Brontotheriidae
Die Brontotheriidae (Synonym Titanotheriidae), im Deutschen teilweise auch Donnerhuftiere genannt (nach der wissenschaftlichen Bezeichnung hergeleitet aus den griechischen Wörtern βροντή (brontḗ) für „Donner“ und θηρίον (theríon) für „Tier“), sind eine ausgestorbene Familie nashornähnlicher Unpaarhufer aus der chronostratigraphischen Serie des Eozän. Der überwiegende Teil der Fossilien wurde in Nordamerika, Asien gefunden, einige wenige auch in Osteuropa. Charakteristisches Kennzeichen war ein massiger Körperbau. Bei größeren und häufig auch bekannteren Gattungen der Gruppe befand sich ein paariges oder rammbockartiges Horn auf der Schnauze oberhalb der Augenhöhle, welches im Gegensatz zu jenem der Nashörner aus Knochensubstanz bestand; das Merkmal kommt jedoch nicht bei allen Mitgliedern der Familie vor. Brontotherien lebten in dichten Wäldern und waren allesamt Pflanzenfresser mit weitgehender Spezialisierung auf Blattnahrung. Über ihr Sozialverhalten ist nur wenig bekannt. Ihre Stammesgeschichte währte fast 20 Millionen Jahre und begann höchstwahrscheinlich in Nordamerika vor rund 53 Millionen Jahren mit noch recht kleinen, tapirgroßen Vertretern. Zum Ende des Eozäns kam es zu klimatischen Veränderungen mit im Vergleich kühleren Bedingungen und damit verbunden zu offeneren Landschaften, was möglicherweise der Grund ihres Aussterbens war. In der biologischen Systematik werden die Brontotherien häufig aufgrund des Zahnbaus in die Nähe der Pferde gestellt, generell sind die Verwandtschaftsverhältnisse der großen ausgestorbenen Unpaarhufergruppen zueinander jedoch noch nicht vollständig geklärt.
| Brontotheriidae | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelettrekonstruktion von Megacerops | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Eozän | ||||||||||||
| 53,4 bis 33,9 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Brontotheriidae | ||||||||||||
| Marsh, 1873 | ||||||||||||
Merkmale
Habitus

Brontotherien waren meist große bis sehr große Tiere, die größten Vertreter wie Megacerops konnten eine Schulterhöhe von 2,0 bis 2,5 m erreichen und wogen möglicherweise 2 bis 3 t.[1] Damit sind sie etwa vergleichbar mit dem heutigen Breitmaulnashorn.[2] Einige der frühen Formen, etwa Eotitanops und Palaeosyops, blieben jedoch deutlich kleiner und wurden mit Höhen von 1 m oder weniger nur etwa tapirgroß. Der Schädel war meist robust und im Vergleich zum Körper recht groß, bei späteren Formen trug er charakteristische knöcherne Hornbildungen. Der übrige Körperbau war allgemein nashornähnlich mit einem kräftigen Rumpf und einer Wirbelsäule mit lang auslaufenden Dornfortsätzen an den vorderen Brustwirbeln. Diese dienten als Ansatzstellen für eine massige Nackenmuskulatur, welche kräftig genug war, um den meist tief hängenden Kopf zu halten. Die kräftigen und kurzen Gliedmaßen glichen denen der heutigen Nashörner, die unteren Extremitäten waren allerdings im Durchschnitt kürzer. Die Vorderfüße wiesen vier und die Hinterfüße drei Zehen auf; ein markantes Merkmal für frühe Unpaarhufer, das heute einzig noch von den Tapiren bekannt ist. Insgesamt waren die Brontotherien nicht ganz so massig gebaut wie die heutigen Nashörner.[3][4][5][6]
Schädel- und Gebissmerkmale
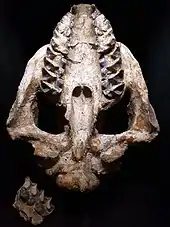
-PaleozoologicalMuseumOfChina-May23-08.jpg.webp)
Der Schädel war bei allen Brontotherien vergleichsweise groß und wurde bei den großen Vertretern 60 cm bis teils über 80 cm lang. Vor allem die evolutiv fortgeschrittenen Formen wiesen zudem weit ausladende und teils deutlich bogenförmig gestaltete Jochbeinbögen auf. Bei vielen Brontotherien-Vertretern war der Hirnschädel recht breit, nur frühe Formen besaßen einen recht schmalen Schädel. Der Schädel wies seitlich an den Scheitelbeinen markante parasagittale Leisten als Muskelansatzstellen auf. Typisch war auch das bisweilen stark verlängerte Hinterhauptsbein, vor allem bei späteren, horntragenden Vertretern, das meist in Seitenansicht spitzwinklig verlief und eine häufig tiefe Kopfhaltung hervorrief. Die für die späten Brontotherien charakteristischen Hörner saßen auf dem Rostrum am Übergang vom Stirn- zum Nasenbein. Der vordere Teil des Nasenbeins stand frei über den Oberkiefer- und Mittelkieferknochen, ähnlich den Nashörnern und Pferden, aber abweichend von den Tapiren mit ihren weit nach hinten versetzten Nasenknochen.[7] Weitere Charakteristiken, die alle Brontotherien teilten, umfassten ein gekürztes Rostrum und ein markant hinter den Augen verlängerter hinterer Schädelbereich.[5]
Der Unterkiefer hatte in der Regel einen kräftigen Bau und wies eine eher kurze Symphyse auf. Der überwiegende Teil der Brontotherien zeigte ein vollständiges Gebiss mit drei Schneidezähnen, einem Eckzahn, vier Prämolaren und drei Molaren je Kieferast. Nur die evolutiv jüngsten Formen besaßen eine um einen Schneidezahn reduzierte vordere Bezahnung und manchmal einen fehlenden vorderen Prämolaren. Dabei hatten die Schneidezähne bei vielen Vertretern eine kleine, kugelige Form. Die generell niederkronigen (brachyodonten) Backenzähne waren mit mondsichelartigen (bunoselenodonten) Zahnschmelzhöckern auf den Kauflächen ausgestattet, die zusätzlich typisch W-förmige Kanten auf den oberen Zähnen aufwiesen und dazu geeignet waren, festeres Pflanzenmaterial zu zermahlen.[5]
Hörner

Im Unterschied zu den Hörnern der Nashörner bestanden die Hornbildungen der Brontotherien nicht aus Keratin, sondern waren Auswüchse von Schädelknochen und blieben deshalb auch fossil erhalten. Sie bildeten sich aus dem hinteren Teil des Nasenbeins und waren von den vorderen Bereichen des Stirnbeins überdeckt.[8] Der überwiegende Teil der horntragenden Brontotherien besaß in der Regel zwei paarige Hörner, die sich häufig oberhalb der Orbita befanden, wobei in diesem Bereich die Breite des Nasenbeins deutlich größer wurde. Ebenfalls abweichend von den Nashörnern endeten die Hörner nicht spitz, sondern waren abgerundet. An der Basis wiesen sie einen runden bis ovalen Querschnitt auf und ragten teilweise schräg nach außen, an den Enden konnten die Hörner wie bei Megacerops auch leicht gegabelt sein.[9] In vielen Fällen bestanden die Hornbildungen aber auch nur aus größeren oder kleineren Knochenschwellungen, was vor allem typisch für die stammesgeschichtlich älteren Formen war.[10] Bei einigen asiatischen Brontotherien standen die Hörner häufig sehr dicht beieinander oder waren miteinander verwachsen und bildeten so eine knöcherne Ramme, die sich am deutlichsten bei Embolotherium zeigte. Es wird meist angenommen, dass die Hörner, ähnlich wie jene der Giraffen, mit Haut bedeckt waren, allerdings weisen sie an der Oberfläche keine Kanälchen für Blutgefäße auf. Jedoch ist bei Embolotherium mittlerweile nachgewiesen, dass die gesamte Ramme in die Weichteilbedeckung des Vorderkopfes mit einbezogen war und dieses so ein markant ausgebildetes Gesicht besaß.[5]
Fossilfunde

Funde von Brontotherien sind in großer Zahl bekannt und stammen aus Nordamerika und Eurasien. Dabei schwankt aufgrund der Beschaffenheit von Fossillagerstätten und unterschiedlicher Bergungs- und Präparationsmethoden die Qualität der Funde sowohl räumlich als auch zeitlich, wodurch einige Vertreter besser bekannt sind als andere. Hervorzuheben sind einige Fundgebiete, in denen Fossilreste der Brontotherien besonders zahlreich beziehungsweise qualitativ hervorragend überliefert wurden. Als wahre „Schatzkammer“ erwies sich der zentral-westliche Bereich Nordamerikas, hauptsächlich der nordwestliche Teil der USA mit den High Plains und den Rocky Mountains von South Dakota, Nebraska und Wyoming. Nach Norden reicht das Fundgebiet bis nach Saskatchewan und British Columbia im Südwesten Kanadas.[11] In diesem fossilreichen Gebiet wurden auch die ersten Funde entdeckt. In diesem Zusammenhang müssen vor allem die Badlands erwähnt werden, wo unzähliges Material aus der Chadron-Formation im Einzugsgebiet des White Rivers stammt. Diese Gesteinsbildung ist dem Oberen Eozän zuzuweisen und enthielt hauptsächlich Reste von Megacerops. Nach dem alten, nicht mehr gebräuchlichen Synonymnamen Titanotherium für diese Gattung, nennt man die fundführenden Schichten bis heute Titanotherium beds.[12] Ebenfalls hervorzuheben ist die als etwas älter einzustufende Duchesne-River-Formation in Utah mit qualitativ hochwertigen Resten von Duchesneodus, einem nahen Verwandten von Megacerops. Weiterhin herausragende Bedeutung hat die Clarno-Formation in Oregon mit gut erhaltenen Fossilfunden von Eubrontotherium.[5] Etwas weiter entfernt gelegen ist zudem zahlreiches Fundmaterial aus Kalifornien bekannt, darunter unter anderem nahezu frisch geborene Jungtiere von Parvicornus.[13] Die bisher nördlichsten Funde in Nordamerika kamen in der Margaret-Formation auf Ellesmere-Insel im Kanadisch-Arktischen Archipel nördlich des Polarkreises zu Tage. Sie datieren in den Übergang vom Unteren zum Mittleren Eozän und werden weitgehend zu Eotitanops und Palaeosyops gestellt.[14][15][16]
Außerhalb Nordamerikas sind sehr gute Fundstellen in Ostasien zu finden, wobei hier die Wüste Gobi herausragend ist. Aus den verschiedenen dort aufgeschlossenen Gesteinsformationen, etwa der Irdin-Manhan-Formation, der Shara-Murun-Formation oder der Ulan-Gochu-Formation des Mittleren und Oberen Eozäns stammen zahlreiche Fossilreste von Gnathotitan und Rhinotitan, aber auch von dem massiven Embolotherium. Die Fundgeschichte begann hier weitgehend in den 1920er Jahren mit Expeditionen des American Museum of Natural History[17], aber auch in der Zeit danach wurden noch zahlreiche Funde entdeckt.[18] Weiterhin wichtig sind die Aufschlüsse von Belutschistan in Südasien, aus denen einige der ältesten Brontotherien-Reste Asiens stammen.[19] Vor allem seit den 1990er Jahren nimmt die Bedeutung Zentralasiens als wichtige Region für die Forschung über diese Unpaarhufergruppe immer weiter zu, zu nennen sind etwa die überaus gut erhaltenen Fossilreste von Aktautitan aus Kasachstan.[4]
Paläobiologie
Ernährungsweise

Das charakteristische bunoselenodonte Zahnschmelzmuster auf den Kauflächen der Backenzähne und die niedrige Kronenhöhe dieser sind üblicherweise Anzeichen dafür, dass sich die Tiere von weicher Pflanzenkost ernährten (browsing).[20] Bestätigt wird dies durch mikroskopische Analysen der Zahnkauoberflächen verschiedener Brontotherien-Gattungen wie Eotitanops, Telmatherium, Metarhinus, Duchesneodus oder Megacerops. Die zahlreichen aufgefundenen engen Kratzer und kleinen Aushöhlungen lassen auf eine stärker auf Blattnahrung spezialisierte Ernährungsweise schließen, bei der nur gelegentlich kleinkörnige Bodensedimente mit aufgenommen wurden. Nur bei den frühesten Formen könnte ein höherer Anteil an Früchten eine Rolle gespielt haben, ähnlich wie es bei den frühesten Pferden der Fall war. Gemischte Pflanzenkost aus Rinde und Ästen oder Samen kann weitgehend ausgeschlossen werden. Da heutige Pflanzenfresser überwiegend ein anderes Kaumuster aufweisen, gehen die Forscher davon aus, dass Brontotherien höchst wählerisch bei der Nahrungssuche vorgingen. Weiterhin konnte ermittelt werden, dass vor allem spätere Vertreter der Brontotherien aus dem Oberen Eozän deutlich mehr Spuren von kleinen Kratzern auf den Zahnkauoberflächen aufwiesen, was entweder mit einer Umstellung der bevorzugten Nahrung oder mit generellen Landschaftsveränderungen zusammenhängen könnte.[21][22] Darüber hinaus ergaben Isotopenanalysen am Zahnschmelz der Backenzähne, dass die spätesten Brontotherien wie Megacerops ihre Nahrung zu nahezu 100 % aus Blättern bezogen und zudem eine recht hohe Wasserabhängigkeit bestand. Vor allem die Abhängigkeit von Flüssigkeit indiziert wiederum ein Verdauungssystem ähnlich zu den heutigen Unpaarhufern, bei denen ein Großteil der Nahrung im Enddarm zersetzt wird (Enddarmfermentierer).[2]
Geschlechtsunterschiede
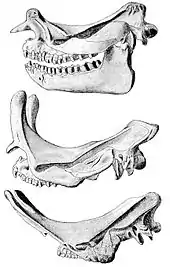
Innerhalb der einzelnen Brontotherien-Gattungen lassen sich Unterschiede im Körperbau feststellen, die häufig als Geschlechtsdimorphismus betrachtet werden. Bei vielen, vor allem hornlosen, Vertretern sind Abweichungen im Zahnbau nachweisbar, der bei einigen Individuen meist einen größeren, die benachbarten Zähne weit überragenden Eckzahn umfasst und der bei anderen Individuen weniger deutlich ausgebildet ist. In Verbindung mit großen Eckzähnen treten häufig auch robustere Schädel auf, die vor allem an den Jochbögen massivere Muskelansatzstellen für die Kaumuskulatur aufweisen, was sich dann in großen Knochenschwellungen ausprägt. Die Kombination von robusteren Schädeln und großen Eckzähnen wird meist mit männlichen Tieren in Verbindung gebracht, wo ähnliche Merkmale zum Beispiel bei den heutigen Pferden ebenfalls zu beobachten sind. Ein weiterer wahrscheinlicher Geschlechtsunterschied ist in der Horngröße bei einigen horntragenden Brontotherien zu bemerken, wobei wiederum Tiere mit größeren Hörnern und teilweise auch damit verbunden robusteren Nasenbeinen als männlich angesehen werden, während weibliche Tiere dagegen grazilere Bildungen aufweisen. Bekannt ist dies von Formen wie Megacerops, Duchesneodus und Embolotherium.[5][23][24] Bemerkenswert in diesem Zusammenhang ist, dass einige der spätesten Brontotherien keinen Unterschied mehr zwischen den Geschlechtern im Zahnaufbau besitzen, so dass die Eckzähne bei allen aufgefundenen Fossilien ähnlich groß sind. Vor allem kann dies bei Embolotherium nachgewiesen werden, das eine markante knöcherne Ramme auf dem Schädel hatte, aber auch anhand der Funde von Duchesneodus, das aber zwei Einzelhörner besaß.[24] Eventuell kam es hier zu einer sekundären Reduktion des Zahndimorphismus im Zusammenhang mit der Entwicklung der Hornbildungen. Ähnliches ist bei heutigen Horn- und Geweihträgern der Paarhufer zu finden: Vertreter mit Kopfwaffen haben in der Regel keine vergrößerten Eckzähne, bei solchen wie den Moschustieren oder den Wasserböcken, die kein Geweih ausbilden, sind diese aber zumindest im Oberkiefer stark verlängert. Vergleichbare Entwicklungen sind auch bei den Nashörnern bekannt.[10]
Sozialverhalten

Über das Sozialverhalten der Brontotherien ist wenig bekannt. Ein deutlicher Geschlechtsunterschied ist in der Regel typisch für die Paarung eines männlichen mit mehreren weiblichen Tieren in einer engen Sozialgruppe oder unter territorialen Tieren. Die Eckzähne und Hörner dienten also möglicherweise dem Imponierverhalten während der Brunft. Auf Rivalenkämpfe könnten auch vereinzelt aufgefundene Rippen mit verheilten Brüchen hinweisen. Einzelne Hörner besitzen auch Hinweise auf ein sekundäres Wachstum, möglicherweise ausgelöst durch Zusammenprall bei Kämpfen. Da aber knöcherne Hörner weniger zäh sind als solche aus Keratin, ist es wahrscheinlicher, dass eventuelle Kämpfe überwiegend eher in Schiebe- oder Druckwettstreiten ausgeführt wurden, als mittels Stoßen und Rammen.[3][10]
Fortbewegung
Der prinzipiell nashornartige Aufbau des Körperskeletts verweist aufgrund der Proportionen der Gliedmaßen auf einen eher schwerfälligen Gang.[6] Es zeigen sich aber einzelne Abweichungen, die weitere Aussagen zur Fortbewegung der Brontotherien zulassen. Vor allem die Hinterbeine besaßen ein annähernd symmetrisch aufgebautes Kniegelenk, so dass beide Gelenkrollen des Oberschenkelknochens ähnlich groß waren, vergleichbar zu denen heutiger Elefanten. Nashörner weisen dagegen stark asymmetrische Kniegelenke auf, die sich aufgrund ihrer extremen Körpermassenzunahme im Verlauf ihrer Stammesgeschichte bildeten. Sie ermöglichen einen sehr schnellen Lauf bis hin zum Galopp – eine Bewegungsart, bei der in einer Phase alle vier Füße gleichzeitig in der Luft sind – im offenen Gelände, wozu Elefanten nicht befähigt sind. Für die sehr großen Brontotherien wie Megacerops wird aufgrund des ähnlichen Aufbaus des Knies und des großen Gewichtes angenommen, dass auch hier ein schneller Lauf die maximale Geschwindigkeit bei der Fortbewegung gewesen sein muss.[25] Ähnliches lässt sich auch für Embolotherium annehmen, das sehr massive Langknochen besaß, welche auf einen eher schwerfälligen Gang schließen lassen. Andererseits hatte zum Beispiel das stammesgeschichtlich ältere Rhinotitan wesentlich schlankere Gliedmaßen, die möglicherweise auch eine flinkere Fortbewegung zuließen.[26]
Biologische Funktionen
Bei einigen Formen wie Metarhinus, Sphenocoelus und Telmatherium sind an der Schädelbasis die eigentlichen internen Nasengänge geschlossen und durch weiter nach hinten Richtung Pflugscharbein verschobene Öffnungen ersetzt. Dadurch wird die Atemluft durch die Nase kommend direkt zur Riechschleimhaut geleitet. Im Bereich des Siebbeins ist eine Art Nasenscheidewand ausgebildet, die die internen Atemwege in zwei separate Röhren unterteilt. Es erscheint dadurch möglich, dass Brontotherien obligat durch die Nase atmeten und nicht durch das Maul. Ähnliches ist auch von den heutigen Pferden bekannt, die als nächste rezente Verwandte der Brontotherien gelten, ebenso in einzelnen Einschränkungen auch von den Nashörnern. Mit weniger Wahrscheinlichkeit könnte auch eine Funktion bei einer möglichen aquatischen Lebensweise in Betracht kommen. Dabei würde theoretisch die Versetzung der internen Atemwege nach hinten die Luftzufuhr bei mit Wasser gefülltem Rachenraum unterstützen, was aber anatomisch bisher nicht eindeutig ist.[27][28]
Paläoumwelt
Aufgrund der Ernährungsgewohnheiten der Brontotherien mit ihrer bevorzugten Nahrung und des benötigten Wassers dürften die Tiere weitgehend in dichten, geschlossenen und von Flüssen sowie Sümpfen durchsetzten Wäldern mit feuchten Klimabedingungen gelebt haben. Darauf lassen unter anderem die Isotopenuntersuchungen an Zähnen aus der Chadron-Formation im Einzugsgebiet des White Rivers, die dem Oberen Eozän angehört, schließen.[2] Unterstützung finden diese durch einen Großteil der geologischen und paläontologischen Fundumstände. Analysen zur Paläoflora der gleichfalls obereozänen Australian-Creek-Formation in British Columbia, die Zahnreste von Brontotherien barg, erlauben eine Rekonstruktion von Nadel-Laub-Mischwäldern unter gemäßigten Klimabedingungen. Die Jahresdurchschnittstemperatur wird auf etwa 13 °C veranschlagt mit einem Durchschnitt für den kältesten Monat von −4 °C. Der Jahresdurchschnittsniederschlag lag bei 115 cm.[11] Dem gegenüber zeigten die Brontotherien aber auch eine gewisse Anpassungsfähigkeit an extreme Umweltbedingungen, wie dies die Hinterlassenschaften der unter-mitteleozänen Margaret-Formation auf Ellesmere-Insel im hohen Norden Kanadas annehmen lassen. Die ehemalige Umwelt kann als vergleichbar zu der der Australian-Creek- und Chadron-Formation angenommen werden, doch unterlag sie den Auswirkungen des Polartags und der Polarnacht mit sich abwechselnder mehrmonatiger Helligkeit und Dunkelheit. Dadurch kam es wahrscheinlich im Jahreszyklus zu regelmäßigen Engpässen bei den Nahrungspflanzen. Inwiefern die Tiere hier das gesamte Jahr im hohen Norden verbrachten, ist bisher ungeklärt, für einige Formen sind aber Jungtiere belegt, die eine permanente Anwesenheit wahrscheinlich machen.[14][15][16]
Systematik
Äußere Systematik
Innere Systematik der Perissodactyla nach Holbrook und Lapergola 2011[29]
|
Innere Systematik der Perissodactyla nach Hooker und Dashzeveg 2004[30]
|
Die Brontotheriidae sind eine ausgestorbene Familie aus der Ordnung der Unpaarhufer (Perissodactyla). Die Unpaarhufer werden traditionell in zwei große Linien unterteilt, die Hippomorpha (die Pferde) und die Ceratomorpha (der Verwandtschaftskomplex aus Nashörnern und Tapiren), wobei mit den Ancylopoda (die ausgestorbenen Chalicotherien) später eine dritte eingeführt wurde. Diese drei Gruppen werden überwiegend als Unterordnungen angesehen. In dieser häufig vertretenen Ansicht unterstreicht das übergeordnete Taxon der Tapiromorpha wiederum die nähere Stellung der Ceratomorpha und Ancylopoda zueinander. Dabei werden die Brontotherien meist zu den Hippomorpha gestellt und sind dadurch trotz ihres abweichenden Äußeren bei den entwickelten Formen näher mit den Pferden als mit den Nashörnern verwandt. Die Verwandtschaft mit den frühen Pferden zeigt sich an zahlreichen Schädelmerkmalen, etwa dem Bau des breiten Nasenbeins oder der kurzen Symphyse des Unterkiefers und weiterhin anhand der Zähne. Die nächstverwandte Gruppe sind aber die ebenfalls ausgestorbenen Lambdotheriidae, mit denen die Brontotherien zusammen die Überfamilie der Brontotherioidea bilden.[7][29] Es gibt aber auch Ansichten, dass die Brontotherien zusammen mit den Lambdotherien eine sehr basale Stellung innerhalb der Unpaarhufer einnehmen, da sie zahlreiche sehr ursprüngliche Merkmale dieser Säugetierordnung vereinen. In diesem Fall werden die Brontotherien und ihre Verwandten der eigenen Unterordnung der Titanotheriomorpha zugewiesen. Allerdings sind die genauen Verwandtschaftsverhältnisse zwischen den Pferden und den Brontotherien und wiederum zu den anderen Unpaarhufern noch nicht genau erforscht.[30]
Innere Systematik
Ursprünglich wurden die einzelnen Brontotherien-Gattungen verschiedenen Unterfamilien wie den Brontotheriinae, Telmatheriinae oder Embolotheriinae zugewiesen. Die innere Systematik der Familie war aber weitgehend nur für die nordamerikanischen Vertreter gut untersucht,[23] während die der eurasischen Formen nur selten berücksichtigt wurde.[31][32] Zwar waren diese auch in einzelnen Unterfamilien organisiert[6][17], doch bestanden Probleme in der Verbindung mit den amerikanischen Vertretern. Dieser Umstand wurde erst während einer großen Revision der Brontotherien durch Matthew C. Mihlbachler im Jahr 2008 behoben[5]. Ihr folgte im Jahr 2010 durch Bryn J. Mader eine Teilrevision der frühesten Vertreter, die Mihlbachler nur wenig berücksichtigt hatte.[33] Nach diesen beiden Überarbeitungen verblieben vorerst drei Unterfamilien innerhalb der Brontotherien, wobei jene der Brontotheriinae alle moderneren Formen beinhaltet. Dadurch wurden alle weiteren höheren Taxa auf die Ebene von Triben, Untertriben und Zwischentriben verschoben. Hier wiederum repräsentieren die Brontotheriina alle Brontotherien mit Hornbildungen, während die Telmatheriina nur Knochenschwellungen und die Rhadinorhinina keine solchen Merkmale aufweisen. Die Zwischentriben der Brontotheriita und Embolotheriita unterscheiden sich dabei im Bau der Hörner. Erstere weisen meist zwei separate Hörner auf, letztere eng stehende oder verwachsene und teils rammbockartig ausgebildete. Die einzelnen Gruppen und Linien der Brontotherien sind nicht geographisch gebunden, sondern zeigen vielfältige Beziehungsgeflechte über die verschiedenen Kontinente ihrer damaligen Verbreitung hinweg. Dies lässt einen mehrfachen und in unterschiedliche Richtungen verlaufenden Faunenaustausch zwischen den einzelnen Landmassen während der Stammesgeschichte der Brontotherien annehmen.[5][34]
Überblick über die Gattungen
|
Insgesamt sind heute über 40 Gattungen aus der Familie der Brontotheriidae bekannt, die letzte umfangreiche Revision der Familie erfolgte wie erwähnt im Jahr 2008. Folgende Gliederung wurde dabei vorgeschlagen, wobei Modifizierungen nach Mader aus dem Jahr 2010 und neuere Entdeckungen mit eingeflossen sind[5][19][33][34][35][36]:
- Familie: Brontotheriidae Marsh, 1873
- Balochititanops Missiaen, Gunnell & Gingerich, 2011
- Pakotitanops West, 1980
- Xylotitan Mihlbachler & Samuels, 2016
- Danjiangia Wang, 1995
- Unterfamilie: Eotitanopinae Osborn, 1914
- Eotitanops Osborn, 1907
- Unterfamilie: Palaeosyopinae Steinmann & Döderlein, 1890
- Palaeosyops (= Limnohyops, Limnohyus, Eometarhinus) Leidy, 1870
- Unterfamilie: Brontotheriinae Marsh, 1873
- Bunobrontops Holroyd & Ciochon, 2000
- Mesatirhinus Osborn, 1908
- Dolichorhinus Hatcher, 1895
- Sphenocoelus Osborn, 1895
- Desmatotitan Granger & Gregory, 1943
- Acrotitan Ye, 1983
- Tribus: Brontotheriini Marsh, 1873
- Microtitan Granger & Gregory, 1943
- Sthenodectes Gregory, 1912
- Epimanteoceras Granger & Gregory, 1943
- Nanotitanops Qi & Beard, 1998
- Untertribus: Rhadinorhinina Osborn, 1929
- Fossendorhinus Mihlbachler, 2008
- Metarhinus Osborn, 1908
- Untertribus: Telmatheriina Osborn, 1914
- Telmatherium (= Leurocephalus, Manteoceras) Marsh, 1872
- Metatelmatherium Granger & Gregory, 1938
- Wickia Mihlbachler, 2008
- Qufutitan Wang & Wang, 1997
- Untertribus: Brontotheriina Marsh, 1873
- Protitan Granger & Gregory, 1943
- Protitanotherium Hatcher, 1895
- Rhinotitan Granger & Gregory, 1943
- Zwischentribus: Embolotheriita Osborn, 1929
- Pollyosbornia Mihlbachler, 2008
- Gnathotitan Granger & Gregory, 1943
- Aktautitan Mihlbachler, Lucas, Emry & Bayshashov, 2004
- Pygmaetitan Miao, 1982
- Brachydiastematherium Böckh & Matyasovski, 1876
- Metatitan Granger & Gregory, 1943
- Maobrontops Averianov, Danilov, Chen & Jin, 2018
- Nasamplus Mihlbachler, 2008
- Protembolotherium Tang, You, Xu, Qiu & Hu, 1974
- Embolotherium (einschließlich Titanodectes) Osborn, 1929
- Zwischentribus: Brontotheriita Marsh, 1873
- Diplacodon (= Eotitanotherium) Marsh, 1875
- Pachytitan Granger & Gregory, 1943
- Parvicornus Mihlbachler & Deméré, 2009
- Parabrontops Granger & Gregory, 1943
- Eubrontotherium Mihlbachler, 2007
- Protitanops Stock, 1936
- Notiotitanops Gazin & Sullivan, 1942
- Dianotitan Chow & Hu, 1959
- Duchesneodus (= Teleodus) Lucas & Schoch, 1982
- Megacerops (= Allops, Ateleodon, Brontops, Brontotherium, Menodus, Menops, Oreinotherium, Titanops) Leidy, 1870
Einige Taxa werden als unsicher in ihrem Gattungsstatus angesehen, aber teilweise in der Literatur so geführt[5]:
- Arctotitan Wang, 1978
- Mulkrajanops Kumar & Sahni, 1985
- Sivatitanops Pilgrim, 1925
Stammesgeschichte
Adaptive Radiation
Generelle Evolutionstrends der Brontotherien sind in der Zunahme der Körpergröße und der Ausbildung knöcherner Hörner am Übergang vom Nasen- zum Stirnbein zu finden. Sehr deutliche Veränderungen erfuhr der Schädel, der bei den ursprünglichsten Vertretern noch sehr flach oder leicht gewölbt verlief. Die zunehmende Verlängerung des Hinterhauptsbeines mit der daraus resultierenden tiefen Kopflage verursachte zusätzlich eine teils deutlich gesattelte Stirnlinie. Weiterhin kam es zu Verkürzungen im vorderen Schnauzenbereich, die dazu führten, dass sich die Lage der Orbita in Bezug auf die Zahnstellung veränderte. So besaßen die frühen Brontotherien noch ein Augenfenster oberhalb oder leicht hinter dem dritten Molar, während spätere dieses oberhalb des ersten oder zweiten Molaren hatten. Dadurch ergab sich auch, dass die modernen Vertreter einen sehr ausgedehnten hinteren Schädel aufwiesen. Im Gebiss kam es zur Modifikation der Prämolaren, die den Molaren immer ähnlicher wurden, außerdem in einigen Linien zur Ausbildung eines Diastemas. Aufgrund der Nahrungsumstellung der frühen Vertreter von eher auf Früchten basierender zu blattreicher Ernährung entstanden im Laufe der Stammesgeschichte aus den bunodonten Molaren deutlich lophodonte (mit quergestellten Zahnschmelzleisten) bis hin zu selenodonten (mit mondsichelartigen Zahnschmelzleisten) gestaltete hintere Backenzähne. Die Zahnanzahl veränderte sich kaum, lediglich einige späte Brontotherien besaßen im vorderen Gebiss je einen Schneidezahn weniger. Der Faunenaustausch fand höchstwahrscheinlich über den hohen Norden statt.[5][37]
Ursprung und Entwicklung
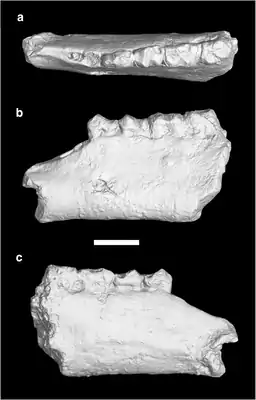
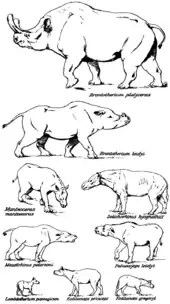
Die Evolution dieser Gruppe ist aufgrund der ausgezeichneten Fossilüberlieferung aus Nordamerika und Ost- sowie Zentralasien gut bekannt, ihre Herkunft ist aber noch nicht restlos geklärt. Einige Forscher nehmen einen Ursprung der Brontotherien in Nordamerika an. Dort traten die frühesten Formen im Unteren Eozän vor rund 53 Millionen Jahren auf (lokalstratigraphisch Wasatchium genannt). Als mögliche Ausgangsform gilt teilweise Lambdotherium, das ursprünglich noch als basalste Form der Brontotherien angesehen wurde, mittlerweile aber in einer eigenen Familie steht. Frühe Funde stammen mit zwei Teilskeletten aus dem Fossil Butte Member der Green-River-Formation im Green-River-Becken im Nordwesten der USA. Diese datieren in die lokalstratigraphische Faunenzone Lostcabinian und sind rund 52 Millionen Jahre alt.[33][37][38] Insgesamt handelte es sich bei den frühen nordamerikanischen Brontotherien um noch recht kleine Formen wie Eotitanops oder Palaeosyops, die eine Schulterhöhe von einem Meter und weniger aufwiesen und noch hornlos waren; für ersteres wird ein Gewicht von rund 140 kg errechnet.[34] Zwar waren sie zu jener Zeit ein relativ seltenes Faunenelement, doch sind Funde auch aus dem sehr hohen Norden Nordamerikas unter anderem aus der Margaret-Formation der heutigen Ellesmere-Insel[14][15][16] und der Buchanan-Lake-Formation der heutigen Axel-Heiberg-Insel[39] nachgewiesen, welche damals noch von Sumpfwäldern bewachsen waren.[15] Nach Ansicht einiger andere Forscher könnte der Ursprung der Gruppe auch in Asien liegen. Hier ist mit Danjiangia ein eventuell sehr früher Angehöriger aus der Lingcha-Formation in der chinesischen Provinz Henan belegt. Er zeigt einige Übereinstimmungen mit Lambdotherium, weist aber mit einer Datierung in den Paläozän-Eozän-Übergangsbereich vor rund 56 Millionen Jahren ein etwas höheres Alter auf (lokalstratigraphisch Bumbanium genannt).[36] Andere frühe Formen aus Asien wurden etwa mit Balochititanops aus der Ghazij-Formation in der Region Kingri in Belutschistan berichtet, sie traten etwa zeitgleich mit ihren nordamerikanischen Verwandten auf.[19]
Die Blütezeit der Brontotherien stellte aber das Mittlere Eozän vor etwa 50 bis 37 Millionen Jahren dar, aus jener Epoche sind etwa zwei Dutzend Gattungen bekannt.[37] Markant war vor allem die starke Körpergrößenzunahme. Für die frühmitteleozänen Funde des immer noch recht urtümlichen Bunobrontops aus der Pondaung-Formation von Myanmar wird ein Gewicht von 510 bis 990 kg veranschlagt, allerdings ist die Gattung weitgehend nur durch Zähne bekannt.[40] Dem gegenüber liegen die Gewichtsschätzungen für die etwa zeitgleich auftretenden, stammesgeschichtlich aber jüngeren Formen Wickia und Metatelmatherium bei etwa 1,6 bis 2,1 t.[34] Entgegen diesem Trend der Körpergrößenzunahme bildeten sich aber gelegentlich auch Zwergformen heraus. Der Prozess ist bisher noch nicht vollständig geklärt, Verzwergungen kamen aber im Verlauf der Stammesgeschichte der Brontotherien mehrfach vor und sind unter anderem mit Nanotitanops[41] und mit Xylotitan[34] belegt. Verbunden war der allgemeine Anstieg der Körpergröße auch mit einer Zunahme der Schädelgröße. Die Schädel entwickelten sich dabei wie bei Dolichorhinus zu sehr langgestreckten Formen, bei denen die Augen weit vorne lagen. Deutliche Hornbildungen sind erstmals im ausgehenden Mitteleozän zu beobachten, die Entwicklung dazu verlief aber in mehreren Stufen. So begann bei einigen Formen zuerst das Stirnbein mit kleinen, dreieckig geformten knöchernen Auswüchsen über das Nasenbein zu wachsen. Bekannt ist dies unter anderem von Telmatherium, das durch ungewöhnlich reiches Fossilmaterial vom Twin Buttes aus dem Bridger-Becken in Wyoming überliefert ist. Die Gattung wurde früher schon oft als das Schwestertaxon zu allen anderen horntragenden Brontotherien angenommen. Später bildeten sich dann kleine knöcherne Auswüchse, wie beim asiatischen Rhinotitan und beim nordamerikanischen Protitanotherium nachgewiesen. Erst dann formten sich die klassischen Hörner. Ebenfalls im ausgehenden Mitteleozän erreichen Brontotherien für kurze Zeit Europa, wo diese Unpaarhufergruppe aber nur selten nachgewiesen ist. In der Nähe von Cluj-Napoca in Rumänien wurde ein Unterkieferfragment von Brachydiastematherium gefunden.[5] Etwas jünger und schon aus dem Obereozän stammend sind die Zahnfunde und Unterkieferreste von Kameno und Tscherno More, beide Bulgarien, deren genaue systematische Stellung aber umstritten ist.[42][43] Insgesamt waren die Brontotherien im Mittleren Eozän die vielfältigste Gruppe an großen Säugetieren sowohl in Nordamerika als auch in Asien. Bemerkenswert ist dabei, dass es auf keinem der beiden Kontinente zu einer unabhängigen Radiation kam, vielmehr ist die phylogenetische Entwicklung stark miteinander verzahnt. Einige Forscher gehen von bis zu einem Dutzend unabhängiger interkontinentaler Ausbreitungswellen aus, die in beide Richtungen verliefen.[5][34][16]
Im Oberen Eozän vor 37 bis 34 Millionen Jahren ging die Anzahl der verschiedenen Brontotherien-Formen allmählich zurück. Etwa zehn Gattungen sind aus dieser Zeit bekannt, die meisten sind aber in die Frühphase der Epoche zu stellen. Alle nachgewiesenen Brontotherien waren horntragend und überwiegend sehr groß, wie das möglicherweise bis zu 3 t schwere Megacerops. Dabei bildeten sich vor allem zwei unterschiedliche Linien aus: Jene mit zwei weit auseinanderstehenden Hörnern mit einer nahen Verwandtschaft zu Megacerops und solche mit zwei eng stehenden oder verwachsenen Hörnern, die in einer näheren Beziehung zu Embolotherium stehen. Gegen Ende des Eozäns kam es zu Klimaveränderungen in Form eines Temperaturrückganges, was sich im Verlauf des Oligozäns noch verstärkte und in deren Zuge sich die ersten Gräser in Verbindung mit offenen Landschaften ausbreiteten. Möglicherweise vermochten die Brontotherien ihre hochspezialisierte Ernährungsweise diesen Landschaftsveränderungen in den mittleren und höheren Breiten, in denen sie hauptsächlich verbreitet waren, nicht anzupassen und wurden von konkurrenzfähigeren Pflanzenfressern verdrängt, etwa den aufsteigenden Gruppen der Nashörner und Chalicotherien.[20] Die meisten Brontotherien waren schon vor dem Ende des Eozän verschwunden, nur Megacerops hielt sich noch bis vor etwa 34 Millionen Jahren.[37]
Forschungsgeschichte

Die Erforschung der Brontotherien reicht bis in die 1840er Jahre zurück und begann in Nordamerika mit der ersten Entdeckung von Überresten dieser Unpaarhufergruppe. Der erste Fund, ein Unterkieferfragment mit den erhaltenen hinteren drei Backenzähnen, geht auf Hiram A. Prout zurück und stammt aus den Badlands des White River in South Dakota. Prout sprach in einer kurzen, 1846 erschienenen Publikation den Unterkieferfund als zu einem riesigen Palaeotherium, einem urtümlichen Pferdeverwandten, gehörig an, ein Jahr später beschrieb er den Unterkiefer genauer.[44][45] Weitere Funde aus der gleichen Region einschließlich des von Prout vorgestellten und abgebildeten Unterkieferrestes verwies Joseph Leidy (1823–1891), der als einer der Begründer der Paläontologie in Nordamerika gilt, später (im Jahr 1852) zu Titanotherium, wobei er den Namen eher am Rande erwähnte.[46] Jedoch hatte der französische Forscher Auguste Pomel (1821–1898) diesen Unterkieferfund bereits 1849 unter dem Namen Menodus beschrieben, wodurch dieser Name gegenüber Titanotherium vordatiert. Pomel benannte dabei nicht nur die Gattung, sondern fügte ihr im Unterschied zu Prout und später Leidy die Art Menodus giganteus bei. Somit handelt es sich um das forschungsgeschichtlich älteste Binomen, das für einen Brontotherien-Vertreter vergeben worden war. Zudem stellt der Unterkiefer und der mit ihm verbundene Artname die erste vergebene wissenschaftliche Bezeichnung für ein Fossil aus dem extrem fundreichen Gebiet der Badlands des White River dar.[12] Lange Zeit galt Prouts Fossilfund als verschollen, er sollte noch im Jahr der Veröffentlichung Pomels während eines großen Stadtbrandes in St. Louis verloren gegangen sein, doch tauchte er 1957 im National Museum of Natural History in Washington, D.C. wieder auf (heute als Exemplarnummer USNM 21820 geführt). Die Bezeichnung Menodus giganteus gilt aber heute als invalid, einerseits aufgrund der wechselvollen Geschichte des ursprünglichen Fundes und der zahlreich danach aufgestellten Gattungen und Arten, andererseits lässt sie sich keiner der heutigen gültigen Arten der Brontotherien (im engeren Sinne von Megacerops) zuordnen.[47] Schon in den 1850er Jahren fand der amerikanische Geologe Ferdinand Hayden (1829–1887) ein außerordentlich reichhaltiges Fossilmaterial in den Badlands, darunter auch zahlreiche Brontotherien, womit er einen wichtigen Beitrag in dieser sehr frühen Forschungsperiode leistete. In den 1870er bis 1890er Jahren stand die Brontotherien-Forschung im Schatten der Cope-Marsh-Fehde, auch „Bone wars“ genannt, die die beiden amerikanischen Paläontologen Edward Drinker Cope (1840–1897) und Othniel Charles Marsh (1831–1899) führten. Marsh war es dabei, der den Namen Brontotherium erstmals 1873 benutzte und anhand dreier Skelette aus dem Peabody Museum of Natural History der Yale University definierte.[48] Allerdings gilt Brontotherium nur als synonym zu Megacerops. Diese Bezeichnung stammt ursprünglich ebenfalls von Leidy, die er im Jahr 1870 veröffentlichte, und wurde von ihm anhand eines Schädels aus Colorado eingeführt, wobei er Ähnlichkeiten zu seiner vorher benannten Gattung Titanotherium erkannte.[49] Cope und Marsh sammelten in dieser Zeit Unmengen an fossilem Material in Nordamerika, vor allem in den beiden Dakota-Staaten und Nebraska, letzterer engagierte in den 1880er Jahren John Bell Hatcher, der ihm von 1886 bis 1888 allein 11.000 t an Fundmaterial zuschickte. Ebenso wurden zahlreiche Gattungen benannt, von denen aber einige – wie sich später herausstellte – identisch mit anderen waren.[3] Neben diesen herausragenden Funden waren die einzigen außeramerikanischen Fossilien, die im 19. Jahrhundert bekannt wurden, einige wenige Gebissreste aus Südosteuropa, über die in den 1870er und 1890er Jahren publiziert wurde.[5][43]
In der gleichen Publikation, in der Marsh die Gattung Brontotherium einführte, bezeichnete er dieses und Leidys Titanotherium als zur Familie der Brontotheriidae gehörig (allerdings als Brontotheridae geschrieben, die fehlerhafte Schreibweise wurde offiziell erst 1902 von Oliver Perry Hay korrigiert), dem heute gültigen Namen. Er erkannte auch die systematische Zugehörigkeit zu den Unpaarhufern,[48] allerdings verwies er nur die jüngeren Formen in diese Gruppe. Ältere Gattungen wie Diplacodon stellte er dagegen in die Familie der Limnohyidae.[50] Erst Cope vereinte beide Familien im Jahr 1879. Zum Ende des 19. Jahrhunderts waren aber mehrere Namen für die Familie der Brontotheriidae in Gebrauch, so unter anderem Menodontidae, welchen Cope 1881 aufstellte, oder Titanotheriidae vom britischen Wissenschaftler William Henry Flower aus dem Jahr 1876. Unabhängig von Flower und wohl in Unkenntnis seiner Schrift etablierte Henry Fairfield Osborn (1857–1935) im Jahr 1889 den Familienbegriff Titanotheriidae erneut und verwendete ihn zeit seines Lebens. Dieser Name ist heute zwar nur noch selten im Gebrauch, da er ein Synonym von Brontotheriidae darstellt, leitet sich jedoch von Leidys Titanotherium her.[5][23]
Zu Beginn des 20. Jahrhunderts und nach dem Tode Marshs war es Osborn, der die Forschungen maßgeblich vorantrieb. Osborn, am American Museum of Natural History tätig, bekam Zugang zu den Aufzeichnungen von Marsh und Cope und erarbeitete in der darauffolgenden Zeit eine Monographie über die Brontotherien (die er generell als Titanotherien bezeichnete). In den 1920er Jahren organisierte er Expeditionen nach Ostasien in die Wüste Gobi, die zwischen 1922 und 1930 stattfanden und als Central Asiatic Expeditions of the American Museum of Natural History bezeichnet werden. Bei diesen Expeditionen, meist geleitet von Roy Chapman Andrews und Walter W. Granger, wurden neben Dinosauriern und anderen fossilen Tieren auch unzählige Reste von Brontotherien mit bis dahin nicht bekannten Formen entdeckt, etwa Embolotherium oder Gnathotitan, womit die bis dahin nur vermutete Verbreitung dieser Unpaarhufergruppe bis nach Asien bewiesen war. Osborn arbeitete das Material in zahlreichen Publikationen nach und nach auf, im Jahr 1929 erschien seine mehr als 890 Seiten umfassende Schrift mit zusätzlich über 230 Tafeln unter dem Titel Titanotheres of ancient Wyoming, Dakota, and Nebraska über die nordamerikanischen Funde; diese enthielt aber noch einen mehr als 40 Seiten langen Anhang über die Funde aus Ostasien.[6] Nach Osborns Tod 1935 setzten Walter W. Granger und William King Gregory dessen Arbeiten fort.[3][5]
Literatur
Einzelnachweise
- Gregory S. Paul und Per Christiansen: Forelimb posture in neoceratopsian dinosaurs: implications for gait and locomotion. In: Paleobiology, 26 (3), 2000, S. 450–465, doi:10.1666/0094-8373(2000)026<0450:FPINDI>2.0.CO;2.
- Alessandro Zanazzi und Matthew J. Kohn: Ecology and physiology of White River mammals based on stable isotope ratios of teeth. In: Palaeogeography, Palaeoclimatology, Palaeoecology 257, 2008, S. 22–37, (PDF).
- Donald R. Prothero und Robert M. Schoch: Horns, tusks, and flippers. The evolution of hoofed mammals. Johns Hopkins University Press, Baltimore, 2003, ISBN 0-8018-7135-2, S. 229–239.
- Matthew C. Mihlbachler, Spencer G. Lucas, Robert J. Emry und Bolat Bayshashov: A New Brontothere (Brontotheriidae, Perissodactyla, Mammalia) from the Eocene of the Ily Basin of Kazakstan and a Phylogeny of Asian “Horned” Brontotheres. In: American Museum Novitates 3439, 2004, S. 1–43, doi:10.1206/0003-0082(2004)439<0001:ANBBPM>2.0.CO;2.
- Matthew C. Mihlbachler: Species taxonomy, phylogeny, and biogeography of the Brontotheriidae (Mammalia: Perissodactyla). In: Bulletin of the American Museum of Natural History 311, 2008, ISSN 0003-0090, S. 1–475.
- Henry Fairfield Osborn: The titanotheres of ancient Wyoming, Dakota, and Nebraska. In: United States Geological Survey Monographs 55, 1929, S. 1–894, (online).
- Luke T. Holbrook: Comparative osteology of early Tertiary tapiromorphs (Mammalia, Perissodactyla). In: Zoological Journal of the Linnean Society 132, 2001, S. 1–54, doi:10.1111/j.1096-3642.2001.tb02270.x.
- Matthew C. Mihlbachler und Nikos Solounias Niko: Anatomy and Evolution of the Bizarre “Battering Ram” of the Brontothere, Embolotherium (Mammalia, Perissodactyla). In: Journal of Morphology Special Issue: Seventh International Congress of Vertebrate Morphology, Boca Raton, Florida, 27 July to 1 August 2004. 260 (3), 2004, S. 274–342 (S. 313).
- Bryn J. Mader und John P. Alexander: Megacerops kuwagatarhinus n. sp., an unusual brontothere (Mammalia, Perissodactyla) with distally forked horns. In: Journal of Paleontology 69, 1995, S. 581–587, (online).
- Matthew C. Mihlbachler: A New Uintan Horned Brontothere from Wyoming and the Evolution of Canine Size and Sexual Dimorphism in the Brontotheriidae (Perissodactyla: Mammalia). In: Journal of Vertebrate Paleontology 31 (1), 2011, S. 202–214, doi:10.1080/02724634.2011.539653.
- Jaelyn J. Eberle und David R. Greenwood: An Eocene brontothere and tillodonts (Mammalia) from British Columbia, and their paleoenvironments. In: Canadian Journal of Earth Sciences 54, 2017, S. 981–992.
- Rachel C. Benton, Dennis O. Terry Jr., Emmett Evanoff und H. Gregory McDonald: The White River Badlands. Geology and Paleontology. Indiana University Press, 2016, S. 1–222 (S. 3 und 165–167).
- Matthew C. Mihlbachler und Thomas A. Deméré: A New Species of Brontotheriidae (Perissodactyla, Mammalia) from the Santiago Formation (Duchesnean, Middle Eocene) of Southern California. In: Proceedings of the San Diego Society of Natural History 41, 2009, S. 1–36.
- Jaelyn J. Eberle: Early Eocene Brontotheriidae (Perissodactyla) from the Eureka Sound Group, Ellesmere Island, Canadian High Arctic - Implications for Brontothere origins and high latitude dispersal. In: Journal of Vertebrate Paleontology, 26 (2), 2006, S. 381–386.
- Jaelyn J. Eberle und David R. Greenwood: Life at the top of the greenhouse Eocene world — A review of the Eocene flora and vertebrate fauna from Canada’s High Arctic. In: Geological Society of America Bulletin; January/February 124 (1/2), 2012, S. 3–23, (PDF).
- Jaelyn J. Eberle und David A. Eberth: Additions to the Eocene Perissodactyla of the Margaret Formation, Eureka Sound Group, Ellesmere Island, Arctic Canada. In: Canadian Journal of Earth Sciences 52, 2015, S. 123–133.
- Walter W. Granger und William K. Gregory: A revision of the Mongolian Titanotheres. In: Bulletin of the American Museum of Natural History 80, 1943, S. 349–389, (PDF).
- Wang Ban Yue: A Skull of Embolotherium (Perissodactyla, Mammalia) from Erden Obo, Nei Mongol, China. In: Vertebrata Palasiatica 38 (3), 2000, S. 237–240.
- Pieter Missiaen, Gregg F. Gunnell und Philip D. Gringerich: New Brontotheridae (Mammalia, Perissodactyla) from the Early and Middle Eocene of Pakistan with implications for Mammalian palaeobiogeography. In: Journal of Paleontology 85 (4), 2011, S. 665–677.
- Christine Janis: An Evolutionary History of Browsing and Grazing Ungulates. In: I. J. Gordon und H. H. T. Prins (Hrsg.): The Ecology of Browsing and Grazing., In: Ecological Studies 195, 2008, S. 21–45.
- Matthew C. Mihlbachler: Body size, dental microwear, and Brontotheres diets through the Eocene. In: Journal of Vertebrate Paleontology 22 (suppl.), 2002, S. 88A.
- Angana Homchaudhuri, Matthew C. Mihlbachler und Nikos Solounias: Dental microwear analysis of Eocene Brontotherioidea and implications for paleodietary interpretations of long extinct species. In: Journal of Vertebrate Paleontology 30 (suppl.), 2010, S. 107A.
- Bryn J. Mader: Brontotheriidae: A systematic revision and preliminary phylogeny of North American genera. In: Donald R. Prothero und Robert M. Schoch (Hrsg.): The evolution of perissodactyls. New York und London, 1989, S. 458–484.
- Spencer George Lucas und Robert M. Schoch: Taxonomy of Duchesneodus (Brontotheriidae) from the late Eocene of North America. In: Donald R. Prothero und Robert M. Schoch (Hrsg.): The evolution of Perissodactyls. New York und Oxford, 1989, S. 490–503.
- Christine M. Janis, Boris Shoshitaishvili, Robert Kambic and Borja Figueirido: On their knees: Distal Femur asymmetry in Ungulates and its relationship to body size and locomotion. In: Journal of Vertebrate Paleontology 32 (2), 2012, S. 433–445, doi:10.1080/02724634.2012.635737.
- Benjamin McLaughin, Matthew C. Mihlbachler und Mick Ellison: The postcranial skeleton of Embolotherium (Brontotheriidae) from the Middle and Late Eocne of Central Asia. In: Journal of Vertebrate Paleontology 30 (suppl.), 2010, S. 132A–133A.
- Bryn J. Mader: The cranial anatomy of Metarhinus (IMammalia, Perissodactyla, Brontotheriidae). In: Journal of Vertebrate Paleontology 29 (4), 2009, S. 1300–1305.
- Bryn J. Mader: The narial morphology of Metarhinus and Sphenocoelus (Mammalia, Perissodactyla, Brontotheriidae). In: Palaeontologia Electronica 22 (1), 2019, S. 8A (S. 1–15) ().
- Luke T. Holbrook und Joshua Lapergola: A new genus of Perissodactyl (Mammalia) from the Bridgerian of Wyoming, with comments on basal Perissodactyl phylogeny. In: Journal of Vertebrate Paleontology 31 (4), 2011, S. 895–901, doi:10.1080/02724634.2011.579669.
- J. J. Hooker und D. Dashzeveg: The origin of chalicotheres (Perissodactyla, Mammalia). In: Palaeontology 47 (6), 2004, S. 1363–1386, (PDF).
- Donald R. Prothero und Robert M. Schoch: Classification of the Perissodactyla. In: Donald R. Prothero und R. M. Schoch (Hrsg.): The evolution of the Perissodactyls. New York 1989, S. 530–537.
- Bryn J. Mader: Brontotheriidae. In: Christine M. Janus, Kathleen M. Scott und Louis L. Jacobs (Hrsg.): Evolution of Tertiary mammals from North America, Vol. 1. Cambridge 1998, S. 525–536.
- Bryn J. Mader: A species-level revision of the North American brontotheres Eotitanops and Palaeosyops (Mammalia, Perissodactyla). In: Zootaxa 2339, 2010, S. 1–43, (PDF).
- Matthew C. Mihlbachler und Joshua X. Samuels: A small-bodied species of Brontotheriidae from the middle Eocene Nut Beds of the Clarno Formation, John Day Basin, Oregon. In: Journal of Paleontology 90 (6), 2016, S. 1233–1244 doi:10.1017/jpa.2016.61.
- Alexander Averianov, Igor Danilov, Wen Chen und Jianhua Jin: A new brontothere from the Eocene of South China. In: Acta Palaeontologica Polonica 63 (1). 2018, S. 189–196 doi:10.4202/app.00431.2017.
- Bin Bai, Yuan-Qing Wang und Jin Meng: The divergence and dispersal of early perissodactyls as evidenced by early Eocene equids from Asia. In: Communications Biology 1, 2018, S. 115 doi:10.1038/s42003-018-0116-5.
- Donald R. Prothero: Evolutionary Transitions in the Fossil Record of Terrestrial Hoofed Mammals. In: Evo Edu Outreach 2, 2009, S. 289–302, doi:10.1007/s12052-009-0136-1.
- Lance Grande: The lost world of Fossil Lake. Snapshot from deep time. University of Chicago Press, Chicago und London, 2013, S. 1–425 (S. 270–273).
- Jaelyn J. Eberle and John E. Store: Northernmost Record of Brontotheres, Axel Heiberg Island, Canada: Implications for Age of the Buchanan Lake Formation and Brontothere Paleobiology. In: Journal of Paleontology 73 (5), 1999, S. 979–983.
- Takehisa Tsubamoto, Naoko Egi, Masanaru Takai, Chit Sein und Maung Maung: Middle Eocene ungulate mammals from Myanmar: A review with description of new specimens. In: Acta Palaeontologica Polonica 50 (1), 2005, S. 117–138, (PDF).
- Qi Tao und K. Christopher Beard: Nanotitan shanghuangensis, gen. et sp. nov.: the smallest known brontothere. In: Journal of Vertebrate Paleontology 16, 1996, S. 578–581.
- Ivan Nikolov und Kurt Heissig: Fossile Säugetiere aus dem Obereozän und Unteroligozän Bulgariens und ihre Bedeutung für die Paläogeographie. In: Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie 25, 1985, S. 61–70, (online).
- Franz Toula: Zwei neue Säugethierfundorte auf der Balkanhalbinsel. In: Akademie der Wissenschaften Wien Sitzungsberichte 101 (1), 1892, S. 608–615.
- Hiram A. Prout: Gigantic Palaeotherium. In: American Journal of Science and Arts 2, 1846, S. 288–289, (online).
- Hiram A. Prout: Description of a fossil maxillary bone of Palaeotherium, from near White River. In: American Journal of Science and Arts 3, 1847, S. 248–250, (online).
- Joseph Leidy: Description of the remains of extinct Mammalia and Chelonia, from Nebraska territory, collected during the Geological Survey under the direction of Dr. D. D. Owen. In: David Dale Owen (Hrsg.): Report of a geological survey of Wisconsin, Iowa, and Minnesota and incidentally of a portion of Nebraska Territory. Philadelphia: Lippincott, Grambo, 1852, S. 534–572 (S. 551–552), (online).
- Matthew C. Mihlbachler, Spencer G. Lucas und Robert J. Emry: The holotype specimen of Menodus giganteus, and the ‘‘insoluble’’ problem of Chadronian brontothere taxonomy. In: Spencer G. Lucas, K. E. Zeigler und P. E. Kondrashov (Hrsg.): Paleogene mammals. New Mexico Museum of Natural History and Science Bulletin 26, 2004, S. 129–135.
- Othniel Charles Marsh: Notice of New Tertiary Mammals. In: The American Journal of Science and Arts 3 (5), 1873, S. 485–488 (online).
- Joseph Leidy: (Description of a new genus and species, Megacerops coloradensis). In: Proceedings of the Academy of Natural Sciences of Philadelphia 22, 1870, S. 1–2, (online).
- Othniel Charles Marsh: Notice of New Tertiary mammals. IV. In: American Journal of Science and Arts 9, 1875, S. 239–250 (online).
Weblinks
- The Paleobiology Database Brontotheriidae