Erdferkel
Das Erdferkel (Orycteropus afer) ist eine Säugetierart und stellt den alleinigen rezenten Vertreter der Gattung Orycteropus in der Familie der Orycteropodidae und der Ordnung der Tubulidentata (Röhrenzähner) dar. Innerhalb der Systematik der heutigen Höheren Säugetiere sind die Röhrenzähner somit die einzige Ordnung, die nur eine Art beinhaltet.
| Erdferkel | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Erdferkel (Orycteropus afer) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name der Ordnung | ||||||||||||
| Tubulidentata | ||||||||||||
| Huxley, 1872 | ||||||||||||
| Wissenschaftlicher Name der Familie | ||||||||||||
| Orycteropodidae | ||||||||||||
| J. E. Gray, 1821 | ||||||||||||
| Wissenschaftlicher Name der Gattung | ||||||||||||
| Orycteropus | ||||||||||||
| É. Geoffroy Saint-Hilaire, 1796 | ||||||||||||
| Wissenschaftlicher Name der Art | ||||||||||||
| Orycteropus afer | ||||||||||||
| (Pallas, 1766) |
Äußerlich markant sind der kräftige Körper des Erdferkels mit aufgewölbtem Rücken und muskulösen Beinen sowie die röhrenförmig verlängerte Schnauze und der fleischige Schwanz. Das Verbreitungsgebiet der Art umfasst das gesamte subsaharische Afrika. Die Tiere bewohnen offene und geschlossene Landschaften. Sie leben einzelgängerisch, sind nachtaktiv und graben im Erdreich, was insgesamt die Beobachtung erschwert. Die Lebensweise des Erdferkels ist daher nur ungenügend untersucht. Seine Nahrung besteht aus staatenbildenden Insekten, was eine hochspezialisierte Anpassung darstellt. Das ausgeprägte Verhalten führt dazu, dass lockere Böden und eine ausreichende Menge an Ameisen und Termiten Grundvoraussetzungen für die Anwesenheit des Erdferkels in einer bestimmten Region sind.
Die Verwandtschaftsverhältnisse des Erdferkels zu anderen Gruppen der Höheren Säugetiere waren lange Zeit ungeklärt. Anfangs wurde die Art aufgrund ihrer spezialisierten Ernährungsweise zusammen mit den ähnlich lebenden Schuppentieren, den Ameisenbären und den Gürteltieren in ein gemeinsames Taxon namens Edentata gestellt. Allerdings beruhen die Ähnlichkeiten der einzelnen Vertreter weitgehend auf konvergenter Evolution, nicht auf Verwandtschaft. Später favorisierten zahlreiche Forscher eine nähere Beziehung zu den Huftieren. Erst molekulargenetische Untersuchungen Ende des 20. Jahrhunderts erbrachten, dass das Erdferkel tatsächlich eine Verwandtschaftsgruppe mit originär afrikanischen Tieren bildet. Die Ergebnisse der Untersuchungen lassen auch eine stammesgeschichtlich frühe Abspaltung des Erdferkels von den anderen Linien dieser Verwandtschaftsgruppe annehmen. Fossilfunde sind allgemein selten und geben momentan nur die letzten 20 Millionen Jahre der Entwicklungsgeschichte wieder. Innerhalb dieser erreichten aber Verwandte des Erdferkels auch das heutige Europa und Asien, wo sie allerdings heute wieder ausgestorben sind. Der einzige rezente Vertreter gilt in seinem Bestand als ungefährdet.
Merkmale
Habitus

Das Erdferkel ist ein mittelgroßer bis großer Vertreter der Säugetiere. Es erreicht allgemein eine Kopf-Rumpf-Länge von 94 bis 142 cm und eine Schulterhöhe von 60 bis 65 cm, der Schwanz misst 44 bis 63 cm. Das Gewicht liegt bei 40 bis 65 kg, da aber der Eintritt der Geschlechtsreife bei Tieren in freier Wildbahn unbekannt ist, sind die Variationen schwer bestimmbar. Ein auffälliger Geschlechtsdimorphismus ist nicht ausgebildet.[1][2] Bei 31 untersuchten Tieren aus Simbabwe konnte eine Gesamtlänge von 149 bis 175 cm und eine Schwanzlänge von 44,3 bis 63,0 cm festgestellt werden. Die entsprechenden Maße bei 10 Tieren aus der Demokratischen Republik Kongo betrugen 161 bis 185 cm beziehungsweise 58 bis 72 cm.[3] Insgesamt stellt das Erdferkel ein kompakt gebautes Tier dar, das einen kräftigen Leib mit markant aufgewölbtem Rücken, einen verhältnismäßig kleinen Kopf und einen langen, fleischigen Schwanz besitzt. Das Rückenfell ist mit rund 1,8 cm Haarlänge relativ kurz, die Bedeckung spärlich und von teils kahlen Flecken durchsetzt. Es zeigt eine grau-braune Grundfärbung, die aber durch lokal vorherrschende Bodenverhältnisse überprägt sein kann. An den Körperseiten und den Beinen wird die Körperbehaarung dichter und länger, die Haare erreichen hier rund 4 cm Länge. Der Nacken des Erdferkels ist kurz und breit. Der Kopf wird durch eine langgestreckte Form charakterisiert, dessen auffälligstes Merkmal die röhrenförmig verlängerte Schnauze ist. Diese endet in eine weiche, flexible Nase und eine kleine Mundöffnung. An den Nasenlöchern, die zumeist schlitzförmig erscheinen, aber weit geöffnet und vollständig geschlossen werden können, sind borstenartige Haare ausgebildet. Die Ohren haben eine langgestreckte, an die von Kaninchen erinnernde Form und werden zwischen 15,8 und 21,0 cm lang. Sie sind zudem extrem beweglich. Die Augen stehen seitlich am Kopf, sind relativ klein und haben eine bräunliche Iris. Sowohl oberhalb als auch unterhalb der Augen und darüber hinaus am Kinn befinden sich zahlreiche borstige Vibrissen. Die Beine sind kurz und äußerst kräftig. Die Vorderfüße besitzen vier, die Hinterfüße fünf Strahlen, die jeweils in massigen Krallen enden, wobei die vorderen größer entwickelt sind als die hinteren. Die einzelnen Zehen werden durch kleine Häutchen verbunden, die dem Graben nützlich sind. Die Länge des Hinterfußes beträgt 22,5 bis 29,0 cm. Weibchen haben zwei Paar Zitzen, die sich in der Brust- und Bauchgegend befinden. Beide Geschlechter verfügen über Drüsen im Genitalbereich.[1][4][2][3]
Schädel- und Gebissmerkmale

(Sammlung Museum Wiesbaden)
Der Schädel des Erdferkels wird zwischen 24,0 und 25,5 cm lang und 9,4 bis 9,8 cm breit. Aufgrund seiner eher langgestreckten Gestalt wirkt er röhrenförmig. Ähnlich den anderen hoch spezialisierten Insektenfressern wie Ameisenbären und Schuppentieren fehlen ihm markante Knochenerhebungen oder -aufrauungen etwa in Form eines Scheitelkamms als Muskelansatzstellen. Im Unterschied zu den beiden anderen Vertretern verfügt das Erdferkel aber über einen geschlossenen, wenn auch schlanken Jochbogen. Der Oberkiefer, Zwischenkieferknochen und das Nasenbein sind gut entwickelt und formen ein verlängertes Rostrum. Das Nasenbein besitzt eine dreieckige Gestaltung mit nach vorn zeigender Spitze und sich verbreiterndem hinteren Teil; der paarige Knochen ist aber nie verwachsen. Das Stirnbein zeigt eine charakteristische Aufwölbung, die durch den außerordentlich entwickelten Riechkolben entsteht; dadurch hat auch der Naseninnenraum ein verhältnismäßig großes Volumen. Das Scheitelbein ist zumeist vollständig verwachsen, es steht im vorderen Bereich mit dem Flügelfortsatz des Keilbeins in Verbindung, sodass das Stirnbein und das Schläfenbein keinen Kontakt zueinander aufweisen. Das Gaumenbein ist langschmal, der seitlich aufsteigende Bereich bildet einen Teil des Randes der Orbita. Im hinteren Teil des Gaumenbeines befinden sich auffällige Öffnungen. Das Mittelohr besitzt einen einfachen Aufbau, eine Paukenblase ist nicht ausgebildet.[5][6][3]
Der insgesamt schlanke Unterkiefer verbreitert sich im Bereich der Backenzähne. Die Symphyse ist bei den heutigen Erdferkeln nur selten verwachsen. An deren hinterem Ende befindet sich das Foramen mentale. Der aufsteigende Ast ragt hoch auf, der Kronenfortsatz (Processus coronoideus) liegt dabei höher als der Gelenkfortsatz (Processus condylaris). Das Unterkiefergelenk ist flach oder leicht eingedellt. Dadurch besitzt auch die Gelenkgrube am Schläfenbein, wo der Unterkiefer einrastet, eine eher schwach entwickelte oder aufgewölbte Gestaltung. Das Gebiss hat bei ausgewachsenen Tieren einen heterodonten und reduzierten Aufbau bestehend aus Prämolaren und Molaren mit allerdings individuell schwankender Zahnanzahl. Die Zahnformel lautet: , die Zahnanzahl variiert somit zwischen 20 und 28. Jungtiere verfügen demgegenüber noch über Schneide- und Eckzähne. Die vorderen Backenzähne sind nagelartig gestaltet, die hinteren zeigen im Querschnitt die Form einer 8 und sind äußerst hochkronig (hypsodont) sowie wurzellos, außerdem fehlt ihnen der harte Zahnschmelz. Der innere Aufbau weicht von den typischen Zähnen der Höheren Säugetiere deutlich ab. So bestehen sie aus säulenartigen Gebilden von vier bis sieben-, häufig aber sechsseitigen Prismen aus Zahnbein, die in einer Matrix aus zahnzementartiger Substanz eingebettet sind. Jede dieser Säulen besitzt eine eigene, rundlich geformte Pulpa. Die Anzahl der Säulchen schwankt individuell und kann von mehreren 100 bis über 1500 reichen.[1] Sie sind teilweise nicht durchgängig ausgebildet, sondern können sich teilen oder vereinen. Die Zähne haben aufgrund dieses Aufbaus keinerlei Krönung.[7][8] Im Milchgebiss weist jeder Backenzahn nur eine Pulpa auf, erst mit dem Wechsel und Wachstum der Zähne entstehen die einzelnen Röhrchen.[9][10][2][3]

Körperskelett

Die Wirbelsäule setzt sich aus 7 Hals-, 13 Brust-, 8 Lenden-, 6 Kreuzbein- und 25 bis 28 Schwanzwirbeln zusammen. Bei nicht ausgewachsenen Tieren und einigen ausgestorbenen Formen kommen nur 5 Kreuzbeinwirbel vor. Der Bewegungsapparat zeigt deutliche Anpassungen an eine grabende Lebensweise. So ist ein Schlüsselbein von kräftiger und stark gebogener Gestalt ausgebildet. An den Langknochen der Arme und Beine kommen zahlreiche erhabene Leisten als Muskelansatzstellen und zusätzlich breite Gelenke vor. Die Schäfte der Langknochen bestehen zum Großteil aus den kompaktierten Knochenbälkchen (Trabekel) des Knocheninnern (Substantia spongiosa), während die äußere Knochenhülle weitgehend resorbiert ist. Dieser besondere, von anderen Großsäugetieren abweichende Knochenaufbau wird mit der starken Anpassung an Grabungsaktivitäten in Verbindung mit teils extremen äußeren Umweltbedingungen und damit begrenztem Nahrungsangebot erklärt.[11] Am Vorderbein ist der Oberarmknochen mit einem äußerst massiven unteren (distalen) Gelenk ausgestattet, dessen Breite über 35 % der Länge des Gesamtknochens erreicht.[12] Am Schaft kommt zudem eine kräftige Knochenleiste, die deltopectorale Leiste, vor. An der Elle ist ein massiver oberer (proximaler) Gelenkfortsatz, das Olecranon, ausgebildet, die Speiche zeigt deutliche Verkürzungen und auffällige Knochenleisten am Schaft. Auch das Becken weist einen wuchtigen Bau auf und ist zudem sowohl Richtung Rücken als auch Schwanz gestreckt. Das Kreuzbein besitzt keine Verbindung zum Sitzbein, die Symphyse des Schambeins ist im Vergleich zu anderen grabenden Säugetieren kaum in ihrer Länge reduziert. Der Oberschenkelknochen ist kurz und robust sowie mit einem kräftigen dritten Rollhügel ausgestattet (Trochanter tertius), der sich am Schaft über eine prominente Rippe erhebt. Am Unterschenkel sind das Schien- und Wadenbein im Bereich des oberen Gelenkendes miteinander verwachsen, der Schaft des Schienbeins ist zudem gebogen. Die Hand- und Fußwurzelknochen zeichnen sich durch eine serielle (taxeopode) Anordnung aus, die auch bei den Rüsseltieren und Schliefern vorherrscht. An der Hand ist das Os centrale mit dem Os scaphoideum (Kahnbein) verwachsen, am Fuß gelenkt das Sprungbein nicht mit dem Würfelbein.[5][3]
Weichteilanatomie
Der Magen des Erdferkels entspricht dem einfachen Bauplan der Säugetiere und ist etwa 15 cm lang bei einem Durchmesser von 7,3 cm. Er besteht aus zwei gleichförmigen Säcken, die durch eine Einschnürung getrennt sind. Als Besonderheit ist aber im hinteren Abschnitt im Bereich des Magenausgangs (Pylorus) eine äußerst kräftige, ringartige Muskulatur von rund 2 cm Dicke ausgebildet. Diese übernimmt das Zerkleinern der Nahrung, das sonst überwiegend über das Zerkauen erfolgt.[13] Auffällig ist auch der für Insektenfresser außergewöhnlich große Blinddarm. Am Gehirn fällt der extrem vergrößerte Riechkolben auf, der zu den größten innerhalb der Höheren Säugetiere gehört und so auch die Dominanz des Geruchssinns anzeigt. Äußerlich wölbt sich durch die Vergrößerung des Riechhirns die Stirn stärker auf.[14] Die Augen stellen Anpassungen an die Dunkelheit dar, Zapfen sind nicht ausgebildet, sodass kein Farbsehen möglich ist. Die Hornhaut weist als Schutz vor Insektenbissen auf der Außenseite eine Keratinschicht auf. Die riemenförmige Zunge ist lang, warzenreich und durch reichliche Speichelabsonderung stets feucht und klebrig. Sie kann 25 bis 30 cm weit aus dem Maul herausgestreckt werden und endet spitz.[14][15][2] Die Nase besteht nur aus zwei Muskeln. Diese sind fleischig und bilden keine Sehnen aus. Dadurch kann die Nase nur eingeschränkt bewegt werden.
Verbreitung und Lebensraum
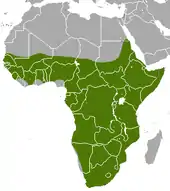
Das Erdferkel ist endemisch in Afrika und dort über den ganzen Kontinent südlich der Sahara verbreitet. Es ist bisher nicht in Lesotho nachgewiesen, kommt dort höchstwahrscheinlich aber ebenfalls vor. Teilweise wird angenommen, dass die Art nicht im Kongobecken auftritt,[1][4] doch liegen von dort zahlreiche Beobachtungen vor, etwa aus dem Ituri forest in der Demokratischen Republik Kongo.[17] Die Verbreitung in Westafrika ist nur wenig untersucht. Aufgrund des weiten Verbreitungsgebietes lebt das Erdferkel in sehr unterschiedlichen Habitaten. Es ist vor allem aus offenen Landschaften wie Savannen, Busch- und Grasländern bis hin zur halbwüstenartigen Karoo nachgewiesen, allerdings ebenso aus Übergangsregionen von offenen zu geschlossenen Landschaften sowie aus dichten Wäldern. Extrem trockene Wüsten und sumpfige Regionen meiden die Tiere aber. Im östlichen Afrika kommt die Art auch in höheren Gebirgslagen vor, etwa in den Udzungwa-Bergen in Tansania[18] oder in den Bale Mountains in Äthiopien, hier konnte sie in bis zu 3200 m Höhe gesichtet werden. Das Erdferkel ist an lockere Böden gebunden und bevorzugt flache Ebenen oder leichte Hänge, die das Anlegen von Erdbauen ermöglichen. In steinigem beziehungsweise felsigem Gelände ist es daher eher selten anzutreffen.[19][20] Grundsätzlich ist das Vorkommen von Ameisen und Termiten eine Voraussetzung für die Anwesenheit des Erdferkels, zugängliche Wasserquellen spielen nur eine untergeordnete Rolle. Die Populationsdichte ist eher niedrig, in für die Tiere zuträglichen Habitaten liegt sie bei rund 10 Individuen auf 10 km².[2][3]
In der geologischen Vergangenheit vom Mittleren Miozän an besiedelten Verwandte des heutigen Erdferkels aus der Gattung Amphiorycteropus, so unter anderem A. depereti und A. gaudryi, auch Eurasien. Hier starben sie aber zu Beginn des Pliozäns und lange vor der Ankunft des Menschen wieder aus.[12] Darstellungen auf Gefäßen und in Grabmalen lassen annehmen, dass das Erdferkel im prädynastischen und im Alten Ägypten auch im mittleren und unteren Niltal anzutreffen war.[21]
Lebensweise
Territorialverhalten
_(1).JPG.webp)
Die Lebensweise des Erdferkels muss trotz seiner weiten Verbreitung als eher wenig erforscht angesehen werden. Die detailliertesten Informationen wurden bisher bei Untersuchungen an Tieren aus dem Naturreservat Tussen die Riviere Nature in der Karoo des südlichen Afrika gewonnen. Das Erdferkel ist nachtaktiv und einzelgängerisch. Die Aktivitätsphase beginnt kurz nach der Abenddämmerung um etwa 19:30 Uhr, wenn die Tiere ihren Bau verlassen, und dauert ununterbrochen bis zur Morgendämmerung gegen 05:30 Uhr an. Am Tage sind sie selten zu sehen. Ausnahmen bilden die kalten Wintermonate im südlichen Afrika, wenn die Temperaturen unter den Gefrierpunkt sinken. Dann treten die Tiere schon in den späten Nachmittagsstunden etwa um 18:00 Uhr in Erscheinung und beenden ihren Tagesrhythmus kurz nach Mitternacht gegen 01:30 Uhr.[20][22] Gründe für diese Verschiebung der Aktivität stellen die geringe Fellbedeckung und eine fehlende Körperfettschicht dar, wodurch die Körperwärme nur mäßig gespeichert wird. Dies ließ sich auch durch eine über rund dreieinhalb Jahre währende Studie in Tswalu in der südafrikanischen Provinz Nordkap zeigen, bei der die untersuchten Tiere unter klimatischen Extrembedingungen ihre Körpertemperatur nicht mehr konstant halten konnten und ihre Aktivitäten stärker in den Tag hinein verlagerten.[23] Den größten Teil des verfügbaren Tagesbudgets verbringt das Erdferkel mit der Suche nach Nahrung, was unabhängig von der vorherrschenden lokalen Witterung erfolgt. Es bewegt sich am Boden im Zehengang fort. Mitunter kann es hohe Geschwindigkeiten von bis zu 40 km/h erreichen. Außerdem vermag das Erdferkel Flüsse und andere Gewässer schwimmend zu durchqueren.[1][4][2][3]



Die einzelnen Tiere nutzen Eigenreviere oder Aktionsräume. In der Karoo sind diese zwischen 1,33 und 3,02 km² groß, ihre Ausdehnung zeigt nur wenige Variationen im Verlauf der Jahreszeiten. Sie können aber bei einem geringeren Nahrungsangebot durchaus größere Flächen einnehmen. Innerhalb der Aktionsräume befindet sich eine größere Anzahl an Erdbauen, die dem Rückzug, dem Schutz und der Aufzucht des Nachwuchses dienen. Die Baue legt das Erdferkel mit Hilfe seiner kräftigen Vorderbeine an, das Erdreich schiebt es mit den Hinterbeinen und teilweise mit dem peitschend über den Boden fegenden Schwanz weg. Die Anlage eines neuen Baues nimmt nur wenig Zeit in Anspruch, ein Tier vermag sich in circa 5 Minuten einzugraben.[15] In vielen Fällen beschränkt sich das Erdferkel darauf, verlassene Baue zu erneuern. Die Dichte an derartigen Unterschlüpfen variiert je nach Landschaftstyp. In der Karoo wurden 101 Baue auf einer Fläche von 1,5 ha gezählt beziehungsweise 58 Baue auf eine Fläche von 200 mal 40 m.[4][2] In den offenen Waldländern im Tal des Mngeni River in der südafrikanischen Provinz KwaZulu-Natal lag die Dichte bei bis zu 29 Bauen auf einem Hektar, in den angrenzenden offenen Grasländern ging sie auf etwa 4 zurück.[19] Dagegen konnten im Ruwenzori-Gebirge in Uganda 15 Baue auf einem Hektar festgestellt werden. Erhebungen im Bui-Nationalpark in Ghana zufolge verteilen sich in den Uferbereichen der Flüsse bis zu 11 Baue auf einem Hektar, während die offenen Savannen nur durchschnittlich 2 Baue auf einer vergleichbar großen Fläche enthalten. Gründe für die unterschiedliche Anzahl von Unterschlüpfen in den verschiedenen Regionen liegen wahrscheinlich in der Beschaffenheit des Untergrundes, wobei weicher Boden eine höhere Anzahl an Erdbauen begünstigt. Sekundär können auch die Bedrohung durch Fressfeinde und durch Buschfeuer eine gewisse Rolle spielen.[24] Die Festigkeit des Bodensubstrats beeinflusst auch die Größe und Tiefe des Unterschlupfes. Die Baue haben einen oder mehrere Eingänge. In Uganda verfügten von 18 untersuchten Bauen 13 über einen Eingang, je zwei über zwei beziehungsweise drei und einer über fünf Eingänge. Bei sieben untersuchten Bauen aus dem Kongobecken besaßen zwei einen Eingang, vier je zwei und einer drei Eingänge. Diese lagen jeweils rund 14 m auseinander.[25][15] Die komplexeren Baue mit mehreren Eingängen bestehen aus bis zu 13 m langen Gängen, bei den ungegabelten sind sie meist zwischen 2 und 3 m lang.[15] Der Eingang reicht steil in den Untergrund bei einem Winkel von rund 40°, später wird der Gang flacher, sein Durchmesser variiert je nach Größe des Tieres zwischen 28 und 45 cm. Er mündet in einer vergrößerten Kammer, die zwischen 70 und 100 cm breit und 74 bis 80 cm hoch ist. Sie kann in bis zu 3 m Tiefe liegen, ihre Ausmaße ermöglichen es einem Tier, sich zu drehen und mit dem Kopf voran den Unterschlupf wieder zu verlassen.[22][4] Die große Tiefe der Wohnkammer hilft auch, die teils empfindlichen Temperaturschwankungen an der Erdoberfläche im Tagesverlauf zu kompensieren. So schwanken die Temperaturen innerhalb des Baus nur um 5 bis 10 °C.[26] Ein einzelnes Tier nutzt einen Unterschlupf nur wenige Tage hintereinander und sucht dann einen neuen auf. In der Karoo wurde eine durchschnittliche Verweildauer von 5 bis 9 Tagen in ein und demselben Bau registriert mit einem Maximum von 38 Tagen hintereinander. Zum Schlafen in der Wohnkammer rollt sich das Erdferkel ein, sodass der Kopf vom Schwanz und den Hinterfüßen bedeckt wird.[1][4][2][3]
Das Sozialsystem ist kaum näher erforscht. Teilweise überschneiden sich die Reviere innerhalb und zwischen den Geschlechtern. Der Grad der Territorialität ist aber unbekannt, Begegnungen zweier gleichgeschlechtlicher Tiere in freier Wildbahn sind bisher äußerst selten beobachtet worden. Bei größerer Populationsdichte kommt es vor, dass Erdferkel beiderlei Geschlechts gemeinsam auftreten. Soziale Interaktionen sind aber auf kurze Treffen von maximal 10 Minuten Dauer beschränkt, in deren Anschluss die Tiere sich wieder der Nahrungssuche widmen. Die Interaktionen werden von Lautäußerungen und dem Beschnüffeln der Genitalgegend begleitet, teilweise erheben sich die Tiere auch auf die Hinterbeine. Das nur gering entwickelte Sozialverhalten ist vermutlich ein Resultat der spezifischen, hochspezialisierten insektenfressenden Lebensweise, da die Nahrungssuche viel Zeit in Anspruch nimmt und so nur wenig Freiraum für ein ausgeprägtes Sozialleben lässt. Die innerartliche Kommunikation findet über das Gehör und den Geruchssinn statt, eine spezielle Körpersprache ist aufgrund des eingeschränkten Sehsinns nicht entwickelt. In der Regel geben einzelne Tiere nur äußerst selten Laute von sich. Zu den bekannten Lautäußerungen gehören ein Grunzen und Blöken. Bedeutend ist das Absetzen von Duftmarken aus Drüsen, die sich bei Weibchen beidseitig der Vulva und beim Männchen hinter der Vorhaut befinden. Diese sondern ein stark riechendes, an Moschus erinnerndes und grünlich gefärbtes Sekret ab, welches über frisch ausgegrabene Erde an Futterplätzen und an Eingängen von Bauen verspritzt oder durch typische Abwärtsbewegungen des Hinterteils abgestrichen wird. Eine spezielle Verhaltensweise stellt auch das Vergraben des Kots dar, der in rund 10 cm tiefe Gruben eingescharrt und teilweise ebenfalls mit Sekreten bedeckt wird.[15] Die Markierungen dienen aber nicht nur der Kommunikation mit Artgenossen, sondern möglicherweise auch der eigenen Orientierung, da die nur wenig gut ausgeprägten Augen, verbunden mit der niedrigen Kopflage und der nächtlichen Aktivität die Bestimmung von Landmarken eher eingeschränkt zulässt.[1][4][2][3][27]
Ernährung
Das Erdferkel ernährt sich vor allem von Insekten. Seine Hauptnahrung besteht aus Ameisen und Termiten, eine Spezialisierung, die als Myrmecophagie bezeichnet wird. Aufgrund der extrem weiten Verbreitung kann die Zusammensetzung der Nahrung lokal sehr unterschiedlich sein. In der Karoo stellen Ameisen die hauptsächliche Grundlage dar und unter diesen wiederum die Vertreter der Gattung Anoplolepis, die durchschnittlich 68,4 % der vertilgten Menge ausmachen. Unter den Termiten gehören die Angehörigen der Gattung Trinervitermes zur bevorzugten Beute; ihr Anteil erreicht im Durchschnitt 15,3 %. Daneben wurden bei Untersuchungen von Kothaufen auch die Ameisengattungen Dorylus, Messor und Crematogaster nachgewiesen, unter den Termiten zudem Hodotermes. Insgesamt umfasst das Beutespektrum in der Karoo mehr als ein Dutzend Arten. Auf die Gesamtmenge bezogen verzehrt das Erdferkel mehr als viermal so viele Ameisen wie Termiten.[28] Im Tal des Mngeni River in KwaZulu-Natal bilden Beobachtungen zufolge Ameisen der Gattung Dorylus etwa 78,2 % der vertilgten Nahrung, unter den Termiten erreicht Odontotermes mit 12,2 % den größten Anteil.[19] Daneben verspeist das Erdferkel gelegentlich auch andere Insekten, etwa Blatthornkäfer oder Grashüpfer, unter Umständen erbeutet es auch kleine Wirbeltiere wie etwa Fettmäuse. Die Nahrungszusammensetzung kann jahreszeitlich schwanken. In der Karoo steigt der Anteil der Termiten im Winter von Mai bis August stark an. In dieser Jahreszeit sind die Termiten weniger aktiv und sammeln sich in ihren Nestern. Bei Trinervitermes kann ein Staat bis zu 80.000 Individuen umfassen, sodass dem Erdferkel durch das Aufbrechen der Baue eine größere Menge an Nahrung zur Verfügung steht. Zudem wachsen in dieser Zeit die Larven heran und bilden so eine energiereiche Nahrungsgrundlage.[28] In Uganda hingegen ist der Anteil an Termiten laut Beobachtungen während der Trockenzeit geringer als während der Regenzeit.[1][4][2][3]
Bei der Nahrungssuche bewegt sich das Erdferkel langsam durch das Gelände, die Durchschnittsgeschwindigkeit beträgt etwa 0,2 bis 0,9 km/h.[20] Dabei läuft es im Zickzack und sucht so einen Streifen von etwa 30 m Breite ab.[15] Die Nase hält das Tier dabei kontinuierlich dicht am Boden und schnüffelt beständig. Sobald ein Nest aufgespürt ist, drückt das Erdferkel die Nase auf den Boden, um die genaue Position der Beute zu erschnüffeln. Bei der gesamten Nahrungssuche hält es die Ohren aufgerichtet, was wahrscheinlich eher dem Aufspüren von Fressfeinden als von Nahrungsressourcen dient. Das Fressen an einem Ameisen- oder Termitennest währt meist nur kurze Zeit, zwischen zehn Sekunden und zwei Minuten. Dadurch kann das Erdferkel rund 25 verschiedene Baue in einer Stunde oder etwa 200 innerhalb einer Nacht aufsuchen. Die gefressene Menge an Insekten beläuft sich auf schätzungsweise 50.000 Individuen. Dazu legt es in der Karoo zwischen 0,5 und 7,0 km zurück.[20] Nach Beobachtungen aus Uganda kann es auf seinen nächtlichen Streifzügen auch Entfernungen von bis zu 14 km überwinden.[15] Die einzelnen Ameisen-/Termitennester werden mit den kräftigen Krallen der Vorderfüße aufgegraben. Die Grabtiefe variiert von 5 bis 30 cm. Für das Anlegen von tiefen Erdlöchern von bis zu 2 m, die etwa bei den Nestern von Hodotermes notwendig sind, erhöht sich dann die Verweildauer des Erdferkels auf bis zu 30 Minuten. In einigen untersuchten Regionen öffnet das Erdferkel die Nester signifikant häufiger an der südlichen und westlichen Seite. Möglicherweise suchen sich die Tiere die wärmsten Bereiche des Termitenbaus, da ihre Beute sich hier in den kühleren Tagesabschnitten der Südhemisphäre konzentriert und dadurch mehr Nahrung zur Verfügung steht.[29][28] Zur Aufnahme der Beute wird die lange Zunge verwendet. Ihre wurmartige Gestaltung ist besonders gut geeignet, um Ameisen und Termiten aus den engen Gängen des Nestes herauszulecken. Dagegen können freilaufende Insekten im offenen Gelände damit nicht so effektiv erbeutet werden. Borstenartige Haare in den Nasenlöchern verhindern das gleichzeitige Eindringen von Erde beim Fressen. Das Erdferkel ist für die chemische Abwehr der Ameisen und Termiten weniger anfällig als andere Insektenfresser. Das gilt unter anderem auch für die stark wirkenden Bisse von Dorylus und Trinervitermes. Die Beute schluckt das Erdferkel in der Regel unzerkaut hinunter.[1][4][2][3]
Neben der tierischen Nahrung gibt es auch Hinweise darauf, dass das Erdferkel pflanzliche Produkte vertilgt. Bekannt sind die Früchte und Samen der Gurkenpflanze Cucumis humifructus, die unterirdisch in bis zu 30 cm Tiefe wachsen und daher ausgegraben werden müssen. Diese sind relativ häufig im Kot des Erdferkels nachgewiesen, sodass die Art zur Verbreitung der Samen der Pflanze beiträgt.[30] Es wird teilweise angenommen, dass die Tiere die Pflanzen aufgrund des hohen Wassergehaltes verspeisen. Da das Erdferkel seinen Flüssigkeitsbedarf aber weitgehend über die Insekten deckt und das Gebiss weniger gut geeignet ist, die harte, wasserabweisende Schale zu zerkleinern, besteht auch die Möglichkeit, dass die Tiere die Samen und Früchte als Beifang beim Verzehr der Ameisen und Termiten schlucken. Darüber hinaus sind einige Pilze von pilzzüchtenden Termiten in Mageninhalten nachgewiesen, die wohl überwiegend in der Trockenzeit verzehrt werden.[1][2][3]
Fortpflanzung

Über die Fortpflanzung des Erdferkels liegen nur wenige Berichte aus Freilandbeobachtungen vor, der überwiegende Teil der Informationen stammt von Tieren in menschlicher Obhut. Geburten in freier Wildbahn wurden in Äthiopien zwischen Mai und Juni, in Uganda im frühen November, in der Demokratischen Republik Kongo im Oktober und November sowie im südlichen Afrika von Mai bis August beobachtet. Die Paarungszeit ist in der Karoo im Oktober und November belegt. Beim Geschlechtsakt setzt sich das Männchen auf das Weibchen, die scharfen Krallen der Vorderfüße verursachen dabei häufig tiefe Kratzer auf dem Rücken des Weibchens. Der Akt selbst besteht aus mehreren Wiederholungen, die durchschnittlich nur 15 Sekunden dauern. Die Tragezeit reicht von 210 bis 266 Tagen, was im Durchschnitt etwa 8 Monaten (243 Tage) entspricht und länger ist als die häufig angenommenen sechs bis sieben Monate.[31] Zumeist kommt pro Wurf ein Junges, selten auch zwei Jungtiere zur Welt. Das Neugeborene ist rund 55 cm lang und wiegt zwischen 1,4 und 1,95 kg. Es besitzt eine nackte, fleischfarbene Haut, gut entwickelte Krallen und geöffnete Augen. Von Geburt an ist es aktiv und aufmerksam. Die Anfangszeit verbringt das Junge in einer Nestkammer in einer Erdhöhle, nach rund 2,5 Wochen verlässt es erstmals den Bau mit seiner Mutter. Nach etwa 12 Wochen bricht der erste Zahn durch, feste Nahrung fängt das Junge mit 14 Wochen an aufzunehmen. Mit sechs Monaten vergräbt das Jungtier erstmals seinen eigenen Kot, davor wird Beobachtungen zufolge dieser vom Muttertier gefressen. Eine Beteiligung des Vatertiers an der Aufzucht findet nicht statt. Der Zeitpunkt der Entwöhnung ist unbekannt, ebenso wie der der vollständigen Unabhängigkeit. Zootiere erreichen mit etwa zwei Jahren die Geschlechtsreife. In Gefangenschaft wachsen Jungtiere relativ schnell und wiegen nach sieben Wochen rund 10 kg, nach sieben Monaten etwa 40 kg. In freier Wildbahn dürfte dieser Prozess aber wesentlich langsamer ablaufen. Die Lebenserwartung freilebender Tiere ist unbekannt, Tiere in menschlicher Gefangenschaft wurden bis zu 30 Jahre alt.[1][2][3]
Fressfeinde, Kommensalen und Sonstiges
Zu den bedeutendsten Fressfeinden gehören Löwen, Leoparden, Geparden, Tüpfelhyänen und Afrikanische Wildhunde. Zuweilen erbeuten Pythons Jungtiere in deren Unterschlüpfen. Bei Sichtung von Beutegreifern flieht das Erdferkel in der Regel in den nächsten Erdbau oder gräbt sich ein, teilweise schlägt es dabei mit den Hinterbeinen aus. Direkt bedrohte Tiere wehren sich häufig auf den Hinterbeinen stehend mit dem Schwanz oder den scharfen Krallen der Vorderfüße oder auf dem Rücken liegend mit allen vieren.[1][3]

Eine Besonderheit findet sich in einem gewissen Kommensalismus mit anderen Tierarten. Hervorzuheben ist dabei der Erdwolf, der sich ebenfalls von Termiten ernährt. Allerdings verfügt er nicht über die körperlichen Voraussetzungen, die harten Baue etwa von Trinervitermes aufzubrechen. Im Winter, wenn sich die Termiten im südlichen Afrika in ihre Nester zurückziehen und somit die Hauptnahrung des Erdwolfes schwindet, folgt dieser häufig dem Erdferkel und profitiert so von den zuvor geöffneten Termitenbauten.[32] Ähnliche Beziehungen bestehen auch zu einigen Vogelarten, etwa zum Termitenschmätzer[33] und zur Grasklapperlerche.[34]
Insgesamt sind Interaktionen des Erdferkels mit anderen Tierarten eher selten. Eine hohe Bedeutung haben jedoch dessen Erdbaue, da diese, wenn verlassen, von zahlreichen anderen Tieren nachgenutzt werden. Allein im südlichen Afrika wurden bei Untersuchungen mehr als zwei Dutzend Arten festgestellt, die so die aufgegebenen Unterschlüpfe des Erdferkels okkupieren. Dazu gehören 21 Vertreter der Säugetiere, 2 der Vögel, 3 der Reptilien und 1 der Amphibien. Zu den größeren Nutzern zählen unter anderem Warzenschwein, Löffelhund, Erdwolf oder Schabrackenschakal. Besonders häufig nutzen auch Kleinsäugetiere wie die Südafrikanische Zwergrennmaus, die Afrikanische Striemengrasmaus, die Kurzschwanz-Hamsterratte oder die Natal-Vielzitzenmaus die Verstecke. Möglicherweise ist das eher ausgeglichene Klima innerhalb der Baue im Vergleich zu den extremeren Außentemperaturen des trockenen südlichen Afrika ein Grund für das Aufsuchen der unterirdischen Gänge durch andere Tierarten.[26] Auch aus anderen Bereichen Afrikas sind zahlreiche Tiere nachgewiesen, die von den verlassenen Bauen profitieren, etwa Stachelschweine, Schuppentiere und Ginsterkatzen beziehungsweise die Schwarzfußkatze. Die Baue werden zur Aufzucht des Nachwuchses oder allgemein als Unterschlupf genutzt.[3]
Parasiten
Zu den bedeutendsten äußeren Parasiten gehören Zecken aus den Gattungen Haemaphysalis und Rhiphicephalus. Daneben kommen Tierläuse wie Haemotopinus und Hybophthirus beziehungsweise Flöhe wie Aucheromyia, Neocordylobia und Pachychoeromyia vor. Unter den inneren Parasiten konnten unter anderem Flagellaten und Amöben festgestellt werden. Zu den ersteren zählen etwa mehrere Arten von Trypanosoma, zu letzteren Entamoeba. Sehr vielgestaltig sind Würmer vertreten, darunter Kratzwürmer, Fadenwürmer und Zungenwürmer. Allein bei den Fadenwürmern sind mehr als ein Dutzend Gattungen belegt, die am Erdferkel parasitieren, unter anderem Angulocirrus, Gendrespirura und Filaria.[1][3]
Systematik
Äußere und Innere Systematik
Innere Systematik der Afrotheria nach Heritage et al. 2021[35]
|
Das Erdferkel bildet als Art unter der wissenschaftlichen Bezeichnung Orycteropus afer das alleinige rezente Mitglied der Gattung Orycteropus, ebenso wie der Familie der Orycteropodidae und der Ordnung der Tubulidentata. Als Ordnungsgruppe stellen die Tubulidentata die einzige innerhalb der Säugetiere dar, die nur eine Art beinhaltet. Die Tubulidentata gehören wiederum in die Überordnung der Afrotheria, welche eine der vier Hauptlinien der Höheren Säugetiere repräsentiert. Die Afrotheria selbst sind eine molekulargenetisch definierte Gruppe, die sich aus überwiegend in Afrika heimischen oder dort ursprünglichen Tieren zusammensetzt. Es können innerhalb der Afrotheria zwei größere Verwandtschaftsgemeinschaften unterschieden werden, die Paenungulata und die Afroinsectiphilia. Die Paenungulata umfassen die heutigen Elefanten, die Schliefer und die Seekühe; sie waren als Gruppe schon länger durch anatomische Übereinstimmungen bekannt. Dagegen konnte die Zusammengehörigkeit der Afroinsectiphilia erst gegen Ende des 20. Jahrhunderts erkannt werden. Sie umschließen die Rüsselspringer, Tenreks, Otterspitzmäuse und Goldmulle. Die drei zuletzt genannten bilden zusammen die Ordnung der Tenrekartigen (Afrosoricida). Zumeist wird auch das Erdferkel als Teil der Afroinsectiphilia angesehen. Die genauen Verwandtschaftsverhältnisse sind aber noch nicht restlos geklärt, da die gegenwärtigen Daten sowohl eine Schwestergruppenbeziehung des Erdferkels mit den Rüsselspringern[36] als auch zu allen anderen Afroinsectiphilia unterstützen (in diesem Fall werden dann die Rüsselspringer und Tenrekartigen unter der Bezeichnung Afroinsectivora zusammengefasst[37])[38][39] ebenso wie zu den Paenungulata.[40] Einige Forscher befürworten daher eine Stellung der Tubulidentata als dritte höhere Gliederungseinheit innerhalb der Afrotheria.[41] Anhand der molekulargenetisch gewonnenen Ergebnisse kann der Ursprung der Afrotheria in die Oberkreide vor 90,4 bis 80,9 Millionen Jahren zurückverfolgt werden. Die Aufspaltung der beiden Hauptgruppen erfolgte etwa 15 Millionen Jahre später. Innerhalb der Afroinsectiphilia stellt das Erdferkel als Ordnungsgruppe möglicherweise eine sehr frühe Abspaltung dar.[39]

Es werden bis zu 18 Unterarten unterschieden, deren Differenzierung zumeist auf einer abweichenden Körperfärbung, Körpergröße und einer variierenden Aufwölbung des Stirnbeins beruht. In einigen Fällen lassen sich Abweichungen im Bau der Ohrregion finden.[6] Die Validität einiger Unterarten ist umstritten und ihre Anzahl wird teilweise als zu hoch angesehen, da beispielsweise nicht davon auszugehen ist, dass allein fünf Unterarten in Tansania auftreten.[42] Ebenso ist aufgrund des nur spärlich verfügbaren Materials die genaue Verbreitung der einzelnen Unterarten nicht bekannt. Folgende Unterarten finden sich in der wissenschaftlichen Literatur:[43][44][2]
- O. a. adametzi Grote, 1921; nordwestliches Kamerun
- O. a. aethiopicus Sundevall, 1843; Sudan, Uganda
- O. a. afer Pallas, 1766; Südafrika (westliche Kapprovinz, Kap der Guten Hoffnung)
- O. a. albicaudus Rothschild, 1907; Angola, westliches Sambia, Namibia, Botswana
- O. a. angolensis Zukowsky & Haltenorth, 1957; westliches Angola
- O. a. erikssoni Lönnberg, 1906; nördliche Demokratische Republik Kongo
- O. a. faradjius Hatt, 1932; nordöstliche Demokratische Republik Kongo, nordwestliches Uganda
- O. a. haussanus Matschie, 1900; Togo
- O. a. kordofanicus Rothschild, 1927; zentraler Sudan
- O. a. lademanni Grote, 1921; zentrales Tansania
- O. a. leptodon Hirst, 1906; Kamerun
- O. a. matschiei Grote, 1921; südöstliches Tansania
- O. a. observandus Grote, 1921; südliches Tansania
- O. a. ruvanensis Grote, 1921; Ruanda, nördliches Tansania
- O. a. senegalensis Lesson, 1840; Senegal
- O. a. somalicus Lydekker, 1908; Somalia
- O. a. wardi Lydekker, 1908; östliche Demokratische Republik Kongo, nordöstliches Sambia
- O. a. wertheri Matschie, 1898; nordöstliches Tansania
Ein bisher wenig untersuchtes Phänomen ist der auffallende Unterschied zwischen Waldbewohnern und Offenlandformen. Demnach besitzen Erdferkel aus dichten Wäldern einen größeren Körperbau mit längeren Schwänzen, darüber hinaus haben sie kleinere Ohren, längere Krallen und kürzere Haare. Zu diesen besonders großen Vertretern gehören beispielsweise die Unterarten O. a. erikssoni[10] und O. a. leptodon,[45] die Gesamtlängen von bis zu 1,98 m aufweisen, während Offenlandformen wie O. a. faradjius nur bis zu 1,77 m Gesamtlänge erreichen.[46][25][2]
Innere Systematik der Tubulidentata nach Lehmann 2009[12]
|
In der rezenten Gattung Orycteropus mit seinem heutigen Vertreter Orycteropus afer sind noch einige ausgestorbene Formen bekannt. Hinzu kommen vier weitere, heute nicht mehr bestehende Gattungen. Die stammesgeschichtlich älteste Form ist Eteketoni aus dem Unteren Miozän. Sie wurde im Jahr 2019 durch Martin Pickford wissenschaftlich eingeführt. Besondere Kennzeichen finden sich in dem flachen Hinterschädel und einem Körperskelett, das bereits dem des heutigen Erdferkels ähnelte und gleichfalls vierstrahlige Hände aufwies. Ebenso war das röhrenzähnige Gebiss schon ausgebildet. Der Bewegungsapparat spricht für einen guten Gräber, die schlanke lange Schnauze ermöglichte es den Tieren wohl in Verbindung mit der geringen Körpergröße, Nahrung aus engen Spalten und Hohlräumen herauszulecken.[41] Zeitlich etwas jünger datiert Myorycteropus, der aber aufgrund der Beschreibung durch D. G. MacInnes im Jahr 1956 wiederum den forschungsgeschichtlich zuerst entdeckten Vertreter repräsentiert. Sein Bewegungsapparat weicht vom heutigen Erdferkel ab und ist durch deutlich größere Hände und Füße und durch einen Oberarmknochen gekennzeichnet, dessen äußerst massives unteres Gelenkende über 46 % der Gesamtlänge des Knochens aufwies. Demgegenüber besaß er aber einen schlankeren Oberschenkelknochen mit einem auf einem markanten Hals sitzenden Gelenkkopf. Die Merkmale sprechen dafür, dass Myorycteropus zwar ein guter Gräber war, aber wohl eine etwas abweichende Grabungsmethodik verwendete. Weitere Unterschiede finden sich im Bau des Unterkiefers mit seinen weniger steil aufsteigenden Gelenkästen. Aufgrund der starken Spezialisierung stellt die Gattung nicht die Stammform der Tubulidentata dar, sondern ist eher als ein Seitenzweig aufzufassen. Im Gegensatz dazu besaß das spätmiozäne Leptorycteropus, dessen wissenschaftliche Beschreibung auf Bryan Patterson aus dem Jahr 1975 zurückgeht, einen generalisierten Körperbau und somit keine guten Grabeigenschaften. Ablesen lässt sich dies unter anderem an der fehlenden deltopectoralen Leiste am Oberarmknochen und den schlankeren Gelenkenden. Zudem ist das Gebiss dieser ausgestorbenen Gattung umfassender und zeigt noch einen großen Eckzahn, der in Verbindung mit der stark nach hinten gezogenen Unterkiefersymphyse wohl noch voll funktionsfähig war. Die Schnauze besaß nicht die auffallende Streckung der heutigen Vertreter, während das anders orientierte Becken einen weniger aufgewölbten Rücken vermuten lässt. Insgesamt erscheint Leptorycteropus weniger deutlich insektenfressend gelebt zu haben und grub wohl auch keine unterirdischen Baue. Trotz des vergleichsweise späten Auftretens ist Leptorycteropus der bisher ursprünglichste Gattungsvertreter der Tubulidentaten. Die dritte anerkannte Gattung ist Amphiorycteropus. Sie wurde 2009 von Thomas Lehmann benannt und beinhaltet weitgehend die eurasischen und einige ältere afrikanische Formen des Mittleren und Oberen Miozäns der Ordnung. Unterschiede zum heutigen Erdferkel finden sich unter anderem im Hand- und Fußskelett, wobei bei Amphiorycteropus die Hand deutlich kleiner als der Fuß ist und der Mittelstrahl der Hand somit kürzer als der des Fußes. Generell sind auch die Gliedmaßenknochen relativ schlank. Ebenso treten einzelne Unterschiede im Schädelbau auf. Insgesamt lassen die Merkmale annehmen, dass Amphiorycteropus nicht in die direkte Vorläuferlinie des heutigen Erdferkels gehört.[47][12]
Teilweise wurde auch die Gattung Plesiorycteropus in die Tubulidentata eingegliedert.[48] Ihre Vertreter lebten bis vor rund 1000 Jahren auf Madagaskar und werden manchmal als „Madagassische Erdferkel“ bezeichnet. Untersuchungen aus dem Jahr 1994 ergaben aber, dass zwischen Plesiorycteropus und den eigentlichen Erdferkeln keine nähere Verwandtschaft besteht, Plesiorycteropus wurde daher in eine eigene Ordnung (Bibymalagasia) gestellt.[49][42][50] Genetische Untersuchungen sprechen dagegen für eine Nahverwandtschaft mit den Tenreks.[40]
Überblick über die fossilen und rezenten Vertreter der Tubulidentata
Die Ordnung der Tubulidentata besteht heute aus fünf Gattungen mit mehr als einem Dutzend Arten:[12][41]
- Ordnung: Tubulidentata Huxley, 1872
- Familie: Orycteropodidae Gray, 1821
- Amphiorycteropus Lehmann, 2009
- A. seni Tekkaya, 1993; Türkei
- A. pottieri Ozansoy, 1965; Türkei, Griechenland
- A. browni Pilgrim, 1933; Pakistan
- A. gaudryi Major, 1888; südliches Europa bis zentrales Asien
- A. mauritanicus Arambourg, 1956; Nordafrika
- A. abundulafus Lehmann, Vignaud, Likius & Brunet, 2005; nördliches Zentralafrika
- A. depereti Helbing, 1933; Südfrankreich
- Eteketoni Pickford, 2019
- E. platycephalus Pickford, 2019; Uganda
- Leptorycteropus Patterson, 1975
- L. guilielmi Patterson 1975; Ostafrika
- Myorycteropus MacInnes, 1956
- M. africanus MacInnes, 1956; Ostafrika
- M. minutus Pickford, 1975; Ostafrika
- M. chemeldoi Pickford, 1975; Ostafrika
- Orycteropus Geoffroy Saint-Hilaire, 1956
- O. djourabensis Lehmann, Vignaud, Mackaye & Brunet, 2004; nördliches Zentral- und Ostafrika
- O. crassidens MacInnes, 1956; Kenia
- O. afer Pallas, 1766; das heute lebende Erdferkel
Einige Formen wie A. seni und A. pottieri haben aufgrund ihres fragmentierten Fundmaterials bisher nur eine unsichere Stellung innerhalb der Gattungen.[12] Die Eigenständigkeit der Gattung Amphiorycteropus wird von manchen Forschern angezweifelt, die sie als synonym zu Leptorycteropus betrachten.[41]
Forschungsgeschichte
Taxonomie und Etymologie

Die wissenschaftliche Erstbeschreibung des Erdferkels erfolgte durch Peter Simon Pallas (1741–1811) im Jahr 1766. Er führte sie anhand eines Fötusses durch, der vom Kap der Guten Hoffnung im südlichen Afrika stammte, das heute auch als Typusregion angesehen wird. Pallas gab dem Erdferkel die wissenschaftlichen Bezeichnung Myrmecophaga afra und stellte es so in die Nähe der Ameisenbären.[51] Später, im Jahr 1780, beschrieb Pallas noch ein ausgewachsenes Tier unter der gleichen Bezeichnung.[52] Johann Friedrich Gmelin stellte dann acht Jahre später in seiner Revision des Systema naturae von Linnaeus die neue Art Myrmecophaga capensis auf. Das Artepitheton capensis galt lange Zeit als gültige Bezeichnung für das Erdferkel. Ob Gmelin dabei die Schriften von Pallas kannte oder durch andere Naturforscher beeinflusst wurde, ist unbekannt.[42]
Der heute gebräuchliche Gattungsname Orycteropus geht auf Étienne Geoffroy Saint-Hilaire (1772–1844) zurück, der ihn im Jahr 1796 in seiner Schrift Extrait d’un mémoire sur le Myrmecophaga capensis erstmals gebrauchte und aufgrund deutlicher Abweichungen zu Myrmecophaga wählte. Der Name ist griechischen Ursprungs und setzt sich aus den Wörtern όρύκτης (oryctes „Grabender“) und πους (pous, „Fuß“) zusammen, bezieht sich also auf die grabende Lebensweise der Tiere.[53] Das Erscheinungsjahr des Aufsatzes wird manchmal auch mit 1795 angegeben, was aber mit Verwechslungen bezüglich des Revolutionskalenders des nachrevolutionären Frankreichs zu begründen ist. Auch die gelegentliche Angabe des Autors mit „C. Geoffroy“[1] ist in diesem Zusammenhang zu sehen, da zu jener Zeit die Bezeichnung Citoyen für „Bürger“ üblich war. Auf Orycteropus basiert des Weiteren die Familienbezeichnung Orycteropodidae. Offiziell gilt heute John Edward Gray (1800–1875) als Erstbenenner, der aber 1821 den Namen Orycteropidae wählte,[54] was erst 1850 durch Charles Lucien Jules Laurent Bonaparte korrigiert wurde. Bryan Patterson nahm im Jahr 1975 eine Aufteilung der Familie in zwei Unterfamilien vor, Orycteropodinae für das eigentliche Erdferkel und Plesiorycteropodinae für die Gattung Plesiorycteropus,[48] letztere bildet heute jedoch keinen Bestandteil der Tubulidentata mehr.[49][40][42][50]
Die Ordnung Tubulidentata für das Erdferkel kreierte Thomas Henry Huxley (1825–1895) im Jahr 1872. Der Name bezieht sich auf die charakteristischen Zähne und besteht aus den lateinischen Wörtern tubus für „Röhre“ und dens für „Zahn“.[55] Der Name setzte sich aber erst in den 1920er Jahren mit den umfangreichen anatomischen Studien von Charles F. Sonntag durch.[5][14][42]
Der deutsche Trivialname leitet sich aus dem Niederländischen beziehungsweise Afrikaans ab, in denen die Tiere Aardvark („Erdschwein“ oder „Erdferkel“) genannt wurden (die heutigen Formen sind „Aardvarken“ im Niederländischen und „Erdvark“ auf Afrikaans[56]). Im englischen Sprachraum wird teilweise auch die Bezeichnung Ant bear gebraucht, was wörtlich „Ameisenbär“ bedeutet.[1][3]
Von den Zahnlosen zu den Röhrenzähnern – Zur systematischen Stellung
Die systematische Stellung des Erdferkels wurde in der forschungsgeschichtlichen Vergangenheit kontrovers diskutiert und ist bis heute nicht restlos geklärt. Die anfängliche Gleichsetzung des Erdferkels mit den südamerikanischen Ameisenbären war 1796 durch Geoffroy Saint-Hilaire mit der Etablierung der Gattung Orycteropus aufgehoben worden. Bereits im Jahr zuvor hatte dieser zusammen mit Georges Cuvier den Begriff der Edentata (Zahnlose) eingeführt, unter denen beide damals die Ameisenbären, die Schuppentiere und die Gürteltiere vereinten[57] und in deren Nähe Geoffroy Saint-Hilaire auch das Erdferkel sah.[53] Im Jahr 1798 gliederte Cuvier neben den Faultieren offiziell auch das Erdferkel in die Edentata ein.[58] Kennzeichnende Merkmale der Edentaten stellten das weitgehende Fehlen der Zähne oder die Ausbildung eines homodonten Gebisses bei gleichzeitigem Verlust des Zahnschmelzes dar. Mit Ausnahme der Faultiere umfassten sie spezialisierte oder generalisierte Insektenfresser. Das Konzept einer systematischen Gruppe namens Edentata hielt sich teilweise bis in das 20. Jahrhundert, auch wenn ihre Zusammensetzung mit der Zeit stark variierte. John Edward Gray beispielsweise führte 1821 das Erdferkel innerhalb der „Oligodontae“ zusammen mit den Megatheriidae und den Gürteltieren. Er setzte die Gruppe von den „Tardigradae“ und den „Edentulae“ ab, in erstere stellte er die heutigen Faultiere, in letztere die Ameisenbären und Schuppentiere.[54] Vier Dekaden darauf wiederum gliederte Gray mit Ausnahme der Faultiere alle diese Formen in die „Edentata Entomophaga“ ein, die seiner Meinung nach zwei Divisionen enthielt: auf der einen Seite die Schuppen- und Gürteltiere, auf der anderen Seite die Ameisenbären und das Erdferkel, zusätzlich aber auch noch das Schnabeltier, welches allerdings nicht zu den Höheren Säugetieren zählt.[59] Thomas Henry Huxley griff diese Ansicht auf und unterteilte die Edentata in die Gruppen der „Phytophaga“ (Blattfresser) und „Entomophaga“ (Insektenfresser).[55] Allerdings kam bereits gegen Ende des 19. Jahrhunderts Kritik an dem Edentata-Konzept auf, da es Tiere zusammenfasse, die originär nicht miteinander verwandt seien. Dies führte unter anderem William Henry Flower im Jahr 1882 aus, indem er auf die altweltliche Verbreitung der Schuppentiere und des Erdferkels gegenüber der neuweltlichen der Gürteltiere, Faultiere und Ameisenbären verwies.[60] In der Folgezeit konnte vor allem Charles F. Sonntag in den 1920er Jahren durch detaillierte anatomische Untersuchungen aufzeigen, dass die verbindenden Merkmale der Edentata eher ursprünglich sind, wogegen die zahlreichen abweichenden Charakteristika sowohl der Skelett- als auch der Weichteilanatomie für eine eigenständige Klassifizierung der Schuppentiere, des Erdferkels und der Nebengelenktiere sprechen. Sonntags Arbeiten führten auch dazu, dass die Tubulidentata als Ordnungsgruppe, bereits 1872 von Huxley vorgeschlagen, Anerkennung fanden.[5][14][15][42]
In der Folgezeit galten die Tubulidentata als zumeist eigenständige Gruppe, deren Verwandtschaftsverhältnis nur ungenügend bekannt war. Sonntag favorisierte einen Ursprung aus dem Kreis der „Condylarthra“, einer umfangreichen, aber nach heutiger Ansicht nicht geschlossenen Gruppe urtümlicher Huftiere, wobei Übereinstimmungen vor allem im Bau der Wirbelsäule und des Fußskelettes bestanden. Er erkannte darüber hinaus zahlreiche Ähnlichkeiten zu den Schliefern und Elefanten.[5] Dem schloss sich Edwin H. Colbert 1941 nach der Untersuchung des umfangreichen Skelettmaterials fossiler Erdferkel der griechischen Insel Samos an,[61] auch George Gaylord Simpson unterstützte diese Ansicht 1945 in seiner generellen Taxonomie der Säugetiere und verwies die Tubulidentata als Ordnungsgruppe zusammen mit den „Condylarthra“ und den „Südamerikanischen Huftieren“ in die Überordnung der Protungulata.[62] Eine ebenfalls ähnliche Position hatte das Erdferkel bei Malcolm C. McKenna, der es in seiner Systematik der Säugetiere 1975 in die Gruppe der Ungulata einschloss,[63] was er später nochmals wiederholte.[64] Erst zum Ende des 20. Jahrhunderts kamen mit molekularbiologischen und biochemischen Analyseverfahren neue Perspektiven in die Verwandtschaftsverhältnisse der Höheren Säugetiere. Eine erste Untersuchung von Strukturproteinen Anfang der 1980er Jahre ordnete das Erdferkel in die Nähe der Schliefer, der Seekühe und der Elefanten.[65] Die Ergebnisse ließen sich in den folgenden Jahren mehrfach reproduzieren und gliederten unter anderem auch die Rüsselspringer in die nähere Verwandtschaft ein.[66] Bestätigt wurden diese ersten Untersuchungen dann durch genetische Analyseverfahren im Übergang vom 20. zum 21. Jahrhundert. Sie erbrachten eine unmittelbare Beziehung von in Afrika heimischen oder dort ursprünglich entstandenen Gruppen, denen neben den genannten zudem noch die Tenreks und Goldmulle angehören. Die dadurch extrem diverse Verwandtschaftsgruppe wurde dann unter dem Namen Afrotheria vereint.[67][68][69][37] Diese Sichtweise wird weiterhin auch durch Untersuchungen unterstützt, wonach das Erdferkel in seinem Genom spezifische Retroposons, sogenannte AfroSINEs, trägt, die es mit anderen Vertretern der Afrotheria gemein hat.[70][71]
Stammesgeschichte
Die Tubulidentata spalteten sich höchstwahrscheinlich schon sehr früh von den anderen Linien der Afrotheria ab, ihre Fossilgeschichte ist bisher aber nur für die letzten 20 Millionen Jahre belegt. Das früheste Auftreten fällt in den Beginn des Unteren Miozäns in Ostafrika, eine Radiation erfolgte aber in weiten Teilen Afrikas, Europas und Asiens. Neben einigen nicht genau zuweisbaren Unterkieferresten aus dem Muhoroni-Agglomerat im westlichen Kenia, die auf rund 23 Millionen Jahre datiert werden, stellen die Funde von Napak in Uganda den frühesten Nachweis dar. Geborgen wurde ein Teilskelett, das Grundlage für die im Jahr 2019 eingeführte Gattung Eteketoni bildete. Das Alter wird mit rund 20 bis 18,5 Millionen Jahren veranschlagt, die Tiere wogen rekonstruiert wohl rund 8,5 kg.[41] Wahrscheinlich nur wenig jünger sind die Fossilreste von Songhor, wie das Muhoroni-Agglomerat ebenfalls im westlichen Kenia gelegen. Sie gehören der Gattung Myorycteropus und genauer der Art M. minutus an. Das hier aufgefundene Material, auf dem die Erstbeschreibung beruht, setzt sich überwiegend aus dem Hand- und Fußskelett nebst einzelnen Zähnen zusammen.[72] Einige Unterkieferreste und Teile des Bewegungsapparates belegen die Art auch aus Arrisdrift im südlichen Namibia, sie weisen auf ein Tier hin, das möglicherweise nur 4 kg wog.[73] Ähnlich alt ist M. africanus, das ursprünglich anhand eines fragmentierten Schädels zuzüglich einiger Elemente des Körperskelettes eines nicht voll ausgewachsenen Tieres von der Insel Rusinga im Victoriasee beschrieben wurde.[74] Weitere Funde sind aus Mfwangano in geographischer Nähe zu Rusinga in Kenia überliefert.[72] Es war ebenfalls relativ klein, zeigte aber die stärksten Anpassungen an grabende Tätigkeiten, wie unter anderem aus dem Bau des Oberarmknochens geschlossen werden kann. Somit handelt es sich bei den Vertretern der Gattung Myorycteropus um relativ kleine Tiere, die im Maximum nur etwa halb so groß wurden wie die heutigen Erdferkel, aber schon zu einer myrmecophagen (auf Ameisen und Termiten spezialisierten) Ernährungsweise und einer fossorialen (unterirdisch grabenden) Lebensweise befähigt waren.[75][76][12]
Aus dem Mittleren Miozän (vor 16,0 bis 11,6 Millionen Jahren) sind nur wenige Funde aus Afrika bekannt. M. chemeldoi ist mit einzelnen Unterkieferresten und Teilen der Hand von den Tugen Hills in Kenia belegt. Die Backenzähne besaßen aber einen auffallend schmalen Bau.[72][75][12] Dafür sind zu dieser Zeit erstmals Vertreter der Tubulidentaten außerhalb Afrikas nachgewiesen. Sie gehören neueren Untersuchungen zufolge der Gattung Amphiorycteropus an. Höchstwahrscheinlich wanderten die frühen Erdferkel nach der Schließung der Tethys und der Entstehung einer Landbrücke nach Eurasien zusammen mit vielen anderen afrikanischen Säugetieren, etwa den Rüsseltieren, aus. Die ältesten Funde stammen mit einigen wenigen Zähnen und Fußelementen aus Paşalar im westlichen Anatolien.[77] Weitere Fossilien konnten unter anderem in der bedeutenden Fundstelle Çandir im zentralen Anatolien nordwestlich von Ankara geborgen werden. Sie werden zur Art A. seni gestellt, da aber das Material nur bruchstückhaft ist, sind die näheren Verwandtschaftsverhältnisse unklar.[78][76][12]
Im Oberen Miozän aus der Zeit vor 11,6 bis 5,3 Millionen Jahren erreichten die Tubulidentaten ihre höchste Vielfalt. Im östlichen Europa und westlichen Asien ersetzte A. pottieri die vorangegangene Form A. seni. Sie ist von einem teilweise erhaltenen Skelett aus Pentalofos in Makedonien und einem Unterkiefer vom Sinap Tepe nordwestlich von Ankara in der Türkei bekannt und stellte ein mittelgroßes Erdferkel dar, das im Gegensatz zu den heutigen Vertretern noch lange Eckzähne besaß.[79] Sehr weit verbreitet war A. gaudryi, das im Jahr 1888 anhand eines Schädels von der griechischen Insel Samos wissenschaftlich beschrieben wurde.[80] Die Art ist nicht nur der zuerst beschriebene, sondern auch der am besten untersuchte fossile Vertreter der Erdferkel.[61] Er besaß etwa drei Viertel der Größe der heutigen Erdferkel, war jedoch schlanker und kurzschnauziger. Sein Verbreitungsgebiet reichte vom südlichen Europa bis zum zentralen Asien. Die Art ist allerdings ein eher seltenes Begleitelement der spätmiozänen Säugetierfauna, bedeutende Funde stammen etwa in Form eines Unterkieferrestes von der griechischen Insel Euböa[81] und von Elementen des Fußskelettes aus der Yatağan-Formation in der südwestlichen Türkei.[82] Weniger bekannt und untersucht ist die zeitgleich auftretende Art A. browni, ein kleiner Vertreter, der über Schädelfragmente aus den Siwaliks von Pakistan bereits aus dem ausgehenden Mittleren Miozän belegt ist.[83] Im Oberen Miozän trat es auch im südöstlichen Europa auf, wo unter anderem ein Schädel aus dem Tal der Struma in Bulgarien zum Vorschein kam.[84] In Afrika sind aus dem Beginn des Oberen Miozäns bis vor 8 Millionen Jahren keine Funde von Tubulidentaten bekannt, was wohl auf die relativ geringe Anzahl von Fundstellen aus dieser Zeit zurückzuführen ist. Im nördlichen Teil des Kontinentes trat dann das größere A. mauretanicus auf, so in Bou Hanifia im Nordwesten Algeriens. Im nördlichen Tschad wurden mehrere Teilskelette eines weiteren Erdferkels entdeckt, die zu der Art A. abundulafus verwiesen werden und bereits in den Übergang zum Pliozän datieren. Ihre Angehörigen waren eher schlechte Gräber und besaßen sehr breite Zähne, zusammen mit den kräftigeren Muskelansatzstellen der Kaumuskeln kann auf festere Insektennahrung geschlossen werden, etwa Blatthornkäfer.[47][85] Leptorycteropus guilielmi stellt eine mittelgroße Form mit einer Schulterhöhe von 24 cm aus dem Übergang vom Oberen Miozän zum Pliozän dar, gefunden wurde ein Teilskelett und einzelne zusätzliche Knochen in der Nawata-Formation von Lothagam im Turkana-Becken im nordwestlichen Kenia. Der Erdferkel-Vertreter besaß im Vergleich zu Orycteropus ein untypisch gut ausgebildetes Gebiss, große Eckzähne, einen kräftigen Kiefer und zeigt keine Anpassungen an eine grabende Lebensweise. Vermutlich ernährte er sich nicht von staatenbildenden Insekten, sondern war eher omnivor.[48][75][76][12]
Das Pliozän (vor 5,3 bis 2,6 Millionen Jahren) markiert das Verschwinden der Erdferkel aus Eurasien. Im Unteren Abschnitt der Phase ist nur noch A. depereti mit einem vollständigen Schädel aus Perpignan in Frankreich nachgewiesen.[86] In Afrika hingegen zeichnet sich der Aufstieg der modernen Erdferkel ab. Der älteste Vertreter der Gattung Orycteropus ist O. djourabensis, der zuerst im nördlichen Tschad gefunden wurde und dessen Beschreibung auf einem teilweise erhaltenen Skelett beruht. Er ähnelte den heutigen Erdferkeln schon stark, besaß aber kürzere Gliedmaßen.[87] Es sind jedoch auch weitere Funde aus Ostafrika bekannt, so zwei Teilskelette von der frühmenschlichen Fundstelle Koobi Fora im nordwestlichen Kenia. Aus dem folgenden Pleistozän (vor 2,6 Millionen bis vor 10.000 Jahren) ist die Art O. crassidens belegt. Sie ist über mehrere Skelette von der Insel Rusinga im Viktoriasee nachgewiesen und erreichte die Größe des heutigen Erdferkels, besaß ihm gegenüber aber auch verhältnismäßig größere Zähne.[88] Darüber hinaus ist noch zahlreiches weiteres Fossilmaterial aus Afrika überliefert. Als einer der möglicherweise frühesten Nachweise des heutigen Erdferkels O. afer wurden einige Schädelreste und postcraniales Skelettmaterial aus der pliozänen Fossillagerstätte Langebaanweg im südwestlichen Südafrika interpretiert,[89] doch konnten dies neuere Untersuchungen nicht bestätigen.[88] Auch andere wichtige Fundstellen, etwa Laetoli in Tansania oder Swartkrans in Südafrika, bargen Funde von Erdferkeln, die der modernen Form sehr ähnlich sehen, sie sind häufig aber nicht aussagekräftig genug, um sie dieser oder einer anderen Art genau zuweisen zu können.[90][91][76]
Erdferkel und Menschen
Lokale Kunst und Kultur
Vereinzelt bildeten lokale Volksgruppen im südlichen Afrika das Erdferkel auf Felszeichnungen ab. Zwei Darstellungen finden sich im Mashonaland in Simbabwe. Sie wurden versteckt in einer Höhle angelegt: eine Darstellung zeigt eine Komposition gemeinsam mit sechs Menschen und einem Kaffernbüffel, die zweite ist isoliert. Beide Zeichnungen sind rötlichbraun gehalten und von kräftigen Punkten umgeben. Im südafrikanischen Glauben gelten Erdferkel als mächtige, aber auch geheimnisvolle Nachttiere, denen die Bantu magische Kräfte zuweisen. Auf letzteres deuten eventuell die Punkte hin.[92]
Bedrohung und Schutz

Das Erdferkel wird in vielen Regionen Afrikas als Nahrungsressource bejagt, sein Fleisch gilt im Geschmack als ähnlich zu Schwein. Dadurch landet es häufig auf lokalen Märkten, die Bushmeat anbieten insbesondere in Kenia, Sambia, Mosambik und dem Kongo. Traditionell gibt es mehrere Methoden, ein Erdferkel zu erbeuten, etwa durch Hineinkriechen in einen besetzten Bau, durch Ausräuchern[17] oder Fluten der Tunnel.[93] Jedoch gilt das Erdferkel als schwer zu erbeuten, da ein bedrängtes Tier mit den Krallen seiner Vorderfüße starke Verletzungen verursachen kann. Neben der Nahrungsgewinnung werden Teile der Tiere auch in der lokalen traditionellen Medizin eingesetzt oder dienen als Rohstoffe für Schmuckstücke, Amulette und Kuriositäten, etwa die Krallen, Zähne oder borstenartigen Haare.[94] Konflikte mit Menschen entstehen vor allem in land- und weidewirtschaftlich genutzten Gebieten, wo etwa das Nutzvieh durch Einbrechen der Decken der unterirdischen Baue verletzt oder Maschinen beschädigt werden können. Allerdings beeinflusst und kontrolliert das Erdferkel lokal die Populationen an Termiten und Ameisen. In der Vergangenheit kam es daher bei örtlicher Ausrottung der Tiere zu einem explosiven Anstieg der staatenbildenden Insekten, die dann teilweise bis zu 60 % Ernteverlust jährlich verursachten. Andererseits führt das Vernichten der örtlichen Termiten- und Ameisenkolonien auch zum Verschwinden der Erdferkelpopulation. Teilweise profitiert die Art darüber hinaus von der Erschließung einiger Regionen durch die Land- und Weidewirtschaft, da in deren Folge die Landschaften geöffnet und die Böden gelockert werden.[15][1][4][2][3]
Aufgrund der sehr weiten Verbreitung in Afrika südlich der Sahara stuft die IUCN das Erdferkel als „nicht gefährdet“ (least concern) ein. Die Populationen im südlichen Afrika werden als stabil angesehen, im westlichen und zentralen Afrika kann es in Folge der Ausdehnung menschlicher Wohn- und Wirtschaftsflächen zu einem starken Rückgang gekommen sein. Die Afrotheria Specialist Group der IUCN erfasst seit 2016 die Größe der Erdferkelpopulation, sowie die genetische Vielfalt der Tiere. Darüber hinaus ist die Aufklärung der lokalen Bevölkerung über die wichtige Funktion des Erdferkels im afrikanischen Ökosystem von vorrangiger Bedeutung. Die Art ist in zahlreichen geschützten Gebieten vertreten, aufgrund ihrer nächtlichen und teils unterirdischen Lebensweise aber schwer zu beobachten, was auch wissenschaftliche Monitoringprogramme beeinflusst.[95][94]
Erstmals wurde ein Erdferkel 1869 im Zoo London gezeigt. Die Haltung der Art kann in drei zeitliche Phasen unterteilt werden, wobei während der ersten im 19. Jahrhundert zumeist Wildfänge dominierten, die nur kurze Zeit in menschlicher Obhut überlebten. In der zweiten Phase, die Mitte der 1950er Jahre begann, gelang 1962 erstmals die erfolgreiche Geburt eines Jungtieres im Zoo in Frankfurt, das Neugeborene überlebte aber nur wenige Tage. Der Wissenszuwachs aus der Zoohaltung führte in den folgenden Jahrzehnten zu weiteren Geburten, Schwierigkeiten bei der Reproduktion in menschlicher Gefangenschaft resultierten aber letztendlich in Inzucht. Die dritte Phase startete im Jahr 2008 mit dem Einkreuzen von nichtverwandten Tieren aus afrikanischen Beständen. Gegenwärtig gibt es fünf Haltungen in Deutschland (Berlin, Halle, Köln, Frankfurt am Main und Saarbrücken), im gesamten Europa mehr als dreißig mit insgesamt über 50 Individuen (Stand 2021).[96] In zahlreichen Einrichtungen werden Erdferkel in speziellen Nachttierhäusern gehalten, in denen die Lichtverhältnisse im umgekehrten Tag-Nacht-Rhythmus geschaltet sind.[97]
Literatur
- Jonathan Kingdon, David Happold, Michael Hoffmann, Thomas Butynski, Meredith Happold und Jan Kalina (Hrsg.): Mammals of Africa Volume I. Introductory Chapters and Afrotheria. Bloomsbury, London, 2013, S. 288–295
- Ronald M. Nowak: Walker's Mammals of the World. Johns Hopkins University Press, 1999, ISBN 0-8018-5789-9
- Jeheskel Shoshani, Corey A. Golden und J. G. M. Thewissen: Orycteropus afer. Mammalian Species 300, 1988, S. 1–8
- W. Andrew Taylor: Order Tubulidentata. In: John D. Skinner und Christian T. Chimimba (Hrsg.): The Mammals of the Southern African Subregion. Cambridge University Press, 2005, S. 35–40
- W. Andrew Taylor: Family Orycteropodidae (Aardvark). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 2: Hooved Mammals. Lynx Edicions, Barcelona 2011, ISBN 978-84-96553-77-4, S. 18–27
Einzelnachweise
- Jeheskel Shoshani, Corey A. Golden und J. G. M. Thewissen: Orycteropus afer. Mammalian Species 300, 1988, S. 1–8
- W. Andrew Taylor: Family Orycteropodidae (Aardvark). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 2: Hooved Mammals. Lynx Edicions, Barcelona 2011, ISBN 978-84-96553-77-4, S. 18–27
- Jonathan Kingdon, David Happold, Michael Hoffmann, Thomas Butynski, Meredith Happold und Jan Kalina (Hrsg.): Mammals of Africa Volume I. Introductory Chapters and Afrotheria. Bloomsbury, London, 2013, S. 288–295
- W. Andrew Taylor: Order Tubulidentata. In: John D. Skinner und Christian T. Chimimba (Hrsg.): The Mammals of the Southern African Subregion. Cambridge University Press, 2005, S. 35–40
- W. E. Le Gros Clark und Charles F. Sonntag: A Monograph of Orycteropus afer. III. The skull. The skeleton of the trunk and Limbs. Proceedings of the Zoological Society of London 96 (2), 1926, S. 445–485
- John R. Wible: The Ear Region of the Aardvark, Orycteropus afer (Pallas, 1766) (Mammalia, Placentalia, Tubulidentata). Annals of Carnegie Museum 80 (2), 2012, S. 115–146
- Hans Virchow: Das Gebiß von Orycteropus aethiopicus. Zeitschrift für Morphologie und Anthropologie 34, 1934, S. 413–435
- P. Adloff: Das Gebiß von Orycteropus aethiopicus. Bemerkungen zu der gleichnamigen Abhandlung von Hans Virchow im Band 103 (Seite 696) dieser Zeitschrift. Zeitschrift für Anatomie und Entwicklungsgeschichte 104 (2), 1935, S. 203–206
- W. J. Schmidt: Polarisationsoptische Untersuchung des Zahnbeins von Orycteropus. Zeitschrift für Zellforschung und Mikroskopische Anatomie Abt. A 30 (4), 1940, S. 598–614
- Einar Lönnberg: On a new Orycteropus from Northern Congo and some remarks on the dentition of the Tubulidentata. Arkiv för Zoologi 3 (3), 1906, S. 1–35 ()
- Lucas J. Legendre und Jennifer Botha-Brink: Digging the compromise: investigating the link between limb bone histology and fossoriality in the aardvark (Orycteropus afer). PeerJ 6, 2018, S. e5216, doi:10.7717/peerj.5216
- Thomas Lehmann: Phylogeny and systematics of the Orycteropodidae (Mammalia, Tubulidentata). Zoological Journal of the Linnean Society 155, 2009, S. 649–702
- A. C. Allison: The structure of the stomach of the South African aardvark, Orycteropus afer. South African Journal of Science 63, 1947, S. 204–209
- Charles F. Sonntag und H. H. Woolland: A Monograph of Orycteropus afer. II. Nervous system, Sense-organs and Hairs. Proceedings of the Zoological Society of London 95 (3), 1925, S. 1185–1235
- Derek A. Melton: The biology of aardvark (Tubulidentata-Orycteropodidae). Mammal Review 6, 1976, S. 75–88
- Giuseppe Carpaneto und Francesco P. Germi: The mammals in the zoological culture of the Mbuti pygmies in North-Eastern Zaire. Hystrix (n.s.) 1, 1989, S. 1–83 (S. 36)
- Francesco Rovero und Daniela W. De Luca: Checklist of mammals of the Udzungwa Mountains of Tanzania. Mammalia 71 (1/2), 2007, S. 47–55
- D. A. Melton und C. Daniels: A note on the ecology of the aardvark Orycteropus afer. South African Journal of Wildlife Research 16 (3), 1986, S. 112–114
- R. J. van Aarde, C. K. Willis, J. D. Skinner und M. A. Haupt: Range utilization by the aardvark, Orycteropus afer (Pallas, 1766) in the Karoo, South Africa. Journal of Arid Environments 22, 1922, S. 387–394
- Gwenola Graff: Le représentations prédynastiques d'oryctéropes. In Jean-Claude Goyon und Christine Cardin (Hrsg.): Proceedings of the ninth International Congress of Egyptologists, Grenoble, 6-12 septembre 2004. Paris 2007, S. 829–835
- W. Andrew Taylor und John D. Skinner: Activity patterns, home ranges and burrow use of aardvarks (Orycteropus afer) in the Karoo. Journal of Zoology 261, 2003, S. 291–297
- Nora Marie Weyer, Andrea Fuller, Anna Jean Haw, Leith Carl Rodney Meyer, Duncan Mitchell, Mike Picker, Benjamin Rey und Robyn Sheila Hetem: Increased Diurnal Activity Is Indicative of Energy Deficit in a Nocturnal Mammal, the Aardvark. Frontiers in Physiology 11, 2020, S. 637, doi:10.3389/fphys.2020.00637
- W. Oduro und O.C. Boateng: Habitat ecology of aardvarks (Orycteropus afer) in Bui National Park, Ghana. Afrotherian Conservation 7, 2009, S. 7–11
- Robert T. Hatt: The pangolins and aardvarks of the American Museum Congo expedition. Bulletin of the American Museum of Natural History 66, 1934, S. 643–672
- G. M. Whittington-Jones, R. T. F. Bernard und D. M. Parker: Aardvark Burrows: A Potential Resource for Animals in Arid and Semi-Arid Environments. African Zoology 46 (2), 2011, S. 362–370
- Nora M. Weyer, W. Andrew Taylor und Thomas Lehmann: Demystifying aardvark tracks and how (not) to determine aardvark sex. Afrotherian Conservation 16, 2020, S. 4–7
- W. Andrew Taylor, P. A. Lindsey und John D. Skinner: The feeding ecology of the aardvark Orycteropus afer. Journal of Arid Environments 50, 2002, S. 135–152
- W. R. J. Dean und W. Roy Siegfried: Orientation of Diggings of the Aardvark. Journal of Mammalogy 72 (4), 1991, S. 823–824
- Jeremy Hollmann und Susan Myburgh: Aardvark and cucumber, a remarkable relationship. Veld & Flora 1995, S. 108–109
- R. Reason, D. Gierhahn und M. Schollhamer: Gestation in Aardvarks Orycteropus afer at Brookfield Zoo, Illinois. International Zoo Yearbook 39, 2005, S. 222–225
- W. Andrew Taylor und John D. Skinner: Associative feeding between Aardwolves (Proteles cristatus) and Aardvarks (Orycteropus afer). Mammal Review 30 (2), 2000, S. 141–143
- W. Andrew Taylor und John D. Skinner: Associative feeding between Ant-eating Chats, Myrmecocichla formicivora, and Aardvarks, Orycteropus afer. Ostrich 72 (3/4), 2001, S. 199–200
- C. J. Vernon und W. R. J. Dean: Further African bird-mammal feeding associations. Ostrich 59 (1), 1988, S. 38–39
- Steven Heritage, Erik R. Seiffert und Matthew R. Borths: Recommended fossil calibrators for time-scaled molecular phylogenies of Afrotheria. Afrotherian Conservation 17, 2021, S. 9–13
- Erik R Seiffert: A new estimate of afrotherian phylogeny based on simultaneous analysis of genomic, morphological, and fossil evidence. BMC Evolutionary Biology 7, 2007, S. 224, doi:10.1186/1471-2148-7-224
- Peter J. Waddell, Hirohisa Kishino und Rissa Ota: A Phylogenetic Foundation for Comparative Mammalian Genomics. Genome Informatics 12, 2001, S. 141–154
- Matjaž Kuntner, Laura J. May-Collado und Ingi Agnarsson: Phylogeny and conservation priorities of afrotherian mammals (Afrotheria, Mammalia). Zoologica Scripta 40 (1), 2011, S. 1–15
- Robert W. Meredith, Jan E. Janečka, John Gatesy, Oliver A. Ryder, Colleen A. Fisher, Emma C. Teeling, Alisha Goodbla, Eduardo Eizirik, Taiz L. L. Simão, Tanja Stadler, Daniel L. Rabosky, Rodney L. Honeycutt, John J. Flynn, Colleen M. Ingram, Cynthia Steiner, Tiffani L. Williams, Terence J. Robinson, Angela Burk-Herrick, Michael Westerman, Nadia A. Ayoub, Mark S. Springer und William J. Murphy: Impacts of the Cretaceous Terrestrial Revolution and KPg Extinction on Mammal Diversification. Science 334, 2011, S. 521–524
- Michael Buckley: A Molecular Phylogeny of Plesiorycteropus Reassigns the Extinct Mammalian Order ‘Bibymalagasia’. PlosOne 8 (3), 2013, S. e59614, doi:10.1371/journal.pone.0059614
- Martin Pickford: Orycteropodidae (Tubulidentata, Mammalia) from the Early Miocene of Napak, Uganda. Münchner Geowissenschaftliche Abhandlungen Reihe A: Geologie und Paläontologie 47, 2019, S. 1–101
- Thomas Lehmann: Amended taxonomy of the order Tubulidentata (Mammalia, Eutheria). Annals of the Transvaal Museum 44, 2007, S. 179–196
- J. A. J. Meester, I. L. Rautenbach, N. J. Dippenaar und C. M. Baker: Classification of southern African mammals. Transvaal Museum Monograph, 1986, S. 1–359 (S. 5)
- D. E. Wilson und D. M. Reeder: Mammal Species of the World. Johns Hopkins University Press, 2005. ISBN 0-8018-8221-4 ()
- Arthur Stanley Hirst: A new species of Orycteropus. The Annals and magazine of natural history 7 (17), 1906, S. 383–384 ()
- Robert T. Hatt: The aardvark of the Haut-Uele. American Museum Novitates 535, 1932, S. 1–2
- Thomas Lehmann, Patrick Vignaud, Andossa Likius und Michel Brunet: A new species of Orycteropodidae (Mammalia, Tubulidentata) in the Mio-Pliocene of northern Chad. Zoological Journal of the Linnean Society 143, 2005, S. 109–131
- Bryan Patterson: The fossil aardvarks (Mammalia: Tubulidentata). Bulletin of the Museum of Comparative Zoology 147, 1975, S. 185–237 (S. 216 und 219–223)
- Ross D. E. MacPhee: Morphology, adaptations, and relationships of Plesiorycteropus, and a diagnosis of a new order of eutherian mammals. Bulletin of the American Museum of Natural History 220, 1994, S. 1–214 (S, 200–203)
- Lars Werdelin: Bibymalagasia. In: Lars Werdelin und William Joseph Sanders (Hrsg.): Cenozoic Mammals of Africa.University of California Press, Berkeley, Los Angeles, London, 2010, S. 113–114
- Peter Simon Pallas: Miscellanea Zoologica. Petrum van Cleef, La Haye, 1766, S. 1–224 (S. 64) ()
- Peter Simon Pallas: Observationes circa Myrmecophagam africanam et Didelphidis novam speciem orientalem, e litteris celeberrimi Petri Camper excerplae et illustratae. Acta Academiae scientiarum imperialis Petropolitanae pro anno 1777, S. 223–231 ()
- Étienne Geoffroy Saint-Hilaire: Extrait d'un mémoire sur le Myrmecophaga capensis. Bulletin de la Société Philomathique de Paris 1, 1796, S. 101–102 ()
- John Edward Gray: On the natural arrangement of vertebrose animals. London Medical Repository 15, 1821, S. 297–310 (305) (PDF)
- Thomas Henry Huxley: A manual of the anatomy of vertebrated animals. Appleton, New York, 1872, S. 1–431 (S. 288–289) ()
- vgl. van Dale Groot Woordenboek, 3. Auflage, Van Dale Lexicografie, Utrecht/Amsterdam 2002 und Pharos Groot Woordeboek, 14. Auflage, Pharos, Kapstadt 1997
- Étienne Geoffroy Saint-Hilaire und Georges Cuvier: Memoire sur une nouvelle division des Mammiferes, et sur les principes qui doivent servir de base dans cette sorte de travail. Magasin Encyclopedique 2, 1795, S. 164–190 (S. 178–179, 188) ()
- Georges Cuvier: Tableau elementaire de l'histoire naturelle des animaux. Paris, Baudouin, 1798, S. 1–710 (S. 142–146) ()
- John Edward Gray: Revision of the genera and species of entomophagous Edentata, founded on the examination of the specimen in the British museum. Proceedings of the Zoological Society of London 1865, S. 359–386 ()
- William Henry Flower: On the mutual affinities of the animals composing the order Edentata. Proceedings of the Zoological Society of London 1882, S. 358–367 ()
- Edwin H. Colbert: A study of Orycteropus gaudryi from the island of Samos. Bulletin of the American Museum of Natural History 78, 1941, S. 305–351
- George Gaylord Simpson: The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History 85, 1945, S. 1–350 (S. 131 und 239)
- Malcolm C. McKenna: Toward a phylogenetic classification of the Mammalia. In W. Patrick Luckett und Frederick S. Szalay (Hrsg.): Phylogeny of the primates: a multidisciplinary approach. New York, London, 1975, S. 21–46
- Malcolm C. McKenna und Susan K. Bell: Classification of mammals above the species level. Columbia University Press, New York, 1997, S. 1–631 (S. 357)
- Wilfried W. de Jong, Anneke Zweers und Morris Goodman: Relationship of aardvark to elephants, hyraxes and sea cows from α-crystallin sequences. Nature 292, 1981, S. 538–540
- Malcolm C. McKenna: The alpha crystallin A chain of the eye lens and mammalian phylogeny. Annales Zoologice Fennici 28, 1992, S. 349–360
- Michael J. Stanhope, Victor G. Waddell, Ole Madsen, Wilfried de Jong, S. Blair Hedges, Gregory C. Cleven, Diana Kao und Mark S. Springer: Molecular evidence for multiple origins of Insectivora and for a new order of endemic African insectivore mammals. PNAS 95, 1998, S. 9967–9972
- Michael J. Stanhope, Ole Madsen, Victor G. Waddell, Gregory C. Cleven, Wilfried W. de Jong und Mark S. Springer: Highly Congruent Molecular Support for a Diverse Superordinal Clade of Endemic African Mammals. Molecular Phylogenetics and Evolution 9 (3), 1998, S. 501–508
- Mark S. Springer, Heather M. Amrine, Angela Burk und Michael J. Stanhope: Additional Support for Afrotheria and Paenungulata, the Performance of Mitochondrial versus Nuclear Genes, and the Impact of Data Partitions with Heterogeneous Base Composition. Systematic Biology 48 (1), 1999, S. 65–75
- Masato Nikaido, Hidenori Nishihara, Yukio Hukumoto und Norihiro Okada: Ancient SINEs from African Endemic Mammals. Molecular Biology and Evolution 20 (4), 2003, S. 522–527
- Hidenori Nishihara, Yoko Satta, Masato Nikaido, J. G. M. Thewissen, Michael J. Stanhope und Norihiro Okada: A Retroposon Analysis of Afrotherian Phylogeny. Molecular Biology and Evolution 22 (9), 2005, S. 1823–1833
- Martin Pickford: New fossil Orycteropodidae (Mammalia, Tubulidentata) from East Africa. Netherlands Journal of Zoology 25, 1975, S. 57–88
- Martin Pickford: Minute species of Orycteropus from the Early Middle Miocene at Arrisdrift, Namibia. Memoir Geological Survey of Namibia 19, 2003, S. 195–198
- D. G. Maclnnes: Fossil Tubulidentata from East Africa. Fossil Mammals of Africa 10, 1956, S. 1–38 ()
- Patricia A. Holroyd: Tubulidentata. In: Lars Werdelin und William Joseph Sanders (Hrsg.): Cenozoic Mammals of Africa.University of California Press, Berkeley, Los Angeles, London, 2010, S. 107–111
- Thomas Lehmann: Biodiversity of the Tubulidentata over geological time Afrotherian Conservation 4, 2006, S. 6–11
- Mikael Fortelius: Less common ungulate species from Paşalar, middle Miocene of Anatolia (Turkey). Journal of Human Evolution 19, 1990, S. 479–487
- Sevket Sen: Dispersal of African mammals in Eurasia during the Cenozoic: Ways and whys. Geobios 46, 2013, S, 159–172
- Louis de Bonis, Genevieve Bouvrain, Denis Geraads, George D. Koufos und Sevket Sen: The first aardvarks (Mammalia) from the Late Miocene of Macedonia, Geece. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 194 (2/3), 1994, S. 243–360
- Charles Immanuel Forsyth Major: Sur un gisement d’ossements fossiles dans l’île de Samos, contemporains de l’âge de Pikermi. Compte Rendu de l’Académie des Sciences de Paris107, 1888, S. 1178–1181 ()
- G. Theodorou, A. Athanassiou, S. Roussiakis und G. Iliopoulos: Preliminary remarks on the Late Miocene herbivores of Kerassiá (Northern Euboea, Greece). Deinsea 10, 2003, S. 519–530
- Tanju T. Kaya, Serdar Mayda, Dimitris S. Kostopoulos, Mehmet Cihat Alcicek, Gildas Mercerond, Aytekin Tan, Seval Karakutuk, Amanda K. Giesler und Robert S. Scott: Şerefköy-2, a new Late Miocene mammal locality from the Yatağan Formation, Muğla, SW Turkey. Comptes Rendus Palevol 11, 2012, S. 5–12
- Edwin H. Colbert: The presence of tubulidentates in the Middle Siwalik beds of Northern India. American Museum Novitates 604, 1933, S. 1–10
- Nikolaï Spassov, Tzank Tzankov und Denis Geraads: Late Neogene stratigraphy, biochronology, faunal diversity and environments of South-West Bulgaria (Struma River Valley). Geodiversitas 28 (3), 2006, S. 477–498
- Thomas Lehmann, Patrick Vignaud, Andossa Likius, Hassane Taïsso Mackaye und Michel Brunet: A sub-complete fossil aardvark (Mammalia, Tubulidentata) from the Upper Miocene of Chad. Comptes Rendus Palevol 5, 2006, S. 693–703
- H. Helbing: Ein Orycteropus-Fund aus dem unteren Pliocaen des Roussillon. Eclogae geologicae Helvetiae 26, 1933, S. 256–267
- Thomas Lehmann, Patrick Vignaud, Hassane Taïsso Mackaye und Michel Brunet: A fossil aardvark (Mammalia, Tubulidentata) from the lower Pliocene of Chad. Journal of African Earth Sciences 40, 2004, S. 201–217
- Thomas Lehmann: Plio-Pleistocene aardvarks (Mammalia, Tubulidentata) from East Africa. Fossil Record 11 (2), 2008, S. 67–81
- Martin Pickford: Orycteropus (Tubulidentata, Mammalia) from Langebaanweg and Baard’s Quarry, Early Pliocene of South Africa. Comptes Rendus Palevol 4, 2005, S. 715–726
- Terry Harrison: Orycteropodidae. In: Terry Harrison (Hrsg.): Paleontology and Geology of Laetoli: Human Evolution in Context. Volume 2: Fossil Hominins and the Associated Fauna. Vertebrate Paleobiology and Paleoanthropology, Springer, 2011, S. 263–274
- Thomas Lehmann: Fossil aardvark (Orycteropus) from Swartkrans Cave, South Africa. South African Journal of Science 100, 2004, S. 311–314
- David Coulson und Alec Campbell: Afrikanische Felsenbilder. Malereien und Gravuren auf Stein. Weingarten, 2001, S. 1–256 (S. 94)
- Galen B. Rathbun: Aardvark hunt in Kenya. Afrotherian Conservation 8, 2011, S. 13–14
- Andrew Taylor und Thomas Lehmann: Orycteropus afer. The IUCN Red List of Threatened Species. Version 2015.2. (); zuletzt abgerufen am 11. November 2021
- Afrotherian Specialist Group: Conservation Fact Sheet. Aardvark (Tubulidentata) (), abgerufen am 11. November 2021
- Zootierliste: Erdferkel Orycteropus afer (Pallas, 1766). () auf Zootierliste.de. Aufgerufen am 4. September 2021
- Astrid Parys, Thomas Lehmann, Wineke Schoo und Thomas M. Wilms: Newcomers enrich the European zoo aardvark population. Afrotherian Conservation 9, 2012, S. 2–5
Weblinks
- Orycteropus afer in der Roten Liste gefährdeter Arten der IUCN 2010. Eingestellt von: A. Taylor & T. Lehmann, 2008. Abgerufen am 11. November 2021.
- The Biology of the Aardvark (Orycteropus afer) Diplomarbeit (PDF-Datei; 2,01 MB)
- Orphaned aardvark at Naankuse Wildlife Sanctuary, Windhoek, Namibia Video in der Encyclopædia Britannica Online