Brasilianische Wanderspinne
Die Brasilianische Wanderspinne (Phoneutria nigriventer), manchmal auch Brasilianische Kammspinne oder selten Giftige Bananenspinne genannt, ist eine Spinne aus der Familie der Kammspinnen (Ctenidae). Sie ist vorwiegend im Südosten Brasiliens, stellenweise im Norden Argentiniens, in Paraguay und mittlerweile ebenso in Uruguay verbreitet. Die Art zählt zu den weltweit größten Vertretern der Echten Webspinnen und ist, wie andere Arten der Bananenspinnen (Phoneutria), für ihre auch für den Menschen gefährlichen Giftbisse bekannt. Ihr Biss kann auch für einen gesunden erwachsenen Menschen tödlich verlaufen. In ihrem teilweise dicht besiedelten Verbreitungsgebiet tritt die Brasilianische Wanderspinne gehäuft in urbanen Räumen und dort auch vermehrt in Gebäuden auf. Aufgrund dessen treffen Menschen oft auf diese Spinne, und ihre hohe Aggressivität begünstigt Bissunfälle. Dennoch sind im Gift der Spinne einzelne Bestandteile enthalten, denen eventuell sowohl im pharmazeutischen als auch im landwirtschaftlichen Bereich Nutzen zugesprochen werden könnte.
| Brasilianische Wanderspinne | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Brasilianische Wanderspinne (Phoneutria nigriventer), Weibchen | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Phoneutria nigriventer | ||||||||||||
| (Keyserling, 1891) |
Die Brasilianische Wanderspinne wird auch zusammen mit anderen Spinnen der Familie schlicht als Wanderspinne oder ebenso wie andere Vertreter der Gattung einschließlich der Gattung selbst als Bananenspinne bezeichnet. Allerdings wird diese Bezeichnung auch für einzelne Arten der Gattung Cupiennius innerhalb der Familie der Fischerspinnen (Trechaleidae) verwendet. Grund dafür ist, dass in der Vergangenheit gelegentlich Individuen beider Gattungen in Bananenstauden in andere Kontinente der Welt unwissentlich mitexportiert wurden. Arten der Bananenspinnen einschließlich der Brasilianischen Wanderspinne können mit der für den Menschen harmloseren Gattung Cupiennius verwechselt werden. Ähnlichkeiten mit der Brasilianischen Wanderspinne bestehen überdies bei vielen anderen Kammspinnen oder weiteren Spinnen der Überfamilie der Lycosoidea.
Die Bezeichnung „Wanderspinne“ rührt von der nomadischen Lebensweise der nachtaktiven Art – Sie fertigt weder dauerhafte Unterschlüpfe noch Spinnennetze zum Fangzweck an, sondern erlegt wie alle Kammspinnen und überdies viele Vertreter der Überfamilie der Lycosoidea freilaufend als Lauerjäger beliebige Beutetiere. Am Tag zieht sich die Brasilianische Wanderspinne zumeist in geeignete und zeitweilig genutzte Versteckmöglichkeiten zurück, ehe sie nachts wieder zum Vorschein kommt. Die englischen und in dieser Sprache mehrdeutigen Trivialnamen Brazilian Wandering Spider und Banana spider sind von der Bedeutung her mit den deutschen Bezeichnungen „Brasilianische Wanderspinne“ und „Bananenspinne“ identisch. Auch hier dient die Bezeichnung Banana spider mitunter ebenso für einzelne Arten der Gattung Cupiennius und im englischen Sprachgebrauch zusätzlich für die ebenfalls harmlosere Warmhaus-Riesenkrabbenspinne (Heteropoda venatoria) aus der Familie der Riesenkrabbenspinnen (Sparassidae), die den Arten der beiden vorherigen Gattungen optisch hingegen nicht ähnelt. Wie diese und auch verschiedene Arten der Bananenspinnen sowie der Gattung Cupiennius wird die Brasilianische Wanderspinne gelegentlich als Heimtier im Bereich der Terraristik gehalten.
Merkmale
_por_Rodrigo_Tetsuo_Argenton_(4).jpg.webp)
Der Körperbau der Brasilianischen Wanderspinne entspricht dem anderer Bananenspinnen (Phoneutria), womit es sich auch bei ihr um eine große und robust gebaute Kammspinne handelt, die wie andere Arten der Familie entfernt an Wolfsspinnen (Lycosidae) erinnert.
Das Weibchen der Art erreicht eine Körperlänge von 25 bis 50 (durchschnittlich 35) Millimetern, während sich die Beinspannweite auf 150 Millimeter belaufen kann.[1] Andere Quellen sprechen von einer maximalen Beinspannweite von gut 180 Millimetern.[2][3] Bei den Maßen der Beinspannweite ist auch der dazwischenliegende Körper der Spinne zu berücksichtigen. Ein einzelnes Bein der Spinne erreicht nach bisherigen Kenntnissen eine Gesamtlänge von 45 etwa 60 Millimetern. Das Männchen ist insgesamt kleiner und weniger kräftig als das Weibchen gebaut, dafür ist es verglichen mit dem Weibchen langbeiniger.[1]
Die Körperlänge lässt die Brasilianische Wanderspinne zusammen mit ähnlich großen Arten der Gattung zu einer der weltweit größten Arten innerhalb der Unterordnung Echten Webspinnen zählen. Auch die Beinspannweite wird innerhalb dieser Unterordnung nur von einigen Arten der ebenfalls zu den Kammspinnen zählenden Gattung Ancylometes sowie einigen Riesenkrabbenspinnen (Sparassidae) übertroffen.[3]
Der gesamte Körper der Brasilianischen Wanderspinne ist dorsal hellbraun über braun bis grau gefärbt und überdies mit kurzen braunen bis grauen Haaren bedeckt. Die Art verfügt wie einige andere Arten der Gattung auch über sichtbare schwarze und gelbe oder weiße Streifen an der Unterseite der beiden vorderen Beinpaare, die bei der Drohgebärde sichtbar werden.[4]
Das dorsal hell- bis dunkelbraun gefärbte Opisthosoma (Hinterleib) wird durch zwei längslaufende Linien, die aus schwachen Pünktchen gebildet werden, ausgezeichnet. Am hinteren Rand dieser Linien sind bei einigen Individuen schwarze Punkte ausgebildet. Die Unterseite ist einheitlich und je nach Geschlecht von orange über braun bis schwarz gefärbt.[5] Auch hierbei handelt es sich um eine Warnfärbung, die bei der Drohgebärde zum Einsatz kommt.
_por_Rodrigo_Tetsuo_Argenton_(2).jpg.webp) Detailaufnahme eines Weibchens
Detailaufnahme eines Weibchens_por_Rodrigo_Tetsuo_Argenton_(1).jpg.webp) Dito, hier mit den gut sichtbaren Warnfarben.
Dito, hier mit den gut sichtbaren Warnfarben._029.jpg.webp) Rückansicht eines drohenden Weibchens
Rückansicht eines drohenden Weibchens
Sexualdimorphismus

Wie viele Spinnen weist auch die Brasilianische Wanderspinne einen ausgeprägten Sexualdimorphismus auf, der sich wie bei den weiteren Vertretern der Gattung zunächst in der Größe zeigt – das Männchen ist kleiner als das Weibchen.[6] Ferner ist die Ventralseite des Opisthosomas (Hinterleibs) beim Männchen heller gefärbt und erscheint zumeist in orangen bis braunen Farbtönen, während sie beim Weibchen dunkelbraun bis schwarz gefärbt ist.[5]
Genitalmorphologische Merkmale
Die Bulbi (männliche Geschlechtsorgane) der Brasilianischen Wanderspinne sind teilweise ellipsenförmig und die Emboli (Einfuhrorgane der Bulbi) sind gerade und schmal aufgebaut. Die Emboli verfügen über eine schmale Basis und auf prolateraler (frontal nach vorne gerichteter) Seite überdies über je einen schwach erhöhten Randkiel.[5]
Die flach aufgebaute Epigyne (weibliches Geschlechtsorgan) der Art weist zylindrisch verlaufende Seitenränder auf. Das Seitenfeld der Epigyne nahe der Kopulationsöffnung ist bei der Brasilianischen Wanderspinne anders als bei den anderen Vertretern der Gattung nicht gerillt und die Seitenapophyse (chitinisierter Fortsatz) verfügen hier über keine Sulci (Furchen).[5]
Ähnliche Arten

Die Brasilianische Wanderspinne ähnelt neben den Wolfsspinnen auch Vertretern weiterer Spinnenfamilien innerhalb der Überfamilie der Lycosoidea.
Ähnlichkeiten mit anderen Bananenspinnen
_(10623228224).jpg.webp)
Die Arten Bananenspinnen (Phoneutria) einschließlich der Brasilianischen Wanderspinne sehen sich optisch allesamt sehr ähnlich, weshalb auch hier Verwechslungen möglich sind. Dies trifft auch auf die Färbungen der verschiedenen Arten zu. Die dorsal angelegte braune Grundfärbung etwa ist bei allen Vertretern der Bananenspinnen vorhanden und auch das Besitzen rot erscheinender Cheliceren teilt sich die Brasilianische Wanderspinne mit der Großen Bananenspinne (P. boliviensis), Phoneutria fera und Phoneutria keyserlingi.[4] Innerhalb der Gattung selbst sieht die Brasilianische Wanderspinne den Arten Phoneutria eickstedtae, Phoneutria keyserlingi und Phoneutria pertyi am ähnlichsten. Von diesen unterscheidet sich aber insbesondere das Weibchen der Brasilianischen Wanderspinne durch die deutlich dunklere Färbung am Ventralbereich des Opisthosomas.[6]
Die sicherste Unterscheidungemethode der vier zuvor genannten Arten ist die Unterscheidung ihrer genitalmorphologischen Merkmale. Die Bulbi des Männchens der Brasilianischen Wanderspinne unterscheiden sich von denen der Männchen von Phoneutria keyserlingi und Phoneutria pertyi durch den gerade geformten Embolus mit dem schwach ausgeprägten und prolateral angelegten Kiel. Bei Phoneutria eickstedtae ist die Basis der Emboli viel breiter. Die Epigyne ist bei Phoneutria keyserlingi anders als bei der Brasilianischen Wanderspinne erhöht und das Seitenfeld der Epigyne nahe der Kopulationsöffnung ist bei der anderen Art außerdem gerillt.[7] Von der Epigyne der Weibchen der beiden anderen Arten unterscheidet sich die des Weibchens der Brasilianischen Wanderspinne durch das flache Zentrum und das Fehlen vom basal angelegten Sulci in der lateralen Apophyse der Epigyne.[8] Bei Phoneutria eickstedtae und Phoneutria pertyi ist das Mittelfeld der Epigyne konvex gebaut und die seitliche Apophyse weist Sulci auf.[5]
Ähnlichkeiten mit Arten der Gattung Ctenus und anderen Kammspinnen
%252C_teneral_penultimate_male.jpg.webp)
Die Arten der Bananenspinnen (Phoneutria) einschließlich der Brasilianischen Wanderspinnen können mit den Arten der ebenfalls zur Familie der Kammspinnen zählenden Gattung Ctenus sowie weiteren der Familie verwechselt werden. Die Arten der Gattung Ctenus sind aber deutlich kleiner und lassen sich mitunter durch die Augenstellungen von den Bananenspinnen unterscheiden. Bei der Gattung Ctenus befinden sich die äußeren Augen nicht auf einer gemeinsamen Erhebung und stehen weiter auseinander.[9]
Ein weiteres sicheres Unterscheidungsmerkmal der Bananenspinnen mitsamt der Brasilianischen Wanderspinne zur Gattung Ctenus und anderen Kammspinnen ist das Vorhandensein der dichten Skopulae an den Pedipalpen. Diese Eigenschaft fehlt allen anderen Kammspinnen.[6]
Ähnlichkeiten mit Arten der Gattung Cupiennius

Die Brasilianische Wanderspinne wird überdies wie andere Bananenspinnen (Phoneutria) gelegentlich mit den für den Menschen deutlich harmloseren Vertretern der Gattung Cupiennius aus der Familie der Fischerspinnen (Trechaleidae) verwechselt. Verwechslungen können auch deshalb entstehen, da gelegentlich Arten beider Gattungen auf Fruchtstauden der Dessertbanane (Musa × paradisiaca) in andere Kontinente mitexportiert werden.[10]
Die Brasilianische Wanderspinne hat allerdings wie andere Arten der Gattung eine wesentlich kontrastärmere Grundfärbung und überdies rote Cheliceren (Kieferklauen), eine Eigenschaft, die fast allen Arten der Gattung Cupiennius fehlt. Außerdem übertrifft die Brasilianische Wanderspinne hinsichtlich ihrer Körpergröße alle Arten der Gattung Cupiennius.[11]
Vorkommen
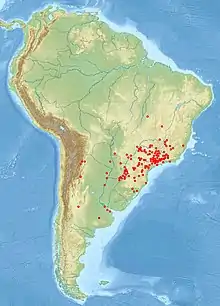
Die Brasilianische Wanderspinne kommt entsprechend ihrer Trivialbezeichnung bevorzugt in Brasilien vor, wo sie in den Bundesstaaten Minas Gerais, Goiás, Mato Grosso do Sul, Espírito Santo, Rio de Janeiro, São Paulo, Paraná, Santa Catarina und Rio Grande do Sul nachgewiesen ist. Außerdem bewohnt die Art die Mata Atlântica und die bewaldeten Gebiete der Cerrados innerhalb Brasiliens.[5] Südlich reicht das Verbreitungsgebiet bis nach Paraguay[12] und den Norden Argentiniens.[5][12] In Uruguay wurde die Brasilianische Wanderspinne überdies durch Bananenexporte eingeschleppt.[4]
Insbesondere Nachweise aus Uruguays Hauptstadt Montevideo und Argentiniens Hauptstadt Buenos Aires gehen vermutlich auf verschleppte und eingebürgerte Tiere zurück, die mittels Bananenimporten in diese Städte gelangten.[12] Durch Unklarheiten in der Synonymie und Namensverwendung (vgl. Abschnitt Systematik) sind ältere Fundortangaben manchmal auf andere Arten zu beziehen.
Lebensräume

Die Brasilianische Wanderspinne bewohnt wie die anderen Arten der Bananenspinnen (Phoneutria) bevorzugt Regenwälder, wo sie sich zumeist auf Bäumen aufhält.[13] Dabei kann die Art jedoch auch am Bodengrund oder in niedriger Vegetation gefunden werden.[14]
Zusammen mit Phoneutria keyserlingi zeigt die Brasilianische Wanderspinne als einzige Art der Gattung eine hohe Anpassungsfähigkeit an menschliche Siedlungen und somit eine gewisse Synanthropie (Bevorzugung menschlicher Lebensräume). So kommen beide Arten sowohl in kleineren Ortschaften als auch in urbaneren Regionen vor. Ebenso sind sowohl Phoneutria keyserlingi als auch die Brasilianische Wanderspinne häufig in Gebäuden anzutreffen.[4]
Einfuhr durch Bananenfrüchte
.jpg.webp)
Es kommt gelegentlich zur unwissentlichen Verschleppung einzelner Individuen der Brasilianischen Wanderspinne durch den Export von in Südamerika geernteten Stauden der Dessertbanane (Musa × paradisiaca). Von daher rührt die sowohl bei den weiteren Vertretern der Bananenspinnen (Phoneutria) und aus dem gleichen Grund bei einzelnen Arten der Gattung Cupiennius mehrdeutige Bezeichnung „Bananenspinne“. Die transportierten Stauden bieten den Spinnen Versteckmöglichkeiten, welche von der tagsüber größtenteils verborgen lebenden Art gerne als solche angenommen werden, wo sie dann übersehen werden können.[15] Allerdings ist die Wahrscheinlichkeit, Exemplare der Brasilianische Wanderspinne an in Europa zu verkaufenden Bananenfrüchten zu finden, sehr gering, da lediglich 0,8 % der dort verkauften Bananenfrüchte aus Brasilien stammen.[16] Auch im deutschen Sprachraum kam es, wenn auch selten, zu Funden der Brasilianischen Wanderspinne, über die in der lokalen Presse berichtet wurde.[16]
Im Allgemeinen sind Funde von Arten der Gattung Cupiennius, insbesondere von C. coccineus und C. getazi an exportierten Bananenstauden häufiger als von jenen der Bananenspinnen mitsamt der Brasilianischen Wanderspinne. Aufgrund der starken Ähnlichkeit der Arten beider Gattungen kommt es dabei nicht selten zu fehlerhaften Identifikationen der jeweiligen Art.[4] Innerhalb der Gattung der Bananenspinnen selbst ist die Brasilianische Wanderspinne neben Phoneutria keyserlingi und der Großen Bananenspinne (P. boliviensis) eine von drei Arten der Gattung, von denen Verschleppungen überliefert sind. Die höchste Wahrscheinlichkeit eines Exports besteht bei Individuen der Großen Bananenspinne, da diese von allen Arten der Gattung das größte Verbreitungsgebiet besitzt und überdies als einzige Art der Gattung auch in Mittelamerika vorkommt. Die Große Bananenspinne ist mit einer Körperlänge von maximal 25 Millimetern allerdings deutlich kleiner als die Brasilianische Wanderspinne und Phoneutria keyserlingi mit Körperlängen von je 30 bis 50 Millimetern, was Verwechslungen dieser Art mit jenen der Gattung Cupiennius jedoch wahrscheinlicher macht. Ferner ist das Gift der Großen Bananenspinne für den Menschen deutlich ungefährlicher als das der beiden anderen Arten.[11]
Lebensweise

Die Brasilianische Wanderspinne ist wie alle Kammspinnen (Ctenidae) überwiegend nachtaktiv und versteckt sich tagsüber oft in Termitenhügeln, dem Blattwerk von Bromeliengewächsen, unter umgestürzten Baumstämmen und Felsen und in Bananenplantagen.[17] Der letztere Aufenthaltsort begünstigt die Verschleppung der Spinne.[13]
Entsprechend ihrem Trivialnamen lebt die Brasilianische Wanderspinne wie alle Arten der Bananenspinnen (Phoneutria) nomadisch und legt keine festen Unterschlüpfe an, sondern nutzt die bereits oben erwähnten und vorhandenen als temporäre Aufenthaltsorte für die Inaktivitätszeit am Tag.[17]
Jagdverhalten und Beutespektrum
Wie alle Spinnen lebt auch die Brasilianische Wanderspinne räuberisch. Ein Spinnennetz zum Jagdzweck wird wie bei allen Kammspinnen (Ctenidae) nicht angelegt.
Beutefang
Das Jagdverhalten der Brasilianischen Wanderspinne entspricht dem anderer Kammspinnen (Ctenidae), und sie jagt dementsprechend als Lauerjäger. Dabei sucht die Art nicht selten in den von ihr bevorzugten Habitaten aktiv nach Beutetieren.[4] In Ihrer Aktivitätszeit in der Nacht unternimmt die Spinne dann auch zu diesem Zweck weitläufige Wanderungen, und ihr Aktivitätsradius kann bis zu 100 Meter betragen.[18]
Bei der Jagd kommen der Vibrations- und der Sehsinn zum Einsatz. Wurde ein Beutetier geortet und ist es in Reichweite, springt die Spinne dieses an und versetzt ihm mittels der Cheliceren einen Giftbiss, ehe es verzehrt wird. In Gefangenschaft wurde bei größeren Beutetieren auch vermerkt, dass die Spinne diese einspann.[19]
Beutespektrum
.jpg.webp)
Die Brasilianische Wanderspinne ist ein opportunistischer Jäger und erlegt alle möglichen Beutetiere, die sie zu überwältigen vermag. Dabei weist die Art aufgrund ihrer Dimensionen ein verglichen mit dem anderer Spinnen großes Beutespektrum auf. Dieses umfasst unter anderem verschiedene Arthropoden, darunter verschiedene Insekten und andere Spinnen.[3] Häufige Beutetiere sind Grillen und Schaben.[20] Ferner ist der Brasilianischen Wanderspinne auch das Erlegen vergleichsweise großer und wehrhafter Gliederfüßer, etwa Skorpionen und Hundertfüßern möglich.[3]

Kleine Wirbeltiere erweitern das Beutespektrum der Art. Dazu zählen kleinere Frösche, Eidechsen,[3] Nagetiere,[21] und Fledermäuse[22]. Darüber hinaus ist das Erlegen von jungen und noch in Brutnesten befindlichen Vögeln und Ratten seitens der Brasilianischen Wanderspinne wie auch bei anderen Arten der Bananenspinnen (Phoneutria) überliefert.[23] Gemieden werden wohl allerdings Tiere, die Gift über ihre Haut absondern können. Dies konnte bei Erbeutungsversuchen der Brasilianischen Wanderspinne bei Baumsteigerfröschen (Dendrobatidae) und giftigen Nachtfaltern aus der Familie der Widderchen (Zygaenidae) beobachtet werden. Diese wurden zwar von der Spinne gepackt, jedoch kurz darauf wieder losgelassen.[3]
Abwehrverhalten

Mit anderen Arten der Bananenspinnen (Phoneutria) teilt die Brasilianische Wanderspinne ihr charakteristisches Drohverhalten, das in dieser Form innerhalb der Familie der Kammspinnen (Ctenidae) einzigartig ist.[6][24]
Die Art gebärdet sich wie die anderen der Gattung bei Störungen recht aggressiv und zieht bei einer Begegnung mit möglichen Prädatoren (Fressfeinden) das Vollführen einer Drohstellung gegenüber einem Rückzug deutlich vor. Bei der Drohstellung stellt sich die Spinne auf die beiden hinteren Beinpaare und richtet den Körper fast senkrecht zum Boden auf. Die beiden vorderen Beinpaare werden in vertikale Richtung erhoben und über dem Körper gehalten. Hier kommen nun die Warnfarben auf der Unterseite der Beine zur Geltung. Gleichzeitig vollführt die Spinne je zu beiden Seiten wiegende Bewegungen mit den erhobenen Extremitäten und richtet sich frontal und dadurch mit den Warnfarben dem Angreifer entgegen. Die Drohgebärde wird seitens der Spinne mit dem Sträuben der Beinstacheln und dem Spreizen der Cheliceren erweitert.[4]
Sollte diese Methode keine Wirkung erzielen, kann sich die Spinne in größter Not auch mit einem Giftbiss verteidigen.[4] Das äußerst wirksame Gift kann auch für den Menschen lebensbedrohliche Auswirkungen haben.
Lebenszyklus
Der Lebenszyklus der Brasilianischen Wanderspinne gliedert sich wie bei anderen Spinnen in mehrere Etappen und ist zudem von den Jahreszeiten abhängig. Er verläuft vom Grundprinzip her wie bei anderen Arten der Bananenspinnen (Phoneutria) und wurde überdies verglichen mit den Lebenszyklen anderer Vertreter der Gattung gut erforscht, was ansonsten lediglich bei der Großen Bananenspinne (P. boliviensis) der Fall ist. Die Entwicklungsstufen beider Arten weisen viele Gemeinsamkeiten auf.
Phänologie
Die Phänologie (Aktivitätszeit) der Brasilianischen Wanderspinne beläuft sich wie bei Regenwaldbewohnern üblich auf das ganze Jahr. Der Schwerpunkt der Aktivitätszeit liegt allerdings in der Trockenzeit ihres natürlichen Vorkommensgebietes. Ausgewachsene Exemplare beider Geschlechter der Brasilianischen Wanderspinne sind bevorzugt von April bis Juni aktiv.[24]
Anlegen eines Spermanetzes
Wie bei anderen Spinnen üblich legt ein geschlechtsreifes Männchen der Brasilianischen Wanderspinne vor der Suche nach einer Geschlechtspartnerin ein sog. „Spermanetz“ an, was in dunklen Bereichen geschieht. Dieses Netz dient der Abgabe von Sperma, das dann wieder aufgenommen wird. Das Netz hat die Form eines kleinen Blattes und nimmt etwa eine halbe Stunde vom Anlegen bis zur Vollendung in Anspruch. Anschließend dauert es etwa 12 bis 15 weitere Minuten, ehe das Männchen das zuvor abgegebene Sperma vollständig in seinen Bulbi aufgenommen hat.[1] Anschließend beginnt es nachts ein Weibchen aufzusuchen.[25]
Balz und Paarung
Das Paarungsverhalten der Brasilianischen Wanderspinne wurde besonders in Gefangenschaft beobachtet, und die Paarungszeit der Art beläuft sich auf die Monate April und Mai. Geschlechtsreife Männchen suchen besonders in dieser Zeitspanne aktiv nach Geschlechtspartnerinnen.[4] Hat ein Männchen ein Weibchen ausfindig machen können, nähert es sich diesem vorsichtig und kommt vor dem Weibchen in einem Abstand, der dem Vierfachen der eigenen Körperlänge des Männchens entspricht, zum Stillstand.[25] Zur Annäherung vollführt das Männchen wie auch andere Angehörige der Überfamilie Lycosoidea einen Balztanz, bei dem es zuerst mit dem Pedipalpen auf den Boden trommelt. Dem Trommeln folgt ein weiteres vorsichtiges Annähern in Richtung des Weibchens. Der nächste Schritt der Balz ist ein Auf- und Abwippen des Opisthosomas, wobei dies nur in Gefangenschaft beobachtet werden konnte.[19] Bei Beobachtungen aus dem Freiland vollführte das Männchen keinen Balztanz, sondern nährt sich dem Weibchen direkt.[26] Beobachtungen, dass sich das Männchen während der Annäherungsversuchs des Weibchens zur Flucht entscheidet, existieren auch nur bei in Gefangenschaft beobachteten Tieren.[19]
Kannibalistisches Verhalten seitens des Weibchens tritt im Allgemeinen nicht auf.[27] Dies konnte lediglich einmalig in Gefangenschaft beobachtet werden.[19]
Vor der Paarung besteigt das Männchen das Weibchen frontal, sodass beide Geschlechtspartner nach der Besteigung in die jeweils entgegengesetzte Richtung blicken. Das Weibchen zieht seine Beine dabei dicht an den Körper und positioniert sie über dem Prosoma. In dieser Position berühren sich die Patellae (Schenkelringe) aller vier Beinpaare des Weibchens fast vollends. Dafür ändert sich das Männchen leicht seitlich, sodass es einen der Bulbi in die Epigyne seiner Partnerin einführen kann.[28] Diese Paarungsstellung ist bei vielen anderen Spinnenarten der Überfamilie der Lycosoidea und anderen frei jagenden Vertretern der Echten Webspinnen ebenfalls präsent.[26]
Beide Partner trennen sich friedlich voneinander. Während sich das Männchen der Brasilianischen Wanderspinne wie bei den anderen Arten der Gattung mehrmals verpaaren kann,[25] vollführt das Weibchen der Art dies in seiner Lebenszeit nur einmalig.[29]
Kokonbau und Schlupf
Von August bis September legt das Weibchen dann wie bei den anderen Vertretern der Gattung seinen ersten Eikokon an.[30] Dieser hat die Form einer Untertasse und hat einen Durchmesser von 30 bis 40 Millimetern und eine Höhe von sieben bis neun Millimetern.[30] Darin werden die Eier in überlappenden Schichten abgelegt. Die Eier sind kugelförmig, gelblich-weiß gefärbt und für Spinnen ungewöhnlicherweise nicht miteinander verklebt. Ihr Durchmesser beträgt gut 1,3 Millimeter. Die Embryonalentwicklung nimmt durchschnittlich 18 bis 23 Tage in Anspruch.[31] Ein Eikokon kann über 3.000 Eier beinhalten. Er wird vom Weibchen stetig mit sich getragen.[4] Dafür hält es ihn mittels seiner Pedipalpen und Cheliceren unter sich. Außerdem wird der Kokon an geeigneten dunklen Plätzen, etwa unter Steinen und Rinde abgelegt. Er wird dort zusätzlich vom Weibchen verteidigt, vor Witterungen geschützt und gleichzeitig der Sonne zur Wärmung entgegengehalten.[30]
Nach dem Schlupf verbleiben die Jungtiere für acht bis zehn Tage in dem Kokon. Anschließend durchbohren sie den Kokon und verlassen ihn.[32] Ein begattetes Weibchen kann hintereinander bis zu vier Eikokons herstellen.[4] Jedoch verschlechtert sich die Qualität jedes folgenden Kokons zunehmend, indem diese nach und nach kleiner und unförmiger werden sowie deutlich weniger befruchtete Eier beinhalten. Dies dürfte mit der steigenden Ermüdung des Weibchens zu begründen sein.[30]
Heranwachsen und Lebenserwartung
Während die Jungtiere den Eikokon verlassen, begeben sie sich von dem Kokonrand, aus dem sie diesen verlassen haben, zu ihrer Mutter. Diese verbleibt auch noch zu diesem Zeitpunkt bei ihren Nachkommen und verhält sich reglos, während sie in einer geduckten Stellung verharrt.[27] Die Jungtiere verfügen bereits über eine räuberische Lebensweise, sobald sie den Kokon verlassen.[4] Sie beginnen sich umgehend vom Muttertier zu trennen und seilen sich dazu mithilfe von Spinnfäden ab. Dabei versuchen sie, in der Vegetation etwa nahegelegene Pflanzenstiele zu erreichen. Für die Ruhephase legen die Jungtiere dann horizontale, lakenartige Gespinste an. Ein solches wird jede Nacht neu angelegt, jeweils etwa fünf Zentimeter über dem vorherigen. Damit befinden sich die Jungtiere bereits nach einigen Tagen einen halben Meter vom Bodengrund entfernt.[27]
Wie bei Spinnen üblich, wachsen die Jungtiere der Brasilianischen Wanderspinne über mehrere Fresshäute (Häutungsstadien) heran.[4] Die in diesem Prozess unbeweglichen Jungtiere fallen in dieser Zeit insbesondere ihren Geschwistern zum Opfer, die bereits eine fortgeschrittene Entwicklung aufweisen. Dies sichert letzteren den Vorteil, eine erste Nahrungsquelle zu erlangen und somit ihr Überleben zu sichern. Die überlebenden Jungtiere beginnen sich ab da fluchtartig zu zerstreuen und beginnen ihre solitäre Lebensweise.[27] Sie durchleben in ihrem ersten Lebensjahr, abhängig von Temperatur und Nahrungsangebot, fünf bis zehn Häutungen. Mit zunehmendem Alter nimmt die Anzahl der Häutungen jedoch ab. Im zweiten Lebensjahr häuten sich die heranwachsenden Spinnen drei- bis siebenmal und im dritten lediglich zwei- bis dreimal. Nach der letzten dieser Häutung, nach der die Spinne demzufolge ausgewachsen ist, tritt auch die Geschlechtsreife ein.[4]
Das Weibchen der Brasilianischen Wanderspinne hat unter natürlichen Umständen eine durchschnittliche Lebensdauer von vier Jahren. Die des Männchens ist mit ungefähr drei Jahren und wenigen Monaten kürzer.[33] Die gesamte Lebensdauer der Art kann bis zu sechs Jahre betragen, wobei dies bislang nur in Gefangenschaft nachgewiesen wurde.[4]
Toxikologie
Toxikologische Analysen konnten die Bestandteile des Giftes der Brasilianischen Wanderspinne sowie deren Wirkung (Toxizität) bisher in Teilen analysieren.
Spinnentoxine
Das komplex aufgebaute Gift der Brasilianischen Wanderspinne enthält neben einigen Enzymen unter anderem neurotoxische (als Nervengift wirkende) Peptide und Proteine (Eiweißkörper). Diese beeinflussen die Ionenkanäle und Rezeptoren des neuromuskulären Systems sowohl von Wirbeltieren als auch von Wirbellosen. Im Giftsekret sind vermutlich bis zu 150 Verbindungen enthalten, von denen bislang 54 dokumentiert wurden. Ferner sind cysteinhaltige arteigene Toxine, sogenannte „PnTxs“ (Phoneutria nigriventer toxins), enthalten. Mehrere Proteine mit hoher Molekülmasse werden vermutet, von denen bislang nur wenige beschrieben wurden. Diese Eigenschaften machen das Gift deutlich wirkungsvoller als etwa die Gifte anderer gefährlicher Spinnen wie der Sydney-Trichternetzspinne (Atrax robustus) oder der Südlichen Schwarzen Witwe (Latrodectus mactans). Ferner kann die vergleichsweise große Brasilianische Wanderspinne Injektionen in achtfacher Menge der wesentlich kleineren Echten Witwen (Latrodectus) verabreichen.[34]
Nomenklatur
Die Nomenklatur der Spinnentoxine der Brasilianischen Wanderspinne erwies sich bisher als problematisch, da in der Vergangenheit dem gleichen Peptid oft mehrere Namen gegeben wurden. Historisch gesehen werden die Toxine der Arten der Bananenspinnen (Phoneutria) basierend auf ihrem Auftreten im Gift annotiert, wenn sie den in den ersten Studien verwendeten Giftreinigungsmethoden folgen, also basierend auf einem bestimmten chromatographischen Schritt und in der Reihenfolge der Elution (Ab-, Herauslösen oder Verdrängen von adsorbierten Stoffen aus Adsorbentien oder Ionenaustauschern) des Toxins in diesem Schritt. In einem Versuch, die Nomenklatur von Peptiden sowohl von Spinnentoxinen als auch den Giftstoffen anderer Tiere zu vereinheitlichen, wurde 2008 von Glenn F. King eine rationale Nomenklatur vorgeschlagen, die das molekulare Ziel, einschließlich der Subtypen des Toxins sowie die Familie, Gattung und Spezies des Tieres berücksichtigt, von der das Toxin ursprünglich stammt.
Gruppierung der Bestandteile
| LD50 (Maus, μg/kg)[35] | |
| Gift, gesamt | 47 |
| PhTx1 | 45 |
| PhTx2 | 1,7 |
| PhTx3 | 137 |
| PhTx4 | 480 |
Durch Gelfiltration kann das Gift in fünf Fraktionen getrennt werden.[36] Drei dieser Fraktionen (P1, P2 und P3) enthalten Proteasen, Moleküle mit hohem Molekulargewicht, die Caseine und Gelatine abbauen können.[37] Die P4-Fraktion enthält die Spinnentoxine, die wiederum in fünf Fraktionen unterteilt werden. Die P5-Fraktion enthält niedermolekulare Substanzen mit Molekulargewichten unter 1 kDa.[36]
Die aus dem ersten Trennungsschritt erhaltene P4-Fraktion kann weiter aufgetrennt werden. Dabei werden fünf neue Fraktionen erhalten, die unterschiedliche Eigenschaften und LD50-Werte aufweisen. Die Fraktionen PhTx1, PhTx2 und PhTx3 zeigen toxische Wirkung auf Säugetiere, während PhTx4 gegenüber Insekten eine höhere Toxizität zeigt als gegenüber Säugetieren. Die fünfte Fraktion PhM zeigt keine letale Wirkung gegenüber Mäusen, wirkt jedoch auf die glatte Muskulatur.[36]
PhTx1 besteht lediglich aus einem Toxin, dem PnTx1, das etwa 0,45 % der Gesamtmasse der Proteine im Gift ausmacht. Mäuse reagierten auf Injektion des Giftes mit Erregung, Schwanzaufrichten und Spastiken, die durch Einwirken auf die Natriumkanäle entstehen.[37][38]
Der PhTx2-Fraktion werden die erregenden Eigenschaften des Giftes, wie Speichelbildung, Tränenfluss, Priapismus (Dauererektion), Krampfanfälle und Spastiken zugeschrieben. Sie besitzt die höchste Toxizität der fünf Fraktionen und wirkt auf die Natriumkanäle. Es konnten mindestens 9 Toxine in dieser Giftfraktion identifiziert werden (PnTx2-1-PnTx2-9). Im Fokus der Forschung stehen vor allem die Toxine PnTx2-5 und PnTx2-6, da sie Priapismus auslösen und somit als Medikamente gegen erektile Dysfunktion in Frage kommen.[39]
In der PhTx3-Fraktion konnten sechs Toxine identifiziert werden (PnTx3-1-PnTx3-6). Die Fraktion wirkt auf Kalium- und Calciumkanäle und verursachen akute schlaffe Lähmung (Acute Flaccid Paralysis, ähnlich wie bei Poliomyelitis).[40] PnTx3-6, auch als Phα1β bekannt, wird als Analgetikum getestet.
Auf Grund ihrer hohen Toxizität gegenüber Insekten wird die PhTx4-Fraktion auch als insektizide Fraktion bezeichnet. Sie zeigt gegenüber Säugetieren nur eine schwache Wirkung, verursacht jedoch in Insekten (Zweiflügler und Rotdeckenkäfer) Hyperaktivität. Es wird vermutet, dass PhTx4 auf das Glutamatsystem wirkt.[40]
Die P5-Fraktion besteht aus niedermolekularen Substanzen. Bis in die 1980er wurde angenommen, dass solche Substanzen keine Rolle in der Toxikologie der Spinnen- und Wespengifte spielen, sondern lediglich Peptide und Proteine toxisch wirken.[41] Im Gift der Brasilianischen Wanderspinne konnte allerdings das Neurotoxin Nigriventrin identifiziert und charakterisiert werden, welches 0,4 % des Giftes ausmacht.[42]
Toxische Wirkungen bei Tieren
Bedingt durch die Forschungen im Rahmen der Toxikologie (Giftkunde) des Gifts der Brasilianischen Wanderspinne, die auch an Versuchstieren durchgeführt wurden, konnten demzufolge auch Resultate der Giftwirkung bei diesen erfasst und dokumentiert werden. Dies geschah ausführlich unter Wolfgang Bücherl im Jahr 1972.
Wirkung bei Hunden
Bei einer subkutanen (in das Gewebe abgegebenen) Zugabe des Gifts bei männlichen Haushunden in kleinen letalen Dosen von 180 bis 200 µg/kg Körgergewicht konnten starke lokale Schmerzen, heftiges Niesen, Tränenfluss, übermäßiger Speichelfluss, Adynamie (allgemeine Erschöpfung bzw. eine ausgeprägte Kraft- und Antriebslosigkeit) oder gravierendere Erschöpfung bis zur Schläfrigkeit, Ataxie (Störungen der Bewegungskoordination), Dyspnoe (erschwerte Atemtätigkeit), Erbrechen, Priapismus (schmerzhafte Erektionen), Ejakulation sowie blutige Exkremente und in einigen Fällen auch das Ableben der Versuchstiere festgestellt werden. Bei Hunden reichen subletale subkutane Dosen im Allgemeinen nicht aus, um Priapismus hervorzurufen. Genauso sind subkutan wirksame Dosen bei intravenöser Injektion sehr toxisch und führen zu einem starken Abfall des Blutdrucks bei Hunden. Mit Ausnahme des Priapismus, der erst in einer wissenschaftlichen Beobachtung 1962 beobachtet wurde, wurden alle oben genannten Effekte bereits bei 1925 stattgefundenen Versuchen gemeldet.[43]
Symptome bei Hunden
Subkutan injiziert löst das Gift bei Hunden große Schmerzen aus und veranlasst diese dazu, fast eine Stunde lang grelle Laute von sich zu geben sowie das injizierte Hinterbein länger als eine Stunde zusammengezogen zu halten. Das spezifische Antivenin (Immunserum bei Tierbissen) neutralisiert die schmerzproduzierende Komponente, wodurch ausgeschlossen werden kann, dass dieser Effekt durch Histamin oder Serotonin im Gift der Spinne verursacht wird. Da der Schmerzfaktor dialysierbar, bzw. eine Blutreinigung möglich ist, muss sein Molekül relativ klein sein. Aus diesem Grund ist es unwahrscheinlich, dass der Faktor ein Bradykinin-Enzym ist.[43]
Bei Hunden sind Niesanfälle eines der ersten Anzeichen einer Vergiftung. Dieses zeitweise auftretende Symptom lässt sich allgemeinen länger als 24 Stunden feststellen. Außerdem treten heftige Anfälle innerhalb der ersten zwei Stunden auf. Dabei wirft der Hund seinen Kopf heftig und unkontrolliert auf den Boden, was häufig in einer Verletzung der Nase und der Lefzen resultiert.[43]
Der durch Gift hervorgerufene Speichelfluss der Hunde äußert sich stark und ähnelt solchem, der durch induziertes Pilocarpin hervorgerufen wird. Demnach tropft über mehrere Stunden nicht geschluckter Speichel aus dem Maul des Tieres. Der Grund dafür sind eine Blockade des Schluckreflexes durch Atropin sowie Dosen von Eresin und Hexamethonium. Obgleich diese Stoffe die Anzeichen einer Vergiftung geringfügig erhöhen, scheinen sie diesen Effekt dennoch nicht weiter zu beeinträchtigen. Subkutane Dosen des Gifts der Brasilianischen Wanderspinne führen bei Hunden aber nicht zu einer Bildung von Speichelfluss, wenn das Gift den Versuchstieren unter Anästhesie mit Chloralose, Chloroform oder Barbituraten verabreicht wird. Dafür erzielt das Gift intravenös (in Venen) abgegeben in gleicher Menge eine hohe Wirkung.[44]
Der Priapismus im Falle einer Induktion des Gifts hat auch ein intermittierendes (zeitweilig aussetzendes) und langanhaltendes Auftreten und wiederholt sich bei Hunden oft über mehrere Stunden (nicht selten über 24). Ist dies der Fall, tritt ein Ödem im Allgemeinen am distalen Endpunkt und korrespondierend mit der Eichel auf. Der Priapismus entwickelt sich schleichend und dauert mehr als 15 Minuten von der Anfangsphase bis zur Endphase an. Ferner erscheint er bei Hunden erst in einem fortgeschrittenen Vergiftungsstadium, bei dem das jeweilige Tier bereits Anzeichen von Adynamie, Ataxie, Schläfrigkeit, Mydriasis (ein- oder beidseitige Weitstellung der Pupille), Sehstörungen, Dyspnoe und Erbrechen aufweist. Priapismus konnte bei anästhesierten (betäubten) Hunden nicht ausgelöst werden. Auch dann nicht, wenn das Gift in größeren Dosen als der subkutan wirksamen Dosis intravenös verabreicht wurde.[45]
Die Priapismus-Induktion scheint nicht von der Erregung höherer Zentren des Nervensystems abzuhängen, da sie bei Hunden provoziert werden könnte, bei denen die Medulla (Kernbereich) am 12. Dorsalnerv geschnitten worden war, was sich unter diesen Umständen bereits 1963 erwies. Im Gegensatz zu Priapismus, der durch Cantharidin ausgelöst wird, scheint der durch das Gift der Brasilianischen Wanderspinne hervorgerufene nicht auf Reflexe zurückzuführen zu sein, die durch eine Reizung der Harnwege verursacht werden. Bei Hunden induziert das Gift Priapismus, bevor es zur Miktion kommt. Außerdem trat Priapismus bei Versuchstieren auf, bei denen der Urinfluss durch die Harnröhre durch Implantation beider Harnleiter in die Haut verhindert wurde. Der Priapismus wird demzufolge nicht durch die Einleitung von Giftlösungen durch die Harnröhre in die Blase induziert.[46]
Bei Hunden induziert das Gift während oder nach dem Einsetzen des Priapismus eine Ejakulation (Ausspritzung der Samenflüssigkeit). Der genaue Auslöser dafür scheint anders als beim Menschen auf die Erregung anderer Organe als der Bläschendrüse zurückzuführen zu sein, da dieses Organ beim Hund nicht vorhanden ist.[46]
Wirkungsstärke bei Hunden
Das Gift der Brasilianischen Wanderspinne wirkt bei Hunden ungefähr viermal giftiger als bei Mäusen. Sollte an diesen 200 µg/kg Gift subkutan verabreicht werden, überleben diese eventuell mehrere Stunden, wobei während der schweren Vergiftungsphase des Tieres Niesanfälle, Speichelfluss und Priapismus auftreten. Kleine intravenös injizierte Giftdosen resultieren bei Hunden in einem starken Abfall des Blutdrucks. Der Anteil an Histamin in dem verabreichten Gift ist in diesen Dosen jedoch zu niedrig, um den Blutdruck zu beeinflussen.[47]
Es wurde 1942 beobachtet, dass das Gift von der Südlichen Schwarzen Witwe (Latrodectus mactans) bei Hunden eine Tachyphylaxie (schnelle Toleranzentwicklung gegenüber Arzneistoffen) hervorruft, was jedoch beim Gift der Brasilianischen Wanderspinne weder von kurzer Dauer noch bei lang anhaltenden und intermittierenden Effekten nicht nachgewiesen werden konnte. Letztgenannter Aspekt würde über mehrere Stunden nicht auftreten, wenn eine Tachyphylaxie vorhanden wäre. Ferner zeigten Hunde, die sich erst von diesen Auswirkungen einer Injektion des Gifts der Spinne erholt hatten, eine identische Reaktion auf das Gift, wenn eine abweichende Dosis bei diesen injiziert wurde. Dadurch ist wahrscheinlich, dass die Tachyphylaxie nicht durchgehend durch Spinnengifte ausgelöst wird. Dies deutet auch auf die unterschiedlichen molekularen Strukturen der beteiligten Wirkstoffe hin.[47]
Wirkung bei Mäusen
Der bei Hunden durch das Gift der Brasilianischen Wanderspinne hervorgerufene Priapismus konnte auch bei Labormäusen mit Albinismus 1963 erstmals nachgewiesen werden. Ferner zeigten die Mäuse bei der Injektion des Gifts alle auch bei Hunden auftretenden und hier leicht erkennbaren Symptome mit Ausnahme von Tränenfluss, Mydriasis, Niesanfällen, Erbrechen und Ejakulationen auf, wobei diese jedoch bei Mäusen schwerer zu verfolgen sind. Mäuse erwiesen sich vermehrt als geeignete Versuchstiere, um das Gift der Brasilianischen Wanderspinne zu erforschen, was eine qualitative und quantitative Untersuchung der Giftwirkung bei geringen angewandten Mengen des Gifts ermöglicht. Ein weiterer Vorteil besteht darin, dass dasselbe Tier über einen Zeitraum von 30 Tagen für mehrere Tests verwendet werden kann, bevor die Immunisierung einen quantitativen Einfluss auf Beobachtungsanalysen zeigt. Dadurch ist eine Forschung des Gifts auf biochemischer Ebene möglich. Es wurde ermittelt, dass bei der Untersuchungen des Spinnengifts anhand von Mäusen die am besten auszuwertenden Resultate erzielt werden, sollte deren Gewicht zwischen 20 bis 25 Gramm liegen. Befindet sich das Gewicht des Versuchstiers unter diesem Wert, treten vermehrt Giftwirkungen im Gegensatz zu Priapismus auf.[48]
Zwar sind Schmerzreaktionen bei Mäusen schwerer nachweisbar als bei Hunden, doch werden diese bei letzteren Tieren durch Konditionierung des nach außen gerichteten Hinterbeines deutlich. Außerdem beißen die Mäuse in die eigene Körperstelle, in die das Gift zuvor injiziert wurde. Diese Manifestationen sind nach dem Einsetzen der distentiven (ausdehnenden) Lähmung nicht einsehbar.[48]
Der bei Mäusen eintretende Speichelfluss als Reaktion auf das Gift lässt sich bei diesen Tieren leicht beobachten. Zunächst sammeln sich am Mund kleine Speichelblasen an. Nach einiger Zeit und abhängig von der verabreichten Menge an Gift können die Tiere später einen großen Teil bis fast das gesamte Fell mit Speichel benetzt haben. Kleine Giftdosen lösen bei Mäusen Speichelfluss aus, ohne, dass bei diesen Anzeichen einer Vergiftung sichtbar werden. Bei größeren Giftmengen tritt Speichelfluss vor den toxischen Manifestationen auf und ist auch während dieser präsent. Die mittlere effektive Dosis (ED50) für den Speichelfluss bei Mäusen beträgt 0,43 mg/kg Körpergewicht.[48]
Die Wechselbeziehung zwischen den verabreichten Giftmengen und dem daraus resultierenden Priapismus bei Mäusen bleibt erhalten, bis ein Maximum der Reaktion bei einer Gruppe von Tieren erreicht ist. Ist dies erreicht, nimmt die Anzahl an den Versuchstieren, bei denen der Priapismus auftritt, ab oder sie sterben, bevor er überhaupt einsetzen kann. Demnach lässt sich die Wirkung des Gifts der Brasilianischen Wanderspinne bei Mäusen am ehesten als funktioneller Antagonismus-Priapismus beschreiben. Ähnlich wie im Falle des Speichelflusses können kleine Dosen Gift bei Mäusen Priapismus ohne toxische Manifestationen hervorrufen und unterscheiden sich somit diesbezüglich von den Hunden, die abweichende Reaktionsmuster zeigten. Die ED50 des Priapismus bei Mäusen beträgt 0,25 mg/kg Körpergewicht.[49]
Die toxischen Reaktionen auf Gift bei Mäusen sind Atemnot, Entkräftigungen, starke Lähmung oder der Tod. Die Letale Dosis (LD50) beträgt hier 0,76 mg/kg Körpergewicht. Rohe Giftproben führen bei Mäusen zu einer starken Lähmung der Hinterbeine und des Schwanzes. Letzterer verbleibt während einer bestimmten Zeit auf dem Rücken des Tieres in einer gebeugten Stellung. Eine Lähmung der Hinterbeine des jeweiligen Versuchstiers tritt auf, nachdem bei diesem der Speichelfluss und der Priapismus bereits auftreten. Die Lähmung dauert bis zum Ableben der Maus an, die sich zu dem Zeitpunkt bereits in einer vorzeitigen Art von Totenstarre befindet.[50]
Wirkung bei anderen Nagetieren
Bei Versuchen mit Meerschweinchen waren fast alle Reaktionen zu verzeichnen, die auch bei Hunden und Mäusen beobachtet wurden. Jedoch ist die Wirkung des Gifts auf Meerschweinchen aufgrund der höheren Menge an benötigtem Gift zum Erreichen dieser Reaktionen weniger untersucht als bei Mäusen. Allerdings macht die gallertartige Konsistenz des Samens der Meerschweinchen diese sehr geeignet für die Untersuchung des Effekts der Samenejakulation, die 1963 untersucht wurde.[51] Dazu wurden zwei Polypeptide aus dem Gift der Brasilianischen Wanderspinne getrennt, die das Ileum (Krummdarm) des jeweiligen Meerschweinchens kontrahieren, bzw. das Zusammenziehen von dessen Muskeln bewirken.[52]
Im Gegensatz zu den zuvor genannten Tieren zeigten Ratten und Kaninchen eine hohe Resistenz auf das Gift der Spinne. Bei einer verabreichten Dosis des Gifts von 500 Mikrogramm (µg) an Ratten mit einem Körpergewicht von 150 Gramm kam es zu einer Sekretion von deren Tränendrüsen, während eine an 2000 Gramm schweren Kaninchen induzierte Giftmenge von 1000 Mikrogramm einen mäßigen Speichelfluss und leichte Vergiftungsanzeichen hervorruft.[51]
Möglicher Nutzen des Giftes für den Menschen
Im wirkungsvollen Gift der Brasilianischen Wanderspinne wird vermehrt ein dem Menschen zugute kommender Nutzen vermutet, da darin befindliche Bestandteile sowohl für die Pharmazie als für die Landwirtschaft Vorteile erbringen können.
Nutzen als Schmerzstiller
Das im Gift der Brasilianischen Wanderspinne enthaltene Neurotoxin Phα1β hat ein großes Potential als Mittel als Analgetikum (Schmerzstillung), da es bei Verabreichung zu einer Schmerzunempfindlichkeit führt. Insbesondere bei neuropathischen Schmerzen ist es hilfreich.[53] Die Wirkung des Neurotoxins wurde 2014 mithilfe von Laborratten erforscht. Injektion des Toxins konnte den Schmerz durch chronische Verengungsverletzung des Ischiasnervs der Ratten verringern. Die mechanische Schmerzschwelle reduzierte sich um etwa das 2,5-fache. Die Nozizeption (objektive Grundlage des Schmerzempfindens) könnte über 14 Tage reduziert werden. Eine einzelne Injektion von Phα1β in einer Menge von 30 oder 100 pmol reduzierte die neuropathische Nozizeption zwischen einer und sechs Stunden nach der Verabreichung, ohne, dass nachweisbare Nebenwirkungen sichtbar wurden. Zu einem ähnlichen Resultat führte eine kontinuierliche Infusion von Phα1β von 60 pmol/μL/h für sieben Tage. Hier blieb die schmerzstillende Wirkung über den gesamten Zeitraum der Infusion erhalten.[53]
Nutzen als Potenzmittel
Das Neurotoxin Tx2-6 könnte als Potenzmittel dienen. Das Toxin wurde 2008 sowohl anhand von Ratten als auch Mäusen getestet. Durch Gabe des Gifts konnte bei den Versuchstieren eine Erektion induziert werden. Der Wirkungsmechanismus ist noch nicht bekannt, jedoch könnte das Freisetzen des Neurotransmitters Stickstoffmonoxid eine Rolle spielen.[54]
Nutzen für die biologische Schädlingsbekämpfung
Weitere 2008 getätigte Forschungen befassten sich mit einem möglichen Nutzen des Gifts der Brasilianischen Wanderspinne im Rahmen der biologischen Schädlingsbekämpfung. Dabei wurde das gegen Insekten wirkende Peptid TX4 (1-6) codiert (verschlüsselt) und in Fusion mit Maltose-Bindungsprotein (MBP) vervielfältigt sowie in Escherichia coli exprimiert (verdeutlicht). Das dadurch geschaffene und affinitätsgereinigte Protein MBP-GlyTX4 wurde mit dem Faktor-Xa-Inhibitor gespalten und für einen Bioassay (Untersuchung der Wirkung von chemischen Stoffen auf lebende Organismen) gegen den Eulenfalter Spodoptera frugiperda verwendet sowie dessen Wirkung auf die Immunisierung von Kaninchen untersucht.[55]
GlyTX4-Proteine mit einem Gewicht von fünf Mikrogramm, die in das Mixocoel (Leibeshöhle) von Raupen und Imagines (ausgewachsenen Tieren) des Falters injiziert wurden, führten bei diesen unmittelbar nach der Injektion zu zitternden und unkoordiniert wirkenden Bewegungsmustern. Außerdem verendeten alle Imagines nach 12 Stunden. Die Raupen waren nach zwei Tagen durch das Protein gelähmt und die die Epidermis (äußere Zellhaut) nahm eine dunkelbraune Färbung an. Darüber hinaus wurde das Entwicklungsstadium der Raupen um zwei Wochen verlängert.[55]
Alternativ wurden in der Versuchsreihe Maisblattscheiben mit 15 Mikrogramm des mit dem Faktor-Xa-Inhibitor gespaltenen rekombinanten Proteins aufgenommen und als Nahrung für Larven verwendet. Dabei starben alle Larven etwa 30 Minuten nach deren Verzehr. Polyklonale Antikörper gegen MBP-GlyTX4 waren wirksam zur Erkennung des Maltose-Bindungsprotein und GlyTX4 in Ganzzellextrakten von Escherichia coli, der das rekombinante Protein exprimierte.[55]
Giftdrüsen
Wie alle Spinnen, die Gift produzieren und außerdem die überwiegende Mehrheit dieser Ordnung darstellen, verfügt die Brasilianische Wanderspinne über Giftdrüsen, die der Produktion und der Speicherung des Gifts dienen. Bei diesen handelt es sich, wie bei Echten Webspinnen (Araneomorphae) üblich, um ein Paar dieser Drüsen, die sich im Prosoma befinden.[56]
Die Giftdrüsen der Brasilianischen Wanderspinne sind zylindrisch und bauchig geformt. Ein zylindrisches Erscheinungsbild besitzen überdies die Giftdrüsen der Labyrinthspinne (Agelena labyrinthica) aus der Familie der Trichterspinnen (Agelenidae) sowie bei der zur Familie der Fischerspinnen (Trechaleidae) zählenden Großen Wanderspinne (Cupiennius salei), während die Giftdrüsen der Spinnenart Loxosceles intermedia innerhalb der Familie der Sechsäugigen Sandspinnen (Sicariidae) mit denen der Brasilianischen Wanderspinne ihre bauchige Form teilen.[57]
Struktureller Aufbau der Giftdrüsen
Der strukturelle Aufbau der Giftdrüsen der Brasilianischen Wanderspinne wurde 2008 näher erforscht. Diese sind äußerlich von einer doppelten Schicht gestreifter Muskeln bedeckt und innerlich von einer Basallamina (Proteinschicht unter der Basis der Epithelzellen) umgeben, die zur Stützung des sekretorischen Epithel dient. Diese gestreiften Muskelzellen weisen je mehrere Nucleolen, bzw. Kernkörperchen auf, die längs ausgerichtet und durch Aktinfasern strukturiert werden. Mittels Phalloidin konnte ermittelt werden, dass die Struktur der Muskeln der Struktur von denen anderer Tiere ähnelt.[58]
Bemerkenswert ist die spiralförmige Anordnung der gestreiften Muskelfasern. Diese Anordnung der äußeren Muskelfasern könnte die Kontraktionsbewegung der Giftdrüse bewirken, während die Spinne Gift abgibt. Eine ähnliche Doppelschicht aus gestreiftem Muskel ist auch bei L. intermedia vorhanden. Bei dieser Spinnenart besteht das Muskelgewebe, das die Giftdrüsen bedeckt, aus zwei Muskelbündeln mit einer großen Anzahl von zusammen ausgerichteten kontraktilen (sich zusammenzuziehen) Mikrofilamenten (fadenförmige Protein-Strukturen). Dies legt die Vermutung nahe, dass Struktur und Funktion der Muskelfasern in den Giftdrüse bei verschiedenen Spinnenarten variieren. Bei der Großen Wanderspinne etwa, deren Giftdrüsen denen der Brasilianischen Wanderspinne ähneln, verlaufen die Muskelfasern ebenfalls in Längsrichtung und bilden eine schwache Spiraldrehung, während die Giftdrüsen der Südlichen Schwarzen Witwe (Latrodectus mactans) aus der Familie der Kugelspinnen (Theridiidae) mit gänzlich längsverlaufenden Fasern umhüllt sind. Im Gegensatz dazu sind bei den Giftdrüsen der Labyrinthspinne eine dicke Schicht gestreifter Muskeln erkennbar, die von einer dünnen Tunica adventitia (aus Bindegewebe und elastischen Fasern bestehende äußere Wand von Hohlorganen) bedeckt sind, die die Muskelbündel miteinander und um die Basallamina bindet und Fasern enthält, die an Kollagene (Strukturproteine des Bindegewebes) erinnern und in einem kreisförmigen Muster ausgerichtet sind.[58]
Ein weiteres Eigenmerkmal der Giftdrüsen der Brasilianischen Wanderspinne, das bisher bei keiner anderen Spinnenart nachgewiesen wurde, ist das Aufweisen von porenartigen Öffnungen zwischen den Muskelfasern, die die Giftdrüsen umschließen. Diese porenartigen Strukturen deuten auf eine mögliche neuronale Kommunikation zwischen Innen- und Außenraum hin, die zur Kontraktion der Giftdrüsen beiträgt. Auch gilt das Vorhandensein axonartiger Strukturen zwischen den Muskelfasern in der Drüsenoberfläche als bemerkenswert. Diese Strukturen enden in je einem synaptischen Vesikel und helfen mit großer Wahrscheinlichkeit bei der neuronalen (Neuronen, bzw. Nervenzellen betreffenden) Reaktion auf die sekretorische Stimulation der Giftdrüsen. Das sekretorische Epithel einer Giftdrüse dringt in das Lumen der Drüse ein, trägt zur Vergrößerung der sekretorischen Oberfläche bei und nimmt auch eine höhere Anzahl von sekretorischen Zellen auf. Diese Eigenschaft tritt auch bei den Giftdrüsen vieler anderer Spinnen, darunter der Labyrinthspinne, der Großen Wanderspinne und L. intermedia auf.[58]
Transmissionselektronenmikroskopien belegten, dass die Granula (körnchenförmige Einlagerungen) der Sekretionszellen in den Giftdrüsen der Brasilianischen Wanderspinnen ein heterogenes Färbungsmuster in den histologischen Schnitten und unterschiedliche Elektronendichten zeigt. Neben den sekretierten Granula konnten innerhalb der sekretorischen Vesikel Gruppen mit einer großen Menge dichter Partikel beobachtet werden. Diese Resultate lassen vermuten, dass verschiedene Substanzen von den einzelnen Zellen ausgeschieden und dann in das Lumen der Drüse zugefügt werden können, damit das Gift entsprechend durch Mischung der Stoffe gebildet werden kann. Eine andere Möglichkeit wäre, dass mehrere sekretorische Vesikel in verschiedenen Reifungsstadien in einer einzelnen sekretorischen Zelle gleichzeitig befindlich sind.[58]
Die 2008 durchgeführten Studien im Bereich der Histologie (Gewebelehre) erwiesen mindestens säulenförmige und subquaderförmige Zellen als identifizierbare Zelltypen. Die Säulenzellen wiesen den Kern in apikaler Position auf und wären ausgereift genug, Giftstoffe abzugeben. Die subquaderförmigen Zellen sind dagegen in der Nähe der Basallamina konzentriert. Ähnlich dazu konnten mittels der Rasterelektronenmikroskopien unterschiedliche Zelltypen analysiert werden. Bei diesen handelt es sich möglicherweise um regenerative unreife Zellen, die bei folgender Reifung eine säulenförmige Gestalt annehmen und anschließend als Ersatz für abgestorbene Zellen im Epithel dienen. Diese Möglichkeit wurde erstmals 1971 erwogen, als Zellinseln in der Basis des säulenförmigen Epithels beschrieben wurden. Dabei wurden die Basalzellen als dazu fähig betrachtet, sich nach ihrer Degeneration zu Epithelzellen zu entwickeln und diese zu ersetzen. Dadurch würde der Sekretionsprozess ununterbrochen aufrechterhalten werden. Diese Hypothese ließ sich aber nie fest bestätigen.[58]
Beobachtungen von histologischen Schnitten und Rasterelektronenmikroskopien erwiesen, dass die Sekretionszellen in den Giftdrüsen der Brasilianischen Wanderspinne als komplexen Strukturen erscheinen. Bei diesen handelt es sich um Sekretionseinheiten, die an der Basis der Drüsen in der Nähe der Muskelschicht entstehen und sich in den zentralen Bereich bis zur Drüsenschicht sowie in den zentralen Bereich bis zu den Lumen der Drüsen erstrecken. Bei den aus Cytoplasma bestehenden Teilen der Epithelzellen wurden Plasmamembranen mit Invaginationen (Einstülpungen) und großen Mengen an sekretorischen Vesikeln deutlich. Ähnliches wurde überdies bei der Labyrinthspinne, der Großen Wanderspinne und L. intermedia beobachtet.[59]
2008 konnte während des „Melkens“ des Versuchstiers, bei dem von diesem dreimal das Abgeben von dessen Gift erfolgte, zumindest beim ersten „Melkprozess“ möglicherweise ein holokriner (Sekrete absondernder und dabei die Zellen der Drüsen auslösender) Sekretionsmechanismus beobachtet werden, da Nucleolen mittels Konfokalmikroskopien im Gift gesichtet wurden, deren Erkennung wiederum durch Färbung mit 4′,6-Diamidin-2-phenylindol (DAPI) möglich war. Da bei den darauf folgenden „Melkprozessen“ allerdings keine Kerne im Gift der Spinne sichtbar wurden, sind alle drei dieser Prozesse kein Indiz dafür, dass der Sekretionsmechanismus weder ausschließlich holo- noch merokrin (einen Teil des Zellinhaltes als Sekret abgebend und somit teilsezernierend) ist.[60]
Volumen der Giftdrüsen
Das Füllvolumen der Giftdrüsen sowie die Menge des unter Narkose abgenommenen Gifts der Brasilianischen Wanderspinne ist vom Alter und vom Geschlecht der Spinne abhängig. Diese Eigenschaften wurden 2002 anhand von 72 Spinnen unterschiedlichen Geschlechts und Stadiums untersucht. Dabei konnten auch unterscheide in der Toxizität der Gifte festgestellt werden. Untersucht wurden Gruppen von 6 bis 21 Tieren. Zur besseren Vergleichbarkeit von Männchen und Weibchen wurde eine Gruppe kleinerer Weibchen untersucht, die eine ähnliche Größe wie die Männchen aufwiesen.
| Stadium und Geschlecht | Anzahl der untersuchten Spinnen | Länge des Prosomas (mm) | Giftmenge (μl) | Giftgewicht (μg) | LD50 (Maus, mg/kg) |
| Ausgewachsenes Weibchen | 21 | 15,8±0,6 | 11,1 | 1.079 | 0,63 |
| Junges Weibchen | 17 | 13,5±0,6 | 6,4 | 548 | |
| Junges Weibchen | 16 | 11,5±0,7 | 2,5 | 296 | |
| Ausgewachsenes Weibchen mit Eikokon | 6 | 13,9±1,7 | 3,2 | 462 | 0,61 |
| Ausgewachsenes Männchen | 12 | 11,8±2,7 | 2,9 | 386 | 1,57 |
Unterschiede des Giftes je nach Geschlecht und Stadium
Die Zusammensetzung des Gifts der Brasilianischen Wanderspinne unterscheidet sich abhängig von Geschlecht und Stadium der Spinne. 2002 wurden bei Untersuchungen über die mögliche Abgabemenge mehrerer Exemplare Unterschiede in der Zusammensetzung des Giftes entdeckt. Mithilfe der Hochleistungsflüssigkeitschromatographie können 21 Peaks im Gift weiblicher Spinnen und 18 im Gift von Weibchen mit Eikokons und Männchen identifiziert werden.[62] Eine methodische Ursache für diese Unterschiede kann die Kontamination des Giftes durch Speichel sein. Bei der Giftentnahme durch Elektrostimulation ist eine Kontamination mit Speichel und darin vorhandenen Verdauungsenzymen nicht unwahrscheinlich.[63][64] Proteasen, die so in das Gift gelangen, können Peptide abbauen und so die Analyseresultate verfälschen.[65][66] Allerdings wiesen bei den 2002 durchgeführten Versuchen, ausgenommen die Giftproben männlicher Tiere, alle ein ähnliches SDS-PAGE-Bandenmuster auf. Das Gift der männlichen Spinnen weist spezifische Proteine im Gewichtsbereich von 66 und 200 kDa auf. Abbauprodukte durch proteolytische Enzyme würden deutlich kleinere Massen aufweisen, was darauf hinweist, dass in dieser Studie kein methodischer Fehler begangen wurde. Ferner wurde über Proteine, die nur im Gift männlicher Spinnen nachweisbar sind, bereits berichtet.[65] Dies deutet darauf hin, dass die unterschiedliche Proteinzusammensetzung des Gifts männlicher Individuen der Brasilianischen Wanderspinne nicht auf eine Kontamination durch proteolytische Enzyme zurückzuführen ist.[61]
Die LD50-Werte des Gifts von Weibchen mit und ohne Eikokons erweisen sich bei Mäusen als fast 2,5 mal kleiner als die des Gifts von Männchen.[65] Die Neurotoxizität beim Gift von Männchen weist einen 1,4 mal so hohen LD50-Wert auf wie das Gift von Weibchen. Daraus lässt sich schließen, dass die Zusammensetzung des Gifts je nach Geschlecht der Spinne variiert und dass das Gift weiblicher Individuen vermutlich eine höhere Menge der für Mäuse toxischen Komponenten enthält.[67] Aus SDS-PAGE-Analysen ergibt sich, dass ein variabler Prozentsatz an Proteinen mit niedrigem Molekulargewicht im Bereich von 7 bis 16 kDa zur Variation der LD50-Werte zwischen den Giften männlicher und weiblicher Exemplare der Brasilianischen Wanderspinne führt. Proteine mit solch einem niedrigen Molekulargewicht spielen eine wichtige Rolle für die tödliche Wirkung des Giftes gegenüber Mäusen.[68] Bei Untersuchungen an Termiten konnte festgestellt werden, dass das Gift der männlichen Spinnen im Vergleich zu dem des Weibchens eine längere Zeit benötigt, um Verhaltensauffälligkeiten oder Tod der Termiten hervorzurufen.[69] Diese geschlechtsspezifische Variation der Gifte könnte durch eine unterschiedliche Verteilung der insektiziden Komponenten in der PhTx4-Fraktion hervorgerufen werden.[70][71] Da 70 % aller nachgewiesenen Bissunfälle der Brasilianischen Wanderspinne von Männchen verursacht werden, die jedoch dreimal niedrigere Giftmengen abgeben, welche zudem eine geringer Toxizität aufweisen, verlaufen viele Unfälle glimpflich. So ist keine Anwendung von Gegengiften erforderlich. Dies ist insbesondere in Anbetracht der möglichen allergischen Reaktionen auf das Gegengift ein wichtiger Aspekt bei der medizinischen Behandlung von durch die Brasilianische Wanderspinne ausgelösten Bissunfällen.[67]
Studienabhängige Unterschiede in den bestimmten LD50-Werten können neben den geschlechts- und stadiumsabhängigen Unterschieden auch auf geografische Unterschiede der Giftzusammensetzung hindeuten.[67][72] Eine systematische Untersuchung dessen steht noch aus.
Systematik
Die Systematik befasst sich im Bereich der Biologie sowohl mit der taxonomischen (systematischen) Einteilung als auch mit der Biologie und mit der Nomenklatur (Disziplin der wissenschaftlichen Benennung) von Lebewesen und damit auch der der Brasilianischen Wanderspinne.
Der wissenschaftliche Artname nigriventer stammt aus der lateinischen Sprache und bedeutet übersetzt etwa „schwarze Ventralseite“, was von der dunklen Unterseite der Brasilianischen Wanderspinne rührt.
Beschreibungsgeschichte
Die Art wurde, als Ctenus nigriventer 1891 durch Eugen von Keyserling, nach einem von Hermann von Ihering im brasilianischen Bundesstaat Rio Grande del Sul gesammelten Weibchen, erstbeschrieben. 1936 transferierte der brasilianische Arachnologe Cândido Firmino de Mello-Leitão die Art, als Phoneutria nigriventer in die von Maximilian Perty bereits 1833 beschriebene Gattung der Bananenspinnen (Phoneutria), in der sie seitdem verblieb. Die Abgrenzung von der Gattung Ctenus war von der Forschung bis zur Revision durch Mello-Leitão als zweifelhaft angesehen worden. Sie wurde in der neueren Forschung aber bestätigt.[2]
Die Abgrenzung und Umschreibung der Arten innerhalb der Gattung ist bis in jüngste Zeit unklar und umstritten gewesen. Wolfgang Bücherl hielt Phoneutria nigriventer irrtümlich für ein Synonym von Phoneutria fera. Die heute wieder als valide angesehenen Arten Phoneutria keyserlingi und Phoneutria pertyi wurden bei einer Revision 2001 mit der Brasilianischen Wanderspinne synonymisiert,[73] diese Synonymie aber 2007 wieder aufgehoben.[74] Die Arten werden vor allem nach Genitalmerkmalen (Details in der Gestalt der weiblichen Epigyne und des männlichen Embolus) unterschieden. Daneben ist ein Färbungsmerkmal charakteristisch: Bei Phoneutria nigriventer ist das Opisthosoma auf der Ventralseite einheitlich braun bis schwarz gefärbt, bei den meisten verwandten Arten heller.
In der heutigen Fassung werden folgende Namen als Synonyme von Phoneutria nigriventer angesehen:[75] Ctenus rufichelis Mello-Leitão, 1917 (= Phoneutria rufichelis), Ctenus paca Mello-Leitão, 1922 (= Phoneutria paca), Ctenus luederwaldti Mello-Leitão, 1927 (= Phoneutria luederwaldti).
Innere Systematik

Die phylogenetischen Stellungen einzelner Arten der Bananenspinnen (Phoneutria) ist bisher in Teilen anhand fünf Arten erforscht worden. Dabei konnten zwei Artengruppen aufgestellt werden. Eine davon ist die der Brasilianischen Wanderspinne mitsamt ihrer am nächsten verwandten Art und somit ihrer Schwesterart, der Großen Bananenspinne (P. boliviensis). Die Andere ist die Phoneutria reidyi-Artengruppe und umfasst neben dieser die Arten Phoneutria bahiensis und Phoneutria fera. Folgendes Kladogramm soll die nähere Beziehung der fünf Arten zueinander verdeutlichen:[76]
| Bananenspinnen (Phoneutria) |
| ||||||||||||||||||||||||
Brasilianische Wanderspinne und Mensch

Die Brasilianische Wanderspinne gilt zusammen mit der Art Phoneutria fera als die giftigste Art der Gattung und gemeinsam mit dieser als die wohl giftigste Spinne überhaupt. Ihr Gift macht die Art zu einer der wenigen, deren Wirkung auch für einen gesunden erwachsenen Menschen lebensgefährliche Folgen haben kann. Der Ruf der Art fällt verschieden aus, sie wird aber nicht selten aufgrund der von ihr ausgehenden Gefahr gefürchtet. Auf andere Personen übt die Spinne mitunter wegen ihrer Gefährlichkeit und auch ihrer für Echte Webspinnen (Araneomorphae) stattlichen Dimensionen eine gewisse Faszination aus. Überdies gelten die Brasilianische Wanderspinne und darunter insbesondere ihr Gift mittlerweile als bedeutende Forschungsobjekte, was mit deren Vielfältigkeit zu begründen ist.
Gefährlichkeit
Bedingt durch ihre recht aggressive Wesensart, hohe Agilität und Giftigkeit geht von der Brasilianischen Wanderspinne eine nicht zu unterschätzende Gefahr für den Menschen aus. Dies liegt vor allem an der relativ hohen Wahrscheinlichkeit eines Zusammentreffens zwischen Mensch und Spinne, da diese im Gegensatz zu der ähnlich giftigen, aber hauptsächlich fernab menschlicher Siedlungsräume verbreiteten Phoneutria fera vorwiegend in dicht besiedelten Bundesstaaten wie São Paulo, Paraná, Santa Catarina und Rio Grande do Sul lebt. Dabei dringen die Spinnen auch in menschliche Behausungen ein, wo sie tagsüber Unterschlüpfe aufsuchen und sich beispielsweise in Wäschekisten, Schränken oder Haushaltsutensilien verstecken, wo sie unbemerkt bleiben können. Bei Störungen oder für die Spinne unerwarteten Annäherungen kann sie sich bedroht fühlen und den vermeintlichen Angreifer beißen. Besonders während der Paarungszeit kommt es zu Bissen durch männliche Tiere, da diese bei der Suche nach Weibchen gehäuft in Häuser gelangen.[77]
Die Gefährlichkeit der Brasilianischen Wanderspinne für den Menschen wird allerdings auch überschätzt. Todesfälle haben heute stark abgenommen, da in Brasilien mittlerweile ein wirksames Gegengift zur Verfügung steht. In den Jahren zwischen 1926 und 1996 starben in Brasilien im Durchschnitt 14 Menschen an Spinnenbissen, die der Brasilianischen Wanderspinne zugerechnet werden. Heute kommt es in den vier oben aufgelisteten Bundesstaaten zu etwa 1200 bis 1300 dokumentierten Bissunfällen bei 0–3 Todesfällen jährlich; in ca. 3–4 % der Fälle ist die Gabe des Gegengiftes notwendig.[19] Die in deutschsprachigen Ländern gängige Annahme, dass die Art über Bananentransporte häufig nach Europa gelange, ist nicht zutreffend. Von den Ländern des Verbreitungsgebiets exportiert lediglich Brasilien Bananen in die Europäische Union, und nur 0,8 % der Bananen auf deutschen Märkten stammen aus Brasilien, was die Wahrscheinlichkeit der oft befürchteten Einfuhr senkt. Bei den in Bananenkisten tatsächlich gefundenen Spinnen handelt es sich meist entweder um heimische Spinnenarten, die während des Transports zwischen die Früchte gelangen, oder um andere exotische Spinnen, etwa ähnlich aussehende Vertreter der Kammspinnen oder der Überfamilie Lycosoidea, beispielsweise die Große Wanderspinne (Cupiennius salei), oder auch andere, meist weniger giftige Arten der Bananenspinnen (Phoneutria). Entsprechend selten sind Bissunfälle mit Phoneutria nigriventer in Europa.[16]
Bissunfälle und Behandlung
.jpg.webp)
Die Brasilianische Wanderspinne besitzt wie alle Arten der Bananenspinnen (Phoneutria) eine verglichen mit der vieler anderer Spinnenarten sehr hohe Aggressivität oder Abwehrbereitschaft.[18] Dadurch und wegen der hohen Wahrscheinlichkeit des Zusammenkommens von Mensch und Spinne sind Bissunfälle dieser Art nicht unwahrscheinlich. Ein Sicherheitsabstand zur Spinne wird deshalb empfohlen.
Ein sich bedroht fühlendes Exemplar der Art nimmt für gewöhnlich eine für einige Spinnen typische Drohgebärde ein (s. Kapitel „Abwehrverhalten“ und Bild). Es gibt allerdings auch Berichte von Bissen, die sich ohne Vorwarnungen seitens der Spinne ereignet haben. Bedingt durch ihre Agilität ist es der Brasilianischen Wanderspinne auch möglich, Fressfeinde oder vermeintliche Fressfeinde im Sprung anzugreifen.[18] Gelegentlich übt die Brasilianische Wanderspinne sogenannte Trockenbisse aus (Bisse, bei denen die Spinne zwar mit den Cheliceren zubeißt, aber kein Gift verabreicht), die dann mit Ausnahme des Schmerzes durch den Biss selbst sowie womöglichen bakteriellen Infektionen keine Komplikationen hervorrufen.[77]
Behandeln lässt sich ein Giftbiss der Brasilianischen Wanderspinne mit dem Antiserum Anti-ctenidico, mit dem sich sowohl der Biss der Brasilianischen Wanderspinne als auch jener anderer Arten der Gattung Phoneutria genauso behandeln lassen wie Bisse von den allerdings wesentlich harmloseren Wolfsspinnen (Lycosidae) der Gattung Lycosa.[78]
Symptome beim Menschen
Ein Biss der Brasilianischen Wanderspinne kann beim Menschen mit verschiedenen Symptomen einhergehen. Als erstes tritt ein großer lokaler Schmerz an der Bissstelle ein. Ebenso können Ödeme (Schwellung von Körpergewebe), Erythem (Hautrötung), Hyperhidrose (übermäßige Schweißproduktion), Parästhesien (starke Empfindungen in den Versorgungsgebieten von Hautnerven) und Muskelfaszikulationen (feine Zuckungen kleiner Muskelfaserbündel) an der Bissstelle auftreten. Tachykardie (Herzrasen), Bluthochdruck, Priapismus (schmerzhafte Erektion), Erbrechen und Sialorrhoe (übermäßiger Speichelfluss) können Anzeichen für akutere durch das Gift hervorgerufene Körperschäden (systemische Effekte) sein. Weitere starke Symptome, die besonders bei Kindern auftreten, sind Erbrechen, Priapismus, Durchfall, Bradykardie (verminderter Herzschlag), arterielle Hypotonie (zu niedriger Blutdruck), Herzrhythmusstörungen, akutes Lungenödem und Schockzustände. Bei ausbleibender Behandlung kann das Gift der Brasilianischen Wanderspinne zum Tod führen.[77]
Prägung des Begriffs „Bananenspinne“
_female_Domestic_Hunting_Spider_(16289066016).jpg.webp)
Die Brasilianische Wanderspinne hat wie keine andere der als „Bananenspinnen“ bekannten Spinnenarten den Begriff geprägt, sodass er zumeist mit dieser Art assoziiert wird. Insbesondere in vielen Medienformaten erscheinen über mehrere Tage andauernde Blickfänge in Form von Titeln, die auf für den Menschen tödliche und in Supermärkten gefundene Spinnen hindeuten. Zwar sind solche Vorkommnisse in der Vergangenheit mehrfach belegt worden, doch handelt es sich zumeist um andere als Bananenspinne bekannte Spinnenarten, etwa welche der Gattung Cupiennius oder der Warmhaus-Riesenkrabbenspinne (Heteropoda venatoria). Bei in Supermärkten gefundenen Spinnenarten wird deren tatsächliche Identifizierung zumeist deutlich weniger bekannt als der Fund der jeweiligen Spinne selbst, wobei es sich in wenigen Fällen tatsächlich um Individuen der Brasilianischen Wanderspinne handelt.[16]
Terraristik
Wie andere Spinnenarten wird die Brasilianische Wanderspinne gelegentlich auch als Haustier im Rahmen der Terraristik gehalten, wobei gewisse Haltungsparameter für eine erfolgreiche Haltung eingehalten werden sollen. Vor dem Kauf einer oder mehrerer Exemplare der Art sollte man sich überdies der von der Spinne ausgehenden Gefahr bewusst sein.[19]
Gefährdung
Über mögliche Bestandsbedrohungen der Brasilianischen Wanderspinne existieren keine Angaben, da ihre Bestände nicht gewertet werden.
Einzelnachweise
- Wolfgang Bücherl, Eleanor E. Buckley: Venomous Animals and Their Venoms: Venomous Invertebrates. Elsevier, 1972, ISBN 978-1-4832-6289-5, S. 237 (englisch).
- Rosana Martins, Rogerio Bertani: The Non-Amazonian Species Of The Brazilian Wandering Spiders Of The Genus Phoneutria Perty, 1833 (Araneae: Ctenidae), With The Description Of A New Species. In: Zootaxa. Band 1562, Nr. 1, 2007, S. 1 (englisch, researchgate.net [abgerufen am 30. August 2020]).
- Claudio Wessloh: Phoneutria. Staatliches Museum für Naturkunde Karlsruhe, abgerufen am 19. September 2020.
- Lawrence E. Reeves, Jennifer L. Gillett-Kaufman: armed Spiders. University of Florida, abgerufen am 19. September 2020.
- Rosana Martins, Rogerio Bertani: The Non-Amazonian Species Of The Brazilian Wandering Spiders Of The Genus Phoneutria Perty, 1833 (Araneae: Ctenidae), With The Description Of A New Species. In: Zootaxa. Band 1562, Nr. 1, 2007, S. 8 (englisch, researchgate.net [abgerufen am 30. August 2020]).
- Miguel Simó, Antonio Domingos Brescovit: Revision and cladistic analysis of the Neotropical spider genus Phoneutria Perty, 1833 (Araneae, Ctenidae), with notes on related Cteninae. In: Bulletin of the British Arachnological Society. Band 12, Nr. 2, Januar 2001, S. 70 (englisch, researchgate.net [abgerufen am 10. September 2020]).
- Rosana Martins, Rogerio Bertani: The Non-Amazonian Species Of The Brazilian Wandering Spiders Of The Genus Phoneutria Perty, 1833 (Araneae: Ctenidae), With The Description Of A New Species. In: Zootaxa. Band 1562, Nr. 1, 2007, S. 73 (englisch, researchgate.net [abgerufen am 30. August 2020]).
- Rosana Martins, Rogerio Bertani: The Non-Amazonian Species Of The Brazilian Wandering Spiders Of The Genus Phoneutria Perty, 1833 (Araneae: Ctenidae), With The Description Of A New Species. In: Zootaxa. Band 1562, Nr. 1, 2007, S. 3 (englisch, researchgate.net [abgerufen am 30. August 2020]).
- Wilhelm Peters: Bericht über die Leistungen der Naturgeschichte der Arachniden während der Jahre 1847 und 1848. In: Archiv für Naturgeschichte. Band 15, Nr. 1, 1849, S. 335 (Textarchiv – Internet Archive).
- Richard S. Vetter, Stefan Hillebrecht: Distinguishing Two Often-Misidentified Genera (Cupiennius, Phoneutria) (Araneae: Ctenidae) of Large Spiders Found in Central and South American Cargo Shipments. In: American Entomologist. Band 40, Nr. 2, 2008, S. 88 (englisch, oup.com [abgerufen am 15. März 2020]).
- Richard S. Vetter, Stefan Hillebrecht: Distinguishing Two Often-Misidentified Genera (Cupiennius, Phoneutria) (Araneae: Ctenidae) of Large Spiders Found in Central and South American Cargo Shipments. In: American Entomologist. Band 40, Nr. 2, 2008, S. 91 (englisch, oup.com [abgerufen am 15. März 2020]).
- Miguel Simó, Antonio Domingos Brescovit: Revision and cladistic analysis of the Neotropical spider genus Phoneutria Perty, 1833 (Araneae, Ctenidae), with notes on related Cteninae. In: Bulletin of the British Arachnological Society. Band 12, Nr. 2, Januar 2001, S. 74 (englisch, researchgate.net [abgerufen am 10. September 2020]).
- Miguel Simó, Antonio Domingos Brescovit: Revision and cladistic analysis of the Neotropical spider genus Phoneutria Perty, 1833 (Araneae, Ctenidae), with notes on related Cteninae. In: Bulletin of the British Arachnological Society. Band 12, Nr. 2, Januar 2001, S. 71 (englisch, researchgate.net [abgerufen am 10. September 2020]).
- Joao Paulo Niemeyer Corbellini, Omar Lupi, Laila Klotz, Livia Montelo: Environmental Causes of Dermatitis. In: Tropical Dermatology. S. 443, doi:10.1016/B978-0-323-29634-2.00036-5.
- Niedersächsisches Landesamt für Verbraucherschutz und Lebensmittelsicherheit: Bananenspinnen – und was dahinter steckt. Niedersächsisches Landesamt für Verbraucherschutz und Lebensmittelsicherheit, abgerufen am 15. März 2020.
- Claudio Wessloh, Hubert Höfer: "BANANENSPINNEN". Arachnologische Gesellschaft e. V., abgerufen am 11. November 2020.
- Rosana Martins, Rogerio Bertani: The Non-Amazonian Species Of The Brazilian Wandering Spiders Of The Genus Phoneutria Perty, 1833 (Araneae: Ctenidae), With The Description Of A New Species. In: Zootaxa. Band 1562, Nr. 1, 2007, S. 4 (englisch, researchgate.net [abgerufen am 30. August 2020]).
- Siegfried Borelli: Dermatologischer Noxen-Katalog: Krankheiten der Haut und Schleimhaut durch Kontakte in Beruf und Umwelt. Springer, 2019, ISBN 978-3-642-70311-9, S. 201.
- Lennart Pettersson: Brazilian wandering spider, (Phoneutria nigriventer, Keyserling 1891) in terrarium. In: Minax Tarantulas. Archiviert vom Original am 6. September 2019; abgerufen am 3. Mai 2019.
- Wolfgang Bücherl: Invertebrados: as aranhas (= Coleção Cientistas de amanha). Edart, 1972, S. 128.
- Steve Peigneur, Maria Elena De Lima, Jan Tytgat: Phoneutria nigriventer venom: A pharmacological treasure. In: Toxicon. Band 151, Nr. 1, Juli 2018, S. 96, doi:10.1016/j.toxicon.2018.07.008 (englisch).
- Gregory H. Bledsoe, Michael J. Manyak, David A. Townes: Expedition and Wilderness Medicine. Cambridge University Press, 2008, ISBN 978-1-139-64385-6 (englisch).
- Wolfgang Bücherl, Eleanor E. Buckley: Venomous Animals and Their Venoms: Venomous Invertebrates. Elsevier, 1972, ISBN 978-1-4832-6289-5, S. 246 (englisch).
- Rosana Martins, Rogerio Bertani: The Non-Amazonian Species Of The Brazilian Wandering Spiders Of The Genus Phoneutria Perty, 1833 (Araneae: Ctenidae), With The Description Of A New Species. In: Zootaxa. Band 1562, Nr. 1, 2007, S. 3 (englisch, researchgate.net [abgerufen am 30. August 2020]).
- Wolfgang Bücherl, Eleanor E. Buckley: Venomous Animals and Their Venoms: Venomous Invertebrates. Elsevier, 1972, ISBN 978-1-4832-6289-5, S. 239 (englisch).
- Nicolas Hazzi: Natural history of Phoneutria boliviensis (Araneae: Ctenidae): habitats, reproductive behavior, postembryonic development and prey-wrapping. In: Journal of Arachnology. Band 42, Nr. 3, November 2014, S. 308, doi:10.1636/Hi13-05.1 (englisch, researchgate.net [abgerufen am 3. Februar 2021]).
- Wolfgang Bücherl: Invertebrados: as aranhas (= Coleção Cientistas de amanha). Edart, 1972, S. 127.
- Nicolas Hazzi: Natural history of Phoneutria boliviensis (Araneae: Ctenidae): habitats, reproductive behavior, postembryonic development and prey-wrapping. In: Journal of Arachnology. Band 42, Nr. 3, November 2014, S. 305, doi:10.1636/Hi13-05.1 (englisch, researchgate.net [abgerufen am 3. Februar 2021]).
- Wolfgang Bücherl, Eleanor E. Buckley: Venomous Animals and Their Venoms: Venomous Invertebrates. Elsevier, 1972, ISBN 978-1-4832-6289-5, S. 242 (englisch).
- Wolfgang Bücherl, Eleanor E. Buckley: Venomous Animals and Their Venoms: Venomous Invertebrates. Elsevier, 1972, ISBN 978-1-4832-6289-5, S. 240 (englisch).
- Wolfgang Bücherl: Invertebrados: as aranhas (= Coleção Cientistas de amanha). Edart, 1972, S. 126.
- Wolfgang Bücherl: Invertebrados: as aranhas (= Coleção Cientistas de amanha). Edart, 1972, S. 126–127.
- Wolfgang Bücherl: Invertebrados: as aranhas (= Coleção Cientistas de amanha). Edart, 1972, S. 127–128.
- Zvonimir Maretic, Drago Lebez: Araneism: With Special Reference to Europe. In: The Quarterly Review of Biology. Band 56, Nr. 1, März 1981, S. 86, doi:10.1086/412130 (englisch).
- Maria Elena de Lima, Suely Gomes Figueiredo, Alessandra Matavel, Kenia Pedrosa Nunes, Carolina Nunes da Silva: Phoneutria nigriventer Venom and Toxins: A Review. In: Spider Venoms. Springer Netherlands, Dordrecht 2015, ISBN 978-94-007-6646-4, S. 5, doi:10.1007/978-94-007-6646-4_6-1 (englisch).
- Maria Elena de Lima, Suely Gomes Figueiredo, Alessandra Matavel, Kenia Pedrosa Nunes, Carolina Nunes da Silva: Phoneutria nigriventer Venom and Toxins: A Review. In: Spider Venoms. Springer Netherlands, Dordrecht 2015, ISBN 978-94-007-6646-4, S. 5–6, doi:10.1007/978-94-007-6646-4_6-1 (englisch).
- Leonides Rezende, Marta N. Cordeiro, Eduardo B. Oliveira, Carlos R. Diniz: Isolation of neurotoxic peptides from the venom of the ‘armed’ spider Phoneutria nigriventer. In: Toxicon. Band 29, Nr. 10, Januar 1991, S. 1225–1233, doi:10.1016/0041-0101(91)90195-W (englisch).
- Maria Elena de Lima, Suely Gomes Figueiredo, Alessandra Matavel, Kenia Pedrosa Nunes, Carolina Nunes da Silva: Phoneutria nigriventer Venom and Toxins: A Review. In: Spider Venoms. Springer Netherlands, Dordrecht 2015, ISBN 978-94-007-6646-4, S. 6–7, doi:10.1007/978-94-007-6646-4_6-1 (englisch).
- Maria Elena de Lima, Suely Gomes Figueiredo, Alessandra Matavel, Kenia Pedrosa Nunes, Carolina Nunes da Silva: Phoneutria nigriventer Venom and Toxins: A Review. In: Spider Venoms. Springer Netherlands, Dordrecht 2015, ISBN 978-94-007-6646-4, S. 7–9, doi:10.1007/978-94-007-6646-4_6-1 (englisch).
- Maria Elena de Lima, Suely Gomes Figueiredo, Alessandra Matavel, Kenia Pedrosa Nunes, Carolina Nunes da Silva: Phoneutria nigriventer Venom and Toxins: A Review. In: Spider Venoms. Springer Netherlands, Dordrecht 2015, ISBN 978-94-007-6646-4, S. 11–14, doi:10.1007/978-94-007-6646-4_6-1 (englisch).
- Paulo Cesar Gomes, Mario Sergio Palma: The Nonpeptide Low Molecular Mass Toxins from Spider Venoms. In: Spider Venoms. Springer Netherlands, Dordrecht 2016, ISBN 978-94-007-6388-3, S. 3–19, doi:10.1007/978-94-007-6389-0_14 (englisch).
- Paulo C. Gomes, Bibiana M. de Souza, Nathalia B. Dias, Lilian M.M. Cesar-Tognoli, Luiz C. Silva-Filho: Nigriventrine: A low molecular mass neuroactive compound from the venom of the spider Phoneutria nigriventer. In: Toxicon. Band 57, Nr. 2, Februar 2011, S. 266–274, doi:10.1016/j.toxicon.2010.11.021 (englisch).
- Wolfgang Bücherl, Eleanor E. Buckley: Venomous Animals and Their Venoms: Venomous Invertebrates. Elsevier, 1972, ISBN 978-1-4832-6289-5, S. 280 (englisch).
- Wolfgang Bücherl, Eleanor E. Buckley: Venomous Animals and Their Venoms: Venomous Invertebrates. Elsevier, 1972, ISBN 978-1-4832-6289-5, S. 281 (englisch).
- Wolfgang Bücherl, Eleanor E. Buckley: Venomous Animals and Their Venoms: Venomous Invertebrates. Elsevier, 1972, ISBN 978-1-4832-6289-5, S. 281–282 (englisch).
- Wolfgang Bücherl, Eleanor E. Buckley: Venomous Animals and Their Venoms: Venomous Invertebrates. Elsevier, 1972, ISBN 978-1-4832-6289-5, S. 282 (englisch).
- Wolfgang Bücherl, Eleanor E. Buckley: Venomous Animals and Their Venoms: Venomous Invertebrates. Elsevier, 1972, ISBN 978-1-4832-6289-5, S. 283 (englisch).
- Wolfgang Bücherl, Eleanor E. Buckley: Venomous Animals and Their Venoms: Venomous Invertebrates. Elsevier, 1972, ISBN 978-1-4832-6289-5, S. 284 (englisch).
- Wolfgang Bücherl, Eleanor E. Buckley: Venomous Animals and Their Venoms: Venomous Invertebrates. Elsevier, 1972, ISBN 978-1-4832-6289-5, S. 284–286 (englisch).
- Wolfgang Bücherl, Eleanor E. Buckley: Venomous Animals and Their Venoms: Venomous Invertebrates. Elsevier, 1972, ISBN 978-1-4832-6289-5, S. 286 (englisch).
- Wolfgang Bücherl, Eleanor E. Buckley: Venomous Animals and Their Venoms: Venomous Invertebrates. Elsevier, 1972, ISBN 978-1-4832-6289-5, S. 287 (englisch).
- Wolfgang Bücherl, Eleanor E. Buckley: Venomous Animals and Their Venoms: Venomous Invertebrates. Elsevier, 1972, ISBN 978-1-4832-6289-5, S. 283–284 (englisch).
- F. Rosa, G. Trevisan, F. K. Rigo, R.l Tonello, E. L. Andrade: Phα1β, a Peptide from the Venom of the Spider Phoneutria Nigriventer Shows Antinociceptive Effects after Continuous Infusion in a Neuropathic Pain Model in Rats. In: Anesthesia & Analgesia. Band 119, Nr. 1, Juli 2014, ISSN 0003-2999, S. 196–202, doi:10.1213/ANE.0000000000000249 (englisch).
- K. P. Nunes, A. Costa-Gonçalves, L. F. Lanza, S.F. Cortes, M. N. Cordeiro: Tx2-6 toxin of the Phoneutria nigriventer spider potentiates rat erectile function. In: Toxicon. Band 51, Nr. 7, Juni 2008, S. 1197–1206, doi:10.1016/j.toxicon.2008.02.010, PMID 18397797, PMC 3019117 (freier Volltext) – (englisch).
- José Edson Fontes Figueiredo, Evanguedes Kalapothakis, Marcus Vinicio Gomez, Wellington Bressan: Heterologous expression of an insecticidal gene from the armed spider (Phoneutria nigriventer). In: Journal of Venomous Animals and Toxins including Tropical Diseases. Band 14, Nr. 2, Januar 2008, S. 274, doi:10.1590/S1678-91992008000200006 (englisch, researchgate.net [abgerufen am 4. März 2020]).
- Luciana Maria Silva, Ana Cristina C Botelho, Rafael Pimenta, Gustavo Ferreira Martins: Structural analysis of the venom glands of the armed spider Phoneutria nigriventer (Keyserling, 1891): Microanatomy, fine structure and confocal observations. In: Toxicon. Band 51, Nr. 4, April 2008, S. 702 (englisch, researchgate.net [abgerufen am 26. März 2021]).
- Luciana Maria Silva, Ana Cristina C Botelho, Rafael Pimenta, Gustavo Ferreira Martins: Structural analysis of the venom glands of the armed spider Phoneutria nigriventer (Keyserling, 1891): Microanatomy, fine structure and confocal observations. In: Toxicon. Band 51, Nr. 4, April 2008, S. 703–704 (englisch, researchgate.net [abgerufen am 26. Dezember 2020]).
- Luciana Maria Silva, Ana Cristina C Botelho, Rafael Pimenta, Gustavo Ferreira Martins: Structural analysis of the venom glands of the armed spider Phoneutria nigriventer (Keyserling, 1891): Microanatomy, fine structure and confocal observations. In: Toxicon. Band 51, Nr. 4, April 2008, S. 704 (englisch, researchgate.net [abgerufen am 7. Dezember 2020]).
- Luciana Maria Silva, Ana Cristina C Botelho, Rafael Pimenta, Gustavo Ferreira Martins: Structural analysis of the venom glands of the armed spider Phoneutria nigriventer (Keyserling, 1891): Microanatomy, fine structure and confocal observations. In: Toxicon. Band 51, Nr. 4, April 2008, S. 704–705 (englisch, researchgate.net [abgerufen am 7. Dezember 2020]).
- Luciana Maria Silva, Ana Cristina C Botelho, Rafael Pimenta, Gustavo Ferreira Martins: Structural analysis of the venom glands of the armed spider Phoneutria nigriventer (Keyserling, 1891): Microanatomy, fine structure and confocal observations. In: Toxicon. Band 51, Nr. 4, April 2008, S. 705 (englisch, researchgate.net [abgerufen am 7. Dezember 2020]).
- Volker Herzig, Richard John Ward, Wagner Ferreira dos Santos: Intersexual variations in the venom of the Brazilian ‘armed’ spider Phoneutria nigriventer (Keyserling, 1891). In: Toxicon. Band 40, Nr. 1, 2002, S. 1403 (englisch).
- Volker Herzig, Richard John Ward, Wagner Ferreira dos Santos: Intersexual variations in the venom of the Brazilian ‘armed’ spider Phoneutria nigriventer (Keyserling, 1891). In: Toxicon. Band 40, Nr. 1, 2002, S. 1403–1404 (englisch).
- Beat A Perret: Venom regeneration in tarantula spiders—I. analysis of venom produced at different time intervals. In: Comparative Biochemistry and Physiology Part A: Physiology. Band 56, Nr. 4, Januar 1977, S. 607–613, doi:10.1016/0300-9629(77)90294-8 (englisch).
- Volker Herzig, Richard John Ward, Wagner Ferreira dos Santos: Intersexual variations in the venom of the Brazilian ‘armed’ spider Phoneutria nigriventer (Keyserling, 1891). In: Toxicon. Band 40, Nr. 1, 2002, S. 1402 (englisch).
- L. Oliveira-Calegário, M. E. De Lima, M. N. Cordeiro, C.R. Diniz: Estudo comparativo da constituição e neurotoxicidade dos venenos de aranhas macho e fêmea da espécie Phoneutria nigriventer. In: Simposio do Sociedade Brasileirade Toxinologia. 2000, S. 11, Abstract (brasilianisches Portugiesisch).
- Volker Herzig, Richard John Ward, Wagner Ferreira dos Santos: Intersexual variations in the venom of the Brazilian ‘armed’ spider Phoneutria nigriventer (Keyserling, 1891). In: Toxicon. Band 40, Nr. 1, 2002, S. 1402–1403 (englisch).
- Volker Herzig, Richard John Ward, Wagner Ferreira dos Santos: Intersexual variations in the venom of the Brazilian ‘armed’ spider Phoneutria nigriventer (Keyserling, 1891). In: Toxicon. Band 40, Nr. 1, 2002, S. 1404 (englisch).
- Marta do Nascimento Cordeiro, Michael Richardson, John Gilroy, Suely Gomes de Figueiredo, Paulo S. L. Beirão: Properties of the Venom from the South American ‘‘Armed'’ Spider Phoneutria Nigriventer (Keyserling, 1891). In: Journal of Toxicology: Toxin Reviews. Band 14, Nr. 3, Januar 1995, ISSN 0731-3837, S. 309–326, doi:10.3109/15569549509019466 (englisch).
- Volker Herzig, Richard John Ward, Wagner Ferreira dos Santos: Intersexual variations in the venom of the Brazilian ‘armed’ spider Phoneutria nigriventer (Keyserling, 1891). In: Toxicon. Band 40, Nr. 1, 2002, S. 1404–1405 (englisch).
- Roberta Amaral Mafra, Suely Gomes Figueiredo, Carlos Ribeiro Diniz, Marta Nascimento Cordeiro, Jáder Dos Santos Cruz: PhTx4, a new class of toxins from Phoneutria nigriventer spider venom, inhibits the glutamate uptake in rat brain synaptosomes. In: Brain Research. Band 831, Nr. 1-2, Juni 1999, S. 297–300, doi:10.1016/S0006-8993(99)01472-9 (englisch).
- Volker Herzig, Richard John Ward, Wagner Ferreira dos Santos: Intersexual variations in the venom of the Brazilian ‘armed’ spider Phoneutria nigriventer (Keyserling, 1891). In: Toxicon. Band 40, Nr. 1, 2002, S. 1405 (englisch).
- S. Schenberg, F. A. Lima: Pharmacology of the polypeptides from the venom of the spider Phoneutria fera. In: Memorias Do Instituto Butantan. Band 33, Nr. 2, 1966, ISSN 0073-9901, S. 627–638, PMID 6012216 (englisch).
- Miguel Simó, Antonio Domingos Brescovit: Revision and cladistic analysis of the Neotropical spider genus Phoneutria Perty, 1833 (Araneae, Ctenidae), with notes on related Cteninae. In: Bulletin of the British Arachnological Society. Band 12, Nr. 2, Januar 2001, S. 73 (englisch, researchgate.net [abgerufen am 10. September 2020]).
- Rosana Martins, Rogerio Bertani: The Non-Amazonian Species Of The Brazilian Wandering Spiders Of The Genus Phoneutria Perty, 1833 (Araneae: Ctenidae), With The Description Of A New Species. In: Zootaxa. Band 1562, Nr. 1, 2007, S. 2 (englisch, researchgate.net [abgerufen am 30. August 2020]).
- Naturhistorisches Museum der Burgergemeinde Bern: World Spider Catalog – Phoneutria nigriventer. Abgerufen am 15. März 2020.
- Miguel Simó, Antonio Domingos Brescovit: Revision and cladistic analysis of the Neotropical spider genus Phoneutria Perty, 1833 (Araneae, Ctenidae), with notes on related Cteninae. In: Bulletin of the British Arachnological Society. Band 12, Nr. 2, Januar 2001, S. 79 (englisch, researchgate.net [abgerufen am 11. Februar 2021]).
- Marcelo Diniz, Ana L. B. Paiva, Clara Guerra-Duarte, Milton Y Nishiyama Jr: An overview of Phoneutria nigriventer spider venom using combined transcriptomic and proteomic approaches. In: PLoS ONE. Band 13, Nr. 8, August 2018, S. 1, doi:10.1371/journal.pone.0200628 (englisch).
- Wolfgang Bücherl, Eleanor E. Buckley: Venomous Animals and Their Venoms: Venomous Invertebrates. Elsevier, 1972, ISBN 978-1-4832-6289-5, S. 273 (englisch).
Literatur
- Wolfgang Bücherl, Eleanor E. Buckley: Venomous Animals and Their Venoms: Venomous Invertebrates. Elsevier, 1972, ISBN 978-1-4832-6289-5 (562 S.).
- Rosana Martins, Rogerio Bertani: The Non-Amazonian Species Of The Brazilian Wandering Spiders Of The Genus Phoneutria Perty, 1833 (Araneae: Ctenidae), With The Description Of A New Species. In: Zootaxa. Band 1562, Nr. 1, 2007, S. 1–36.
- Miguel Simó, Antonio Domingos Brescovit: Revision and cladistic analysis of the Neotropical spider genus Phoneutria Perty, 1833 (Araneae, Ctenidae), with notes on related Cteninae. In: Bulletin of the British Arachnological Society. Band 12, Nr. 2, Januar 2001.
- Richard S. Vetter, Stefan Hillebrecht: Distinguishing Two Often-Misidentified Genera (Cupiennius, Phoneutria) (Araneae: Ctenidae) of Large Spiders Found in Central and South American Cargo Shipments. In: American Entomologist. Band 40, Nr. 2, 2008, S. 88–93.
- Steve Peigneur, Maria Elena De Lima, Jan Tytgat: Phoneutria nigriventer venom: A pharmacological treasure. In: Toxicon. Band 151, Nr. 1, Juli 2018, S. 96–110, doi:10.1016/j.toxicon.2018.07.008.
- Wolfgang Bücherl: Invertebrados: as aranhas (= Coleção Cientistas de amanha). Edart, 1972 (158 S.).
- Maria Elena de Lima, Suely Gomes Figueiredo, Alessandra Matavel, Kenia Pedrosa Nunes, Carolina Nunes da Silva: Phoneutria nigriventer Venom and Toxins: A Review. In: Spider Venoms. Springer Netherlands, Dordrecht 2015, ISBN 978-94-007-6646-4, S. 1–24, doi:10.1007/978-94-007-6646-4_6-1.
- Leonides Rezende, Marta N. Cordeiro, Eduardo B. Oliveira, Carlos R. Diniz: Isolation of neurotoxic peptides from the venom of the ‘armed’ spider Phoneutria nigriventer. In: Toxicon. Band 29, Nr. 10, Januar 1991, S. 1225–1233, doi:10.1016/0041-0101(91)90195-W.
- Paulo C. Gomes, Bibiana M. de Souza, Nathalia B. Dias, Lilian M.M. Cesar-Tognoli, Luiz C. Silva-Filho: Nigriventrine: A low molecular mass neuroactive compound from the venom of the spider Phoneutria nigriventer. In: Toxicon. Band 57, Nr. 2, Februar 2011, S. 266–274, doi:10.1016/j.toxicon.2010.11.021.
- F. Rosa, G. Trevisan, F. K. Rigo, R.l Tonello, E. L. Andrade: Phα1β, a Peptide from the Venom of the Spider Phoneutria Nigriventer Shows Antinociceptive Effects after Continuous Infusion in a Neuropathic Pain Model in Rats:. In: Anesthesia & Analgesia. Band 119, Nr. 1, Juli 2014, ISSN 0003-2999, S. 196–202, doi:10.1213/ANE.0000000000000249.
- K. P. Nunes, A. Costa-Gonçalves, L. F. Lanza, S.F. Cortes, M. N. Cordeiro: Tx2-6 toxin of the Phoneutria nigriventer spider potentiates rat erectile function. In: Toxicon. Band 51, Nr. 7, Juni 2008, S. 1197–1206, doi:10.1016/j.toxicon.2008.02.010, PMID 18397797, PMC 3019117 (freier Volltext).
- Luciana Maria Silva, Ana Cristina C Botelho, Rafael Pimenta, Gustavo Ferreira Martins: Structural analysis of the venom glands of the armed spider Phoneutria nigriventer (Keyserling, 1891): Microanatomy, fine structure and confocal observations. In: Toxicon. Band 51, Nr. 4, April 2008, S. 702.
- José Edson Fontes Figueiredo, Evanguedes Kalapothakis, Marcus Vinicio Gomez, Wellington Bressan: Heterologous expression of an insecticidal gene from the armed spider (Phoneutria nigriventer). In: Journal of Venomous Animals and Toxins including Tropical Diseases. Band 14, Nr. 2, Januar 2008, S. 274–293, doi:10.1590/S1678-91992008000200006.
- Volker Herzig, Richard John Ward, Wagner Ferreira dos Santos: Intersexual variations in the venom of the Brazilian ‘armed’ spider Phoneutria nigriventer (Keyserling, 1891). In: Toxicon. Band 40, Nr. 1, 2002, S. 1399–1406.
- L. Oliveira-Calegário, M. E. De Lima, M. N. Cordeiro, C.R. Diniz: Estudo comparativo da constituição e neurotoxicidade dos venenos de aranhas macho e fêmea da espécie Phoneutria nigriventer. In: Simposio do Sociedade Brasileirade Toxinologia. Abstract, 2000, S. 11.
- Marta do Nascimento Cordeiro, Michael Richardson, John Gilroy, Suely Gomes de Figueiredo, Paulo S. L. Beirão: Properties of the Venom from the South American ‘‘Armed'’ Spider Phoneutria Nigriventer (Keyserling, 1891). In: Journal of Toxicology: Toxin Reviews. Band 14, Nr. 3, Januar 1995, ISSN 0731-3837, S. 309–326, doi:10.3109/15569549509019466.
- Roberta Amaral Mafra, Suely Gomes Figueiredo, Carlos Ribeiro Diniz, Marta Nascimento Cordeiro, Jáder Dos Santos Cruz: PhTx4, a new class of toxins from Phoneutria nigriventer spider venom, inhibits the glutamate uptake in rat brain synaptosomes. In: Brain Research. Band 831, Nr. 1-2, Juni 1999, S. 297–300, doi:10.1016/S0006-8993(99)01472-9.
- Marcelo Diniz, Ana L. B. Paiva, Clara Guerra-Duarte, Milton Y Nishiyama Jr: An overview of Phoneutria nigriventer spider venom using combined transcriptomic and proteomic approaches. In: PLoS ONE. Band 13, Nr. 8, August 2018, S. 1–29, doi:10.1371/journal.pone.0200628.
- Celio J Castro-Junior, Julie Milano, Alessandra Hugo de Souza, Juliana Figueira da Silva: Phα1β toxin prevents capsaicin-induced nociceptive behavior and mechanical hypersensitivity without acting on TRPV1 channels. In: Neuropharmacology. Band 71, doi:10.1016/j.neuropharm.2013.04.001.
- Wolfgang Bücherl: Biology and Venoms of the Most Important South American Spiders of the Genera Phoneutria, Loxosceles, Lycosa, and Latrodectus. In: American Zoologist. Band 9, Nr. 1, Februar 1969, S. 157–159.
Weblinks
- Phoneutria nigriventer im World Spider Catalog
- Phoneutria nigriventer bei Global Biodiversity Information Facility
- Claudio Wessloh: Phoneutria. Staatliches Museum für Naturkunde Karlsruhe, abgerufen am 19. September 2020.
- Lawrence E. Reeves, Jennifer L. Gillett-Kaufman: armed Spiders. University of Florida, abgerufen am 19. September 2020.
- Niedersächsisches Landesamt für Verbraucherschutz und Lebensmittelsicherheit: Bananenspinnen – und was dahinter steckt. Niedersächsisches Landesamt für Verbraucherschutz und Lebensmittelsicherheit, abgerufen am 15. März 2020.
- Claudio Wessloh, Hubert Höfer: "BANANENSPINNEN". Arachnologische Gesellschaft e. V., abgerufen am 11. November 2020.
- Lennart Pettersson: Brazilian wandering spider, (Phoneutria nigriventer, Keyserling 1891) in terrarium. In: Minax Tarantulas. Archiviert vom Original am 6. September 2019; abgerufen am 3. Mai 2019.
- wissenschaft.de: Erektion nach Spinnenbiss. In: Wissenschaft.de. Konradin Medien GmbH, 18. September 2007, abgerufen am 10. März 2021.