludovic Orlando, Ian Barnes und Ross D. E. MacPhee: Ancient proteins resolve the evolutionary history of Darwin’s South American ungulates. Nature 522, 2015, S. 81–84, doi:10.1038/nature14249
- Richard A. Fariña, Sergio F. Vizcaíno und Gerardo De Iuliis: Megafauna. Giant beasts of Pleistocene South America. Indiana University Press, 2013, S. 1–436 (S. 86–87) ISBN 978-0-253-00230-3
- Federico L. Agnolin und Nicolás R. Chimento: Afrotherian affinities for endemic South American “ungulates”. Mammalian Biology 76, 2011, S. 101–108
- Michael Buckley: Ancient collagen reveals evolutionary history of the endemic South American ‘ungulates’. Proceedings of the Royal Society B 282, 2015, S. 20142671, doi:10.1098/rspb.2014.2671
- Michael Westbury, Sina Baleka, Axel Barlow, Stefanie Hartmann, Johanna L. A. Paijmans, Alejandro Kramarz, Analıá M. Forasiepi, Mariano Bond, Javier N. Gelfo, Marcelo A. Reguero, Patricio López-Mendoza, Matias Taglioretti, Fernando Scaglia, Andrés Rinderknecht, Washington Jones, Francisco Mena, Guillaume Billet, Christian de Muizon, José Luis Aguilar, Ross D. E. MacPhee und Michael Hofreiter: A mitogenomic timetree for Darwin’s enigmatic South American mammal Macrauchenia patachonica. Nature Communications 8, 2017, S. 15951, doi:10.1038/ncomms15951
- Ross MacPhee, Frido Welker, Jessica Thomas, Selina Brace, Enrico Cappellini, Samuel Turvey, Ian Barnes, Marcelo Reguero, Javier Gelfo und Alejandro Kramarz: Ancient protein sequencing resolves litoptern and notoungulate superordinal affinities. In: Esperanza Cerdeño (Hrsg.): 4th International Palaeontological Congress. The History of Life: A view from Southern Hemisphere. September 28 – October 3, 2014 Mendoza, Argentina. Mendoza, 2014, S. 186
- Nicolás R. Chimento und Federico L. Agnolin: Phylogenetic tree of Litopterna and perissodactyla indicates a complex early history of hoofed mammals. Scientific Reports 10, 2020, S. 13280, doi:10.1038/s41598-020-70287-5
- Leonardo S. Avilla und Dimila Mothé: Out of Africa: A New Afrotheria Lineage Rises From Extinct South American Mammals. Frontiers in Ecology and Evolution 9, 2021, S. 654302, doi:10.3389/fevo.2021.654302
- Kenneth D. Rose, Luke T. Holbrook, Kishor Kumar, Rajendra S. Rana, Heather E. Ahrens, Rachel H. Dunn, Annelise Folie, Katrina E. Jones und Thierry Smith: Anatomy, Relationships, and Paleobiology of Cambaytherium (Mammalia, Perissodactylamorpha, Anthracobunia) from the lower Eocene of western India. Journal of Vertebrate Paleontology 39 (suppl.), 2020, S. 1–147, doi:10.1080/02724634.2020.1761370
- Christelle Tougard, Thomas Delefosse, Catherine Hänni und Claudine Montgelard: Phylogenetic Relationships of the Five Extant Rhinoceros Species (Rhinocerotidae, Perissodactyla) Based on Mitochondrial Cytochrome b and 12S rRNA Genes. Molecular Phylogenetics and Evolution 19, 2001, S. 34–44
- Cynthia C. Steiner und Oliver A. Ryder: Molecular phylogeny and evolution of the Perissodactyla. Zoological Journal of the Linnean Society 163, 2011, S. 1289–1303
- Mario A. Cozzuol, Camila L. Clozato, Elizete C. Holanda, Flávio H. G. Rodrigues, Samuel Nienow, Benoit de Thoisy, Rodrigo A. F. Redondo und Fabrício R. Santos: A new species of tapir from the Amazon. Journal of Mammalogy 94 (6), 2013, S. 1331–1345 ()
- Colin P. Groves und Peter Grubb: Ungulate Taxonomy. Johns Hopkins University Press, 2011, S. 1–317 (S. 16)
- Donald R. Prothero: Evolutionary Transitions in the Fossil Record of Terrestrial Hoofed Mammals. Evo Edu Outreach 2, 2009, S. 289–302
- Haibing Wang, Bin Bai, Jin Meng und Yuanqing Wang: Earliest known unequivocal rhinocerotoid sheds new light on the origin of Giant Rhinos and phylogeny of early rhinocerotoids. Scientific Reports 6, 2016, S. 39607, doi:10.1038/srep39607
- Bin Bai, Jin Meng, Chi Zhang, Yan-Zin Gong und Yuan-Qing Wang: The origin of Rhinocerotoidea and the phylogeny of Ceratomorpha (Mammalia, Perissodactyla). Communications Biology 3, 2020, S. 509, doi:10.1038/s42003-020-01205-8
- Leonard B. Radinsky: Early Tertiary Tapiroidea of Asia. Bulletin of the American Museum of Natural History 129 (2), 1965, S. 183–263
- Bin Bai, Yuan-Qing Wang, Fang-Yuan Mao und Jin Meng: New material of Eocene Helaletidae (Perissodactyla, Tapiroidea) from the Irdin Manha Formation of the Erlian Basin, Inner Mongolia, China and comments on Related Localities of the Huheboerhe Area. American Museum Novitates 3878, 2017, S. 1–44
- Bin Bai, Jin Meng, Fang-Yuan Mao, Zhao-Qun Zhang und Yuan Qing Wang: A new early Eocene deperetellid tapiroid illuminates the origin of Deperetellidae and the pattern of premolar molarization in Perissodactyla. PLoS ONE 14 (11), 2019, S. e0225045, doi:10.1371/journal.pone.0225045
- Luke T. Holbrook und Joshua Lapergola: A new genus of Perissodactyl (Mammalia) from the Bridgerian of Wyoming, with comments on basal Perissodactyl phylogeny. Journal of Vertebrate Paleontology 31 (4), 2011, S. 895–901
- Jerry J. Hooker und Demberelyin Dashzeveg: The origin of chalicotheres (Perissodactyla, Mammalia). Palaeontology 47 (6), 2004, S. 1363–1386
- George Gaylord Simpson: The Principles of Classification and a Classification of Mammals. Bulletin of the American Museum of Natural History 85, 1945, S. 1–350 (S. 252–258)
- Horace Elmer Wood: Revision of the Hyrachyidae. Bulletin of the American Museum of Natural History 67, 1934, S. 181–295 (S. 260)
- Horace Elmer Wood: Perissodactyl suborders. Journal of Mammalogy 18 (1), 1937, S. 106
- William Berryman Scott: Part V: Perissodactyla. In: William Berryman Scott und Glenn Lowell Jepsen (Hrsg.): The Mammalian Fauna of the White River Oligocene. Transactions of the American Philosophical Society New Series 28 (5), 1941, S. 747–964 (S. 747)
- Leonard B. Radinsky: Paleomoropus, a new early Eocene chalicothere (Mammalia, Perissodactyla), and a revision of Eocene chalicotheres. American Museum Nouitates 2179, 1964, S. 1–28
- J. J. Hooker: A primitive ceratomorph (Perissodactyla, Mammalia) from the early Tertiary of Europe. Zoological Journal of the Linnean Society of London 82, 1984, S. 229–244
- Robert Milton Schoch: Two unusual specimens of Helaletes in the Yale Peabody Museum collections, and some comments on the ancestry of the Tapiridae (Perissodactyla, Mammalia). Postilla, Peabody Museum, Yale University 193, 1984, S. 1–20
- Luke T. Holbrook: Comparative osteology of early Tertiary tapiromorphs (Mammalia, Perissodactyla). Zoological Journal of the Linnean Society 132, 2001, S. 1–54
- Malcolm C. McKenna und Susan K. Bell: Classification of mammals above the species level. Columbia University Press, New York, 1997, S. 1–631 (S. 469–490)
- Leonard B. Radinsky: The Adaptive Radiation of the Phenacodontid Condylarths and the Origin of the Perissodactyla. Evolution 20 (3), 1966, S. 408–417
- Malcolm C. McKenna, Chow Minchen, Ting Suyin und Luo Zhexi: Radinskya yupingae, a Perissodactyl-like mammal from the Late Palaeocene of China. In: Donald R. Prothero und Robert M. Schoch (Hrsg.): The evolution of perissodactyls. New York und London 1989, S. 24–36
- K. Christopher Beard: East of Eden: Asia as an important center of taxonomic origination in mammalian evolution. Bulletin of the Carnegie Museum of Natural History 34, 1998, S. 5–39
- Kenneth D. Rose: The beginning of the age of mammals. Johns Hopkins University Press, Baltimore, 2006, S. 1–431 (S. 242–244 und S. 266–267)
- Sunil Bajpai, Vivesh Kapur, Debasis P. Das, B. N. Tiwari, N. Saravanan und Ritu Sharma: Early Eocene land mammals from Vastan Lignite Mine, District Surat (Gujarat), western India. Journal of the Palaeontological Society of India 50 (1), 2005, S. 101–113
- Sunil Bajpai, Vivesh Kapur, J. G. M. Thewissen, Debasis P. Das und B. N. Tiwari: New Early Eocene cambaythere (Perissodactyla, Mammalia) from the Vastan Lignite Mine (Gujarat, India) and an evaluation of cambaythere relationships. Journal of the Palaeontological Society of India 51 (1), 2006, S. 101–110
- Kenneth D. Rose, Thierry Smith, Rajendra S. Rana, Ashok Sahni, H. Singh, Pieter Missiaen und A. Folie: Early Eocene (Ypresian) continental vertebrate assemblage from India, with description of a new anthracobunid (Mammalia, Tethytheria). Journal of Vertebrate Paleontology 26 (1), 2006, S. 219–225
- Kishor Kumar: Comments on ‘Early Eocene land mammals from Vastan Lignite Mine, District Surat (Gujarat), western India’ by Bajpai, S. et al. published in Journal of the Palaeontological Society of India 50, 1: 101-113, 2005. Palarch 1 (2), 2006, S. 7–13
- Kenneth D. Rose, Luke T. Holbrook, Rajendra S. Rana, Kishor Kumar, Katrina E. Jones, Heather E. Ahrens, Pieter Missiaen, Ashok Sahni und Thierry Smith: Early Eocene fossils suggest that the mammalian order Perissodactyla originated in India. Nature Communications 5 (5570), 2014 doi:10.1038/ncomms6570
- Thierry Smith, Floréal Solé, Pieter Missiaen, Rajendra S. Rana, Kishor Kumar, Ashok Sahni und Kenneth D. Rose: First early Eocene tapiroid from India and its implication for the paleobiogeographic origin of perissodactyls. Palaeovertebrata 39 (2), 2015, S. e5, doi:10.18563/pv.39.2.e5
- Ross Secord, Jonathan I. Bloch, Stephen G. B. Chester, Doug M. Boyer, Aaron R. Wood, Scott L. Wing, Mary J. Kraus, Francesca A. McInerney und John Krigbaum: Evolution of the Earliest Horses Driven by Climate Change in the Paleocene-Eocene Thermal Maximum. Science 335, 2012, S. 959–962
- David J. Froehlich: Quo vadis eohippus? The systematics and taxonomy of the early Eocene equids (Perissodactyla). Zoological Journal of the Linnean Society, 134, 2002, S. 141–256
- Bin Bai, Yuan-Qing Wang und Jin Meng: The divergence and dispersal of early perissodactyls as evidenced by early Eocene equids from Asia. Communications Biology 1, 2018, S. 115, doi:10.1038/s42003-018-0116-5
- Pieter Missiaen and Philip D. Gingerich: New Early Eocene Tapiromorph Perissodactyls from the Ghazij Formation of Pakistan, with Implications for Mammalian Biochronology in Asia. Acta Palaeontologica Polonica 57 (1), 2012, S. 21–34
- Pieter Missiaen, Gregg F. Gunnell und Philip D. Gingerich: New Brontotheriidae (Mammalia, Perissodactyla) from the Early and Middle Eocene of Pakistan with Implications for Mammalian Paleobiogeography. Journal of Paleontology 85 (4), 2011, S. 665–677
- Bin Bai, Yuanqing Wang, Jin Meng, Qian Li und Xun Jin: New Early Eocene Basal tapiromorph from Southern China and Its Phylogenetic Implications. PlosONE 9 (10), 2014, S. e110806
- Kerstin Hlawatsch und Jörg Erfurt: Zahnmorphologie und stratigraphische Verbreitung von Hyrachyus minimus (Perissodactyla, Mammalia) in den eozänen Geiseltalschichten. In: Jörg Erfurt, Lutz Christian Maul (Hrsg.): 34. Tagung des Arbeitskreises für Wirbeltierpaläontologie der Paläontologischen Gesellschaft 16.3 bis 18.3.2007 in Freyburg/Unstrut. Hallesches Jahrbuch für Geowissenschaften BH 23, 2007, S. 161–173
- Christine Janis: An Evolutionary History of Browsing and Grazing Ungulates. In: I. J. Gordon und H. H. T. Prins (Hrsg.): The Ecology of Browsing and Grazing. Ecological Studies 195, 2008, S. 21–45
- Mikael Fortelius und John Kappelmann The largest land mammal ever imagined. Zoological Journal of the Linnean Society 107, 1993, S. 85–101
- Jérémy Tissier, Damien Becker, Vlad Codrea, Loïc Costeur, Cristina Fărcaş, Alexandru Solomon, Marton Venczel und Olivier Maridet: New data on Amynodontidae (Mammalia, Perissodactyla) from Eastern Europe: Phylogenetic and palaeobiogeographic implications around the Eocene-Oligocene transition. PLoS ONE 13 (4), 2018, S. e0193774, doi:10.1371/journal.pone.0193774
- Alexander Averianov, Igor Danilov, Jianhua Jin und Yingyong Wang: A new amynodontid from the Eocene of South China and phylogeny of Amynodontidae (Perissodactyla: Rhinocerotoidea). Journal of Systematic Palaeontology 15 (11), 2017, S. 927–945, doi:10.1080/14772019.2016.1256914
- Matthew Colbert: New Fossil Discoveries and the History of Tapirus. Tapir Conservation 16 (2), 2007, S. 12–14
- Ludovic Orlando, Jessica L. Metcalf, Maria T. Alberdi, Miguel Telles-Antunes, Dominique Bonjean, Marcel Otte, Fabiana Martin, Véra Eisenmann, Marjan Mashkour, Flavia Morello, Jose L. Prado, Rodolfo Salas-Gismondi, Bruce J. Shockey, Patrick J. Wrinn, Sergei K. Vasil’ev, Nikolai D. Ovodov, Michael I. Cherry Blair Hopwood, Dean Male, Jeremy J. Austin, Catherine Hänni und Alan Cooper: Revising the recent evolutionary history of equids using ancient DNA. PNAS 106, 2009, S. 21754–21759
- Étienne Geoffroy Saint-Hilaire und Georges Cuvier: Memoire sur une nouvelle division des Mammiferes, et sur les principes qui doivent servir de base dans cette sorte de travail. Magasin Encyclopedique 2, 1795, S. 164–190 (S. 178, 189) ()
- Georges Cuvier: Le regne animal distribue d'apres son organisation pour servir de base a l'histoire naturelle des animaux et d'introduction a l'anatomie comparee. (Band 1) Paris, 1817, S. 1–540 (S. 227–242)
- Johann Friedrich Blumenbach: Handbuch der Naturgeschichte. Göttingen, 1779, S. 168–448 (S. 109–112)
- Georges Cuvier: Tableau elementaire de l'histoire naturelle des animaux. Paris, Baudouin, 1798, S. 1–710 (S. 168–170) ()
- Henri Marie Ducrotay de Blainville: Prodrome d’une nouvelle distribution systèmatique du règne animal. Bulletin des Sciences 3 (3), 1816, S. 105–124 ()
- Richard Owen: Description of teeth and portions of jaws of two extinct anthrocotherioid quadrupeds (Hyopotamus vectianus and Hyop. bovinus) discovered by the Marchioness of Hastings in the Eocene deposits on the N.W. coast of the Isle of Wight: with an attempt to develope Cuvier’s idea of the classification of pachyderms by the number of their toes. The Quarterly journal of the Geological Society of London 4, 1848, S. 103–141 ()
- Othniel Charles Marsh: Dinocerata: a monograph of an extinct order of gigantic mammals. Washington, 1884/1886, S. 1–243 (S. 9, 177 und 186) ()
- Donald R. Prothero und Robert M. Schoch: Classification of the Perissodactyla. In: Donald R. Prothero und Robert M. Schoch (Hrsg.): The evolution of perissodactyls. New York und London 1989, S. 530–537
- Rodolphe Tabuce, Laurent Marivaux, Mohammed Adaci, Mustapha Bensalah, Jean-Louis Hartenberger, Mohammed Mahboubi, Fateh Mebrouk, Paul Tafforeau und Jean-Jacques Jaeger: Early Tertiary mammals from North Africa reinforce the molecular Afrotheria clade. Proceedings of the Royal Society B 274, 2007. S. 1159–1166
- Daten nach IUCN Red List of Threatened Species, Version 2012.2, abgerufen am 23. Februar 2013
Weblinks
- Detailliertes Kladogramm der Unpaarhufer
- Informationen, Abbildungen und Verbreitungskarten auf The Ultimate Ungulate Page (englisch)
- Informationen auf Animal Diversity Web (englisch)