L-Gulonolactonoxidase
L-Gulonolactonoxidase (GULO, Gulo oder GLO), auch als L-Gulono-γ-lacton-Oxidase bezeichnet, ist ein Enzym aus der Gruppe der Oxidasen, das für die Herstellung von Ascorbinsäure (Vitamin C) in höheren Organismen sehr wichtig ist. Es katalysiert mit der selektiven Oxidation von L-Gulonolacton (auch L-Gulono-1,4-lacton oder L-Gulono-γ-lacton genannt) den letzten Schritt der Biosynthese von Ascorbinsäure. Die L-Gulonolactonoxidase findet sich bei nahezu allen Wirbeltieren (Vertebrata) und – nach gegenwärtigem Kenntnisstand (2013) – auch bei sehr vielen Wirbellosen (Invertebrata).
| L-Gulonolactonoxidase | ||
|---|---|---|
| Eigenschaften des menschlichen Proteins | ||
| Masse/Länge Primärstruktur | beim Menschen ohne Translation | |
| Bezeichner | ||
| Externe IDs |
| |
| Enzymklassifikation | ||
| EC, Kategorie | 1.1.3.8, Oxidoreduktasen | |
| Reaktionsart | Oxidation | |
| Substrat | L-Gulonolacton + O2 | |
| Produkte | Ascorbinsäure + Wasserstoffperoxid | |
| Vorkommen | ||
| Übergeordnetes Taxon | Wirbeltiere | |
| Ausnahmen | Trockennasenprimaten, Meerschweinchen, Echte Knochenfische, einige Sperlingsvögel- und Fledertier-Familien | |
| Orthologe | ||
| Mensch | Maus | |
| Entrez | 2989 | 268756 |
| Ensembl | ENSG00000234770 | ENSMUST00000059970 |
| UniProt | P58710 | P58710 |
| Refseq (mRNA) | NG_001136.2 | NM_178747.3 |
| Refseq (Protein) | NP_848862.1 | |
| Genlocus | Chr 8: 27.42 – 27.45 Mb | Chr 14: 65.99 – 66.01 Mb |
| PubMed-Suche | 2989 | 268756 |
Die L-Gulonolactonoxidase wird durch Expression eines Gens, des Gulo-Gens, produziert. Ein durch eine Mutation ausgelöster Gendefekt führt bei dem betroffenen Organismus dazu, dass er keine Ascorbinsäure mehr herstellen kann. Ohne ausreichende Vitamin-C-Zufuhr über die Nahrung erkranken solche Organismen an Hypovitaminose C – beim Menschen Skorbut genannt. Beim Menschen sowie zahlreichen anderen Wirbeltiergruppen, u. a. bei allen Echten Knochenfischen (Teleostei), den meisten Taxa der Fledertiere (Chiroptera) und einigen Taxa der Sperlingsvögel (Passeriformes) sowie allen Meerschweinchen (Caviidae) entspricht die genetisch bedingte Unfähigkeit, Ascorbinsäure herstellen zu können, allerdings einem im Laufe der Evolution erworbenen Normalzustand. Vitamin-C-Mangelerscheinungen treten bei ihnen aufgrund einer allgemein Vitamin-C-reichen Nahrung nur in Ausnahmesituationen auf. Während beim Menschen und den übrigen betroffenen Amnioten Gulo als Pseudogen vorliegt und deshalb auch GULOP oder GuloP (P steht für ‚Pseudo‘) genannt wird, ist es bei Echten Knochenfischen gar nicht mehr nachweisbar.
Erst der Funktionsverlust der L-Gulonolactonoxidase macht Ascorbinsäure für die betroffenen Spezies definitionsgemäß zu einem ‚Vitamin‘. Für alle anderen Arten mit funktionsfähiger L-Gulonolactonoxidase ist Ascorbinsäure nur ein Metabolit.[1]
Funktion und Beschreibung
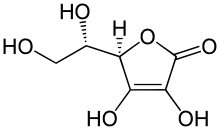
Ascorbinsäure ist für alle Pflanzen[2][3] und Tiere[4] lebensnotwendig (essenziell).[5] Als autotrophen Organismen stehen Pflanzen keine exogenen Quellen zur Deckung des Ascorbinsäurebedarfs zur Verfügung. Sie sind daher alle auf die Eigensynthese von Ascorbinsäure angewiesen. Dagegen können Tiere, die grundsätzlich heterotroph sind, ihren Bedarf an Ascorbinsäure prinzipiell über die Nahrungsaufnahme beispielsweise von Pflanzen decken. Dennoch sind die weitaus meisten Wirbeltiere in der Lage, Ascorbinsäure selbst zu synthetisieren. Bei der sehr großen Anzahl wirbelloser Tiere ist das Wissen darüber, welche Arten in der Lage sind, Ascorbinsäure zu synthetisieren, noch sehr lückenhaft und zum Teil widersprüchlich.[5] Die Biosynthese von Ascorbinsäure in Pflanzen unterscheidet sich grundlegend von der in Tieren.[6] So ist beispielsweise bei höheren Pflanzen im letzten Syntheseschritt L-Galactono-1,4-lacton das Substrat für das Enzym L-Galactono-1,4-γ-lacton-Dehydrogenase (GLDH).[7][8] L-Gulonolactonoxidase spielt bei der Biosynthese von Ascorbinsäure bei Pflanzen keine Rolle.[9]
Bei Tieren beginnt die Biosynthese mit der D-Glucose (Traubenzucker). Sie wird auf enzymatischem Weg in vier Stufen über die Zwischenprodukte D-Glucuronsäure, L-Gulonsäure und L-Gulono-1,4-lacton in Ascorbinsäure umgewandelt.[10] Für den letzten Schritt der Biosynthese in Tieren wird das Enzym L-Gulonolactonoxidase benötigt. Es katalysiert die Oxidation von L-Gulono-1,4-lacton zur Ascorbinsäure. Für diese Reaktion wird zudem Sauerstoff benötigt, der über die Blutgefäße den ascorbinsäureproduzierenden Zellen zugeführt wird. Zusammen mit zwei Wasserstoffatomen, die bei der Reaktion aus dem Ringsystem des L-Gulonolacton in 3,4-Position entfernt werden, bildet sich so als Nebenprodukt der Reaktion Wasserstoffperoxid.
Organismen, denen das Enzym L-Gulonolactonoxidase fehlt oder bei denen es durch eine Mutation nicht funktionsfähig ist, können selbst keine Ascorbinsäure produzieren. Diese Organismen sind auf die Aufnahme ausreichender Mengen von Ascorbinsäure über die Nahrung angewiesen. Andernfalls erkranken sie bei einem Vitamin-C-Mangel.
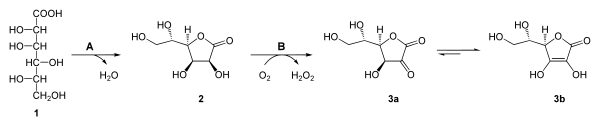
L-Gulonsäure (1) wird unter dem katalytischen Einfluss einer Glucono-Lactonase (A) zu L-Gulonolacton (L-Gulono-1,4-lacton) (2) umgewandelt. Im letzten Schritt katalysiert L-Gulonolactonoxidase (B) die selektive Oxidation des L-Gulonolacton zu 2-Keto-L-Gulonlacton (3a),[11] das spontan zur Ascorbinsäure (3b) tautomerisiert. Als Nebenprodukt der Oxidation entsteht Wasserstoffperoxid (H2O2).[12]
Fehlt einem Organismus das Enzym L-Gulonolactonoxidase, so kann er keine Ascorbinsäure selbst herstellen. Dann ist er zum Überleben auf die exogene Aufnahme von Ascorbinsäure angewiesen. Andernfalls erkrankt er aufgrund des Vitamin-C-Mangels, was nach mehreren Monaten zum Tod führt.
L-Gulonolactonoxidase ist ein mikrosomales Enzym. Bei der Ratte und anderen Gulo-positiven Säugern findet es sich in den Mikrosomen der Hepatozyten (Leberzellen). Es ist ein membranständiges Enzym, dessen aktive Seite in das Lumen der Mikrosomen hineinragt. Das oxidierte Substrat – die Ascorbinsäure – wird dagegen extraluminal in Richtung des Endoplasmatischen Retikulums abgegeben.[13] Das bei der Reaktion ebenfalls entstehende Wasserstoffperoxid wird durch äquivalente Mengen von Glutathion reduziert.[14] Das bevorzugte Substrat der L-Gulonolactonoxidase ist L-Gulono-1,4-lacton. Darüber hinaus ist es auch in der Lage die Oxidation von L-Galactonolacton, D-Mannonolacton und D-Altronolacton zu katalysieren.[15] Dagegen wird die Oxidation von anderen γ-Lactonen, wie beispielsweise L-Idonolacton oder D-Gluconolacton, nicht katalysiert. Offensichtlich müssen die geeigneten Substrate eine Hydroxygruppe am zweiten Kohlenstoffatom aufweisen. Die Michaeliskonstante (Km-Wert) von L-Gulonolactonoxidase liegt im Bereich von 0,007 bis 0,15 mM.[15][16] Prinzipiell ist der Elektronentransfer von der L-Gulonolactonoxidase nicht auf Sauerstoff als Elektronenakzeptor beschränkt. Auch andere Oxidationsmittel, wie beispielsweise Phenazinmethosulfat oder Kaliumhexacyanidoferrat(III) können mittels L-Gulonolactonoxidase L-Gulono-1,4-lacton zu Ascorbinsäure oxidieren.[17][18] Aus der Rattenleber isolierte L-Gulonolactonoxidase besteht aus 440 Aminosäuren und hat eine molare Masse von 50.605 g/mol. Das für dieses Enzym codierende Gen hat einen offenen Leserahmen von 1320 Nukleotiden.[19]
Vorkommen und Nachweis

Das für das Enzym L-Gulonolactonoxidase codierende Gulo-Gen findet sich bei fast allen Wirbeltieren. Es wird vor allem von Zellen in der Leber oder in den Nieren exprimiert. Diese beiden Organe sind die Hauptproduzenten für Ascorbinsäure bei Wirbeltieren. Im Laufe der Evolution fand in verschiedenen Entwicklungslinien der Wirbeltiere unabhängig voneinander ein Wechsel der Ascorbinsäuresynthese von den Nieren zur Leber statt. So wird bei Fischen, Amphibien, Reptilien und entwicklungsgeschichtlich älteren Vogel-Ordnungen sowie bei den eierlegenden Säugetieren (Kloakentiere, Monotremata) Ascorbinsäure in den Nieren produziert. Dagegen findet die Ascorbinsäureproduktion bei entwicklungsgeschichtlich jüngeren Vogel-Ordnungen und bei den höheren Säugetieren (Placentalia) in der Leber statt.[21][1] Beuteltiere (Marsupialia) produzieren Ascorbinsäure sowohl in den Nieren als auch in der Leber.[1] Der Übergang zur größeren Leber ist möglicherweise das Ergebnis eines höheren Selektionsdrucks, um unter Stressbedingungen die Homöostase besser aufrechterhalten zu können.[4][22][23]
Gulo wird von vielen Organismen erst in einer späteren Phase ihrer Individualentwicklung exprimiert. Rattenföten sind beispielsweise erst ab dem 16. Tag in der Lage Ascorbinsäure zu produzieren. Die Expression von Gulo kann durch verschiedene Stimuli erhöht werden. Dazu gehört beispielsweise die Glykogenolyse (der Abbau von Glykogen). Auch verschiedene Medikamente, wie zum Beispiel Barbiturate, Phenazon oder Aminophenazon, sowie Karzinogene, wie Methylcholanthren oder Benzo[a]pyren, erhöhen die Gulo-Expression bei Versuchstieren. Die Ursache hierfür ist vermutlich der erhöhte Bedarf an Glucuronsäure zur Entgiftung dieser Xenobiotika. Dabei werden offensichtlich alle Enzyme des Glucuronsäure-Wegs hochreguliert.[1]
Einige Spezies sind nicht in der Lage, Ascorbinsäure selbst zu synthetisieren. Nach dem gegenwärtigen Stand ist die Ursache hierfür immer ein Defekt des Gulo-Gens oder dessen Deletion.[23]
Bis in die 1970er Jahre hinein bestand die klassische Nachweismethode für ein defektes oder fehlendes Gulo-Gen darin, Versuchstiere möglichst ascorbinsäurefrei zu ernähren und dann auf Symptome des Vitamin-C-Mangels hin zu untersuchen. Danach wurden In-vitro-Techniken entwickelt, bei denen man Gewebehomogenisate, beispielsweise aus Leber oder Nieren der zu untersuchenden Spezies, mit L-Gulono-1,4-lacton, – dem Vorläufermolekül der Ascorbinsäure bei der Biosynthese – versetzte und die unter dem katalytischen Einfluss der L-Gulonolactonoxidase gebildete Menge an Ascorbinsäure bestimmte.[24] Beides sind indirekte Nachweismethoden für das Vorhandensein von L-Gulonolactonoxidase. Moderne Verfahren der Genexpressionsanalyse von Gulo basieren beispielsweise auf Gulo-spezifischen Antikörpern und Western Blot,[25] sowie auf der Fluoreszenz-in-situ-Hybridisierung.[26]
Wirbellose und basale Wirbeltiere

Bei stichprobenartigen Untersuchungen an Wirbellosen („Invertebrata“) und Fischen fand man zunächst keine Hinweise auf eine Aktivität von L-Gulonolactonoxidase oder allgemein auf die Fähigkeit dieser Tiere, Ascorbinsäure synthetisieren zu können. Eine dieser Spezies ist beispielsweise die Wüstenheuschrecke (Schistocerca gregaria).[27] In den 1970er Jahren erwuchs aus diesen Ergebnissen die Ansicht, dass Fische sowie Insekten und andere Wirbellose grundsätzlich nicht in der Lage seien, Ascorbinsäure zu produzieren.[24] Da bekannt war, dass bei den modernen Amphibien L-Gulonolactonoxidase in aktiver Form vorhanden ist, wurde die Hypothese ergänzt durch das Postulat, dass die L-Gulonolactonoxidase ein erst im Zuge des Landgangs der Wirbeltiere, der für den Zeitraum vor ca. 416 bis 359 Millionen Jahren vermutet wird,[28][5] neu erworbenes Merkmal sei. Der Bedarf an Ascorbinsäure, so die Argumentation, sei durch den mit dem Landgang verbundenen erhöhten oxidativen Stress deutlich höher.[29][4]
Bei der Aufstellung dieser Hypothese sind allerdings ältere Untersuchungen, die in direktem Widerspruch dazu standen, nicht berücksichtigt worden. So wurde bereits 1922 beim „Modellorganismus“ Drosophila melanogaster (Schwarzbäuchige Taufliege) festgestellt, dass dieser ohne Ascorbinsäure in der Nahrung auskommt.[30] Gleiches gilt für den Roten Baumwollkapselwurm (Pectinophora gossypiella)[31] und die Motte Argyrotaenia velutinana.[32] Zudem wurden systematische Fehler begangen. So gehörten die untersuchten Fische alle zu den Echten Knochenfischen (Teleostei), bei denen es sich jedoch um eine relativ stark abgeleitete Gruppe der Strahlenflosser (Actinopterygii) handelt. Nachdem in den folgenden Jahren bei ursprünglicheren Strahlenflossern, Lungenfischen (Dipnoi),[33] Haien (Selachii)[34] und Rochen (Batoidea)[35] die Fähigkeit zur Ascorbinsäuresynthese nachgewiesen werden konnte, war die Annahme, dass diese Fähigkeit bei Wirbeltieren erst durch den Landgang evolutionär erworben wurde, nicht mehr haltbar.[36][5] Nachdem 1998 auch bei dem Meerneunauge (Petromyzon marinus), einem sehr ursprünglichen Wirbeltier, aktive L-Gulonolactonoxidase nachgewiesen wurde,[37] kann zudem davon ausgegangen werden, dass die Ascorbinsäuresynthese ein ursprüngliches Merkmal aller Wirbeltiere ist, das lediglich in einigen Entwicklungslinien wieder abhandenkam. Bei Wirbellosen ist das Wissen um die Fähigkeit der Ascorbinsäuresynthese jedoch noch zu lückenhaft, als dass sich derzeit (2013) bestimmen ließe, wann diese durch L-Gulonolactonoxidase ermöglichte Fähigkeit im Laufe der Evolution erstmals in Erscheinung trat.
„Höhere“ Wirbeltiere ohne L-Gulonolactonoxidase
Bei allen Wirbeltieren, die nicht in der Lage sind, Ascorbinsäure selbst zu synthetisieren, ist die Ursache hierfür immer das Gulo-Gen, dessen Genprodukt den letzten Schritt der Biosynthese zur Ascorbinsäure katalysiert. Bei keinem dieser Tiere ist ein Gendefekt in einem der anderen drei in die Ascorbinsäurebiosynthese involvierten Enzyme die Ursache. Die Erklärung hierfür ist, dass ein Defekt bei Gulo nur die Synthese von Ascorbinsäure betrifft,[18] während ein Gendefekt hinsichtlich anderer Enzyme noch die Biosynthese weiterer Substanzen unterbrechen würde. Beispielsweise würde ein Gendefekt, durch den keine Produktion von Glucono-Lactonase erfolgte, nicht nur die Synthese von L-Gulonolacton unterbrechen, sondern unter anderem auch den Pentosephosphatweg und den Abbau von Caprolactam. Das Gulo-Gen unterliegt, im Vergleich zu den anderen Genen der Ascorbinsäurebiosynthese, einem deutlich geringeren Selektionsdruck. Ein Funktionsverlust hat weniger fatale Folgen und ist bei manchen Organismen offensichtlich sogar ohne negative Auswirkungen.[23] Mehrere Entwicklungslinien der Wirbeltiere sind bezüglich L-Gulonolactonoxidase negativ. Dies sind alle Echten Knochenfische (Teleostei), einige Familien der Sperlingsvögel (Passeriformes) und Fledertiere (Chiroptera), alle Arten aus der Familie der Meerschweinchen (Caviidae) und alle zur Unterordnung der Trockennasenprimaten (Haplorhini) gehörenden Arten, einschließlich der des Menschen. Bei den Echten Knochenfischen, Meerschweinchen und Trockennasenprimaten ist der Gendefekt so schwerwiegend, dass er evolutionsgeschichtlich als irreversibel einzustufen ist. Dagegen ist das ursprüngliche Gulo-Pseudogen in einigen Fledertier- und Sperlingsvogelarten im Laufe der Evolution offensichtlich wieder reaktiviert worden. Bei dieser ‚Gen-Reaktivierung‘ spielte nach derzeitigem Kenntnisstand die Nahrung der betroffenen Spezies offensichtlich keine Rolle. Man vermutet daher, dass der Verlust der Fähigkeit, Ascorbinsäure zu synthetisieren, ein neutrales Merkmal ist.[23]
Echte Knochenfische (Teleostei)

Die Landwirbeltiere (Tetrapoda) sind eine prinzipiell Gulo-positive Großgruppe, enthalten jedoch einige Taxa – vor allem unter den Säugetieren – die ebenfalls keine funktionsfähige L-Gulonolactonoxidase besitzen. Diese werden nachfolgend gesondert aufgeführt.


Ursprünglich ging man davon aus, dass Fische generell nicht in der Lage sind, Ascorbinsäure zu synthetisieren, und dass sich diese Fähigkeit im Laufe der Evolution erstmals bei den frühen Landwirbeltieren entwickelte habe.[4][29] Aufgrund umfangreicher Untersuchungen weiß man heute, dass alle Fische, mit Ausnahme der Echten Knochenfische (Teleostei), Ascorbinsäure in ihrem Körper produzieren. Dies tun sie mit Hilfe der L-Gulonolactonoxidase, die bei allen Gulo-positiven Fischen in den Nieren produziert wird.[23] Die Ascorbinsäuresynthese ist ein angestammtes Merkmal von Wirbeltieren, das bei dem gemeinsamen Vorfahren der Teleostier vor etwa 200 bis 210 Millionen Jahren verloren ging.[39][40] Der Gen-Verlust, der dieses Merkmal bewirkt, ist offensichtlich vollständig. Mittels BLAST-Algorithmus konnte in keinem der vollständig sequenzierten Genome eines Teleostiers die Gulo-Sequenz, beziehungsweise Reste davon, gefunden werden.[41] Im Vergleich dazu findet man, ausgehend von der Proteinsequenz der L-Gulonolactonoxidase des Haushuhns (Gallus domesticus), eine 74%ige Übereinstimmung zu der des Weißen Störs (Acipenser transmontanus) und selbst zur Schlauchseescheide (Ciona intestinalis) noch eine 48%ige Übereinstimmung. Das Gulo-Gen, das die L-Gulonolactonoxidase codiert, ist somit über viele Taxa hoch konserviert. Die Ursache dafür, dass man keine Reste des Gulo-Gens im Genom der Teleostier findet, ist entweder, dass das Pseudogen über die etwa 200 Millionen Jahre bis zur Unkenntlichkeit mutierte oder dass es zu einer Gendeletion kam.[23]
Sperlingsvögel (Passeriformes)

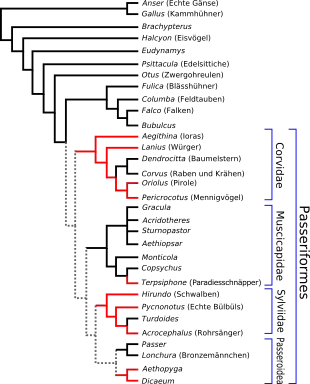
Die Ordnung der Sperlingsvögel (Passeriformes) ist evolutionsgeschichtlich betrachtet ein vergleichsweise junges Taxon.[23] Einige Arten sind nicht in der Lage, Ascorbinsäure selbst zu synthetisieren.[43] Andere wiederum synthetisieren die Ascorbinsäure in der Leber und nicht, wie in vielen anderen Vogelarten, in den Nieren. Der Übergang zur Synthese in der Leber, und der Funktionsverlust bei einigen Arten der Sperlingsvögel, wird von einigen Autoren als „evolutionärer Fortschritt“ gewertet.[23]
Genauere Untersuchungen der Stammesgeschichte machen deutlich, dass diejenigen Sperlingsvögel, die nicht in der Lage sind, Ascorbinsäure zu synthetisieren, nicht monophyletisch sind. Geht man davon aus, dass die Unfähigkeit zur Ascorbinsäuresynthese der angestammte Zustand der Sperlingsvögel ist, so wurde die Fähigkeit viermal in verschiedenen Linien zurückerlangt, und ging einmal erneut verloren (bei Terpsiphone). Nimmt man dagegen an, dass die Fähigkeit zur Ascorbinsäuresynthese der angestammte Zustand ist, so ging diese Fähigkeit dreimal in verschiedenen Linien verloren und wurde dreimal erneut wiedererlangt.[42][23]
Fledertiere (Chiroptera)

Nachdem bei Untersuchungen an der Fledermausart Vesperugo abramus und der Flughunde-Gattung Pteroptus festgestellt wurde, dass diese nicht in der Lage sind Ascorbinsäure zu synthetisieren,[45][46] wurden 1976 insgesamt 34 Fledertierarten aus 6 unterschiedlichen Familien eingehend auf diese Fähigkeit untersucht. Nachdem man keine Gulo-Aktivität bei diesen Tieren fand, schloss man daraus 1976 voreilig, dass dies bei Fledertieren generell der Fall sei.[47] Diese Annahme musste 2011 revidiert werden: Bei der Flughundart Rousettus leschenaultii und der Fledermausart Himalaja-Rundblattnase (Hipposideros armiger) stellte man zunächst überraschenderweise fest, dass bei diesen Tieren das Gulo-Gen kein Pseudogen ist. Mit einem fledertierspezifischen polyklonalen Gulo-Antikörper konnte dann bei diesen beiden Fledertierarten schließlich auch L-Gulonolactonoxidase nachgewiesen werden – sie sind also in der Lage Ascorbinsäure zu produzieren.[25] Verglichen mit einer Maus ist die Produktion von L-Gulonolactonoxidase etwa um den Faktor sechs beziehungsweise vier reduziert. Basierend auf der heute allgemein akzeptierten Phylogenie der Fledertiere lässt sich folgern, dass bei diesen beiden Spezies das ursprünglich inaktive Gulo-Gen evolutionär wieder reaktiviert wurde. Im Gegensatz zu beispielsweise den Teleostiern ist dies möglich, weil die Sequenz des Gulo-Gens in beiden Arten sehr gut erhalten ist und sich von dem Gulo-positiver Säugetiere nur wenig unterscheidet. Die Reaktivierung des Gens benötigte wahrscheinlich nur Mutationen in Bereichen, die an der Regulierung der Expression des Gens beteiligt sind. Die Tatsache, dass die Aktivität deutlich geringer als bei einer Maus ist, lässt darauf schließen, dass weitere Mutationen zur Erhöhung der Expressionsrate erforderlich wären. Andererseits kann die weitere evolutionäre Entwicklung des Gulo-Gens in diesen beiden Spezies auch in genau die andere Richtung verlaufen, nämlich, dass es auf dem Weg zu einem nicht mehr aktiven Pseudogen ist.[23] Beim Kalong-Flughund (Pteropus vampyrus), der Gulo-negativ ist, wurden im Genom die Exons 3 bis 8 sowie 11 und 12 gefunden. Die Sequenz ist frei von Indels und Stopcodons, sodass die Genstruktur noch weitgehend intakt ist. Allerdings weist die dazugehörige Aminosäuresequenz acht Mutationen an Positionen auf, die bei elf anderen Säugetierarten vollständig konserviert sind. Es wird daher vermutet, dass selbst im Fall einer möglichen Expression dieses Gens das Genprodukt – L-Gulonolactonoxidase – nicht funktionsfähig ist. Der Zustand des Gulo-Gens beim Kalong-Flughund ist möglicherweise ein Beispiel für ein Gen, das im Laufe der Evolution nicht mehr reaktiviert werden kann, da zu viele Rückmutationen notwendig wären.[23] Die Veränderungen im Gulo-Gen der Fledertiere sind evolutionsgeschichtlich vergleichsweise jung. Beispielsweise fand die Loss-of-function-Mutation bei der Gattung Pteropus erst vor etwa 3 Millionen Jahren statt.[48]
Meerschweinchen (Caviidae)

Meerschweinchen sind Gulo-negativ, wobei mit diesem Merkmal eine besondere medizinhistorische Episode verbunden ist: Bereits 1907 entdeckten die beiden norwegischen Ärzte Axel Holst und Theodor Frølich,[49] dass Meerschweinchen bei einer bestimmten Diät, die ausschließlich aus Getreide oder Brot bestand,[50] ein Krankheitsbild entwickeln, das dem des Skorbuts beim Menschen entspricht. Ihnen gelang es damit erstmals die Vitamin-C-Mangelerkrankung gezielt auf ein Versuchstier zu übertragen. Darüber hinaus konnten sie zeigen, dass bei einer einseitigen Ernährung mit Weißkohl, Karotte oder Löwenzahn die Versuchstiere nicht erkrankten. Ließen sie den verfütterten Hafer oder die Gerste zuvor keimen, erkrankten die Meerschweinchen ebenfalls nicht. Trockneten sie das gekeimte Getreide vor der Verfütterung oder erwärmten sie es auf 37 °C, so gingen die anti-skorbutischen Eigenschaften wieder verloren.[51] Holst und Frølich gelang mit ihren Versuchen der Beweis, dass Skorbut eine Mangelerkrankung ist.[52] 19 Jahre nach den Versuchen von Holst und Frølich wurde von Albert von Szent-Györgyi Nagyrápolt die Ascorbinsäure entdeckt.
Durch vergleichende Sequenzanalysen des Gulo-Gens von Ratten, Mäusen und Meerschweinchen wurde, ausgehend von einem Zeitpunkt der Trennung (Divergenzzeit[53]) der Meerschweinchen-Linie (entspricht der Großgruppe der Stachelschweinverwandten[54]) von der Ratte-Maus-Linie (entspricht der Großgruppe aus Biberverwandten, Mäuseverwandten und Gleithörnchen[54]) bei etwa 72 mya, der Zeitpunkt der Loss-of-function-Mutation des Gulo-Gens bei Meerschweinchen auf etwa 14 mya datiert.[41] Das vergleichsweise geringe Alter, sowie die Art der weiteren Mutationen im Gulo-Pseudogen, zeigen eindeutig, dass dieser Funktionsverlust unabhängig von dem bei anderen Säugetieren, beispielsweise der Trockennasenprimaten, entstanden sein muss. So sind im Gulo-Pseudogen der Meerschweinchen die Exons 1 und 5 vollständig und Exon 6 teilweise verloren gegangen,[55] während bei den Trockennasenprimaten von den ursprünglichen zwölf Exons sieben verloren gingen.[56] Die Art der ersten Mutation, die zum Funktionsverlust der L-Gulonolactonoxidase geführt hat, ist jedoch sowohl bei den Meerschweinchen als auch bei den Trockennasenprimaten noch völlig unklar.[23]
ODS-Ratten und sfx-Mäuse
ODS-Ratten (Osteogenic Disorder Shionogi) sind ein mutierter Stamm von Albino-Ratten (Wistar-Ratten), bei denen durch eine Punktmutation die Funktion von L-Gulonolactonoxidase völlig zum Erliegen gekommen ist.[57][58] Eine einzige G-A-Mutation (Guanin gegen Adenin) im Nukleotid 182 führt im Genprodukt dazu, dass die Aminosäure Cystein in Position 61 der L-Gulonolactonoxidase durch Tyrosin ersetzt wird, was den vollständigen Funktionsverlust (Loss-of-function-Mutation) der Oxidase zur Folge hat.[55]
Im Jahr 2000 wurde erstmals über einen Mäusestamm berichtet, der zu spontanen Knochenbrüchen neigt.[59] Bei diesen als sfx-Mäuse (engl. spontaneous bone fractures) bezeichneten Tieren wurde zunächst ein Gendefekt auf Chromosom 14 als Ursache gefunden. 2005 wurde entdeckt, dass es sich um eine Deletion des Gulo-Gens auf diesem Chromosom handelt.[60] Erhalten sfx-Mäuse in ihrer Nahrung eine ausreichende Menge an Vitamin C, so geht die Neigung zu spontanen Knochenbrüchen verloren.[61]
ODS-Ratten und sfx-Mäuse werden – neben Meerschweinchen – als Modellorganismen, vor allem für Versuche zum Vitamin-C-Stoffwechsel, verwendet.[62][63][61]
Allgemein


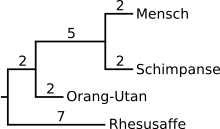
Derzeit (2013) wird davon ausgegangen, dass der Funktionsverlust von Gulo bei den Trockennasenprimaten (Haplorhini) im Zeitraum vor etwa 74 bis 61 Millionen Jahren stattfand, relativ kurz nach Trennung der Linie der Trockennasenprimaten (Altweltaffen, Neuweltaffen und Koboldmakis) von der Lemuren-Linie (77,5 mya).[41]
Da funktionslose Pseudogene keinem Selektionsdruck unterliegen, und Mutationen in diesen Genen für den betroffenen Organismus keinen Evolutionsvorteil oder -nachteil haben, weisen sie typischerweise eine hohe Mutationsrate auf. Deshalb lassen sich durch den Vergleich identischer Genabschnitte Verwandtschaftsbeziehungen zwischen einzelnen Entwicklungslinien der Trockennasenprimaten analysieren. Eine japanische Forschergruppe verglich dazu 1999 einen Genabschnitt mit 164 Basenpaaren auf Exon 10 von GuloP bei mehreren Primatenspezies.[5][64] Je weniger Basenpaare in diesem Abschnitt sich beim Vergleich zweier Arten unterscheiden, desto näher verwandt sind sie miteinander. Bei Schimpansen, den nächsten Verwandten des Menschen, sind tatsächlich auch die Unterschiede zum GuloP-Gen des Menschen am kleinsten.
Für den US-amerikanischen Biologen Jerry Coyne ist GuloP eines der wichtigsten Beweisstücke für die Evolution und ein Argument gegen das sogenannte „Intelligent Design“. Der Verlust der Funktion von Gulo und die Mutationsunterschiede zwischen den Primaten, die mit ihrem Verwandtschaftsgrad korrelieren, lassen sich seiner Meinung nach nur durch die Evolution und gemeinsame Vorfahren dieser Spezies erklären. Coyne fragt unter anderem, weshalb ein „Designer“ beim Menschen einen Mechanismus zur Ascorbinsäuresynthese einbauen würde, ihn aber dann durch die Veränderung eines der dafür verantwortlichen Gene wieder abschaltet.[65]
“Why would a creator put a pathway for making vitamin C in all these species, and then inactivate it?”
Menschen (Homo sapiens)
GuloP ist eines von etwa 80 Pseudogenen, die bisher beim Menschen gefunden und charakterisiert werden konnten.[66][67] Es liegt auf Chromosom 8 Genlocus 21.1.[68] GuloP besteht aus etwa sechs Exons, die aber alle nicht codieren. Das heißt, dass dieses Gen nicht als Vorlage für die Biosynthese eines dem genetischen Code entsprechenden Proteins – dem Enzym L-Gulonolactonoxidase – dient, was wiederum der Grund dafür ist, dass es als Pseudogen bezeichnet wird. Im Vergleich dazu besteht das voll funktionsfähige Gulo-Gen der Ratten aus zwölf Exons.[69] Die Länge des Transkriptes beträgt beim Menschen 748 Basenpaare.[70] Von den zwölf Exons im Gulo-Gen der Ratte finden sich beim Menschen nur die Exons 7, 9, 10 und 12. Für die Exons 8 und 11 liegt wahrscheinlich eine Deletion vor. In den erhaltenen Exons findet sich die für Pseudogene allgemein typische hohe Anzahl an Mutationen.[56][71]
Bis in die 1970er Jahre hinein gab es Spekulationen darüber, dass bestimmte Populationen – speziell die Eskimos – möglicherweise in der Lage sind, Ascorbinsäure in ihrem Körper synthetisieren zu können. Aus der täglichen Nahrung, die seinerzeit fast ausschließlich aus Fisch und Fleisch bestand, schien es, könne der tägliche Bedarf an Vitamin C nicht gedeckt werden.[72] Heute weiß man, dass Eskimos – wie alle anderen Menschen auch – keine L-Gulonolactonoxidase in ihrem Organismus haben und folglich auch keine Ascorbinsäure synthetisieren können. Die Zubereitung von Fleisch, häufig roh, höchstens aber nur mild gekocht, sorgt für einen weitgehenden Erhalt des enthaltenen Vitamin C. Man geht heute davon aus, dass etwa 15 bis 20 mg Vitamin C so über die tägliche Nahrung aufgenommen werden. Eine Menge, die ausreichend hoch ist, um Skorbut zu verhindern. Dazu kommen noch regelrechte Vitamin-C-Schübe durch den Verzehr von roher Robben- oder Rentier-Leber. Der Verzehr von Mengen um 100 Gramm ist ausreichend, um den täglichen Bedarf an Vitamin C zu decken. Von den Eskimos wird Maktaaq (Walhaut) sehr geschätzt und dies lange bevor man durch Analysen einen hohen Gehalt an Vitamin C nachweisen konnte.[73] Maktaaq enthält ca. 35 mg Vitamin C pro 100 Gramm[74][75] – eine höhere Konzentration an Vitamin C als sie einige Zitrusfrüchte aufweisen. Alles in allem geht man davon aus, dass ein Eskimo bei traditioneller Ernährung etwa 40 mg Vitamin C pro Tag aufnimmt.[73]
Ursachen für einen Funktionsverlust
Aus evolutionärer Sicht konnten nur solche Arten die Funktion von L-Gulonolactonoxidase verlieren, die über ihre Nahrung dauerhaft ausreichende Mengen an Ascorbinsäure aufnehmen. Andernfalls wäre eine Loss-of-function-Mutation bei Gulo ein signifikanter Selektionsnachteil. Alle Tierarten, die nicht in der Lage sind, Ascorbinsäure selbst zu produzieren, ernähren sich von Natur aus Vitamin-C-reich. Dies zeigen Untersuchungen an verschiedenen Spezies, die Gulo-negativ sind. Während die empfohlene Vitamin-C-Tagesdosis erwachsener Menschen in den Vereinigten Staaten bei 1 mg pro kg Körpergewicht und Tag liegt, nehmen in freier Wildbahn beispielsweise Gorillas 20 bis 30, Mantelbrüllaffen 88 und Geoffroy-Klammeraffen 106 mg Vitamin C pro kg Körpergewicht und Tag auf. Die Jamaika-Fruchtfledermaus (Artibeus jamaicensis) kommt gar auf einen Wert von 258 mg/kg/Tag.[76] Ein weiteres Indiz für den fehlenden Selektionsdruck bei Gulo-negativen Arten ist, dass diese Tiere zwar eine sehr unterschiedliche, aber immer Vitamin-C-reiche Ernährung haben.[77] Umgekehrt wurde bisher noch keine Gulo-negative Spezies gefunden, die sich Vitamin-C-arm ernährt, beispielsweise durch den ausschließlichen Verzehr von Pflanzensamen.[47] Ein Mehr an Ascorbinsäure durch körpereigene Synthese, zusätzlich zur Ascorbinsäure, die über die Nahrung aufgenommen wird, bietet offenbar keinen Selektionsvorteil. Die Nahrungsergänzung mit Vitamin C zu der normalen, Vitamin-C-reichen Nahrung zeigt bei Meerschweinchen keine positiven Effekte.[4][78][23] Der Selektionsdruck ist bei vielen Wirbeltieren, sowohl was den Verlust, als auch den Wiedergewinn der Gulo-Aktivität betrifft, offensichtlich sehr klein.
Der zweifache Nobelpreisträger Linus Pauling beschäftigte sich intensiv mit der Frage, warum einige Arten die Fähigkeit zur Ascorbinsäure-Synthese verlieren konnten, obwohl diese potenziell doch so lebensnotwendig ist. Er stellte dabei für den Menschen die These auf, dass ein direkter, früher Vorfahre des Menschen vor etwa 25 Millionen Jahren in einem Gebiet lebte, in dem die Nahrung dieser Tierart reich an Ascorbinsäure gewesen sei. Durch eine Mutation sei die Fähigkeit zur körpereigenen Ascorbinsäure-Synthese verlorengegangen. Möglicherweise sei dies durch den Funktionsverlust eines Enzyms geschehen. Da über die Nahrung ausreichend Vitamin C zur Verfügung stand, habe diese Mutation nicht nur keine negativen Auswirkungen, sondern im Gegenteil einen Selektionsvorteil bedeutet. Dieser habe sich dadurch ergeben, dass diese Mutanten keine Ressourcen mehr in Aufbau und Betrieb der Ascorbinsäure-Biosynthese hätten stecken müssen.
“These mutant animals would, in the environment that provided an ample supply of ascorbic acid have an advantage over the ascorbic-acid-producing animals, in that they had been relieved of the burden of constructing and operating the machinery for producing ascorbic acid.”
Die durch den Verlust der Ascorbinsäure-Synthese freigewordene Energie habe den betroffenen Organismen nun für andere Zwecke zu Verfügung gestanden, wodurch sie einen Vorteil gegenüber den Nicht-Mutanten gehabt hätten.[79] Pauling folgte mit diesem Denkansatz weitgehend der Life-history-Theorie[80] und der Less-is-more-Hypothese.[66] Letztere besagt, dass Genverluste eine wichtige Rolle in der Evolution spielen und einen Evolutionsvorteil bedeuten können.[81][82]
Ascorbinsäure regelt in höheren Organismen den Hypoxie-induzierten Faktor 1α (HIF-1α). Bei erhöhten Ascorbinsäurespiegeln wird die Produktion und Aktivität von HIF-1α deutlich reduziert.[83] Im aktivierten Zustand reguliert HIF-1α die Expression von hunderten von Stress-Genen hoch. Aus diesen experimentellen Beobachtungen wurde die Hypothese entwickelt, dass Organismen, die die Fähigkeit zur Ascorbinsäure-Synthese verloren haben, einen evolutionären Vorteil haben, weil sie die HIF-1α-Aktivität über die exogene Aufnahme von Ascorbinsäure regulieren können.[84] Ist die Versorgung mit Ascorbinsäure ausreichend, so ist der Transkriptionsfaktor HIF-1α weniger aktiv, als bei einem Ascorbinsäure-Defizit. Auf diese Weise wird der Organismus offensichtlich in die Lage versetzt, den Versorgungsstatus von Ascorbinsäure zu erkennen.[85] Aus Untersuchungen anderer Pseudogene weiß man, dass diese zwar keine Genprodukte (= Proteine) liefern, aber wichtige epigenetische Funktionen bei der Expression anderer Gene haben.[86] Welche Rolle dabei GuloP spielt, und ob diese einen evolutionären Vorteil bietet, ist bisher noch weitgehend unbekannt.[85]
Eine andere Hypothese geht davon aus, dass der Vorteil einer Ascorbinsäure-Autarkie die Nachteile der Ascorbinsäure-Synthese nicht überwiegt. Bei der durch L-Gulonolactonoxidase katalysierten Oxidation von L-Gulonolacton entsteht als Nebenprodukt Wasserstoffperoxid. Für ein erzeugtes Molekül des Antioxidans Ascorbinsäure entsteht ein Molekül des Oxidationsmittels Wasserstoffperoxid.[87] Dies wiederum erhöht den oxidativen Stress und den Bedarf an Glutathion in den Ascorbinsäure produzierenden Zellen. Glutathion ist – neben Ascorbinsäure – das wichtigste intrazelluläre Antioxidans. Dieser Hypothese folgend, war bei ausreichender Versorgung mit exogener Ascorbinsäure der Verlust der L-Gulonolactonoxidase-Aktivität ein evolutionärer Vorteil.[14] Gegen diese Hypothese spricht allerdings der Fakt, dass bei einigen Spezies das Gulo-Gen zurückmutiert ist. Nach dem gegenwärtigen Stand (2013) wird deshalb eher davon ausgegangen, dass der mehrfache Verlust und die Wiedererlangung der Ascorbinsäure-Synthese zufällig ist, wie es für ein neutrales Merkmal zu erwarten ist. Dieses Merkmal ist allerdings nur so lange neutral, wie ausreichend Vitamin C in der Nahrung enthalten ist.[23]
Der Funktionsverlust der L-Gulonolactonoxidase führt zu einer Einschränkung der Ernährungsweise. Speziell für die Trockennasenaffen wird angenommen, dass es mit dem Funktionsverlust zur Weiterentwicklung der sensorischen Fähigkeiten, zu Verhaltensänderungen und Veränderungen des Stoffwechsels kam, um sich der notwendigen Ernährungsweise besser anzupassen. Möglicherweise hat dies bei den Affen zur Entwicklung des trichromatischen Sehens geführt, das für die Nahrungssuche, unter anderem zur Farbdifferenzierung von Früchten, einen evolutionären Vorteil bietet.[88]
Einzelnachweise
- Gerald F. Combs: The Vitamins. 4. Auflage, Academic Press, 2012, ISBN 0-12-381981-4, S. 236. eingeschränkte Vorschau in der Google-Buchsuche.
- P. L. Conklin: Recent advances in the role and biosynthesis of ascorbic acid in plants. In: Plant, Cell and Environment. Band 24, 2001, S. 383–394, doi:10.1046/j.1365-3040.2001.00686.x.
- R. D. Hancock, D. McRae u. a.: Synthesis of L-ascorbic acid in the phloem. In: BMC plant biology. Band 3, November 2003, S. 7, doi:10.1186/1471-2229-3-7, PMID 14633288, PMC 317296 (freier Volltext).
- I. B. Chatterjee: Evolution and the biosynthesis of ascorbic acid. In: Science. Band 182, Nummer 4118, Dezember 1973, S. 1271–1272, PMID 4752221.
- D. Glaubitz: Ascorbinsäure und Ascorbinsäuresynthese bei Invertebraten – eine vergleichende Analyse. Dissertation, Freien Universität Berlin, 2004, S. 1–20.
- N. Smirnoff, P. L. Conklin, F. A. Loewus: Biosynthesis of ascorbic acid in plants: a renaissance. In: Annual review of plant physiology and plant molecular biology. Band 52, Juni 2001, S. 437–467, doi:10.1146/annurev.arplant.52.1.437, PMID 11337405.
- G. L. Wheeler, M. A. Jones, N. Smirnoff: The biosynthetic pathway of vitamin C in higher plants. In: Nature. Band 393, Nummer 6683, Mai 1998, S. 365–369, doi:10.1038/30728, PMID 9620799.
- C. L. Linster, T. A. Gomez u. a.: Arabidopsis VTC2 encodes a GDP-L-galactose phosphorylase, the last unknown enzyme in the Smirnoff-Wheeler pathway to ascorbic acid in plants. In: The Journal of biological chemistry. Band 282, Nummer 26, Juni 2007, S. 18879–18885, doi:10.1074/jbc.M702094200, PMID 17462988, PMC 2556065 (freier Volltext).
- V. Locato, S. Cimini, L. D. Gara: Strategies to increase vitamin C in plants: from plant defense perspective to food biofortification. In: Frontiers in plant science. Band 4, 2013, S. 152, doi:10.3389/fpls.2013.00152, PMID 23734160, PMC 3660703 (freier Volltext).
- I. B. Chatterjee, G. C. Chatterjee u. a.: Biological synthesis of L-ascorbic acid in animal tissues: conversion of D-glucuronolactone and L-gulonolactone into L-ascorbic acid. In: The Biochemical journal. Band 76, August 1960, S. 279–292, PMID 13692610, PMC 1204705 (freier Volltext).
- John M. C. Gutteridge, Naoyuki Taniguchi: Experimental Protocols for Reactive Oxygen and Nitrogen Species, 2000, Oxford University Press, ISBN 0-19-850668-6.
- N. Smirnoff: L-ascorbic acid biosynthesis. In: Vitamins and hormones. Band 61, 2001, S. 241–266, PMID 11153268 (Review).
- F. Puskás, L. Braun u. a.: Gulonolactone oxidase activity-dependent intravesicular glutathione oxidation in rat liver microsomes. In: FEBS letters. Band 430, Nummer 3, Juli 1998, S. 293–296, PMID 9688558.
- G. Bánhegyi, M. Csala u. a.: Ascorbate synthesis-dependent glutathione consumption in mouse liver. In: FEBS letters. Band 381, Nummer 1–2, Februar 1996, S. 39–41, PMID 8641435.
- K. Kiuchi, M. Nishikimi, K. Yagi: Purification and characterization of L-gulonolactone oxidase from chicken kidney microsomes. In: Biochemistry. Band 21, Nummer 20, September 1982, S. 5076–5082, PMID 7138847.
- M. Nishikimi, B. M. Tolbert, S. Udenfriend: Purification and characterization of L-gulono-gamma-lactone oxidase from rat and goat liver. In: Archives of biochemistry and biophysics. Band 175, Nummer 2, August 1976, S. 427–435, PMID 822334.
- G. L. Eliceiri, E. K. Lai, P. B. McCay: Gulonolactone oxidase. Solubilization, properties, and partial purification. In: J Biol Chem. Band 244, Nummer 10, 1969, S. 2641–2645, PMID 5769996.
- C. L. Linster, E. Van Schaftingen: Vitamin C. Biosynthesis, recycling and degradation in mammals. In: The FEBS journal. Band 274, Nummer 1, Januar 2007, S. 1–22, doi:10.1111/j.1742-4658.2006.05607.x, PMID 17222174 (Review).
- T. Koshizaka, M. Nishikimi u. a.: Isolation and sequence analysis of a complementary DNA encoding rat liver L-gulono-gamma-lactone oxidase, a key enzyme for L-ascorbic acid biosynthesis. In: J Biol Chem. Band 263, Nummer 4, 1988, S. 1619–1621, PMID 3338984.
- S. Z. Wong, B. Ching u. a.: Ascorbic Acid Biosynthesis and Brackish Water Acclimation in the Euryhaline Freshwater White-Rimmed Stingray, Himantura signifer. In: PloS one. Band 8, Nummer 6, 2013, S. e66691, doi:10.1371/journal.pone.0066691, PMID 23825042, PMC 3688944 (freier Volltext).
- S. D. Gupta, C. Sen Gupta u. a.: Enzymic synthesis of L-ascorbic acid from synthetic and biological D-glucurono-1,4-lactone conjugates. In: Analytical biochemistry. Band 38, Nummer 1, November 1970, S. 46–55, PMID 5478251.
- E. C. Birney, R. Jenness, H. D. Hume: Evolution of an enzyme system: ascorbic acid biosynthesis in monotremes and marsupials. In: Evolution. Nummer 34, 1980, S. 230–239.
- G. Drouin, J. R. Godin, B. Pagé: The genetics of vitamin C loss in vertebrates. In: Current genomics. Band 12, Nummer 5, August 2011, S. 371–378, doi:10.2174/138920211796429736, PMID 22294879, PMC 3145266 (freier Volltext).
- I. B. Chatterjee, A. K. Majumder u. a.: Synthesis and some major functions of vitamin C in animals. In: Annals of the New York Academy of Sciences. Band 258, September 1975, S. 24–47, PMID 1106297 (Review).
- J. Cui, Y. H. Pan u. a.: Progressive pseudogenization: vitamin C synthesis and its loss in bats. In: Molecular biology and evolution. Band 28, Nummer 2, Februar 2011, S. 1025–1031, doi:10.1093/molbev/msq286, PMID 21037206.
- L. Hasan, P. Vögeli u. a.: The L-gulono-gamma-lactone oxidase gene (GULO) which is a candidate for vitamin C deficiency in pigs maps to chromosome 14. In: Animal genetics. Band 30, Nummer 4, August 1999, S. 309–312, PMID 10467707.
- R. H. Dadd: Ascorbic Acid and Carotene in the Nutrition of the Desert Locust, Schistocera gregaria Forsk. In: Nature Band 179, 1957, S. 427, doi:10.1038/179427a0.
- G. Niedźwiedzki, P. Szrek u. a.: Tetrapod trackways from the early Middle Devonian period of Poland. In: Nature. Band 463, Nummer 7277, Januar 2010, S. 43–48, doi:10.1038/nature08623, PMID 20054388.
- A. Nandi, C. K. Mukhopadhyay u. a.: Evolutionary significance of vitamin C biosynthesis in terrestrial vertebrates. In: Free radical biology & medicine. Band 22, Nummer 6, 1997, S. 1047–1054, PMID 9034244.
- A. W. Bacot, A. Harden: Vitamin Requirements of Drosophila. I Vitamins B and C. In: The Biochemical journal. Band 16, Nummer 1, 1922, S. 148–152, PMID 16743060, PMC 1259065 (freier Volltext).
- E. S. Vanderzant, R. Reiser: Aseptic rearing of the pink bollworm on synthetic media. In: Journal of Economic Entomology. Band 49, 1956, S. 7–10.
- G. C. Rock: Aseptic rearing of the codling moth on synthetic diets: ascorbic acid and fatty acids requirements. In: Journal of Economic Entomology. Band 60, 1967, S. 1002–1005.
- D. E. Dykhuizen, K. M. Harrison, B. J. Richardson: Evolutionary implications of ascorbic acid production in the Australian lungfish. In: Experientia. Band 36, Nummer 8, August 1980, S. 945–946, PMID 7439328.
- Y. K. Nam, Y. S. Cho u. a.: Isolation and transient expression of a cDNA encoding L-gulono-gamma-lactone oxidase, a key enzyme for L-ascorbic acid biosynthesis, from the tiger shark Scyliorhinus torazame. In: Aquaculture. Band 209, 2002, S. 271–284, doi:10.1016/s0044-8486(01)00731-1.
- D. M. Fracalassi, M. E. Allen u. a.: Ascorbic acid biosynthesis in amazonian fishes. In: Aquaculture. Band 192, 2001, S. 321–332, doi:10.1016/S0044-8486(00)00455-5.
- R. Moreau, K. Dabrowski: Fish acquired ascorbic acid synthesis prior to terrestrial vertebrate emergence. In: Free radical biology & medicine. Band 25, Nummer 8, November 1998, S. 989–990, PMID 9840745.
- R. Moreau, K. Dabrowski: Body pool and synthesis of ascorbic acid in adult sea lamprey (Petromyzon marinus): an agnathan fish with gulonolactone oxidase activity. In: PNAS. Band 95, Nummer 17, August 1998, S. 10279–10282, PMID 9707638, PMC 21499 (freier Volltext).
- Y. S. Cho, S. E. Douglas u. a.: Isolation and characterization of cDNA sequences of L-gulono-gamma-lactone oxidase, a key enzyme for biosynthesis of ascorbic acid, from extant primitive fish groups. In: Comparative biochemistry and physiology. Part B, Biochemistry & molecular biology. Band 147, Nummer 2, Juni 2007, S. 178–190, doi:10.1016/j.cbpb.2007.01.001, PMID 17317254.
- R. Moreau, K. Dabrowski: Biosynthesis of ascorbic acid by extant actinopterygians. In: J Fish Biol. Band 57, 2000, S. 733–745, doi:10.1111/j.1095-8649.2000.tb00271.x.
- K. Dabrowski: Primitive Actinopterigian fishes can synthesize ascorbic acid. In: Experientia. Band 50, Nummer 8, 1994, S. 745–748, doi:10.1007/BF01919376.
- M. Y. Lachapelle, G. Drouin: Inactivation dates of the human and guinea pig vitamin C genes. In: Genetica. Band 139, Nummer 2, Februar 2011, S. 199–207, doi:10.1007/s10709-010-9537-x, PMID 21140195.
- C. Martinez del Rio: Can Passerines Synthesize Vitamin C? In: The Auk. Band 114, Nummer 3, S. 513–516, doi:10.2307/4089257.
- C. R. Chaudhuri, I. B. Chatterjee: L-ascorbic acid synthesis in birds: phylogenetic trend. In: Science. Band 164, Nummer 3878, April 1969, S. 435–436, PMID 5777214.
- E. C. Teeling, M. S. Springer u. a.: A molecular phylogeny for bats illuminates biogeography and the fossil record. In: Science. Band 307, Nummer 5709, Januar 2005, S. 580–584, doi:10.1126/science.1105113, PMID 15681385.
- S. Dutta Gupta, P. K. Choudhury, I. B. Chatterjee: Synthesis of l-ascorbic acid from d-glucurono-1,4-lactone conjugates by different species of animals. In: International Journal of Biochemistry. Band 4, Nummer 21, 1973, S. 309–314, doi:10.1016/0020-711X(73)90053-0.
- R. N. ROY, B. C. GUHA: Species difference in regard to the biosynthesis of ascorbic acid. In: Nature. Band 182, Nummer 4631, August 1958, S. 319–320, PMID 13577829.
- E. C. Birney, R. Jenness, K. M. Ayaz: Inability of bats to synthesise L-ascorbic acid. In: Nature. Band 260, Nummer 5552, April 1976, S. 626–628, PMID 1264230.
- J. Cui, X. Yuan u. a.: Recent loss of vitamin C biosynthesis ability in bats. In: PloS one. Band 6, Nummer 11, 2011, S. e27114, doi:10.1371/journal.pone.0027114, PMID 22069493, PMC 3206078 (freier Volltext).
- A. Holst, T. Frölich: Experimentel studies relating ship beri-beri to scurvy. II. On the etiology of scurvy. In: The Journal of Hygiene. Band 7, 1907, S. 634–671, PMID 4606855.
- Adolphe-Auguste Lesage, Rudolf Fischl: Lehrbuch der Krankheiten des Säuglings. Georg Thieme, 1912, S. 375.
- H. Schaumann: Die Ätiologie der Beriberi II. Kapitel 3: Insuffiziente Nahrungsmittel. Archiv für Schiffs- und Tropenhygiene, 1914, S. 125. eingeschränkte Vorschau in der Google-Buchsuche.
- Tina König: Entwicklung der Ernährungsforschung beim Schwein (bis 1930). (PDF; 3,6 MB) Dissertation, Tierärztliche Hochschule Hannover, 2004, S. 165.
- Wolfgang Schaumann: Charles Darwin – Leben und Werk John Wiley & Sons, 2012, ISBN 3-527-66072-0, S. 51. eingeschränkte Vorschau in der Google-Buchsuche.
- P. H. Fabre, L. Hautier u. a.: A glimpse on the pattern of rodent diversification: a phylogenetic approach. In: BMC Evolutionary Biology. Band 12, 2012, S. 88, doi:10.1186/1471-2148-12-88, PMID 22697210, PMC 3532383 (freier Volltext).
- T. Kawai, M. Nishikimi u. a.: A missense mutation of L-gulono-gamma-lactone oxidase causes the inability of scurvy-prone osteogenic disorder rats to synthesize L-ascorbic acid. In: The Journal of biological chemistry. Band 267, Nummer 30, Oktober 1992, S. 21973–21976, PMID 1400508.
- M. Nishikimi, R. Fukuyama u. a.: Cloning and chromosomal mapping of the human nonfunctional gene for L-gulono-gamma-lactone oxidase, the enzyme for L-ascorbic acid biosynthesis missing in man. In: The Journal of biological chemistry. Band 269, Nummer 18, Mai 1994, S. 13685–13688, PMID 8175804.
- F. Horio, K. Ozaki u. a.: Requirement for ascorbic acid in a rat mutant unable to synthesize ascorbic acid. In: The Journal of nutrition. Band 115, Nummer 12, Dezember 1985, S. 1630–1640, PMID 4067654.
- F. Horio, K. Ozaki u. a.: Ascorbic acid requirement for the induction of microsomal drug-metabolizing enzymes in a rat mutant unable to synthesize ascorbic acid. In: The Journal of nutrition. Band 116, Nummer 11, November 1986, S. 2278–2289, PMID 3098936.
- W. G. Beamer, C. J. Rosen u. a.: Spontaneous fracture (sfx): a mouse genetic model of defective peripubertal bone formation. In: Bone. Band 27, Nummer 5, November 2000, S. 619–626, PMID 11062347.
- Y. Jiao, X. Li u. a.: A deletion causing spontaneous fracture identified from a candidate region of mouse Chromosome 14. In: Mammalian genome. Band 16, Nummer 1, Januar 2005, S. 20–31, doi:10.1007/s00335-004-2414-0, PMID 15674730.
- Y. Jiao, J. Zhang u. a.: Differential gene expression between wild-type and Gulo-deficient mice supplied with vitamin C. In: Genetics and molecular biology. Band 34, Nummer 3, Juli 2011, S. 386–395, doi:10.1590/S1415-47572011005000031, PMID 21931508, PMC 3168176 (freier Volltext).
- C. Vergely, F. Goirand u. a.: Vitamin C deficiency exerts paradoxical cardiovascular effects in osteogenic disorder Shionogi (ODS) rats. In: The Journal of nutrition. Band 134, Nummer 4, April 2004, S. 729–735, PMID 15051818.
- D. Smith, G. Asmundsson u. a.: The osteogenic disorder Shionogi (ODS) rat: a new model for the study of vitamin C metabolism. Bei: United States Department of Agriculture – Agricultural Research Service Juni 1995.
- Y. Ohta, M. Nishikimi: Random nucleotide substitutions in primate nonfunctional gene for L-gulono-gamma-lactone oxidase, the missing enzyme in L-ascorbic acid biosynthesis. In: Biochimica et Biophysica Acta. Band 1472, Nummer 1–2, Oktober 1999, S. 408–411, PMID 10572964.
- Jerry A. Coyne: Why Evolution is True. Oxford University Press, 2009, ISBN 0-19-923084-6, S. 72–73. eingeschränkte Vorschau in der Google-Buchsuche.
- X. Wang, W. E. Grus, J. Zhang: Gene losses during human origins. In: PLoS biology. Band 4, Nummer 3, März 2006, S. e52, doi:10.1371/journal.pbio.0040052, PMID 16464126, PMC 1361800 (freier Volltext).
- Z. D. Zhang, A. Frankish u. a.: Identification and analysis of unitary pseudogenes: historic and contemporary gene losses in humans and other primates. In: Genome biology. Band 11, Nummer 3, 2010, S. R26, doi:10.1186/gb-2010-11-3-r26, PMID 20210993, PMC 2864566 (freier Volltext).
- GULOP.
- Gen-Datenbank des National Center for Biotechnology Information (NCBI): GULOP gulonolactone (L-) oxidase, pseudogene (Homo sapiens (human)) Abgerufen am 1. September 2013.
- Ensembl:Transcript: GULOP-001 ENST00000454030 Abgerufen am 1. September 2013.
- L-Gulonolactonoxidase. In: Online Mendelian Inheritance in Man. (englisch).
- R. A. Gibson, A. J. Sinclair: Are Eskimos obligate carnivores? In: Lancet. Band 1, Nummer 8229, Mai 1981, S. 1100, PMID 6112464.
- Kenneth J. Carpenter: The History of Scurvy and Vitamin C. Cambridge University Press, 1988, ISBN 0-521-34773-4, S. 231. eingeschränkte Vorschau in der Google-Buchsuche.
- J. R. Geraci, T. G. Smith: Vitamin C in the Diet of Inuit Hunters From Holman, Northwest Territories. (PDF; 288 kB) In: Arctic. Band 32, Nummer 2, 1979, S. 135–139.
- K. Fediuk, N. Hidiroglou u. a.: Vitamin C in Inuit traditional food and women’s diets. In: Journal of Food Composition and Analysis. Band 15, Nummer 3, 2002, S. 221–235, doi:10.1006/jfca.2002.1053.
- K. Milton, R. Jenness: Ascorbic acid content of neotropical plant parts available to wild monkeys and bats. In: Experientia. Band 43, Nummer 3, März 1987, S. 339–342, PMID 3104078.
- J. I. Pollock, R. J. Mullin: Vitamin C biosynthesis in prosimians: evidence for the anthropoid affinity of Tarsius. In: American journal of physical anthropology. Band 73, Nummer 1, Mai 1987, S. 65–70, doi:10.1002/ajpa.1330730106, PMID 3113259.
- M. Levine: New concepts in the biology and biochemistry of ascorbic acid. In: The New England Journal of Medicine. Band 314, Nummer 14, April 1986, S. 892–902, doi:10.1056/NEJM198604033141407, PMID 3513016 (Review).
- Tom Hager: Linus Pauling: And the Chemistry of Life. Oxford University Press, 2000, ISBN 0-19-513972-0, S. 122. eingeschränkte Vorschau in der Google-Buchsuche.
- D. A. Roff: Contributions of genomics to life-history theory. In: Nat Rev Genet 8, 2007, S. 116–125, PMID 17230198 (Review).
- NPO: Menschwerdung: Vorteil durch Genverlust? Bei: scinexx vom 14. Januar 2006.
- M. V. Olson: When less is more: gene loss as an engine of evolutionary change. In: American Journal of Human Genetics. Band 64, Nummer 1, Januar 1999, S. 18–23, doi:10.1086/302219, PMID 9915938, PMC 1377697 (freier Volltext) (Review).
- H. J. Knowles, R. R. Raval u. a.: Effect of ascorbate on the activity of hypoxia-inducible factor in cancer cells. In: Cancer Research. Band 63, Nummer 8, April 2003, S. 1764–1768, PMID 12702559.
- A. Grano, M. C. De Tullio: Ascorbic acid as a sensor of oxidative stress and a regulator of gene expression: the Yin and Yang of vitamin C. In: Medical Hypotheses. Band 69, Nummer 4, 2007, S. 953–954, doi:10.1016/j.mehy.2007.02.008, PMID 17376607.
- M. C. De Tullio: The Mystery of Vitamin C. In: Nature Education. Band 3, Nummer 9, 2010, 48.
- L. Poliseno, L. Salmena u. a.: A coding-independent function of gene and pseudogene mRNAs regulates tumour biology. In: Nature. Band 465, Nummer 7301, Juni 2010, S. 1033–1038, doi:10.1038/nature09144, PMID 20577206, PMC 3206313 (freier Volltext).
- B. Halliwell: Vitamin C and genomic stability. In: Mutation research. Band 475, Nummer 1–2, April 2001, S. 29–35, PMID 11295151 (Review).
- Terrence W. Deacon: A Role for Relaxed Selection in the Evolution of the Language Capacity. In: John C. Avise, Francisco J. Ayala (Hrsg.): In the Light of Evolution IV: The Human Condition. Band 4, National Academy of Sciences, National Academies Press, 2010, ISBN 0-309-18528-9, S. 287 eingeschränkte Vorschau in der Google-Buchsuche.