Otterspitzmäuse
Die Otterspitzmäuse (Potamogalidae) sind eine Familie der Säugetiere bestehend aus drei Arten in zwei Gattungen: die Große Otterspitzmaus sowie die zwei Arten der Kleinen Otterspitzmäuse, die Zwerg- und die Ruwenzori-Otterspitzmaus. Alle drei Vertreter kommen nur auf dem afrikanischen Festland vor und bewohnen die tropischen Regenwälder beiderseits des Äquators. Die Tiere bevorzugen Landschaften mit kleinen, klaren Gewässern. Sie sind an das Wasserleben angepasst und gehen in der Nacht schwimmend auf Nahrungssuche. Dabei ernähren sie sich hauptsächlich von Krabben und Fischen. Allgemein leben sie einzelgängerisch und verbringen den Tag in einem Bau. Die genaue Lebensweise der Otterspitzmäuse ist bisher kaum erforscht. in ihrem Erscheinungsbild sind sie an das Leben im Wasser angepasst. Sie haben eine otterähnliche Gestalt mit einer abgeplatteten Schnauze, die zahlreiche Tasthaare aufweist, und einem muskulösen Schwanz. Die Fortbewegung im Wasser erfolgt hauptsächlich durch Schlängelbewegungen des Schwanzes, eine Art besitzt allerdings Schwimmhäute und setzt auch ihre Beine paddelnd ein.
| Otterspitzmäuse | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Große Otterspitzmaus (Potamogale velox) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Potamogalidae | ||||||||||||
| Allman, 1865 |
Die nächsten Verwandten der Otterspitzmäuse stellen die Tenreks (Tenrecidae) dar, die endemisch auf Madagaskar verbreitet sind. Die beiden Gruppen trennten sich bereits vor nahezu 50 Millionen Jahren. Teilweise wurden die Otterspitzmäuse auch als afrikanischer Zweig der Tenreks angesehen, aus molekulargenetischer Sicht befürwortet der lange Abspaltungszeitraum neben einigen anatomischen Unterschieden einen eigenständigen Familienstatus. Die Otterspitzmäuse sind bisher nur selten im Fossilbericht belegt, einige wenige Funde stammen aus dem südwestlichen Afrika und weisen ein Alter von rund 40 Millionen Jahre auf. Die ersten Berichte von heutigen Vertretern der Familie datieren in die 1860er Jahre. Sie beziehen sich auf die Große Otterspitzmaus, die beiden Angehörigen der Kleinen Otterspitzmäuse wurden erst rund einhundert Jahre später entdeckt. In der Regel wurden die Otterspitzmäuse in ihrer forschungsgeschichtlichen Vergangenheit als Teil der Insektenfresser angesehen. Im Übergang vom 20. zum 21. Jahrhundert erbrachten aber genetische Untersuchungen eine nähere Beziehung zu anderen originär afrikanischen Tieren. Von den drei heutigen Arten wird eine als in ihrem Bestand gefährdet eingestuft.
Merkmale
Habitus
Otterspitzmäuse sind kleine Vertreter der Säugetiere. Sie besitzen eine Kopf-Rumpf-Länge von 12 bis 20 cm bei den Kleinen Otterspitzmäuse (Micropotamogale) und von 30,5 bis 33,7 cm bei der Großen Otterspitzmaus (Potamogale). Der Schwanz wird zwischen 9,5 und 29,0 cm lang, er erreicht zwischen 77 und 84 % der Länge des übrigen Körpers. Das Körpergewicht variiert von 32 bis 780 g. Wie der Name schon andeutet, haben die Tiere in ihrem äußeren Erscheinungsbild eine entfernte Ähnlichkeit mit Ottern. Ihr Körper ist stromlinienförmig, die Schnauze breit und abgeflacht mit unterständigem Maul. Der Körper wird von einem dichten Unterfell bedeckt, das von glänzenden Überhaaren überlagert wird. Der Glanz wird durch die abgeflachten Spitzen der Haare hervorgerufen. Das Fell ist an der Oberseite dunkelbraun und an der Unterseite weißlich gefärbt. Eine Ausnahme bildet die Zwerg-Otterspitzmaus (Micropotamogale lamottei), deren Körper einheitlich dunkel gefärbt erscheint. Ein markantes Merkmal bildet der lederige Nasenspiegel, der zumeist herzförmig gestaltet ist und von einer Längsfurche geteilt wird. Die sich seitlich-oberhalb befindenden Nasenlöcher können durch kleine Klappen verschlossen werden. An der Oberlippe besteht ein Kranz aus steifen Vibrissen, weitere Tasthaare finden sich über den Augen und am Unterkiefer. Die Augen selbst sind klein, die Ohrmuscheln haben eine gestreckt-ovale Form und ragen aus dem Fell heraus. Der Schwanz ist kräftig und lang gebaut, bei der Großen Otterspitzmaus zeigt er eine seitlich deutlich abgeplattete Form, bei den Ruwenzori-Otterspitzmaus (Micropotamogale ruwenzorii) ist er nur leicht abgeplattet, während er bei der Zwerg-Otterspitzmaus einen runden Querschnitt aufweist. Die kurzen und kräftigen Gliedmaßen enden in jeweils fünf Strahlen. Schwimmhäute zwischen den Fingern und Zehen kommen lediglich bei der Ruwenzori-Otterspitzmaus vor. Typisch für alle Otterspitzmäuse bilden der zweite und dritte Strahl des Hinterfußes eine funktionale Einheit, die Zehen sind also syndactyl. Alle Finger und Zehen tragen gebogene Krallen.[1][2][3][4]
Schädel- und Gebissmerkmale
Der Schädel der Otterspitzmäuse ist allgemein schmal, die größte Breite wird am Hirnschädel erreicht. Dadurch wirkt der Schädel in der Aufsicht tropfenförmig mit einem lang ausgezogenen Rostrum. In Seitenansicht verläuft die Stirnlinie nahezu gerade. Auf dem Scheitelbein besteht ein Scheitelkamm.[5][6] Der Jochbogen ist nicht geschlossen. Unter dem hinteren Bogenansatz am Schläfenbein befindet sich die Glenoidgrube, der Ansatzstelle des Unterkiefergelenks. Der Tränen-Nasen-Gang (Ductus nasolacrimalis) ebenso wie das Tränenloch (Foramen lacrimale) fehlen, was mit der Reduktion des Geruchssinnes zusammenhängt.[7] Auffallend an der Schädelbasis ist weiterhin eine verknöcherte Paukenblase.[8][3]
Das Gebiss setzt sich aus insgesamt 40 Zähnen mit folgender Zahnformel zusammen: . Ein charakteristisches Merkmal bilden der vergrößerte erste obere und der zweite untere Schneidezahn. Sie erinnern in ihrem Aussehen an Eckzähne und wirken als Gegenspieler beim Ergreifen der Beute. Die nachfolgenden Zähne einschließlich des Eckzahns und der vorderen Prämolaren zeigen einen einfachen Bau. Zwischen den einzelnen Arten bestehen Unterschiede in der Größenausprägung dieser Zähne. So sind bei der Zwerg-Otterspitzmaus deutliche Variationen erkennbar, während bei der Großen Otterspitzmaus alle Zähne in diesem Abschnitt etwa die gleichen Ausmaße haben. Dadurch wirkt das Gebiss bei den jeweiligen Arten unterschiedlich stark spezialisiert. Der obere Eckzahn weist darüber hinaus immer zwei Wurzeln auf. Die Molaren verfügen über ein markantes Kauflächenmuster, das drei Haupthöcker umfasst: den Para-, Meta- und Protoconus (bezogen auf die Oberkieferzähne). Der Protoconus ist gut ausgebildet, der Metaconus verschmilzt abweichend von den Tenreks nicht so deutlich mit dem Paraconus, letzterer stellt den Haupthöcker der Mahlzähne dar. Auch hier sind Unterschiede zwischen den Arten ausgeprägt, so ist die Trennung zwischen den beiden letztgenannten Höckern bei der Großen Otterspitzmaus auffallender als bei den Kleinen Otterspitzmäusen. Charakteristisch ist überdies eine V-förmige Scherleiste (Ectoloph), deren Spitze der Paraconus bildet. Die Mahlzähne können daher als weitgehend zalambdodont aufgefasst werden. Der hinterste obere Molar zeigt eine starke Größenreduzierung.[2][9][3]

Skelettmerkmale
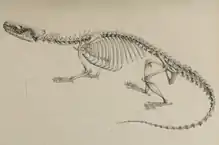
Die Wirbelsäule der Otterspitzmäuse besteht aus 7 Hals-, 16 bis 17 Brust-, 5 bis 6 Lenden-, 5 Kreuzbein- und 31 bis 34 Schwanzwirbeln. Untersuchte Jungtiere der Zwerg-Otterspitzmaus besitzen 24 bis 25 Schwanzwirbel.[10] An den vorderen Schwanzwirbeln treten kräftig entwickelte Dorn- und Querfortsätze sowie robuste Hämalbögen auf. Ihre Größe nimmt nach hinten ab[11][12][13]
Ein auffälliger Unterschied zu den Tenreks besteht im Fehlen des Schlüsselbeins bei den Otterspitzmäusen. Das Schulterblatt ist bei der Großen Otterspitzmaus sehr lang und schmal, allerdings besitzt es abweichend von den Tenreks kein ausgeprägtes Acromion, wo normalerweise Teile des Deltamuskels ansetzen. Ebenfalls im Gegensatz zu den Tenreks weist der Oberarmknochen gegenüber den Knochen des Unterarms eine sehr langgestreckte Form auf. Der große Tuberkel des Humerus ragt als krallenartiger Fortsatz steil auf und erinnert hier weniger an Tenreks als eher an die Fledermäuse. Er blockiert gegen das Schulterblatt und verhindert so zu weit ausholende Armbewegungen. Das untere Humerusgelenk (Ellenbogen) ist sehr schlank, allerdings ist das Capitulum breit und rechteckig. An der Elle, die eine kurze, kompakte Gestalt aufweist, fällt der langgestreckte obere Gelenkfortsatz, das Olecranon, auf. Es nimmt rund 19 % der Länge des Gesamtknochens ein. Der gesamte Bau des Ellenbogengelenks zeigt, dass es auf die Stabilisierung des Vorderarms ausgerichtet ist und nur eingeschränkte seitliche Bewegungen zulässt. Die Handwurzelknochen sind in ihrer Anzahl reduziert, eventuell ist kein Os centrale ausgebildet oder mit dem Kahnbein verwachsen, teilweise treten auch Verwachsungen zwischen dem Kahn- und dem Mondbein auf. Die Verringerung der Anzahl der Handwurzelknochen ähnelt der der grabenden Tenreks und hilft wie bei diesen, die Hand beim Schwimmen zu stabilisieren. Dagegen ist ein Präpollex, eine fingerartige Verlängerung am Großen Vieleckbein, abweichend von den Tenreks nicht vorhanden oder stark verkleinert. Dafür tritt ein Fortsatz am Kahnbein auf, der wohl dessen Position einnimmt. Im Vergleich zu den Handwurzelknochen sind die Mittelhandknochen ausgesprochen langgestreckt.[14][15]
Der Oberschenkelknochen ist in Bezug auf das Becken eher kurz, was in Verbindung mit der nach oben orientierten Hüftgelenkpfanne eine erhöhte Beinhebelkraft bewirkt. Am Femurschaft besteht nur ein kleiner dritter Rollhügel, der im Unterschied zu den Tenreks weit nach unten verlagert ist und so einen kräftigen Gesäßmuskel unterstützt. Schein- und Wadenbein sind im unteren Drittel miteinander verwachsen. Beide Knochen verlaufen weitgehend gerade und sind am Schaft nicht deutlich gebogen. Das Fußgelenk ist in seiner Struktur stark abweichend von dem der Tenreks. Dies betrifft etwa die langgestreckten und schmalen Knöchel, die die Rotationsfähigkeit des Fußes einschränken. Dies bewirkt auch der scharfe Übergang von der Sprungbeinrolle zum Gelenkhals am Sprungbein. Letzterer ist außerdem extrem kurz. Weitere markante Unterschiede finden sich am Fersenbein mit seinem extrem langen Fersenhöcker. Am Fuß sind die Mittelfußknochen des inneren und äußeren Strahls vergleichsweise länger geformt als bei den Tenreks.[16][17]
Weichteile
Wie bei den Tenreks auch bilden die Geschlechts-, Verdauungs- und Ausscheidungsorgane eine gemeinsame Austrittsöffnung, die Kloake. Der Magen ist einfach gebaut und von ovaler Form. Der Darm bestehend aus dem Dünndarm und dem Dickdarm erreicht bei der Ruwenzori-Otterspitzmaus eine Gesamtlänge von 55 cm,[1] anderen Angaben zufolge bis 68 cm,[18] was in etwa der vierfachen Körperlänge des Tieres entspricht, der Blinddarm fehlt. Die Bauchspeicheldrüse ist stark verästelt.[18] Die Hoden befinden sich bei den Otterspitzmäusen in der Beckengegend auf Höhe des Schambeins. Sie liegen in faserigen Hauttaschen (Cremastersäcken), die einem inneren Pseudo-Hodensack entsprechen. Die Position der Hoden unterscheidet sich von einigen Tenreks wie den Reistenreks, wo sie in der Nähe der Nieren verbleiben. Der Penis ist vergleichsweise lang.[10][1] Weibchen besitzen eine zweihörnige Gebärmutter, die Plazenta ist im Unterschied zu den Tenreks endotheliochorial.[19]
Das Gehirn der Otterspitzmäuse ist relativ einfach gebaut. Es fällt durch eine Verkleinerung des Riechkolbens und durch eine Vergrößerung der Medulla oblongata auf. Von letzterer geht der Trigeminus-Nerv aus, der unter anderem die Tasthaare steuert. Es handelt sich hier wahrscheinlich um eine Anpassung an das Wasserleben, bei dem der Tastsinn als wichtigstes Sinnesorgan den Geruchssinn ablöst. Die Merkmale sind bei der Zwergotterspitzmaus am geringsten, bei der Großen Otterspitzmaus am stärksten ausgebildet, was als graduelle Anpassung an die semi-aquatische Lebensweise gedeutet wird und sich auch in anderen Körpermerkmalen wiederfindet. Das Volumen des Gehirns ist im Vergleich zu den nahe verwandten Tenreks größer. Es variiert zwischen 720 bis 1270 mg bei den Kleinen Otterspitzmäusen und zwischen 3720 und 4640 mg bei der Großen Otterspitzmaus.[20][21][22]
Verbreitung und Lebensraum
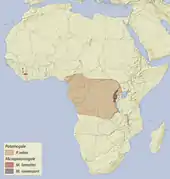
Die Otterspitzmäuse kommen endemisch im zentralen und westlichen Afrika vor. Die Große Otterspitzmaus hat das größte Verbreitungsgebiet, es umfasst das gesamte Kongobecken und schließt die angrenzenden Gebiete vom östlichen Nigeria im Norden bis nach Uganda im Osten und bis in das nördliche Angola und Sambia im Süden ein. Im Osten überlappt es sich mit dem der Ruwenzori-Otterspitzmaus, das allerdings nur aus einem schmalen Streifen westlich des Ostafrikanischen Grabens vom Kivusee bis zum Ruwenzori-Gebirge besteht. Die Zwerg-Otterspitzmaus lebt abgetrennt von den beiden anderen Arten im Grenzgebiet von Liberia, der Elfenbeinküste und Guinea. Die Tiere bevorzugen tropische Regenwälder vom Meeresspiegelniveau bis in teils höher gelegene Areale bis 2200 m. Der Lebensraum umfasst Waldgebiete mit kleinen, langsam fließenden und klaren Flüssen oder Bächen sowie mit Tümpeln und Sumpfgebieten. Die Wassertemperaturen liegen meist über 20 °C, in Gebirgslagen können sie aber auch auf bis zu 12 °C absinken. In der Regel werden größere Flüsse gemieden.[18][3][4]
Lebensweise
Territorialverhalten und Fortbewegung
Die Lebensweise der Otterspitzmäuse ist kaum untersucht. Alle drei Arten gelten als nachtaktiv, die aktive Phase wird von mehreren kurzen Ruheperioden unterbrochen.[18][23] Tagsüber ziehen sich die Tiere in Baue in den Uferhängen zurück. Die Baue enthalten eine Nestkammer, die mit Blättern und anderem Pflanzenmaterial ausgepolstert ist. Die Tiere leben höchstwahrscheinlich einzelgängerisch und kommen außer zur Paarungszeit nicht oder nur selten mit anderen Artgenossen zusammen. Möglicherweise herrscht unter den einzelnen Individuen Territorialität vor, worauf unter anderem die Ablage von Kot in speziellen Latrinen hindeutet.[18][23][24] Charakteristisch ist das Kratzen mit den syndactylen Zehen des Hinterfußes, was als Komfortverhalten interpretiert wird.[3][4]
Otterspitzmäuse sind mit ihren charakteristischen Körperbau gut an ein Leben im Wasser angepasst. Sie bewegen sich schnell schwimmend vorwärts. Die Schwimmbewegungen der drei Arten unterscheiden sich leicht voneinander. Bei der Großen Otterspitzmaus sorgen seitlich schlängelnde Bewegungen des Körpers und vor allem des seitlich abgeplatteten Schwanzes für den Vortrieb, die Beine werden an den Körper angelegt, die Vorderbeine haben nur einen eingeschränkten Bewegungsspielraum.[23][15] Dagegen paddelt die Ruwenzori-Otterspitzmaus mit den mit Schwimmhäuten versehenen Vorder- und Hinterfüßen. Der Schwanz hat bei ihr einen eher rundlichen Querschnitt, die undulierenden Körperbewegungen bleiben aber trotzdem erhalten.[18] Die Zwerg-Otterspitzmaus mit ihren eher kleinen Füßen ohne Schwimmhäute und ebenfalls rundlichen Schwanz kombiniert beide Möglichkeiten, sie unduliert mit dem Körper sowie dem Schwanz und paddelt gelegentlich mit den Füßen. Teilweise wird ihre Schwimmbefähigung auch als eher ursprünglich im Vergleich zu den beiden anderen Vertretern bewertet.[25] In der Regel trocknet das Fell nach jedem Wassergang innerhalb kürzester Zeit.[24] An Land laufen die Otterspitzmäuse auf den Sohlen, der Schwanz schleift dabei auf dem Boden und zieht eine gerade Linie.[18][23][3][4]
Ernährung
Alle Otterspitzmäuse ernähren sich fleischfresserisch, sie sind in erster Linie auf Krabben und Fische spezialisiert. Darüber hinaus verzehren sie auch Insekten und Frösche. letztere gehören aber nicht zur bevorzugten Nahrung. Die Beute wird vorwiegend mit dem Tastsinn aufgespürt, untergeordnet auch über den Geruch. Zum Beutefang unternehmen die Tiere kurze Tauchgänge von wenigen Sekunden Dauer.[18][23][25] In der Regel erlegen sie ihre Beute mit Bissen, eine Manipulation mit den Vorderbeinen erfolgt nicht. Kleinere Beute fressen sie direkt im Wasser, größere schaffen sie aber auch an Land. Vor allem die kleineren Otterspitzmäuse nehmen täglich eine Nahrungsmenge zu sich, die nahezu ihrem Körpergewicht entspricht.[3][4]
Fortpflanzung
Über die Fortpflanzung der Otterspitzmäuse ist wenig bekannt. Lediglich bei der Zwerg-Otterspitzmaus wurde bisher Geburten beobachtet, bei denen bis zu vier Jungen zur Welt kamen. Diese sind als Nesthocker nackt und blind.[26] Bei den beiden anderen Arten wurden bisher nur trächtige Weibchen dokumentiert, die durchschnittlich zwei Embryonen austrugen.[18][23] Ein trächtiges Weibchen der Großen Otterspitzmaus war dabei in Begleitung von zwei Jungtieren, so dass von wenigstens zwei Würfen im Jahr ausgegangen werden kann. Weibliche Otterspitzmäuse besitzen vier Paare an Zitzen: zwei in der Brust und jeweils eins in der Bauch- und in der Leistengegend.[3] Analog zu den Tenreks entstehen bei der Entwicklung der Eibläschen keine Graafschen Follikel.[27][4]
Systematik
Äußere Systematik
Innere Systematik der Afrotheria nach Heritage et al. 2021[28]
|
Die Otterspitzmäuse bilden eine Familie innerhalb der Ordnung der Tenrekartigen (Afrosoricida), zu der außerdem noch die Tenreks (Tenrecidae) und die Goldmulle (Chrysochloridae) gestellt werden. Als nächste Verwandte sind die Tenreks aufzufassen, beide Gruppen zusammen formen das übergeordnete Taxon der Tenrecomorpha.[29] Die Tenrekartigen wiederum gehören zur Überordnung der Afrotheria, einer der vier Hauptlinien innerhalb der Höheren Säugetiere. Die Afrotheria als Verwandtschaftsgemeinschaft umfassen verschiedene, überwiegend in Afrika heimische oder von dort stammende Tiergruppen. Ihre Zusammengehörigkeit beruht vor allem auf molekulargenetischen Untersuchungen weniger auf anatomischen Gemeinsamkeiten. Es können innerhalb der Afrotheria zwei größere Linien unterschieden werden: die Paenungulata und die Afroinsectiphilia. Zur ersteren gehören die heutigen Elefanten, die Schliefer und die Seekühe, die Gruppe galt schon länger als eine gemeinsame Abstammungseinheit. Die letztere Linie vereint neben den Tenrekartigen auch die Rüsselspringer und das Erdferkel. Die genaueren Verwandtschaftsverhältnisse der drei letztgenannten Gruppen sind noch nicht vollständig geklärt. In einigen genetischen Untersuchungen besteht eine Schwestergruppenbeziehung zwischen den Afrosoricida und den Rüsselspringern, beide Gruppen werden dann als Afroinsectivora zusammengefasst.[30][31][32] Andere wiederum sehen die Tenrekartigen in einer Schwestergruppenposition zu allen anderen Afrotheria, die Rüsselspringer zeigen dann eine engere Beziehung zum Erdferkel.[33] Laut den molekulargenetischen Untersuchungen zufolge entstanden die Afrotheria bereits in der Oberkreide vor 90,4 bis 80,9 Millionen Jahren, ihre Aufspaltung in die beiden Hauptgruppen begann etwa 15 Millionen Jahre später. Die Afrosoricida wiederum bildeten sich dann kurz vor der Kreide-Tertiär-Grenze vor etwa 68 Millionen Jahren heraus.[32] Rund zehn Millionen später spalteten sich die Goldmulle von der gemeinsamen Linie mit den Tenreks und den Otterspitzmäusen ab. Die Trennung der beiden letztgenannten erfolgte im Unteren Eozän vor etwa 48 Millionen Jahren.[29]
Innere Systematik
Innere Systematik der Otterspitzmäuse nach Everson et al. 2016[29]
|
Die Familie der Otterspitzmäuse ist im Gegensatz zu den nahe verwandten, aber auf Madagaskar beschränkten Tenreks wenig divers. Sie besteht heute aus drei Arten in zwei Gattungen. In der Regel wird folgende Gliederung unternommen:[29]
- Familie Potamogalidae Allman, 1865
- Potamogale Du Chaillu, 1860
- Potamogale velox (Du Chaillu, 1860) (Große Otterspitzmaus)
- Micropotamogale Heim de Balsac, 1954 (Kleine Otterspitzmäuse)
- Micropotamogale lamottei Heim de Balsac, 1954 (Zwerg-Otterspitzmaus)
- Micropotamogale ruwenzorii (de Witte & Frechkop, 1955) (Ruwenzori-Otterspitzmaus)
Allen Otterspitzmäusen gemein ist das Fehlen des Schlüsselbeins, der syndactyle zweite und dritte Strahl des Hinterfußes, der zweiwurzelige Eckzahn und die eher deutliche Trennung von Paraconus und Metaconus an den Molaren des Oberkiefers, was sie von den Tenreks unterscheidet. In ihrer generellen Morphologie stimmen die drei Arten der Otterspitzmäuse überein, dennoch gibt es einige bedeutende Abweichungen im Detail, die sich neben allgemeinen Größenunterschieden in der Ausformung des Schwanzes, dem Vorhandensein oder Fehlen von Schwimmhäuten und in speziellen Gebissmerkmalen ausdrücken. Einige der genannten Merkmalsunterschiede bedingen Abweichungen in der Lebensweise, etwa bei der schwimmenden Fortbewegung der Tiere. Die charakteristischen Schwimmhäute der Ruwenzori-Otterspitzmaus verleiteten Henri Heim de Balsac im Jahr 1956 dazu, die Art in die eigene Gattung Mesopotamogale zu stellen,[34] was sich aber nicht allgemein durchsetzte.[10][13] Teilweise wird aber heute noch auf die Berechtigung von Heim de Balsacs Einordnung hingewiesen, womit jede Art der Otterspitzmäuse einen eigenständigen Gattungsstatus besäße.[3]
Fossil wird gegenwärtig nur eine Gattung anerkannt:[35]
- Namagale Pickford, 2015
Der Status der Otterspitzmäuse wurde in der Vergangenheit unterschiedlich bewertet. Teilweise galten sie als eigenständige Familie,[36][16][35] teilweise sah man sie als Unterfamilie der Tenreks an.[37][38][39] Einige Wissenschaftler nahmen auch eine engere Beziehung zum Wassertenrek (Microgale mergulus) an, der ebenfalls an ein Leben im Wasser angepasst ist und wie die Ruwenzori-Otterspitzmaus über Schwimmhäute verfügt. So wurde 1957 von Serge Frechkop die Unterfamilie Limnogalinae für die Otterspitzmäuse und den Wassertenrek vorgeschlagen.[40] In skelettanatomischen Untersuchungen aus dem Jahr 1999 kam Robert J. Asher zu dem Schluss, dass der Wassertenrek das Schwestertaxon zu den Otterspitzmäuse bildet.[41] Genetische Studien bestätigten aber die Zugehörigkeit des Wassertenreks zu den madagassischen Tenreks und somit die Monophylie der gesamten Gruppe.[42][43] Dies zeigt auch eine Analyse aus dem Jahr 2016, die alle bisher bekannten Vertreter der Tenreks und der Otterspitzmäuse einschloss. Sie kommt überdies zu dem Schluss, dass aufgrund der frühen Abspaltung der Otterspitzmäuse von den madagassischen Tenreks und genügend morphologisch-anatomischer Merkmalsunterschiede ein eigener Familienstatus für erstere gerechtfertigt ist.[29]
Forschungsgeschichte
Taxonomie

Der heute gebräuchliche wissenschaftliche Name Potamogalidae für die Otterspitzmäuse geht auf George James Allman zurück, der ihn im Jahr 1866 prägte. Allman gab dabei in seinem Aufsatz eine umfassende Beschreibung der morphologischen sowie skelettanatomischen Merkmale der Großen Otterspitzmaus ab und berief sich auf die Gattungsbezeichnung Potamogale für die Art. Diese wiederum stammt von Paul Belloni Du Chaillu aus dem Jahr 1860. Du Chaillu definierte Potamogale allerdings nicht, sondern merkte den Namen nur als mögliche Gattungszuordnung an.[44] Erst durch die Veröffentlichung von Allmans Aufsatz sechs Jahre später konnte Potamogale somit wissenschaftlich etabliert werden, im gleichen Zug wies er der Gattung einen eigenständigen Familienstatus zu.[5] Nicht unumstritten ist der Verweis auf Allman 1865 als erste Namensnennung. Hierbei handelt es sich lediglich um eine Ankündigung des neuen Aufsatzes, der erst ein Jahr später herausgegeben wurde.[45] Da darin aber der Familienname angegeben wird, führen in der jüngeren Zeit die meisten Autoren 1865 als Ursprungsjahr.[35][29]
Ein eher selten gebrauchtes Synonym ist Mystomyidae von Edward Drinker Cope, das er im Jahr 1883 aufgrund der abweichenden Molarengestaltung im Vergleich zu den Tenreks einführte.[46] Mystomyidae wiederum geht auf Mystomys zurück, eine Alternativbezeichnung zu Potamogale von John Edward Gray aus dem Jahr 1861. Gray etablierte die neue Gattungsbezeichnung, da er Du Chaillus Erstbeschreibung der Großen Otterspitzmaus als fehlerhaft ansah.[47] Sowohl Mystomys als auch Mystomyidae sind heute nicht anerkannt.[38]
Zur systematischen Stellung

Du Chaillus Reise in das zentrale Afrika in den 1850er Jahren führte zur Entdeckung der Großen Otterspitzmaus. Ihre systematische Stellung war allerdings unklar. So sah Du Chaillu noch Verbindungen zu den Schleichkatzen und damit zu den Raubtieren,[44][48] Gray dagegen verschob sie zu den Glires (der gemeinsamen Gruppe der Nagetiere und Hasenartigen).[47] Beide Einschätzungen fußten auf unzureichendem Material. Erst die Untersuchungen von vollständigen Individuen durch José Vicente Barbosa du Bocage und durch Allman in der Mitte der 1860er Jahre brachten die Erkenntnis, dass es sich hierbei um ein insektenfresserartiges Tier handelt.[49][5] Barbosa du Bocage formulierte eine nähere Verwandtschaft mit den madagassischen Tenreks (Tenrecidae), Allman führte dagegen eine engere Beziehung zu den auf einigen karibischen Inseln verbreiteten Schlitzrüsslern (Solenodontidae) aus. In seiner umfassenden Übersicht A Monograph of the Insectivora aus dem Jahr 1883 bestätigte George Edward Dobson die Verbindungen zu den Tenreks und den Schlitzrüsslern. Er fand dafür Übereinstimmungen in den Schädelmerkmalen und in der Struktur der Mahlzähne.[11]
In der Regel wurde dies im Verlauf des 19. und des 20. Jahrhunderts kaum in Frage gestellt. Die Tenreks galten schon bei der Einführung des Taxons Insectivora durch Thomas Edward Bowdich im Jahr 1821 als Teil dieser.[50] Wilhelm Peters und Ernst Haeckel folgten dem in den 1860er Jahren in ihrer feineren Aufgliederung der Insektenfresser in die Lipotyphla (ohne Blinddarm) und die Menotyphla (mit Blinddarm), wobei die Tenreks und die Schlitzrüssler zu ersteren gehörten.[51][52] Im Jahr 1885 versuchte Theodore Gill eine Neugliederung der Insektenfresser und unterschied diese nach der Ausprägung des Kaumusters der Mahlzähne. In seiner Auffassung bestanden zwei große Formengruppen, einerseits die Zalambdodonta, die eine V-förmige Zahnschmelzleiste auf den Backenzähnen besitzen (von griechisch ζα (za-) für eine Übersteigerung und dem griechischen Buchstaben Λ (Lambda)), andererseits die Dilambdodonta mit einer W-förmigen Leiste (von griechisch δι (di „zwei“) und dem griechischen Buchstaben Λ (Lambda)). Er beachtete dabei auch die Otterspitzmäuse und stellte sie zusammen mit den Tenreks, den Goldmullen und den Schlitzrüsslern zu den Zalambdodonta.[53]
Zweieinhalb Dekaden später kombinierte William K. Gregory in seinem Werk The orders of Mammals die Gliederungen von Haeckel und Gill. Er verwies die Menotyphla aus den Insektenfressern und führte den Rest der Gruppe unter der Bezeichnung Lipotyphla. Die Lipotyphla wiederum untergliederte er in die Zalambdodonta mit den Otterspitzmäusen, den Tenreks, den Schlitzrüsslern und den Goldmulle sowie in die beiden Großgruppen Erinaceomorpha (Igel) und Soricomorpha (Spitzmäuse und Maulwürfe).[54] Percy M. Butler variierte dieses Schema im Jahr 1956. Er unterschied dabei nur zwischen den Erinaceomorpha und den Soricomorpha, die Tenreks (einschließlich der Otterspitzmäuse) und die Goldmulle ordnete er zu den Soricomorpha.[55] Ein völlig andere Auffassung dagegen vertrat Leigh Van Valen. Er gliederte die Zalambdodonta, also die Tenreks, Otterspitzmäuse, Goldmulle und Schlitzrüssler, aus den Insektenfressern aus und verwies sie in die von ihm geschaffene Ordnung der Deltatheridia. Innerhalb dieser Gruppe standen die Zalambdodonta den sogenannten Hyaenodonta gegenüber. Letztere gehören aber in einer eher klassischen Sicht zu den „Creodonta“ („Urraubtiere“), einer großen, aber in sich nicht geschlossenen Gruppe räuberisch lebender Säugetiere.[37] Van Valens Neuordnung wurde jedoch kaum geteilt. Nachfolgende Autoren behielten die zalambdodonten Formen zumeist bei den Insektenfressern und dort bei den Soricomorpha, so etwa Malcolm C. McKenna in den 1970er Jahren[56] und Michael J. Novacek in den 1980er Jahren.[57] McKenna wiederholte diese Auffassung zusammen mit Susan K. Bell noch einmal in seinem Überblickswerk zur Systematik der Säugetiere in den 1990er Jahren, im Unterschied zu vorher hatte er aber die Goldmulle hier aber aus den Soricomorpha ausgeschlossen.[38]
Erst im Übergang vom 20. zum 21. Jahrhundert wurde mittels molekulargenetischen Untersuchungen erkannt, dass die Insektenfresser keine monophyletische Gruppe bilden. Demnach stand der afrikanische Zweig mit den Tenreks und den Goldmullen, zuzüglich der Rüsselspringer (Macroscelidea) und des Erdferkels (Tubulidentata) anderen originär afrikanischen Tieren wie den Elefanten und den Schliefern näher als dem eurasischen Teil der Insektenfresser. Michael J. Stanhope und Kollegen führten daher 1998 die afrikanischen Tiere in dem Taxon Afrotheria zusammen.[58] Nur wenige Jahre später vereinte ein Forscherteam um Peter J. Waddell die afrikanischen Insektenfresser innerhalb der Afrotheria unter der Bezeichnung Afroinsectiphilia.[30] Die Otterspitzmäuse waren in diesen frühen genetischen Untersuchungen noch nicht einbezogen. Die ersten Gensequenzierungen hier erfolgten unter Christophe J. Douady im Jahr 2002 und bestätigten die nahe Verwandtschaft mit den Tenreks.[59] Weitere Analysen unterstützten diese Ansicht.[60][61]
Stammesgeschichte
Gemäß den molekulargenetischen Untersuchungen reicht die Familie der Otterspitzmäuse bis wenigstens in das Untere Eozän zurück.[29] Trotz der langen Zeitspanne sind Fossilfunde äußerst selten.[62] Im Jahr 2015 beschrieb Martin Pickford Schädel- und Gebissreste sowie einzelne Teile des Bewegungsapparates von Namagale, einer sehr urtümlichen Otterspitzmaus, die noch einen deutlich getrennten Paraconus und Metaconus an den Oberkiefermahlzähnen aufweist. Wie bei den anderen Vertretern der Familie ist der Eckzahn zweiwurzelig und der hinterste Mahlzahn verkleinert. Die Funde stammen von der Fundstelle Eocliff im Sperrgebiet von Namibia. Es handelt sich hierbei um eine Travertinlagerstätte, die sich im Oberen Eozän an der Austrittstelle einer kalkhaltigen Quelle gebildet hatte. Namagale wäre damit der älteste bekannte Angehörige der Otterspitzmäuse.[35][63]
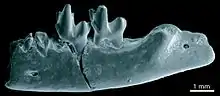
Weitere eindeutige Fossilfunde sind nicht bekannt. Aus dem Fayyum-Gebiet in Ägypten wurden einzelne Formen wie Dilambdogale, Widanelfarasia, Jawharia und Qatranilestes bekannt, die in der Regel nur über Gebissreste überliefert sind. Für sie ist eine nähere Verwandtschaft mit den Afrosoricida anzunehmen, was sich unter anderem in dem zalambdodonten Kauflächenmuster der hinteren Oberkiefermolaren ausdrückt. Allerdings wies der vorderste Mahlzahn ein weitgehend dilambdodontes Muster auf, was an die eigentlichen Insektenfresser erinnert. Die Datierung der Funde aus dem Fayyum reicht vom Oberen Eozän bis zum Unteren Oligozän.[64][65][66] Jünger sind Funde aus dem östlichen Afrika, die Protenrec und Erythrozootes zugewiesen werden und in das Untere Miozän gehören. Das Fundmaterial setzt sich aus verschiedenen Schädel- und Zahnresten zusammen. In ihrem weniger entwickelten zalambdodonten Molarenmuster weisen sie Übereinstimmungen mit den Otterspitzmäusen, speziell mit der Großen Otterspitzmaus auf, gelegentlich wurden sie auch in deren systematische Nähe gestellt.[37] Die meisten Autoren sehen aber aufgrund des einwurzligen Eckzahns und einiger Schädelmerkmale eine nähere Stellung zu den Tenreks.[67][68][62] Allerdings wurde auch ähnlich zu den Funden aus Fayyum eine sehr basale Stellung innerhalb der Afrosoricida vorgeschlagen.[65][66]
Bedrohung und Schutz
Bedrohungen für die einzelnen Bestände der Otterspitzmäuse stellen der Verlust des Lebensraums durch Waldrodungen und Ausweitung landwirtschaftlicher Nutzflächen sowie die Verminderung der Wasserqualität durch die damit verbundene Erosion, aber auch durch Bautätigkeit dar. Dazu kommt noch die Jagd auf ihr Fell und die Tatsache, dass sich Tiere manchmal in Fischernetzen verfangen und ertrinken. Die IUCN listet eine Art, die Zwerg-Otterspitzmaus, als „potenziell gefährdet“ (near threatened). Diese bewohnt ein kleines Gebiet im westlichen Afrika, in welchem intensiv nach Eisenerzen geschürft wird, was den Lebensraum immer weiter einschränkt. Die anderen beiden Arten gelten hingegen als nicht unmittelbar bedroht. Für alle drei Vertreter der Otterspitzmäuse merkt die IUCN aber auch eine potentiell höhere Gefährdungskategorie an, was die Organisation mit dem Rückgang der Population begründet. Alle drei Arten sind in Schutzgebieten präsent.[69][70][71]
Literatur
- Ara Monadjem: Potamogalidae (Otter-shrews). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos. Lynx Edicions, Barcelona 2018, S. 174–179 ISBN 978-84-16728-08-4
- Ronald M. Nowak: Walker's Mammals of the World. The Johns Hopkins University Press, Baltimore 1999, ISBN 0-8018-5789-9
- Peter Vogel: Subfamily Potamogalinae Otter Shrew. In: Jonathan Kingdon, David Happold, Michael Hoffmann, Thomas Butynski, Meredith Happold und Jan Kalina (Hrsg.): Mammals of Africa Volume I. Introductory Chapters and Afrotheria. Bloomsbury, London, 2013, S. 216–222
Einzelnachweise
- W. N. Verheyen: Recherches anatomiques sur Micropotamogale ruwenzorii. 1. La morphologie externe, les viscères et l’organe génital mâle. Bulletins de la Société Royale de Zoologie d’Anvers 21, 1961, S. 1–16
- Hans-Jürg Kuhn: Zur Kenntnis von Micropotamogale lamottei. Zeitschrift für Säugetierkunde 29, 1964, S. 152–173
- Peter Vogel: Subfamily Potamogalinae Otter Shrew. In: Jonathan Kingdon, David Happold, Michael Hoffmann, Thomas Butynski, Meredith Happold und Jan Kalina (Hrsg.): Mammals of Africa Volume I. Introductory Chapters and Afrotheria. Bloomsbury, London, 2013, S. 216–222
- Ara Monadjem: Potamogalidae (Otter-shrews). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos. Lynx Edicions, Barcelona 2018, S. 174–179 ISBN 978-84-16728-08-4
- George J. Allman: On the characters and affinities of Potamogale, a genus of insectivorous mammals. Transactions of the Zoological Society of London 6, 1866, S. 1–16 ()
- Hans-Jürg Kuhn: An Adult Female Micropotamogale lamottei. Journal of Mammalogy 52 (2), 1971, S. 477–478
- M. R. Sanchez-Villagra und R. J. Asher: Cranio-sensory adaptations in small faunivorous semiaquatic mammals, with special reference to olfaction and the trigeminal system. Mammalia 66 (1), 2002, S. 93–109
- Ch. Guth, Henri Heim de Balsac und M. Lamotte: Recherches sur la morphologie de Micropotamogale lamottei et l’evolution des Potamogalinae. I. Ecologie, denture, anatomie crânienne. Mammalia 23, 1959, S. 423–447
- Robert J. Asher und Marcelo R. Sánchez-Villagra: Locking Yourself Out: Diversity Among Dentally Zalambdodont Therian Mammals. Journal of Mammalian Evolution. 12 (1/2), 2005, S. 265–282
- Ch. Guth, Henri Heim de Balsac und M. Lamotte: Recherches sur la morphologie de Micropotamogale lamottei et l’evolution des Potamogalinae. II. Rachis, viscéris, position systématique. Mammalia 24, 1960, S. 190–216
- George Edward Dobson: A Monograph of the Insectivora, systematic and anatomical. Part II. London, 1883, S. 67–86 ()
- Wilhelm Leche: Zur Entwicklungsgeschichte des Zahnsystems der Säugetiere, zugleich ein Beitrag zur Stammesgeschichte dieser Tiergruppe. Zweiter Teil: Phylogenie. Zweites Heft: Familien der Centetidae, Solenodontidae und Chrysochloridae. Zoologica 20, 1906/1908, S. 1–157 ()
- W. N. Verheyen: Recherches anatomiques sur Micropotamogale ruwenzorii. 4. Observations ostéologiques et considérations générales. Bulletins de la Société Royale de Zoologie d’Anvers 22, 1961, S. 1–7
- Justine A. Salton und Eric J. Sargis: Evolutionary morphology of the Tenrecoidea (Mammalia) carpal complex. Biological Journal of the Linnean Society, 93, 2008, S. 267–288
- Justine A. Salton und Eric J. Sargis: Evolutionary Morphology of the Tenrecoidea (Mammalia) Forelimb Skeleton. In: E.J. Sargis und M. Dagosto (Hrsg.): Mammalian Evolutionary Morphology: A Tribute to Frederick S. Szalay, Springer Science, 2008, S. 51–71
- Justine A. Salton und Frederick S. Szalay: The Tarsal Complex of Afro-Malagasy Tenrecoidea: A Search for Phylogenetically Meaningful Characters. Journal of Mammalian Evolution 11 (2), 2004, S. 73–104
- Justine A. Salton und Eric J. Sargis: Evolutionary Morphology of the Tenrecoidea (Mammalia) Hindlimb Skeleton. Journal of Morphology 270, 2009, S. 367–387
- U. Rahm: Beobachtungen an der ersten in Gefangenschaft gehaltenen Mesopotamogalen ruwenzorii (Mammalia-Insectivora). Revue Suisse de Zoologie 68 (4), 1961, S. 73–90
- A. M. Carter, T. N. Blankenship, A. C. Enders und P. Vogel: The Fetal Membranes of the Otter Shrews and a Synapomorphy for Afrotheria. Placenta 27, 2006, S. 258–268
- H. Stephan und H.-J. Kuhn: The brain of Micropotamogale lamottei Heim de Balsac, 1954. Zeitschrift für Säugetierkunde 47, 1982, S. 129–142
- H. Stephan, Kabongo Ka Mubalamata und M. Stephan: The Brain of Micropotamogale ruwenzorii (De Witte and Frechkop, 1955). Zeitschrift für Säugetierkunde 51, 1986, S. 193–204
- Jason A. Kaufman, Gregory H. Turner, Patricia A. Holroyd, Francesco Rovero und Ari Grossman: Brain Volume of the Newly-Discovered Species Rhynchocyon udzungwensis(Mammalia: Afrotheria: Macroscelidea): Implications for Encephalization in Sengis. PlosOne 8 (3), 2013, S. e58667 doi:10.1371/journal.pone.0058667
- Gérard Dubost: Quelques renseignements biologiques sur Potamogale velox. Biologia Gabonica 1, 1965, S. 257–272
- Martin E. Nicoll: The biology of the Giant otter-shrew Potamogale velox. National Geographic Society Research Report 21, 1985, S. 331–337
- Peter Vogel: Contribution a l’écologie et a la zoogéographie de Micropotamogale lamottei (Mammalia, Tenrecidae). Revue d‘Ecologie (Terre Vie) 38, 1983, S. 37–49
- Peter Vogel: Highlights and disappointments during 40 years of research on otter-shrews. Afrotherian Conservation 10, 2014, S. 9–11
- A. C. Enders, A. M. Carter, H. Künzle und P. Vogel: Structure of the Ovaries of the Nimba Otter Shrew, Micropotamogale lamottei, and the Madagascar Hedgehog Tenrec, Echinops telfairi. Cells Tissues Organs 179, 2005, S. 179–191
- Steven Heritage, Erik R. Seiffert und Matthew R. Borths: Recommended fossil calibrators for time-scaled molecular phylogenies of Afrotheria. Afrotherian Conservation 17, 2021, S. 9–13
- Kathryn M. Everson, Voahangy Soarimalala, Steven M. Goodman und Link E. Olson: Multiple loci and complete taxonomic sampling resolve the phylogeny and biogeographic history of tenrecs (Mammalia: Tenrecidae) and reveal higher speciation rates in Madagascar’s humid forests. Systematic Biology 65 (5), 2016, S. 890–909 doi: 10.1093/sysbio/syw034
- Peter J. Waddell, Hirohisa Kishino und Rissa Ota: A Phylogenetic Foundation for Comparative Mammalian Genomics. Genome Informatics 12, 2001, S. 141–154
- Matjaž Kuntner, Laura J. May-Collado und Ingi Agnarsson: Phylogeny and conservation priorities of afrotherian mammals (Afrotheria, Mammalia). Zoologica Scripta 40 (1), 2011, S. 1–15
- Robert W. Meredith, Jan E. Janečka, John Gatesy, Oliver A. Ryder, Colleen A. Fisher, Emma C. Teeling, Alisha Goodbla, Eduardo Eizirik, Taiz L. L. Simão, Tanja Stadler, Daniel L. Rabosky, Rodney L. Honeycutt, John J. Flynn, Colleen M. Ingram, Cynthia Steiner, Tiffani L. Williams, Terence J. Robinson, Angela Burk-Herrick, Michael Westerman, Nadia A. Ayoub, Mark S. Springer und William J. Murphy: Impacts of the Cretaceous Terrestrial Revolution and KPg Extinction on Mammal Diversification. Science 334, 2011, S. 521–524
- Erik R Seiffert: A new estimate of afrotherian phylogeny based on simultaneous analysis of genomic, morphological, and fossil evidence. BMC Evolutionary Biology 7, 2007, S. 224 doi:10.1186/1471-2148-7-224
- Henri Heim de Balsac: Morphologie divergente des Potamogalinae (Mammifères Insectkivores) en milieu aquatique. Comptes rendus hebdomadaires des séances de l’Académie des sciences 242, 1956, S. 2257–2258 ()
- Martin Pickford: Late Eocene Potamogalidae and Tenrecidae (Mammalia) from the Sperrgebiet, Namibia. Communications of the Geological Survey of Namibia 16, 2015, S. 114–152
- George Gaylord Simpson: The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History 85, 1945, S. 1–350 (S. 48–49 und 175–179)
- Leigh van Valen: New Paleocene Insectivores and Insectivore classification. Bulletin of the American Museum of Natural History 135, 1967, S. 217–284
- Malcolm C. McKenna und Susan K. Bell: Classification of mammals above the species level. Columbia University Press, New York, 1997, S. 1–631 (S. 293–295)
- Don E. Wilson und DeeAnn M. Reeder: Mammal Species of the World. A taxonomic and geographic Reference. Johns Hopkins University Press, 2005 ()
- Serge Frechkop: A propos de nouvelles espèces de Potamogalines. Mammalia 21, 1957, S. 226–234
- Robert J. Asher: A morphological basis for assessing the phylogeny of the „Tenrecoidea“ (Mammalia, Lipotyphla). Cladistics 15, 1999, S. 231–252
- Link E. Olson und Steven M. Goodman: Phylogeny and biogeography of tenrecs. In: Steven M. Goodman und Jonathan P. Benstead (Hrsg.): The natural history of Madagascar. University of Chicago Press, 2003, S. 1235–1242
- Robert J. Asher und Robert Hofreiter: Tenrec Phylogeny and the Noninvasive Extraction of Nuclear DNA. Systematic Biology 55 (2), 2006, S. 181–194
- Paul Belloni Du Chaillu: Descriptions of new species of mammals discovered in western equatorial Africa. Proceedings of the Boston Society of Natural History 7, 1860, S. 296–304 und 358–367 (S. 361–363) ()
- George J. Allman: [ohne Titel]. Proceedings of Zoological Society of London, 1865, S. 467 ()
- Edward Drinker Cope: On the Mutual Relations of the Bunotherian Mammalia. Proceedings of the Academy of Natural Sciences of Philadelphia 35, 1883, S. 77–83 ()
- John Edward Gray: Zoological notes on perusing M. Du Chaillu's "Adventures in Equatorial Afrika". Annals and Magasin of Natural History 8, 1861, S. 60–65 ()
- Paul Belloni Du Chaillu: Explorations and adventures in Equatorial Africa. London, 1861, S. 1–479 (S. 422) ()
- José Vicente Barbosa du Bocage: Sur quelques Mammifères rares et peu connus, d'Afrique occidentale, qui se trouvent au Muséum de Lisbonne. Proceedings of the Zoological Society of London, 1865, S. 401–404 ()
- Thomas Edward Bowdich: An analysis of the natural classifications of Mammalia for the use of students and travellers. Paris, 1821, S. 1–115 (S. 31–33) ()
- Wilhelm Peters: Über die Säugethirgattung Selenodon. Abhandlungen der Königlichen Akademie der Wissenschaften zu Berlin 1863, S. 1–22 ()
- Ernst Haeckel: Generelle Morphologie der Organismen. Band 2. Berlin, 1866, S. 1–462 (S. CLX) ()
- Theodore Gill: Insectivora. In: John Sterling Kingsley (Hrsg.): Standard natural history. Volume 5. Boston, 1885, S. 1–535 (S. 134–158) ()
- William King Gregory: The orders of mammals. Bulletin of the American Museum of Natural History 27, 1910, S. 1–524 (S. 464–465)
- Percy M. Butler: The skull of Ictops and the classification of the Insectivora. Proceedings of the Zoological Society of London 126, 1956, S. 453–481
- Malcolm C. McKenna: Toward a phylogenetic classification of the Mammalia. In W. Patrick Luckett und Frederick S. Szalay (Hrsg.): Phylogeny of the primates: a multidisciplinary approach. New York, London, 1975, S. 21–46
- Michael J. Novacek: The skull of leptictid insectivorans and the higher-level classification of eutherian mammals. Bulletin of the American Museum of Natural History 183, 1986, S. 1–111
- Michael J. Stanhope, Victor G. Waddell, Ole Madsen, Wilfried de Jong, S. Blair Hedges, Gregory C. Cleven, Diana Kao und Mark S. Springer: Molecular evidence for multiple origins of Insectivora and for a new order of endemic African insectivore mammals. PNAS 95, 1998, S. 9967–9972 ()
- Christophe J. Douady, Francois Catzeflis, Diana J. Kao, Mark S. Springer und Michael J. Stanhope: Molecular Evidence for the Monophyly of Tenrecidae (Mammalia) and the Timing of the Colonization of Madagascar by Malagasy Tenrecs. Molecular Phylogenetics and Evolution 22 (3), 2002, S. 357–363
- Céline Poux, Ole Madesen, Elisabeth Marquard, David R. Vieites, Wilfried W. de Jong und Miguel Vences: Asynchronous Colonization of Madagascar by the Four Endemic Clades of Primates, Tenrecs, Carnivores, and Rodents as Inferred from Nuclear Genes. Systematic Biology 54 (5), 2005, S. 719–730
- Céline Poux, Ole Madesen, Julian Glos, Wilfried W. de Jong und Miguel Vences: Molecular phylogenetic and divergence times of Malagasy tenrecs: Influence of data partitioning and taxon sampling on dating analyses. BMC Evolutionary Biology, 2008, S. 102
- Robert J. Asher: Tenrecoidea. In: Lars Werdelin und William Joseph Sanders (Hrsg.): Cenozoic Mammals of Africa. University of California Press, Berkeley, London, New York, 2010, S. 99–106
- Robert J. Asher: Recent additions to the fossil record of tenrecs and golden moles. Afrotherian Conservation 15, 2019, S. 4–13
- Erik R. Seiffert und Elwyn L. Simons: Widanelfarasia, a diminutive placental from the late Eocene of Egypt. PNAS 97 (6), 2000, S. 2646–2651
- Erik R. Seiffert, Elwyn L. Simons, Timothy M. Ryan, Thomas M. Bown und Yousry Attia: New remains of Eocene and Oligocene Afrosoricida (Afrotheria) from Egypt, with implications for the origin(s) of afrosoricid zalambdodonty. Journal of Vertebrate Paleontology 27 (4), 2007, S. 963–972
- Erik R. Seiffert: The oldest and youngest records of afrosoricid placentals from the Fayum Depression of northern Egypt. Acta Palaeontologica Polonica 55 (4), 2010, S. 599–616
- Percy M. Butler: Insectivora and Chiroptera. In: V. J. Maglio und H. B. S. Cooke (Hrsg.): Evolution of African Mammals. Harvard University Press, 1978, S. 56–68
- Percy M. Butler: Macroscelidea, Insectivora and Chiroptera from the Miocene of East Africa. Palaeovertebrata 14 (3), 1984, S. 117–200
- Peter J. Stephenson, Steven M. Goodman und Voahangy Soarimalala: Potamogale velox. The IUCN Red List of Threatened Species 2016. e.T18095A97203526 (); zuletzt abgerufen am 18. Oktober 2017
- Peter J. Stephenson: Micropotamogale ruwenzorii. The IUCN Red List of Threatened Species 2016. e.T13394A21287768 (); zuletzt abgerufen am 18. Oktober 2017
- Peter J. Stephenson: Micropotamogale lamottei. The IUCN Red List of Threatened Species 2016. e.T13393A21287657 (); zuletzt abgerufen am 18. Oktober 2017
Weblinks
- Potamogale velox in der Roten Liste gefährdeter Arten der IUCN 2017. Eingestellt von: J. Stephenson, Steven M. Goodman und Voahangy Soarimalala, 2015. Abgerufen am 18. Oktober 2017.
- Micropotamogale ruwenzorii in der Roten Liste gefährdeter Arten der IUCN 2017. Eingestellt von: P. J. Stephenson, 2015. Abgerufen am 18. Oktober 2017.
- Micropotamogale lamottei in der Roten Liste gefährdeter Arten der IUCN 2017. Eingestellt von: P. J. Stephenson, 2015. Abgerufen am 18. Oktobero0 2017.