Gürteltiere
Die Gürteltiere (Dasypoda) bilden die einzige überlebende Säugetiergruppe der Gepanzerten Nebengelenktiere (Cingulata). Sie werden heute in zwei Familien mit insgesamt rund 20 gegenwärtig vorkommenden Arten unterschieden, darüber hinaus sind zahlreiche weitere, ausgestorbene Vertreter bekannt. Die Gruppe ist über den südamerikanischen Kontinent und die südöstlichen Teile Nordamerikas verbreitet; ein Großteil der heutigen Arten konzentriert sich im zentralen und nördlichen Südamerika.
| Gürteltiere | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Neunbinden-Gürteltier (Dasypus novemcinctus) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Dasypoda | ||||||||||||
| Quenstedt, 1885 (Hoernes, 1886) |
Als einzige bekannte Säugetiergruppe der heutigen Zeit verfügen die Gürteltiere über eine knöcherne äußere Panzerbildung. Die mit Hornschuppen gepanzerten Schuppentiere galten früher irrtümlich als nahe mit ihnen verwandt. Gürteltiere treten weitgehend einzelgängerisch auf und leben in unterschiedlichem Maße unterirdisch in Erdbauen, wobei sie an diese grabende Lebensweise durch mehrere Skelettmodifikationen angepasst sind. Als Hauptnahrung werden verschiedenste Insekten konsumiert, einige Arten verzehren aber auch pflanzliche Nahrung oder vertilgen kleinere Wirbeltiere. Die Lebensgewohnheiten der Gürteltiere sind nicht in allen Fällen gut untersucht, vor allem was die Fortpflanzung betrifft.
Das Fleisch wird von Menschen teilweise als Nahrungsressource genutzt, manche Skelettteile werden lokal auch als handwerkliche Rohstoffe verwendet. Einige Arten gelten aufgrund ihrer bodenwühlenden Aktivitäten als Landplage. In Europa bekannt wurden die Gürteltiere erstmals am Anfang des 16. Jahrhunderts, die heute anerkannte Erstbeschreibung der Gruppe der Dasypoda stammt aus dem Jahr 1885/1886.
Die über Fossilfunde belegte stammesgeschichtliche Entwicklung kann bis in das Paläozän vor mehr als 56 Millionen Jahren zurückverfolgt werden. Die gegenwärtig bestehenden Arten traten jedoch erst relativ spät in Erscheinung. Die nächsten Verwandten der Gürteltiere sind die Ameisenbären und Faultiere. Alle drei Gruppen weisen besondere Skelettbildungen an der Wirbelsäule auf, die sie gemeinsam die Überordnung der Nebengelenktiere (Xenarthra) formen lassen, einer der vier großen Hauptlinien der Höheren Säugetiere.
Körperbau
Habitus
.jpg.webp)
Gürteltiere sind überwiegend kleine bis selten mittelgroße Säugetiere mit einer Kopf-Rumpf-Länge von 11 bis 100 cm. Hinzu kommt ein Schwanz, der mit Ausnahme der Kugelgürteltiere (Tolypeutes) und der Gürtelmulle häufig etwa die Hälfte der Körperlänge ausmacht, manchmal jedoch deutlich länger ist. Das Gewicht variiert ähnlich der Körpergröße sehr stark und reicht von 100 g beim Gürtelmull (Chlamyphorus truncatus) bis zu 45 kg beim Riesengürteltier (Priodontes giganteus).[1][2] Ausgestorbene Gürteltiere konnten allerdings auch erheblich größer werden und erreichten mit Macroeuphractus, das schätzungsweise zwischen 90 und 120 kg wog, das Gewicht heutiger kleinerer Hausschweine.[3][4] Ein Großteil der bekannten Arten verblieb aber in der Größenvariabilität der rezenten Vertreter.[5]
Der Kopf hat eine schmale, spitz zulaufende Form, die Ohren stehen mausartig nach oben ab, die Augen sind sehr klein. Bei einigen Arten ist die Schnauze röhrenartig verlängert. Auf der Oberseite ist der Kopf mit Knochenplättchen besetzt, die einen Kopfpanzer formen. Charakteristisch ist der meist kräftig gewölbte Rückenpanzer, der fast die gesamte Körperoberfläche der Tiere bedeckt. An Vorder- und Hinterleib ist er häufig zu starren Rückenschilden verwachsen (Schulter- und Beckenschild, auch Buckler genannt), dazwischen befindet sich eine variierende Anzahl an zur Bauchseite offenen, querverlaufenden Ringen, die Gürtel oder Bänder, nach denen die Gruppe ihren deutschen Trivialnamen erhielt. Verbindungen mit überlappenden Hautfalten gewährleisten eine hohe Flexibilität des Panzers im Bereich der Bänder. Der spitz endende Schwanz ist bei den meisten Arten ebenfalls von Knochenringen umgeben. Auf dem Bauch dagegen findet sich keine Panzerung, manchmal aber einzelne Knochenplättchen. Eine Körperbehaarung ist nicht bei allen Arten ausgebildet, sofern sie vorhanden ist, sprießt sie in borstigen Haaren aus den Rückenschildchen oder an den Körperseiten und ist wollig dicht am Bauch.[6][1][2]
Die kurzen Beine haben hinten immer fünf, vorne jedoch vier bis fünf, selten auch drei Zehen, die stark gebogene Krallen tragen, wobei jene der Hinterfüße bei allen Arten deutlich kleiner als jene der vorderen sind. Dabei halten die Krallen an den mittleren Zehen der Vorderfüße des Riesengürteltiers mit bis zu 20 cm Länge den Rekord der größten Krallen im gesamten heutigen Tierreich.[6][1][2]
Schädel- und Gebissmerkmale
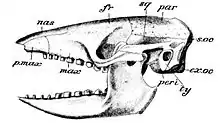
Die verschiedenen Gürteltierarten besitzen variierende Schädelformen, die zumeist die Ernährungsweise widerspiegeln. Arten, die sich überwiegend von Insekten ernähren, besitzen meist einen leichter gebauten Schädel und Unterkiefer mit geringer entwickelten Muskelansatzstellen. Jene aber, die weitgehend als Allesfresser leben und zum Teil auch Pflanzen verzehren, sind durch massivere Schädel mit deutlich entwickelter Kaumuskulatur charakterisiert.[7] Neben dem generellen Aufbau des Schädels der Säugetiere weisen die Gürteltiere zwei anatomische Besonderheiten auf. Im vorderen Bereich der Nasenhöhle tritt eine als Septomaxilla (Os nariale) bezeichnete Knochenbildung auf, die sonst nur bei stammesgeschichtlich älteren Säugetieren und Reptilien ausgebildet ist. Die Funktion dieses Knochens bei den Gürteltieren ist weitgehend unbekannt, wahrscheinlich dient er zum Verschließen der Nasenlöcher während des Grabens, damit kein Staub eingeatmet wird. Bei zahlreichen, aber nicht allen Gürteltierarten ist am Gehörgang anstatt einer Paukenblase ein tympanischer Ring ausgebildet.[8][1][2]

Der Unterkiefer ist bei vielen Arten zumeist lang, schmal und am Knochenkörper niedrig gebaut. Auffallend ist die nicht fest verknöcherte Symphyse, die die beiden Unterkieferhälften im vorderen Bereich verbindet.[9] Im Zahnbau zeichnen sich ebenfalls die Unterschiede in der Ernährungsweise ab mit kleinen Zähnen bei den Insektenfressern und großen bei den Allesfressern. Zudem weichen die Zähne von denen der anderen Säugetiere ab: Sie sind homodont gestaltet, Schneide- oder Eckzähne beziehungsweise Prämolaren werden nicht ausgebildet. Das gesamte Gebiss besteht aus stiftartig einfachen, an Molaren erinnernde Zahnbildungen. Diese weisen dabei keinerlei Zahnschmelz auf – nur die Langnasengürteltiere (Dasypus) und einige ausgestorbene Gattungen besitzen eine sehr dünne Zahnschmelzschicht, die sehr schnell abgekaut wird – und bestehen fast nur aus Zahnbein, hauptsächlich Orthodentin, dem härtesten Bestandteil des Zahnbeins. Einige Arten besitzen anstelle des Zahnschmelzes eine sehr dünne äußere Schicht an Zahnzement, was etwa beim Riesengürteltier oder beim Zwerggürteltier (Zaedyus pichiy) der Fall ist. Die Anzahl der Zähne variiert von Art zu Art und häufig auch innerhalb der Arten und reicht insgesamt von 7 bis 25 je Kieferbogen. Die Zähne wachsen in der Regel ein Leben lang, lediglich bei den Langnasengürteltieren ist ein Zahnwechsel vom Milch- zum Dauergebiss nachgewiesen. Dieser kann mitunter sehr spät erfolgen, teilweise erst im ausgewachsenen Alter und variiert offensichtlich individuell.[8][10][1][2]
Panzerung
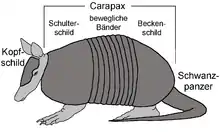

Die Ausbildung eines äußeren, knöchernen Körperpanzers, der den Kopf (Kopfschild) sowie den Rücken (Carapax) und mit Ausnahme der Nacktschwanzgürteltiere (Cabassous) auch den Schwanz (Schwanzpanzer) sowie Teile der Beine bedeckt, ist einzigartig unter den Säugetieren und gibt den Gürteltieren und ihren ausgestorbenen Verwandten dadurch eine besondere Stellung innerhalb dieser Tierklasse. Er besteht aus Knochenplättchen, sogenannte Osteoderme, die in der Haut gebildet werden. Sie sind sowohl am Rückenpanzer als auch am Schwanz in Reihen angeordnet. Je nach Art variiert die Anzahl der panzerbildenden Osteoderme. Bei den Kugelgürteltieren liegt sie bei über 660, beim Zwerggürteltier bei rund 950 und bei den Borstengürteltieren (Chaetophractus) bei über 1040.[11] Die Osteoderme werden von Hornschildchen überzogen, die in zwei unterschiedlichen Weisen angeordnet sind: Einerseits bedeckt ein Hornschildchen mehrere, teilweise bis zu vier benachbarte Osteoderme gleichzeitig, andererseits nur ein oder maximal zwei. Erstere Variante wird als eher urtümlich für die Gürteltiere angesehen und ist fast ausschließlich nur bei den Langnasengürteltieren ausgebildet, letztere trifft auf die meisten anderen Arten zu.[12][6][1][2]
Die Osteoderme der starren Teile des Rückenpanzers besitzen häufig eine quadratische, vier- bis vieleckige Form. An den beweglichen Bändern sind sie dagegen zumeist länglich-rechteckig gestaltet und verfügen am vorderen Ende über eine Gelenkfläche, die sie mit dem entsprechenden Element der benachbarten Reihe verbindet. Ein einzelnes Osteoderm eines beweglichen Bandes unterlagert dadurch mit der Gelenkfläche sein Gegenstück der vorhergehenden Reihe und überdeckt im hinteren Abschnitt die jeweilige Gelenkfläche des nachfolgenden Knochenplättchens. Die Oberfläche der Osteoderme ist in der Regel ornamentiert. Dabei umfasst diese Oberflächenzeichnung in der Regel eine größere, zentrale Musterung, um die herum kleinere Muster konzentrisch oder seitlich angeordnet sind. Form und Oberflächengestaltung der Knochenplättchen variieren zwischen den Arten, beide Merkmale haben somit taxonomischen Wert. Die Hornschildchen sind zumeist rundlich in ihrer Gestalt und überlagern die Knochenplättchen. Bei den Langnasengürteltieren, deren Hornschildchen mehrere Osteoderme bedecken, entsteht so am Schulter- und Beckenschild mitunter ein irreguläres Muster, bei den anderen Gürteltieren hingegen paust sich der linear angeordnete Aufbau des Panzers durch.[13]
Die Knochenplättchen sind im Querschnitt mehrlagig aufgebaut, mit einer festen Knochenschicht oben und unten sowie weicherem Material innen, in dem sich Hohlräume für Schweiß- und Talgdrüsen sowie Knochenmark und bei behaarten Arten auch für Haarfollikel befinden. Generell haben die einzelnen Hohlräume je nach Funktion einen eigenen Charakter und können so auch bei fossilen Formen relativ sicher zugewiesen werden. Solche Haarfollikel sind zumeist langgestreckt und gruppieren sich am Rand des Osteoderms, Drüsen haben häufig eine kugelige Form und sitzen in der Mitte, während die für Knochenmark in der Regel keine Öffnung zur Oberfläche aufweisen.[14][15][16][17] Die knöcherne Substanz setzt sich aus hartem, stark mineralisiertem Material und kollagenreichen, faserigen Strukturen (Sharpeysche Fasern) zusammen, die den einzelnen Knochenplättchen eine hohe Elastizität geben.[18] Im Innern wird der Panzer durch meist 11 Paare an sehr breiten Rippen abgestützt.[7]
Die Anlage des Panzers beginnt bereits in der Fetalphase im Mutterleib. Neugeborene besitzen allerdings einen weichen, ledrigen Panzer, der nach und nach durch Verknöcherung der Lederhaut aushärtet. Dabei bilden sich die typischen, je nach Art unterschiedlich geformten, mehreckigen bis runden Knochenplättchen, auf denen die Hornplättchen sitzen und sich zu den festen Panzerteilen und Gürteln sortieren. Die Aushärtung des Panzers erfolgt aber je nach Art unterschiedlich während des Jugendwachstums, was als Anpassung an die jeweiligen Umweltbedingungen interpretiert wird.[19]
Ob sich der Panzer als Schutz vor Fressfeinden bildete, ist wissenschaftlich nicht vollständig geklärt. Allerdings schützt er gut vor der dornigen Vegetation trockener Landschaften, in denen zahlreiche Gürteltierarten leben. Gleichzeitig vermindert er den Befall mit äußeren Parasiten.[7][20]
Körperskelett
.jpg.webp)
Spezielle, von anderen Säugetieren abweichende Skelettmerkmale finden sich vor allem im Bereich der Wirbelsäule. Die sogenannten xenarthrischen Gelenke (Nebengelenke), die der Überordnung Xenarthra ihren Namen gaben, sind vor allem an den hinteren Brustwirbeln und den Lendenwirbeln ausgebildet. Diese zusätzlichen Gelenke finden sich an den seitlichen Fortsätzen der Wirbel und verbinden den vorhergehenden mit dem nachfolgenden (genauer den Processus accessorius mit dem Processus transversus und dem Processus mammillaris), allerdings ist ihre Funktion weitgehend unklar. Auch die Gelenkflächen der Schwanzwirbel sind kräftig ausgebildet und greifen tief ineinander, was die Gürteltiere befähigt, den Schwanz beim Aufrichten auf die Hinterbeine als Stütze zu benutzen. Im Bereich der Halswirbelsäule gibt es bei einigen Arten Verwachsungen der hinteren Hals- mit den vorderen Brustwirbeln, was als postcervicaler Knochen bezeichnet wird und höchstwahrscheinlich mit der grabenden Lebensweise der Tiere in Verbindung steht. Darüber hinaus besitzen Gürteltiere nur durchschnittlich 11 Brustwirbel und meist zwischen 3 und 4 Lendenwirbel, was deutlich geringer ist als bei ihren nächsten Verwandten, den Ameisenbären und Faultieren und vor allem im Fall der geringeren Anzahl an Lendenwirbeln dem Rücken mehr Stabilität beim Aufrichten verleiht. Eine derartige Reduktion der Anzahl aller Rückenwirbel im Vergleich zu nahe verwandten Gruppen scheint zudem typisch für panzertragende Wirbeltiere zu sein, da ähnliches bei Schildkröten und einigen ausgestorbenen Parareptilien mit Panzerbildung ebenfalls nachgewiesen ist.[21][22]
Auch der sonstige Bewegungsapparat zeigt einige Besonderheiten. Die kräftigen Vorderbeine mit den großen Klauen sind eine hervorragende Anpassung an eine grabende Lebensweise. Im Knochenbau zeigt dies vor allem die Ulna, deren proximaler (= rumpfnaher) Gelenkfortsatz (das Olecranon) stark ausgeprägt ist; seine Länge macht zwischen 37 (Riesengürteltier) und 54 % (Gürtelmull) der gesamten Knochenlänge aus.[23] Der prozentuale Anteil des Olecranons an der Ulna ist umso größer, je stärker die einzelnen Arten an eine grabende Lebensweise angepasst sind. Proximales wie distales (= rumpffernes) Ende des Oberarmknochens zeigen bei grabenden Arten ebenfalls Verstärkungen, die dem Ansatz entsprechender Muskulatur dienen.[24]
Die Hintergliedmaßen sind dagegen nicht übermäßig kräftig ausgebildet, was daran liegt, dass diese nicht zum Graben verwendet werden, sondern vornehmlich der Fortbewegung bei der Futtersuche dienen.[25][23] Der Große Rollhügel (Trochanter major) des Oberschenkelknochens befindet sich etwa auf der Höhe des Oberschenkelkopfes, was die Streckung des Beins im Hüftgelenk begünstigt. Dies geschieht unter anderem in den Bauen beim Festkrallen in den Boden, wenn ein Tier von einem Fressfeind attackiert wird. Neben dem Großen und dem Kleinen Rollhügel (Trochanter minor) besitzt der Oberschenkelknochen typischerweise einen Dritten Rollhügel (Trochanter tertius); letzterer ist bei Säugetieren nicht regelhaft zu finden, stellt aber ein charakteristisches Merkmal der Nebengelenktiere dar. Bei Gürteltierarten mit höherer Körpermasse liegt der Trochanter tertius weiter distal (zum Kniegelenk hin) als bei solchen mit geringerer.[26]
Innere Organe und Sinnesleistung
Die Nahrungsaufnahme erfolgt mit Hilfe der langen, weit ausstreckbaren Zunge, die zudem bei den meisten untersuchten Gürteltierarten eine durch pilz-, kreis- oder fadenförmige Papillen aufgeraute Oberfläche besitzt.[27] Die Verdauung wird dabei durch einen sialinsäurehaltigen Speichel unterstützt, der in drei unterschiedlichen Hauptgruppen von Speicheldrüsen gebildet wird (Ohr-, Unterzungen- und Unterkieferspeicheldrüsen).[28] Ein Großteil der Nahrungszersetzung findet im Magen statt, dessen Pylorus stark von Muskeln durchsetzt ist. Der Magen ist zumeist sackartig einfach gestaltet,[29] bei einigen Gürteltierarten, etwa dem Südlichen Siebenbinden-Gürteltier, wirkt er aber durch eine Einziehung fast zweilappig. Überdies ist er recht groß und kann beim Neunbinden-Gürteltier rund 150 cm³ umfassen.[7] Die Milz erreicht zwischen 0,21 und 0,38 % des Körpergewichtes und hat über die gesamte Lebenszeit eines Tieres eine blutzellenbildende Funktion.[30]
Die Gebärmutter ist bei den Langnasen- und den Kugelgürteltieren einfach gestaltet und trapezförmig, womit sie jener der Primaten gleicht. Alle anderen Gürteltiere verfügen über eine zweihörnige Gebärmutter.[31][32] Weibliche Tiere besitzen ein einzelnes Paar Milchdrüsen, mit Ausnahme der Langnasengürteltiere, die zwei Paare aufweisen. Der Penis ist im Vergleich zur Körpergröße einer der längsten unter allen Säugetieren und erreicht bei einigen Arten im erigierten Zustand bis zu 50 % der Körperlänge. Wegen des Körperpanzers wäre die Begattung ohne einen derart großen Penis gar nicht möglich.[20][7]
Vor allem der Geruchssinn ist stark ausgeprägt und wird häufig bei der Nahrungssuche eingesetzt. Dies spiegelt sich auch am Gehirn wider, das ein großes Riechhirn besitzt. Der Sehsinn ist dagegen unterentwickelt, was ebenfalls durch eine weniger ausgeprägte Area optica am Gehirn diagnostiziert werden kann.[7]
Verbreitung und Lebensraum
Gürteltiere leben ausschließlich auf dem amerikanischen Doppelkontinent. Das Vorkommen der meisten Arten ist auf Südamerika beschränkt, wo sie aber über den größten Teil der Landfläche verbreitet sind. Hier ist auch der Ursprung der Gürteltiere anzusiedeln, der bis in das ausgehende Paläozän vor rund 58 Millionen Jahre zurückreicht. In Mittelamerika finden sich lediglich zwei Arten, darunter die bekannteste Gürteltierart, das Neunbinden-Gürteltier (Dasypus novemcinctus), das große Teile von Süd- und Mittelamerika bis einschließlich der südöstlichen USA bewohnt. Die Vorfahren der nord- und mittelamerikanischen Arten sind vermutlich erst nach dem Entstehen der Landbrücke zwischen Nord- und Südamerika am Isthmus von Panama und dem damit verbundenen Großen Amerikanischen Faunenaustausch im Pliozän vor rund 3 Millionen Jahren aus Südamerika eingewandert.[1][2]
Zahlreiche Gürteltierarten bevorzugen trockene und offene Lebensräume wie Halbwüsten, Savannen und Steppen mit dorniger Gebüschvegetation oder Trockenwäldern und benötigen zudem einen lockeren Untergrund zum Anlegen ihrer Baue. Dadurch sind viele Vertreter im Gran Chaco, aber auch in den Pampa-Gebieten des zentralen Südamerikas vom südlichen Brasilien über Bolivien und Paraguay bis ins nördliche Argentinien anzutreffen; allein in Paraguay sind 12 der 21 heute noch lebenden Arten nachgewiesen.[6] Einige Arten leben aber auch im tropischen Regenwald, in den Yungas-Wäldern der Hochgebirgsflanken oder in Feuchtgebieten wie dem Pantanal. Die direkten Hochlagen der Anden haben aber nur einzelne Gürteltierarten erschlossen, etwa das Kleine Borstengürteltier („Andenborstengürteltier“; Chaetophractus vellerosus).[33][1][2]
Lebensweise
Territorialverhalten

Die Lebensweise der meisten Gürteltierarten ist nur wenig erforscht. Verlässliche Daten beziehen sich in der Regel auf die weiter verbreiteten und häufigeren Arten, etwa das Neunbinden- oder das Sechsbinden-Gürteltier (Euphractus sexcinctus). Gürteltiere sind überwiegend nachtaktive Einzelgänger; einige Arten erscheinen jedoch auch tagsüber. Insgesamt können drei Lebensmodelle unterschieden werden:[23]
- überwiegende Bodenbewohner, die nur selten unterirdische Baue graben, etwa die Kugelgürteltiere (Tolypeutes);
- überwiegend grabende Tiere, die aber oberflächlich auf Nahrungssuche gehen, etwa die Langnasengürteltiere (Dasypus) und Borstengürteltiere (Chaetophractus);
- generell grabende Tiere, die auch unterirdisch Nahrung aufnehmen, etwa die Nacktschwanzgürteltiere (Cabassous) und beide Vertreter der Gürtelmulle (Chlamyphorus und Calyptophractus).
Einen Großteil der Wachphase verbringen Gürteltiere mit der Nahrungssuche, meist behände und permanent, wobei der bestens entwickelte Geruchssinn die Beute bis zu 20 cm tief im Erdboden aufspürt, die daraufhin meist ausgegraben wird. Dabei sind einige Vertreter der Gürteltiere in der Lage, bis zu sechs Minuten lang die Luft anzuhalten, um die Atemwege freizuhalten. Trotz des plumpen und scheinbar starren Körperbaus können sich die Gürteltiere zudem erstaunlich flink fortbewegen. Einige Arten sind sogar gute Schwimmer; damit sie in ihrer Panzerung nicht untergehen, pumpen sie vorher Luft in Magen und Darm. Zum Schlafen graben sie sich im Boden ein oder suchen einen bereits vorhandenen Bau auf, die Wiedernutzung von derartigen Bauen ist aber von Art zu Art deutlich unterschiedlich. Die Wohnhöhlen selbst sind ebenfalls je nach Art unterschiedlich und können klein sein, aber auch verzweigt und mehrere Meter lang im Untergrund verlaufen. Zum Teil werden am Ende der Gänge auch vergrößerte Nistplätze angelegt.[1][2]
Ernährung

Die Gürteltiere haben ein weites Nahrungsspektrum, allerdings werden Insekten je nach Menge mehr oder weniger von allen Vertretern gefressen. Es können hier jedoch verschiedene, artabhängige Ernährungsstrategien unterschieden werden:[7][1][2]
- überwiegende Carnivoren und Omnivoren, etwa das Sechsbinden-Gürteltier (Euphractus), das Zwerggürteltier (Zaedyus) und die Borstengürteltiere (Chaetophractus); diese vertilgen neben Insekten auch kleine Wirbeltiere wie Eidechsen und Mäuse, seltener Aas und Pflanzenkost;
- opportunistische Insektenfresser, etwa die Kugelgürteltiere (Tolypeutes), die Langnasengürteltiere (Dasypus) und der Gürtelmull (Chlamyphorus); diese ernähren sich von den verschiedensten Insekten wie Ameisen, Termiten, aber auch Käfer und deren Larven sowie Heuschrecken und Spinnen;
- spezialisierte Insektenfresser, etwa das Riesengürteltier (Priodontes) und die Nacktschwanzgürteltiere (Cabassous); deren Nahrung umfasst weitgehend Ameisen und Termiten.
Ebenso ernährten sich die ausgestorbenen Gürteltiere anhand untersuchter Zähne weitgehend omnivor oder insektivor, allerdings gibt es auch Hinweise auf eine deutlich fleischhaltigere Nahrung, etwa beim großen Macroeuphractus, dessen zweiter Zahn markant an einen Eckzahn erinnerte und der möglicherweise bis zu hasengroße Tiere erbeutete.[3] Vertreter aus der Gruppe der Eutatini, etwa Eutatus, wiederum waren mehr auf pflanzliche Nahrung spezialisiert, was anhand des Baus des Unterkiefers und der Abrasionsspuren der Zähne ermittelt werden konnte.[34]
Die Nahrungsaufnahme erfolgt in der Regel mit der langen, klebrigen Zunge. Die langen Krallen der Vordergliedmaße dienen zum Aufreißen der Ameisen- und Termitenhügel, die mitunter sehr hart sein können, oder zum Graben von Löchern, in denen nach Nahrung gesucht wird. Bei der Nahrungssuche richten sich Gürteltiere häufig auf die Hinterbeine auf, um eine Duftspur zu verfolgen.[1][2]
Energiehaushalt und Konsequenzen des Panzers
Generell haben die Gürteltiere eine sehr niedrige Stoffwechselrate; sie liegt bei etwa 40 bis 60 % derer, die bei gleich großen Säugetieren zu erwarten wäre. Neben der der Kloakentiere gehört sie zu der niedrigsten unter den Säugetieren.[35] Dabei ist der Metabolismus umso geringer, je stärker die Tiere an Ameisen und Termiten als primäre Nahrungsressourcen angepasst sind. Verursacht wird der langsame Stoffwechsel durch die geringe Energiedichte der Nahrung, durch das zusätzlich aufgenommene Erdmaterial beim Herumwühlen in den Insektennestern vermindert sich diese zusätzlich. Der Effekt wirkt sich bei größeren Arten mitunter stärker aus als bei kleineren, da unter anderem beim Riesengürteltier die Stoffwechselrate teils nur 29 % erreicht.[36][37] Der sich je nach Aktivität unter Umständen ergebende höhere Sauerstoffbedarf wird anders als bei den meisten Säugetieren nicht durch tiefere, sondern durch schnellere Atemzüge gedeckt. Die Notwendigkeit dieses Verhaltens ergibt sich aus der Ausbildung des Panzers, der den Brustkorb einengt und so dessen Bewegungsspielraum beschränkt.[38][20]
Die langsame Stoffwechselrate geht des Weiteren mit einer niedrigen sowie schwankenden Körpertemperatur und einer hohen Wärmeleitfähigkeit einher. Die durchschnittliche Körpertemperatur liegt bei tropisch verbreiteten Formen bei etwa 33 bis 34,5 °C, sie ist etwas niedriger als bei Tieren aus kühleren Regionen. Die hohe Wärmeleitfähigkeit wird durch die Ausbildung des Panzers und das Fehlen eines dichten Fells begünstigt, sie fördert die Abgabe der Körperwärme an die Umgebung und verhindert dadurch eine zu starke Aufheizung unter warmklimatischen Bedingungen. Zur Verhinderung einer zu schnellen Körperwärmeabgabe ziehen sich die Tiere bei lokalen starken Temperaturschwankungen in ihre unterirdischen Baue mit deutlich ausgeglicheneren Umgebungsbedingungen zurück. In kälteren Regionen auftretende Arten wie das Zwerggürteltier reagieren auf unwirtliche Phasen mit einem Torpor oder begegnen diesen wie beim Neunbinden-Gürteltier mit verminderter Aktivität. Teilweise kommt es bei diesen kälteresistenteren Vertretern auch zum Ansammeln eines Fettpolsters. Eine derartige Speicherstrategie der Nahrung ist bei vielen tropisch verbreiteten Arten kaum belegt. So würde ein zusätzliches Fettpolster etwa die Kugelgürteltiere nicht mehr befähigen, sich vollständig in ihren Panzer einzurollen.[39][40][37][41][20]
Die generelle Konstitution und der Energiehaushalt bedingen, dass Gürteltiere häufiger in warmklimatischen Landschaften verbreitet sind und verhindern eine stärkere Ausbreitung in kühlklimatische Breiten. Als weiterer limitierender Faktor kommt die Verfügbarkeit der Ameisen und Termiten zur Deckung des Nahrungsbedarfs hinzu. Die staatenbildenden Insekten gedeihen unter winterkalten Bedingungen schlecht. Dies alles begrenzt beispielsweise die weitere Nordexpansion des Neunbinden-Gürteltiers erheblich.[40] Des Weiteren haben die Ernährung und der daraus resultierende Stoffwechsel unmittelbaren Einfluss auf die Lebensstrategien verschiedener Tiergruppen. Bei den Gürteltieren führen die energiearme Kost und der niedrige Metabolismus zu einer unterirdischen Lebensweise. Der ausgebildete Panzer bietet wiederum einen gewissen Schutz vor sowohl Beutegreifern als auch einstürzenden Tunneln und ähnlichem. Sie behielten dadurch auch eine mehr oder weniger „urtümliche“ Fortbewegung mit Sohlengang und kurzen Gliedmaßen bei, die zwar eine prinzipiell schnelle, durch kurze Schritte aber weniger effektive Fortbewegung zulassen. Ähnliches ist auch von anderen insektenfressenden Säugetieren mit äußeren Schutzbildungen bekannt, seien es die Schuppentiere oder die vielfältigen, mit stacheligem Fellkleid ausgestatteten Tiere wie die Igel oder verschiedene Vertreter der Tenreks. Dagegen bildete sich bei den zu den Gürteltieren vergleichbar großen Hasenartigen durch ihre energiereiche Ernährung und ihrer hohen Stoffwechselrate ein hochmobiler Bewegungsapparat heraus, der durch lange Gliedmaßen charakterisiert ist. Sie stellen mit ihren hohen Geschwindigkeiten und großen Schrittlängen effektive Fluchttiere dar, so dass hier eine Körperpanzerung nicht erforderlich wurde.[35][42][20]
Fortpflanzung
Die Fortpflanzung ist nur bei den wenigsten und vor allem am weitesten verbreiteten Arten gut erforscht. Generell ist die Brunftzeit die einzige Phase, in der mehrere Individuen der Gürteltiere zusammenkommen. Die Tragzeit ist recht unterschiedlich und kann bei den bekannteren Arten zwischen zwei und vier Monaten variieren. Beim Neunbinden-Gürteltier ist nachgewiesen, dass die befruchteten Eizellen mehrere Monate im Körper aufbewahrt werden, bevor deren Nidation stattfindet und die weitere Entwicklung folgt. Dadurch wird verhindert, dass die Weibchen in ungünstigen Zeiten chancenlose Nachkommen gebären. Darüber hinaus ist beim Neunbinden-Gürteltier und zusätzlich beim Südlichen Siebenbinden-Gürteltier Polyembryonie bekannt, so dass diese genetisch identische Nachkommen erzeugen, deren Anzahl bei zwei bis zwölf pro Wurf liegt. Für alle anderen Vertreter der Langnasengürteltiere wird diese Form der Reproduktion ebenfalls vermutet. Es ist allerdings bisher unklar, ob die Polyembryonie mit der Entwicklung der einfach gebauten Gebärmutter bei diesen Gürteltiervertretern zusammenhängt. Bei vier weiteren Gattungen ist die Geburt von nicht identischen Zwillingen bezeugt, die restlichen Arten bringen jeweils nur einen Nachkommen pro Wurf zur Welt.[32][43] Die Jungen haben anfangs noch eine weiche, ledrige Haut, die nach und nach zum Panzer aushärtet, zudem werden sie meist nur wenige Wochen gesäugt. Die Lebenserwartung der Gürteltiere beträgt in freier Wildbahn bis zu 18 Jahre, in menschlicher Obhut erreichen einige Arten sogar mehr als 30 Jahre, allerdings konnte bisher nicht bei allen Arten ein Zuchterfolg verzeichnet werden.[20][1][2]
Fressfeinde und Feindverhalten

Bei Gefahr suchen Gürteltiere meist den nächstgelegenen Bau auf oder graben sich ein, wo sie sich dann mit den Krallen in den Boden rammen sowie die Knochenplatten abspreizen und in die Tunnelröhre pressen und sich so fest im Erdboden verankern. Im Freien werden ebenfalls die Krallen in den Untergrund gedrückt, zusätzlich pressen sich die Tiere dabei fest an den Boden, so dass nur die Panzerung attackiert werden kann und der weiche Bauch geschützt ist. Allerdings bietet dann der Panzer keinen vollständigen Schutz, da er bei einigen Vertretern nur 2 bis 3 mm dick ist und von einigen größeren Raubtieren leicht geknackt werden kann. Zu einer rundum geschützten Kugel können sich nur die Kugelgürteltiere zusammenrollen; dabei ist der Panzer so lückenlos verzahnt und der Muskelschluss so fest, dass kaum ein Fressfeind diese Schale zerbeißen kann. Die Krallen lassen sich zudem gut zur Verteidigung einsetzen.[20] Zu den potenziellen Fressfeinden gehören die größeren Raubkatzen wie Jaguar und Puma, aber auch kleinere Beutegreifer, etwa Kojote und Ozelot oder Vögel, so der Aguja und der Zaunadler. Ebenfalls stellen freilebende Hunde und Katzen Gürteltieren nach. Jungtiere können auch von zahlreichen weiteren Beutegreifern erlegt werden.[7][44]
Ökologische Bedeutung
Gürteltiere üben einen großen Einfluss auf ihr jeweiliges lokales Habitat aus. Die grabenden Vertreter leisten einen Beitrag zur Bioturbation und damit zur Durchlüftung und Auflockerung von Bodensubstraten. Die dabei bewegte Erdmenge beträgt, abhängig von der Größe des Verursachers, je Bau zwischen 0,002 und 0,6 m³. Für eine untersuchte Region in Kolumbien ergab dies bei 157 registrierten Bauen des Riesengürteltiers ein Volumen von schätzungsweise rund 99 m³. Die Baue dienen aber nicht nur den Gürteltieren als Unterschlupf, sondern werden nachfolgend auch von zahlreichen anderen Tieren genutzt. Da die Gürteltiere somit Lebensraum für diverse Lebewesen schaffen, stufen Wissenschaftler sie als ecosystem engineers ein. Besonders gut ist dies beim Riesengürteltier untersucht, dessen Baue teils mehr als zwei Dutzend verschiedenen Arten als Lebensraum dienen. Ebenso haben Gürteltiere durch ihre grabende und wandernde Lebensweise einen Anteil am Nährstoffkreislauf. Dies betrifft nicht nur beispielsweise den Stickstoffgehalt, der bei entsprechender Populationsdichte durch Defäkation lokal deutlich ansteigen kann. Zusätzlich nehmen Gürteltiere durch ihre Ernährungsweise nennenswerte Mengen an Erde auf, die dann andernorts wieder ausgeschieden werden. Einige Arten, die mitunter pflanzenfressend leben, fungieren darüber hinaus als Verbreiter von Samen. Dies schließt mehr als drei Dutzend Pflanzengattungen ein, deren Früchte nachweislich zum Nahrungsspektrum der Gürteltiere gehören. Weitere Aspekte betreffen die Kontrolle der Ausbreitung von Krankheiten durch das Vertilgen zahlreicher unterschiedlicher Insektenarten. Allerdings sind Gürteltiere auch Träger mehrerer Krankheitserreger, die dem Menschen gefährlich werden können. Zu nennen wären hier unter anderem die Lepra und die Chagas-Krankheit. Nicht zuletzt bilden Gürteltiere einen Teil des Nahrungsnetzes, da sie nicht nur selbst im gewissen Maße räuberisch leben, sondern auch von anderen Predatoren erbeutet werden.[44]
Systematik
Äußere Systematik
Verwandtschaftsverhältnis der Gürteltiere laut morphologischen Daten nach Billet et al. 2011[45]
|

Die Gürteltiere (Dasypoda) sind eine Gruppe aus der Ordnung der Gepanzerten Nebengelenktiere (Cingulata) und der Überordnung der Nebengelenktiere (Xenarthra). Als gemeinsames Merkmal der ansonsten sehr vielgestaltigen Nebengelenktiere gelten die xenarthrischen Gelenke an der Wirbelsäule. Ebenfalls zu den Xenarthra zu stellen sind die Faultiere (Folivora) und die Ameisenbären (Vermilingua), die beide eine enger verwandte Gruppe darstellen und als Ordnung der Zahnarmen (Pilosa) den Gürteltieren gegenüberstehen. Molekulargenetische Analysen zeigten auf, dass die Trennung der gemeinsamen Linie der Faultiere und Ameisenbären von der der Gürteltiere bereits zu Beginn des Paläozäns vor rund 65 Millionen Jahren erfolgt war. Die Xenarthra, deren Ursprung bis in die Oberkreide zurückreicht, bilden eine der vier großen Überordnungen der Höheren Säugetiere, die drei anderen werden teilweise in einem übergeordneten Taxon zusammengefasst, den Epitheria, das den Xenarthra als Schwestergruppe gegenübersteht. Alle drei rezenten Ordnungen sind heute auf Amerika begrenzt.[46][47]
Die Zusammensetzung und systematische Gliederung der Gepanzerten Nebengelenktiere ist komplex und in Diskussion. Allgemein können neben den Gürteltieren einige andere größere, nur fossil überlieferte Linien unterschieden werden.[48] Von Bedeutung sind hier unter anderem die Pampatheriidae und Glyptodontidae, die gemäß phylogenetischen Untersuchungen als sehr nah miteinander verwandt gelten und daher in der höheren taxonomischen Einheit der Glyptodonta zusammengefasst werden. Die Pampatherien traten erstmals im Mittleren Miozän auf und verfügen ebenfalls über einen starren Schulter- und Beckenpanzer, zwischen denen sich einzelne, zumeist drei, bewegliche Bänder befinden. Allerdings waren die Vertreter vor allem in der Spätphase ihrer Stammesgeschichte wesentlich größer als die heutigen Gürteltiere. So erreichte die nordamerikanische Gattung Holmesina, die erst am Ende des Pleistozän vor rund 10.000 Jahren ausstarb, eine Länge von zwei 2 m und ein Körpergewicht von bis zu 220 kg; ähnliche Ausmaße besaß das zur gleichen Zeit in Südamerika verbreitete Pampatherium. Ursprünglich hielt man die Pampatherien als nahe verwandt mit den Gürteltieren, teilweise wurden sie auch als Unterfamilie innerhalb dieser geführt.[49] Die nähere Beziehung der Pampatherien und Glyptodonten zueinander ergibt sich jedoch unter anderem aus der Struktur des Gehörganges und dem Bau des Kauapparates, wie der hohe Unterkiefer sowie die komplexer gestalteten Zähne.[50] Die Glyptodonten wiederum bilden eine der umfangreichsten Gruppen innerhalb der Cingulata. Sie sind durch einen starren Panzer und durch das Fehlen der xenarthrischen Gelenke gekennzeichnet, aufgrund ihrer hochkronigen Zähne verfolgten sie wohl ähnlich wie die Pampatherien eine eher grasfressende Lebensweise. Glyptodonten traten erstmals im Mittleren Eozän auf und waren anfänglich noch recht klein, so wog das aus dem Unteren Miozän stammende Propalaehoplophorus nur rund 74 kg.[5] Sie erreichten aber im Pleistozän riesige Ausmaße mit einem Gewicht von über 2 t wie bei Doedicurus.[51][10]
Im Verlauf des 20. Jahrhunderts sahen die meisten Wissenschaftler die Glyptodonten- und die Gürteltierverwandtschaft als zwei getrennte Entwicklungslinien an. Skelettanatomische Untersuchungen aus dem Jahr 2006, durchgeführt von Timothy J. Gaudin und John R. Wible, verwiesen die Glyptodonta jedoch in eine verwandtschaftliche Nähe der Gürteltiere. Hier erwies sich vor allem das unmittelbaren Umfeld des Sechsbinden-Gürteltiers (Euphractus) und der Borstengürteltiere (Chaetophractus) als bedeutend. Allerdings ergaben sich dadurch erste Hinweise auf einen paraphyletischen Ursprung der Gürteltiere.[50] Dies wurde durch weitere anatomische Studien im Jahr 2011 untermauert, die die Glyptodonta noch tiefer in die Gürteltierverwantschaft hineinrückten.[45] Neuere genetischen Analysen aus dem Jahr 2016 an fossilem Erbgut der Glyptodontidae, namentlich von Doedicurus, kamen zu einem vergleichbaren Ergebnis. Demnach gruppieren sich zumindest die Glyptodonten innerhalb der Chlamyphoridae, anders als bei den anatomischen Studien ist hier aber eine engere Bindung an die Verwandtschaftsgruppe um die Kugelgürteltiere (Tolypeutes) und die Gürtelmulle wahrscheinlicher. Dadurch bestätigte sich einerseits die paraphyletische Herkunft der Gürteltiere als gesamte Einheit unter Einbeziehung ausgestorbener Gruppen, sie zeigten aber auch auf, dass die Glyptodonten keine eigenständige Entwicklungslinie innerhalb der Gepanzerten Nebengelenktiere darstellen. Inwiefern eine ähnliche Stellung auch für die Pampatherien zutrifft, ist momentan aus genetischer Sicht unklar, da noch kein DNA-Material sequenziert wurde.[52][53]
Des Weiteren bestehen innerhalb der Cingulata noch einige andere ausgestorbene Gruppen. Hierzu gehören die Peltephilidae, die Palaeopeltidae und die Pachyarmatheriidae. Ihre Verwandtschaftsverhältnisse zu den Gürteltieren sind nicht ganz eindeutig geklärt. Erstere könnten sehr basal im Stammbaum der Gepanzerten Nebengelenktiere stehen,[45] letztere formen eventuell die Schwestergruppe der Glyptodonten und Pampatherien.[54]
Innere Systematik
Innere Systematik der Gürteltiere nach Gibb et al. 2015[55]
|
Die heutigen Gürteltiere werden nach molekulargenetischen Untersuchungen aus dem Jahr 2015 in zwei Familien untergliedert, die sich aus neun Gattungen und 20 Arten zusammensetzen. Die eine Familie umfasst mit den Dasypodidae die Langnasengürteltiere (Dasypus), die andere, Chlamyphoridae genannt, schließt alle anderen bekannten Gattungen ein. Die Aufspaltung der heutigen Gürteltiere in diese beiden Entwicklungslinien begann laut den genetischen Daten bereits sehr früh und fand im Verlauf des Mittleren Eozäns vor rund 45 Millionen Jahren statt.[56][57][58][55] Die gegenwärtigen Gürteltiere bilden allerdings nur einen Teil der einstigen Artenvielfalt ab. Es bestehen innerhalb der beiden Familien insgesamt vier Hauptlinien (Unterfamilien), zu denen auch ein nennenswerter Teil der fossil überlieferten Vertreter zugeordnet wird. Die vier Hauptlinien sind sowohl genetisch als auch anatomisch gut abgrenzbar. Die älteste Linie stellen dabei die Dasypodinae mit den Langnasengürteltieren (Dasypus) dar. Sie bilden außerdem die Tribus der Dasypodini. Weiterhin sind hier noch die ausgestorbenen Linien der Stegotheriini und der Astegotheriini hinzuzufügen, letztere umfassen die ursprünglichste Gruppe der Gürteltiere, die bereits im ausgehenden Paläozän fossil belegt ist.[59][60]
Die drei anderen Linien erschienen relativ schnell hintereinander. Noch im Oberen Eozän vor 37 Millionen Jahren formten sich die Euphractinae heraus,[56][57][58][55] fossil sind diese aber seit mindestens 42 Millionen Jahren nachgewiesen. Ihr gehören neben dem namensgebenden Sechsbinden-Gürteltier (Euphractus) auch das Zwerggürteltier (Zaedyus) und die Borstengürteltiere (Chaetophractus) an. Sie werden zusammen in der Tribus der Euphractini vereint, denen auch mehrere ausgestorbene Vertreter angehören. Ihnen gegenüber stehen die Eutatini, eine erloschene Linie mit zahlreichen Gattungen.[61] Die Eutatini gehörten zu den formenreichsten Vertretern der Gürteltiere, sie lebten allerdings ausschließlich im südlichen Südamerika, wo sie anhand teilweise gut erhaltener Fossilfunde nachgewiesen sind, die zudem die Feststellung erlauben, dass ein äußerst dichtes Haarkleid als Adaption an kühles Klima ausgebildet war.[62]
Im Oligozän vor rund 33 Millionen Jahren spalteten sich dann die Chlamyphorinae und die Tolypeutinae voneinander ab, die beiden letzten großen Linien.[56][57][58][55] Den Chlamyphorinae werden die beiden Vertreter der Gürtelmulle (Chlamyphorus und Calyptophractus) zugeordnet. Von beiden Arten liegt kein nennenswertes Fossilmaterial vor, eine ausgestorbene Gattung ist mit Chlamydophractus aus dem Oberen Miozän belegt.[58][63] Die Tolypeutinae führen die Kugelgürteltiere (Tolypeutes), das Riesengürteltier (Priodontes) und die Nacktschwanzgürteltiere (Cabassous) zusammen. Dabei formen die letzten beiden die Tribus der Priodontini, ersteres steht dagegen in der Tribus der Tolypeutini. Ausgestorbene Linien werden hier nicht unterschieden, doch ist mit Kuntinaru ein sehr basaler Vertreter der Tolypeutinae aus dem Oligozän dokumentiert.[45][6]
Überblick über die rezenten und fossilen Gattungen der Gürteltiere
Innerhalb der Gürteltiere werden rund 60 Gattungen unterschieden, einige dieser Gattungen, etwa Dasypus, Chasicotatus,[62] Eutatus[64] oder Stegotherium,[59] sind sehr formenreich und umfassen teils über ein halbes Dutzend Arten. Die Gliederung basiert auf dem Überblick von Malcolm C. McKenna und Susan K. Bell aus dem Jahr 1997,[65] berücksichtigt aber auch neuere Entwicklungen zur höheren Systematik der Gürteltiere[50][55][66][67] ebenso wie danach neu eingeführte Gattungen:[68][69][12][70][45][71][60][72][73][74][75][76][63][77][78][79]
- Dasypoda Quenstedt, 1885 (Hoernes, 1886)
- ? Yuruatherium Ciancio, Carlini, Campbell & Scillato-Yané, 2013
- ? Proeocoleophorus Sedor, Oliveira, Silva, Fernandes, Cunha, Ribeiro & Dias, 2017
- ? Saltatherium Fernicola, Zimicz, Chornogubsky, Cruz, Bond, Arnal, Cárdenas & Fernández, 2021
- Lumbreratherium Herrera, Powell, Esteban & del Papa, 2017
- Pucatherium Herrera, Powell & del Papa, 2012
- Punatherium Ciancio, Herrera, Aramayo, Payrola & Babot, 2016
- Noatherium Fernicola, Zimicz, Chornogubsky, Ducea, Cruz, Bond, Arnal, Cárdenas & Fernández, 2021
- Eocoleophorus Oliveira, Ribeiro & Bergqvist, 1997
- Familie Dasypodidae Gray, 1821
- Unterfamilie Dasypodinae Gray, 1821
- Tribus Astegotheriini Ameghino, 1906
- Riostegotherium Oliveira & Bergqvist, 1998
- Prostegotherium Ameghino, 1902
- Astegotherium Ameghino, 1902
- Pseudostegotherium Ameghino, 1902
- Parastegosimpsonia Ciancio, Carlini, Campbell & Scillato-Yané, 2013
- Nanoastegotherium Carlini, Vizcaíno & Scillato-Yané, 1997
- Stegosimpsonia Oliveira & Vizcaíno, 1994
- Tribus Stegotheriini Gray, 1821
- Stegotherium Ameghino, 1887
- Tribus Dasypodini Gray, 1821
- Plesiodasypus Barasoain, González-Ruiz, Zurita & Villarroel, 2021
- Anadasypus Carlini, Vizcáıno & Scillato-Yané, 1997
- Propraopus Ameghino, 1881
- Pliodasypus Castro, Carlini, Sánchez & Sánchez-Villagra, 2014
- Dasypus Linnaeus, 1758 (Langnasengürteltiere; einschließlich sieben rezenter Arten)
- Familie Chlamyphoridae Bonaparte, 1850
- Unterfamilie Tolypeutinae Gray, 1865
- Kuntinaru Billet, Hautier, de Muizon & Valentin, 2011
- Tribus Tolypeutini Gray, 1865
- Pedrolypeutes Carlini, Vizcaíno. & Scillato-Yané, 1997
- Tolypeutes Illiger, 1811 (Kugelgürteltiere; einschließlich zwei rezenter Arten)
- Tribus Priodontini Gray, 1873
- Vetelia Ameghino, 1891
- Priodontes Cuvier, 1825 (Riesengürteltier; einschließlich einer rezenten Art)
- Cabassous McMurtrie, 1831 (Nacktschwanzgürteltiere; einschließlich vier rezenter Arten)
- Unterfamilie Chlamyphorinae Bonaparte, 1850
- Chlamydophractus Barasoain, Tomassini, Zurita, Montalvo & Superina, 2019
- Chlamyphorus Harlan, 1825 (Gürtelmull; einschließlich einer rezenten Art)
- Calyptophractus Fitzinger, 1871 (Burmeister-Gürtelmull; einschließlich einer rezenten Art)
- Unterfamilie Euphractinae Winge, 1923
- Coelutaetus Ameghino, 1902
- Dasypodon Castellani, 1925
- Tribus Utaetini Simpson, 1945
- Utaetus Ameghino, 1902
- Tribus Eutatini Bordas, 1933
- Meteutatus Ameghino, 1902
- Barrancatatus Carlini, Ciancio & Scillato-Yané, 2010
- Paraeutatus Scott, 1933
- Sadypus Ameghino, 1902
- Stenotatus Ameghino, 1891
- Pseudeutatus Ameghino, 1891
- Ringueletia Reig, 1958
- Chasicotatus Scillato-Yané, 1979
- Doellotatus Bordas, 1932
- Proeutatus Ameghino, 1891
- Eutatus Gervais, 1867
- Tribus Euphractini Winge, 1923
- Parutaetus Ameghino, 1902
- Orthutaetus Ameghino, 1902
- Archaeutatus Ameghino, 1902
- Anteutatus Ameghino, 1902
- Mazzoniphractus Carlini, Ciancio & Scillato-Yané, 2010
- Paleuphractus Kraglievich, 1934
- Archeuphractus Kraglievich, 1934
- Paraeuphractus Scillato-Yané, 1980
- Prozaedyus Ameghino, 1891
- Chorobates Reig, 1958
- Macrochorobates Scillato-Yané, 1980
- Amblytatus Ameghino 1902
- Paraeuphractus Scillato-Yané, 1980
- Anutaetus Ameghino 1902
- Eodasypus Ameghino 1894
- Hemiutaetus Ameghino 1902
- Isutaetus Ameghino 1902
- Macroeuphractus Ameghino, 1887
- Proeuphractus Ameghino, 1886
- Zaedyus Ameghino, 1889 (Zwerggürteltier; einschließlich einer rezenten Art)
- Euphractus Wagler, 1830 (Sechsbinden-Gürteltier; einschließlich einer rezenten Art)
- Chaetophractus Fitzinger, 1871 (Borstengürteltiere; einschließlich zwei rezenter Arten)
Taxonomie

Gürteltiere wurden in Europa erstmals Anfang des 16. Jahrhunderts mit der Eroberung Amerikas durch die spanischen Conquistadoren bekannt. Francisco Hernández de Córdoba, der 1517 die Halbinsel Yucatán entdeckte, benutzte als Bezeichnung für das Neunbinden-Gürteltier den lokalen aztekischen Namen Azotochtli, was übersetzt so viel wie „Schildkrötenhase“ bedeutet und sich auf das Aussehen des Tieres bezieht.[80] Im Jahr 1553 veröffentlichte Pierre Belon eine der ältesten bekannten Zeichnungen eines Gürteltiers und benannte die Tiere als Tatou. Er stellte die Tiere dabei an die Seite der Igel.[81][82] Mehr als 200 Jahre später, 1763, gab Georges-Louis Leclerc de Buffon in seinem mehrbändigen Werk Histoire naturelle, générale et particuliére einen umfassenden Überblick über die damals bekannten Arten.[83] Nur fünf Jahre zuvor hatte aber Linnaeus in der 10. Ausgabe des Systema Naturae die wissenschaftliche Bezeichnung Dasypus für das Neunbinden-Gürteltier eingeführt. Dabei verwies er für die Wortherkunft auf Hernández. Es wird angenommen, dass Linnaeus den aztekischen Namen Azotochtli als ungeeignet für eine Latinisierung und damit für eine Verwendung in der binären Nomenklatur erachtete und daher Dasypus verwendete (das Wort dasypus ist griechischer Herkunft (δασύπους) und bedeutet direkt übersetzt „Rauhfuß“, wurde aber teilweise auch auf den Hasen bezogen[84]).[85][86] Gleichzeitig belegte er das Siebenbinden-Gürteltier (Dasypus septemcinctus), das Sechsbinden-Gürteltier (Euphractus sexcinctus), das Nördliche Kugelgürteltier (Tolypeutes tricinctus) und das Südliche Nacktschwanzgürteltier (Cabassous unicinctus) ebenfalls mit dem Gattungsnamen Dasypus. Die einzelnen Arten unterschied er anhand der Anzahl der beweglichen Gürtel.[87]

Einen von Dasypus abgeleiteten Familiennamen führte John Edward Gray im Jahr 1821 ein, womit er heute als Erstbeschreiber der Familie angesehen wird. Er wählte damals Dasipidae als Name der Familie und erkannte als einzige Gattung Dasypus an. Seine kurze Beschreibung lautete:
Face long, conical: teeth, grinders cylindrical, separate from one another, seven or eight in each jaw, inside without enamel: tongue smooth, slightly extensible: body covered with shelly, scaly armour: ear and tail long: cœecum none.
„Gesicht lang, konisch. Zähne, Mahlzähne zylindrisch, voneinander getrennt, sieben oder acht in jedem Kiefer, innen ohne Zahnschmelz. Zunge glatt, leicht ausstreckbar. Körper bedeckt mit schaliger, schuppiger Panzerung. Ohr und Schwanz lang. Blinddarm fehlt.“[88]
Die heute korrekte Namensform Dasypodidae wurde erstmals 1838 von Charles Lucien Jules Laurent Bonaparte verwendet, er galt teilweise auch als Erstbeschreiber der Familie.[49] Die Bezeichnung wurde lange Zeit für die gesamte Gruppe der Gürteltiere verwendet. Die Aufteilung der Gürteltiere in zwei Familien (Dasypodidae und Chlamyphoridae), die aufgrund der Ergebnisse molekulargenetischer Untersuchungen im Jahr 2015 erfolgte, erforderte eine neue Bezeichnung für die übergeordnete Gruppe. Gewählt wurde der Name „Dasypoda“. Die Erstverwendung des Namens ist nicht eindeutig. Friedrich August Quenstedt nutzte ihn im Jahr 1885 in seinem Handbuch der Petrefaktenkunde.[89] Allerdings gebrauchte ihn Rudolf Hoernes im selben Zeitraum. Dabei wird häufig auf das Werk Manuel de paléontologie verwiesen, das 1886 in Paris erschien.[90] Die Bezeichnung Dasypoda findet sich jedoch bereits im zwei Jahre zuvor verlegten deutschsprachigen Original Elemente der Paläontologie.[91] Beide Autoren schlossen die Glyptodonten in die Dasypoda mit ein, aufgrund dessen galt die Bezeichnung zumeist als synonym zu Cingulata.[65] Ende der 1970er Jahre verwendete George F. Engelmann die Bezeichnung Dasypoda, um die Gürteltiere von den Glyptodonta zu trennen, in letzteren fasste er die Glyptodontidae und die Pampatheriidae zusammen. Dieser Ansatz hatte vorerst aber nur kurze Zeit Bestand, unter anderem auch dadurch, dass Engelmann nur wenige trennende Merkmale zwischen den beiden Großgruppen aufführen konnte. Dass die Dasypoda und die Glyptodonta im Sinne Engelmanns eine Entwicklungslinie bilden, konnte später dann durch die skelettanatomischen Untersuchungen von Gaudin und Wible aus dem Jahr 2006 aufgezeigt werden.[50] Unterstützung fand dies auch durch die bereits erwähnten genetischen Untersuchungen an fossiler DNA der Glyptodonten aus dem Jahr 2016.[52][53]
Dasypoda ist außerdem ein höherrangiges Homonym zur Gattung der Hosenbienen (Dasypoda), welche von Pierre André Latreille bereits 1802 eingeführt worden war. Die Etablierung von Dasypoda als Überbegriff für die Gürteltiere stieß später aufgrund der wechselhaften Verwendung in der Vergangenheit auf Kritik. Stattdessen wurde die Bezeichnung Dasypodoidea vorgeschlagen, die wiederum auf Gray zurückgeht.[92]
Etymologie
Der deutsche Trivialname „Gürteltier“ bezieht sich auf die beweglichen Bänder oder Gürtel zwischen den beiden festen Panzerteilen. Die im englischen Sprachraum genutzte Bezeichnung armadillo leitet sich vom spanischen Wort armado ab und bedeutet „kleiner Gepanzerter“. Im spanisch sprechenden Raum, vor allem in Südamerika, wird allerdings überwiegend das Guaraní-Wort tatu als Bezeichnung für das Gürteltier genutzt.[6]
Stammesgeschichte
Ursprung und Adaptive Radiation

Insgesamt ist die Stammesgeschichte der Gürteltiere nur wenig erforscht und es bestehen noch zahlreiche Lücken. Umfangreichere Funde liegen hauptsächlich aus dem Eozän, dem Miozän und dem Oberpleistozän vor. Das überwiegende Fossilmaterial beschränkt sich auf die Reste der Panzerung, weitaus rarer sind Schädel- und Skelettreste erhalten.[93] Insgesamt treten Vertreter der Gruppe vergleichsweise eher selten im Fossilbericht auf, dies betrifft auch zahlreiche der heutigen Gürteltierarten, deren Nachweise in geologischer Vergangenheit nur vereinzelt gelang.[45]
Die frühesten Gürteltiere zeigen einige bedeutende Unterschiede zu den heutigen Arten. Ein auffallendes Merkmal ist die abweichende Panzergestaltung. Der Rückenpanzer bestand häufig nur aus einem festen Teil, der im Beckenbereich ausgebildet war, während sich der gesamte vordere Abschnitt aus beweglichen Bändern zusammensetzte. Einige Formen wiesen auch einen Panzer nur aus beweglichen Bändern auf. Der heutige Panzer der Gürteltiere mit einem festen vorderen und hinteren Abschnitt getrennt durch bewegliche Bänder entstand damit erst im Verlauf der Stammesgeschichte. Sofern fossil erhalten, war bei manchen frühen Formen im Gebiss noch Zahnschmelz ausgebildet. Dieser reduzierte sich im Verlauf der Evolutionsgeschichte. Heute kommt eine extrem dünne und schnell abgekaute Zahnschmelzschicht nur noch bei einigen Langnasengürteltieren vor. Ebenso waren die Zähne nicht typisch nagelartig geformt, wie heute bekannt, sondern hatten teils eine flache, eher lappige (lobate) Kauoberfläche. Dies gilt als ursprünglich bei den Nebengelenktieren und findet sich ebenfalls bei den Glyptodontidae, den Pampatheriidae und einigen der ausgestorbenen großen Bodenfaultiere.[59][94]
Fossil fassbar werden die ersten Gürteltiere im Oberen Paläozän vor rund 58 Millionen Jahren. Bedeutend sind hier vor allem die Funde aus einer Spaltenfüllung der Itaboraí-Formation nahe São José de Itaboraí in Itaboraí im südöstlichen Brasilien, die einige Knochenplättchen und weniges postcraniales Skelettmaterial umfassen. Diese werden heute zur Gattung Riostegotherium gestellt, die in die systematische Nähe der Langnasengürteltiere (Dasypus) gehört. Die Tiere besaßen bereits einen Panzer, der sich aus einem festen und einem beweglichen Teil zusammensetzte.[95][96][69][97]
Eozän
Im Unteren Eozän verblieben die Gürteltiere vorerst im zentralen und südlichen Südamerika, das zu dieser Zeit unter tropischen Klimaeinfluss stand. Aus jener Phase sind hauptsächlich Vertreter der Astegotheriini nachgewiesen. Im heutigen Patagonien Argentiniens liegen zwei Fundstellen, Laguna Fría und La Barda am Mittellauf des Río Chubut. Hier treten vor allem Prostegotherium und Stegosimpsonia auf, zwei eher kleine Vertreter der frühen Gürteltiere.[98] Möglicherweise im Mittleren Eozän sind erstmals Angehörige der Euphractinae nachgewiesen. Von herausragender Bedeutung ist hier Gran Barranca im zentralen Patagonien. Die überaus komplexe Fundstelle barg unter anderem Reste bisher seltener Vertreter wie Mazzoniphractus und Orthutaetus, aber auch häufiger dokumentierte Formen wie Meteutatus, Parutaetus und Utaetus. Letzteres kann zahlreiche Reste des Körperskeletts vorweisen, die ein rund 2 bis 3 kg schweres Tier rekonstruieren lassen, das noch Zähne mit Zahnschmelz besaß. Sein Panzer verfügte darüber hinaus lediglich im Beckenbereich über eine feste Struktur, während der gesamte vordere Abschnitt aus beweglichen Bändern bestand.[70][4] Sowohl Utaetus als auch Meteutatus, Parutaetus fanden sich auch in der zu den unteren Abschnitten von Gran Barranca gleichalten Guabirotuba-Formation im brasilianischen Bundesstaat Paraná. Dort verweisen auch mehrere atypische Osteoderme auf Proeocoleophorus, dessen verwandtschaftliche Verhältnisse aber unklar sind.[76][99]
In der Lumbrera-Formation im nordwestlichen Argentinien, die in das Untere und Mittlere Eozän datiert, fehlen hingegen die aus Patagonien bekannten Gürteltierformen bisher weitgehend. Dafür stammt von dort Noatherium, dessen Verwandtschaftsverhältnis innerhalb der Gürteltiere noch nicht eindeutig ist.[78] Als Besonderheit wurde des Weiteren ein für die frühen Gürteltiere sehr seltener Schädel von Lumbreratherium aufgefunden. Dessen Gebiss war heterodont mit einem vorderen eckzahnähnlichen Zahn, der von den restlichen molarenartigen Zähnen durch eine große Lücke (Diastema) getrennt war.[75] Die Gesteinseinheit barg zudem auch ein Teilskelett von Pucatherium, das aus dem Körperpanzer und darin eingebetteten Teilen des Bewegungsapparates besteht. Sowohl bei Lumbrerartherium als auch bei Pucatherium wurde der Panzer nur aus beweglichen Bändern gebildet, während er bei Noatherium übereinstimmend mit Utaetus im hinteren Abschnitt, am Becken, einen fest verwachsenen Bereich besaß. Dadurch lassen sich im Verlauf des Eozän verschiedene Konstruktionstypen des Panzers nachweisen.[100][78] Pucatherium wurde in der Region des Weiteren auch aus der Casa-Grande-, der Quebrada-de-los-Colorados- und der Geste-Formation beschrieben. Letztere erbrachte unter anderem zusätzlich Punatherium. In der Regel sind aber aus den genannten Fundlokalitäten nur Knochenplättchen überliefert. Zeitlich können sie etwas jünger angesetzt werden als die Lumbrera-Formation.[74][93][79]
Zu jener Zeit kam es auch zu den ersten Ausbreitungen in weiter nördlich gelegene Teile Südamerikas. So sind aus der Nähe von Santa Rosa am Río Yurúa im Osten Perus mehrere Arten nachgewiesen, etwa Yuruatherium und Parastegosimpsonia, die in das späte Eozän datieren, letzteres gehört den Astegotheriini an, ersteres ist noch nicht genau zuordenbar.[60][93]
Oligozän
Abkühlende Klimaverhältnisse im Übergang vom Eozän zum Oligozän führten zu einem deutlichen Anstieg euphractiner Gürteltiere, die offensichtlich mit den vorherrschenden Temperaturen besser zurecht kamen. Im frühen Abschnitt der Zeitphase traten in Termas del Flaco im zentralen Chile unter anderem zwei Gattungen auf, Parutaetus und Meteutatus, die dort anhand der Panzerplättchen der Rückenschilde, aber auch durch zahlreiches Schädel- und postcraniales Skelettmaterial nachgewiesen wurden, darunter auch ein nahezu vollständiger Panzer.[12] Eine vergleichbare Gürteltier-Gemeinschaft bestand in Quebrada Fiera im westlichen Argentinien, wo unter anderem neben Meteutatus zusätzlich auch Stenotatus dokumentiert ist.[101] Die weiterfolgende Sukzession in Gran Barranca im zentralen Patagonien zeigt wiederum eine hohe Vielfalt an Gürteltieren. Dies schließt unter anderem Sadypus, Meteutatus und Barrancatatus aus der Gruppe der Eutatini sowie Parutaetus und Archaeutatus aus der Gruppe der Euphractini ein.[70] Aus dem Oberen Oligozän ist Kuntinaru überliefert, ein basales Mitglied der Tolypeutinae ohne spezielle Gruppenzuweisung, das mit Hilfe eines Schädelfundes aus den Salla Beds von Salla-Luribay in Bolivien in der Nähe von La Paz beschrieben werden konnte und rund 26 Millionen Jahre alt ist.[45] Mit Eocoleophorus ist weiterhin ein früher Repräsentant der Dasypodini im Tabauté-Becken Brasiliens entdeckt worden, so dass zu jener Zeit drei der vier Hauptlinien der Gürteltiere fassbar sind.[72][93]
Miozän

Für das Untere und Mittlere Miozän ist die sehr fossilreiche Santa-Cruz-Formation im südlichsten Teil Südamerikas von großer Bedeutung. Die dort damals vorherrschenden Landschaften unterlagen einem feuchten subtropischen Klima und bestanden aus einer Mixtur aus offenen Savannen und teils geschlossenen Wäldern, was eine formenreiche Gürteltierfauna hervorrief. Recht häufig belegt ist Stegotherium aus der Gruppe der Stegotheriini und deren einziger Vertreter, der hier einen seiner frühesten Nachweise hat, möglicherweise aber schon im Oligozän vorkam. Er ist unter anderem in mehreren Fundstellen nahe dem Lago Argentino aufgefunden worden, aber auch aus den küstennahen Fundgebieten entlang des Atlantiks dokumentiert. Die markant verlängerte Schnauze und die in ihrer Größe deutlich reduzierten Zähne zeichnen Stegotherium als eine stark spezialisierte Form aus, die überwiegend Ameisen und Termiten als Nahrungsgrundlage nutzte. Es handelt sich um den bisher am deutlichsten an diese Ernährungsweise angepassten Vertreter der Gürteltiere.[59][102] Die Gesteinseinheit barg darüber hinaus ein hervorragend erhaltenes Skelett von Prozaedyus, das etwa die Größe des heutigen Zwerggürteltiers erreichte. Auch Proeutatus und Stenotatus sind von hier bekannt, ersteres wurde dabei bis zu 15 kg schwer. Alle drei Vertreter gehören den Euphractinae an, wobei Prozaedyus die Euphractini repräsentiert, die beiden anderen aber die Eutatini. Hervorzuheben ist hierbei, dass es innerhalb der Euphractinae zu Beginn des Miozäns einen Austausch gab, da fast alle älteren Formen verschwanden und durch neue ersetzt wurden. Die Gattung Vetelia, von der einzelne Knochenplättchen vorliegen, repräsentierte ein großes Gürteltier vergleichbar mit dem heutigen Riesengürteltier. Ursprünglich ebenfalls zu den Euphractinae gezählt, verweist seine kurze und robuste Schnauze und die Ornamentierung der Osteoderme eher auf eine Beziehung zu den Tolypeutinae mit einer engeren Bindung an das erwähnte Riesengürteltier und die Nacktschwanzgürteltiere. Als lang andauernde Form ist Vetelia noch bis in das Obere Miozän präsent.[67][5][103][102]
Die aus der Santa-Cruz-Formation belegten Gürteltiere bilden auch Bestandteil der Faunen anderer, gleichalter Gesteinseinheiten in Südamerika, so der Chucal-Formation im nördlichen und der Río-Zeballos-Formation im südlichen Chile.[104][105] Sie sind dann noch bis ins Mittlere Miozän nachgewiesen, wo Funde aus den Cerdas Beds im südlichen Bolivien stammen.[61] Einige Formen des frühen Miozäns überlebten jedoch bis in jüngere Zeitphasen. Dies betrifft etwa Prozaedyus, das mit einem sehr kleinen Angehörigen aus der Loma-de-Las-Tapias-Formation im nordwestlichen Argentinien dokumentiert ist. Diese entstand im beginnenden Oberen Miozän. Der aufgefundene nahezu vollständige Schädel zeigt mit seiner kurzen Schnauze einen Allesfresser an.[106] Eine herausragende Fundstelle bildet La Venta am Mittellauf des Río Magdalena in Kolumbien. Die hier aufgeschlossene Honda-Gruppe umfasst mehrere Gesteinseinheiten, die in der Spätphase des Mittleren Miozäns entstanden waren. Mit Plesiodasypus, aufgefunden in der La-Victoria-Formation, tritt unter anderem ein erster früher Vertreter der Dasypodini auf. Belegt ist die Form über einen Schädel und mehrere Panzerfragmente. Das Alter der Funde beträgt etwa 13,3 Millionen Jahre.[77] Rund 12,5 Millionen Jahre alt und damit nur unwesentlich jünger ist Anadasypus aus der gleichen Verwandtschaftsgemeinschaft. Dessen Reste in Form eines Rückenpanzers wurden aus der auflagernden Villavieja-Formation zu Tage gefördert. Gleiches ist für Nanoastegotherium aus der Gruppe der Astegotheriini anzugeben, von dem weitgehend nur isolierte Knochenplättchen vorliegen.[68] Eine Besonderheit bilden Panzerreste und Teile des Körperskelettes von Pedrolypeutes. Dieser Vorläufer der Kugelgürteltiere kam noch in den obersten Abschnitten der La-Victoria-Formation zum Vorschein.[68] Somit ließ sich erstmals im nördlichen Südamerika ein Angehöriger der Tolypeutinae dokumentieren.[45]

Das ausgehende Mittlere und das Obere Miozän ist vor allem durch das Vorkommen zahlreicher neuer Mitglieder der Eutatini und Euphractini gekennzeichnet. So erscheinen hier Macroeuphractus, Proeuphractus und Chasicotatus. Alle drei Gattungen sind vom Conglomerado osifero nachgewiesen, einer äußerst fossilreichen Lagerstätte im Nordosten Argentiniens,[107] ebenso wie von der Cerro-Azul-Formation in der Pampa-Region. Die rund 5200 Gürteltierfragmente aus dieser Gesteinseinheit erbrachten zusätzlich Reste von Doellotatus, Ringueletia, Chorobates und Macrochorobates ebenso wie von Vetelia als Vertreter einer überlieferten älteren Linie.[108] Von Chasicotatus und Macrochorobates stammen einzelne Knochenplättchen auch von der Salicas-Formation im Nordwesten Argentiniens.[109] Auf Chasicotatus wiederum geht unter anderem ein nahezu vollständiger Rückenpanzer aus der Arroyo-Chasicó-Formation in der argentinischen Provinz Buenos Aires zurück.[110] Die gleiche Gesteinseinheit barg mit Chlamydophractus auch den bisher einzigen Nachweis eines fossilen Gürtelmulls, von dem neben Panzerfragmenten ein Unterkiefer und verschiedene Elemente des Bewegungsapparates überliefert sind.[63] Neben diesen stammesgeschichtlich jüngeren Formen sind zusätzlich noch einzelne ältere gefunden worden, so etwa Vetelia. Ihr können mehrere Unterkiefer zugesprochen werden.[67] Zudem gibt es aus der Arroyo-Chasicó-Formation einen der frühesten Hinweise auf Parasitismus von Flöhen an Gürteltieren. Dabei handelt es sich um Fraßspuren der Insekten, die sich im Panzer der damals lebenden Tiere einnisteten.[111] Die meisten Linien der frühen Gürteltiere starben weitgehend zum Ende des Miozäns oder im Verlauf des folgenden Pliozäns wieder aus.[45]
Plio- und Pleistozän

Im Pliozän schloss sich der Isthmus von Panama, wodurch eine Landbrücke nach Nordamerika entstand. Daraufhin setzte der Große Amerikanische Faunenaustausch ein. Zunächst trat im Pliozän weiterhin mit Macroeuphractus einer der größten Gürteltiervertreter auf, der die Größe eines heutigen Hausschweins erreichte. Dieser ist unter anderem aus dem nördlichen Argentinien nahe Buenos Aires nachgewiesen.[3] Zudem sind aus dieser Zeit die ersten Belege heutiger Gürteltiere bekannt. So war Tolypeutes mit der ausgestorbenen Art Tolypeutes pampaeus in der Pampa-Region noch bis in das ausgehende Unterpleistozän verbreitet,[112] in der gleichen Region entwickelte sich Chaetophractus mit dem Braunborsten-Gürteltier. Diese Gattung erreichte erst sehr spät ihr heute weit südliches Auftreten.[113][114] Auch der Ursprung von Dasypus fällt in das Pliozän, wobei der nördliche Teil Südamerikas aufgrund der Verbreitung zahlreicher nahe verwandter Gattungen dort zu jener Zeit als Herkunftsgebiet angenommen wird.[73] Zuerst entstand die fossilen Art Dasypus bellus, die bereits im ausgehenden Pliozän in Nordamerika anzutreffen war und dort recht häufig belegt ist, etwa in Florida, starb aber zum Ende des Pleistozäns dort wieder aus.[115] Dieses Schicksal traf auch zahlreiche andere Gürteltiervertreter in Südamerika wie Propraopus, einem nahen Verwandten von Dasypus, oder Eutatus aus der Linie der Eutatini, einem großen Tier, dass die Ausmaße des heutigen Riesengürteltiers erreichte.[116][64] Erst im 19. Jahrhundert erreichte Dasypus mit dem Neunbinden-Gürteltier wieder Nordamerika.[7]
Gürteltiere und Menschen
.jpg.webp)
In Südamerika werden Gürteltiere oft wegen ihres wohlschmeckenden Fleisches gejagt, was neben der Lebensraumvernichtung durch den Menschen bei einigen Arten bereits zur bedrohlichen Dezimierung geführt hat. Dagegen vermehrt sich beispielsweise das Neunbinden-Gürteltier beständig und breitet sich weiter nach Nordamerika aus. Mancherorts werden die Tiere zur Schädlingsbekämpfung angesiedelt oder als Heimtiere gehalten. Einige Arten gelten als Landplage; sie richten durch Wühlen nach Nahrung oder Anlegen von Erdbauen Schäden in landwirtschaftlich genutzten Gebieten an oder rufen bei großen Weidetieren Verletzungen hervor, wenn diese in die Baue einbrechen. Die Panzer der toten Tiere werden teils als skurrile Körbe an Touristen verkauft. Auch traditionelle Musikinstrumente wie die Charangos bestehen aus Panzerteilen einiger Gürteltierarten; deren Export aus den Herkunftsländern Bolivien und Peru ist jedoch verboten.[33]
Laut IUCN ist nur knapp die Hälfte aller Arten im Bestand nicht gefährdet. Als gefährdet gelten vor allem das Pelzgürteltier (Dasypus pilosus), das Riesengürteltier (Priodontes maximus) und das Nördliche Kugelgürteltier (Tolypeutes tricinctus). Drei weitere Arten – der Gürtelmull (Chlamyphorus retusus), der Burmeister-Gürtelmull (Calyptophractus truncatus) und das Yungas-Gürteltier (Dasypus mazzai) – können aufgrund fehlender Daten in ihrer Bestandsbedrohung nicht eingestuft werden.[33]
Neben dem Menschen sind Gürteltiere eine der wenigen Säugetiergruppen,[117] die das Bakterium der Leprakrankheit in sich tragen können. Ein Zusammenhang zwischen ihrer ungewöhnlich niedrigen Körpertemperatur und der Vermehrung der Mykobakterien gilt als wahrscheinlich. Das macht sie bei der Erforschung von Impfstoffen und neuen Antibiotikakombinationen unentbehrlich. Es bedeutet aber auch, dass Menschen, die Gürteltiere essen, Gefahr laufen, sich mit dieser Krankheit zu infizieren. Zumindest ein Teil der Leprafälle in den Vereinigten Staaten von Amerika wird auf den Kontakt zu Gürteltieren zurückgeführt.[118] Weiterhin sind einige Gürteltierarten auch Träger des Einzellers Trypanosoma cruzi, der die in Südamerika häufige Chagas-Krankheit verursacht und ebenfalls beim Menschen auftreten kann.[7]
Das Gürteltier Fuleco war Maskottchen der Fußball-Weltmeisterschaft 2014 in Brasilien. Der Name setzt sich aus den portugiesischen Wörtern futebol („Fußball“) und ecologia („Ökologie“) zusammen und sollte symbolisch für den Schutz der Gürteltiere stehen. Ein damit verbundener Aufruf von Wissenschaftlern zum besseren Erhalt der lokalen und überregionalen Biodiversität ist weitgehend verhallt.[119][120]
Literatur
- Alfred L. Gardner (Hrsg.): Mammals of South America, Volume 1: Marsupials, Xenarthrans, Shrews, and Bats. University of Chicago Press, 2008, S. 128–157, ISBN 0-226-28240-6
- C. M. McDonough und W. J. Laughry: Dasypodidae (Long-nosed armadillos). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos. Lynx Edicions, Barcelona 2018, S. 30–47 ISBN 978-84-16728-08-4
- Ronald M. Nowak: Walker’s Mammals of the World. Johns Hopkins University Press, 1999, ISBN 0-8018-5789-9
- Mariella Superina und Agustín Manuel Abba: Chlamyphoridae (Chlamyphorid armadillos). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos. Lynx Edicions, Barcelona 2018, S. 48–71 ISBN 978-84-16728-08-4
- Sergio F. Vizcaíno und W. J. Loughry (Hrsg.): The Biology of the Xenarthra. University Press of Florida, 2008, S. 1–370, ISBN 978-0-8130-3165-1
Einzelnachweise
- C. M. McDonough und W. J. Laughry: Dasypodidae (Long-nosed armadillos). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos. Lynx Edicions, Barcelona 2018, S. 30–47 ISBN 978-84-16728-08-4
- Mariella Superina und Agustín Manuel Abba: Chlamyphoridae (Chlamyphorid armadillos). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos. Lynx Edicions, Barcelona 2018, S. 48–71 ISBN 978-84-16728-08-4
- Sergio E Vizcaíno und Gerardo De Iuliis: Evidence for advanced carnivory in fossil armadillos (Mammalia: Xenarthra: Dasypodidae). In: Paleobiology, 29 (1), 2003, S. 123–138
- Darin A. Croft und Velizar Simeonovski: Horned armadillos and rafting monkeys. The fascinating fossil mammals of South America. Indiana University Press, 2016, S. 1–304 (S. 56–57 und 208–209)
- Sergio F. Vizcaíno, M. Susana Bargo, Richard F. Kay und Nick Milne: The armadillos (Mammalia, Xenarthra, Dasypodidae) of the Santa Cruz Formation (early–middle Miocene): An approach to their paleobiology. In: Palaeogeography, Palaeoclimatology, Palaeoecology 237, 2006, S. 255–269
- Paul Smith: The Xenarthra famalies Myrmecophagidae and Dasypodidae. Fauna Paraguay Handbook of the Mammals of Paraguay 2012, S. 1–35
- Mariella Superina: Biologie und Haltung von Gürteltieren (Dasypodidae). Universität Zürich, 2000, S. 1–248
- Sergio F. Vizcaíno: The teeth of the “toothless”: novelties and key innovations in the evolution of xenarthrans (Mammalia, Xenarthra). In: Paleobiology 35 (3), 2009; S. 343–366
- Gerardo De Iuliis, María S. Bargo und Sergio F. Vizcaíno: Variation in skull morphology and mastication in the fossil giant armadillos Pampatherium spp. and allied genera (Mammalia: Xenarthra: Pampatheriidae), with comments on their systematics and distribution. In: Journal of Vertebrate Paleontology 20 (4), 2000, S. 743–754
- Daniela C. Kalthoff: Microstructure of Dental Hard Tissues in Fossil and Recent Xenarthrans (Mammalia: Folivora and Cingulata). In: Journal of Morphology 272, 2011, S. 641–661
- Sergio F. Vizcaíno und María S. Bargo: Los armadillos (Mammalia, Dasypodidae) de La Toma (Partido de Coronel Pringles) y otros sitios arqueológicos de la provincia de Buenos Aires. Consideraciones paleoambientales. In: Ameghiniana 30, 1993, S. 435–443
- Alfredo A. Carlini, Martín Ricardo Ciancio, John J. Flynn, Gustavo J. Scillato‐Yané und André R. Wyss: The phylogenetic and biostratigraphic significance of new armadillos (Mammalia, Xenarthra, Dasypodidae, Euphractinae) from the Tinguirirican (early oligocene) of Chile. In: Journal of Systematic Palaeontology 7 (4), 2009, S. 489–503
- Matthew K. Vickaryous und Brian K. Hall: Osteoderm Morphology and Development in the Nine-Banded Armadillo, Dasypus novemcinctus (Mammalia, Xenarthra, Cingulata). In: Journal of Morphology 267 (11), 2006, S. 1273–1283
- Robert V. Hill: Comparative Anatomy and Histology of Xenarthran Osteoderms. In: Journal of Morphology 267, 2006, S. 1441–1460
- C. M. Krmpotic, M. R. Ciancio, C. Barbeito, R. C. Mario und A. A. Carlini: Osteodermmorphology in recent and fossil euphractine xenarthrans. In: Acta Zoologica (Stockholm) 90, 2009, S. 339–351
- C. M. Krmpotic, M. R. Ciancio, A. A. Carlini, M. C. Castro, A. C. Scarano und C. G. Barbeito: Comparative histology and ontogenetic change in the carapace of armadillos (Mammalia: Dasypodidae). In: Zoomorphology 134, 2015, S. 601–616
- M. R. Ciancio, C. M. Krmpotic, A. C. Scarano und M. B. Epele: Internal Morphology of Osteoderms of Extinct Armadillos and Its Relationship with Environmental Conditions. In: Journal of Mammalian Evolution 26 (1), 2019, S. 71–83, doi:10.1007/s10914-017-9404-y
- Irene H. Chen, James H. Kiang, Victor Correa, Maria I. Lopez, Po-Yu Chen, Joanna McKittrick und Marc A. Meyers: Armadillo armor: Mechanical testing and micro-structural evaluation. In: Journal of the Mechanical Behavior of Biomedical Materials 4 (5), 2011, S. 713–722
- Cecilia M. Krmpotic, Fernando C. Galliari, Claudio G. Barbeito und Alfredo A. Carlini: Development of the integument of Dasypus hybridus and Chaetophractus vellerosus, and asynchronous events with respect to the postcranium. In: Mammalian Biology 77, 2012, S. 314–326
- Mariella Superina und W. J. Loughry: Life on the Half-Shell: Consequences of a Carapace in the Evolution of Armadillos (Xenarthra: Cingulata). In: Journal of Mammalian Evolution 19, 2012, S. 217–224
- Timothy J. Gaudin: The morphology of the xenarthrous vertebrae (Mammalia: Xenarthra). In: Fieldiana 1505 (41), 1999, S. 1–38
- Fernando C. Galliari, Alfredo A. Carlini, Marcelo R. Sánchez-Villagra: Evolution of the axial skeleton in armadillos (Mammalia, Dasypodidae). In: Zeitschrift für Säugetierkunde 75, 2010, S. 326–333
- S. F. Vizcaíno und N. Milne: Structure and function in armadillo limbs (Mammalia: Xenarthra: Dasypodidae). In: Journal of Zoology 257, 2002, S. 117–127
- N. Milne, S. F. Vizcaíno und J. C. Fernicola. A 3D geometric morphometric analysis of digging ability in the extant and fossil cingulate humerus. In: Journal of Zoology 278 (1), 2009, S. 48–56
- Sergio F. Vizcaíno, Richard A. Fariña und Gerardo V. Mazzetta: Ulnar dimensions and fossoriality in armadillos. In: Acta Theriologica 44 (3), 199, S. 309–320
- Nick Milne, Nestor Toledo und Sergio F. Vizcaíno: Allometric and Group Differences in the Xenarthran Femur. In: Journal of Mammalian Evolution 19, 2012, S. 199–208
- M. Ciuccio, S. Estecondo und E. B. Casanave: Scanning Electron Microscopy Study of the Dorsal Surface of the Tongue in Zaedyus pichiy (Mammalia, Xenarthra, Dasypodidae). In: International Journal of Morphology 26 (1), 2008. S. 13–18
- Silvia Estecondo, Stella Maris Codón und Emma Beatriz Casanave: Histological Study of the Salivary Glands in Zaedyus pichiy (Mammalia, Xenarthra, Dasypodidae). In: International Journal of Morphology 23 (1), 2005. S. 19–24
- Sergio F. Vizcaíno, M. Susana Bargo und Guillermo H. Cassini: Dental occlusal surface area in relation to body mass, food habits and other biological features in fossil xenarthrans. In: Ameghiniana 43 (1), 2006, S. 11–26
- E. J. Galíndez, S. Estecondo und E. B. Casanave: The Spleen of Zaedyus pichiy, (Mammalia, Dasypodidae): a Light and Electron Microscopic Study. In: Anatomy, Histology, Embryology 32, 2003, S. 194–199
- Kurt Benirschke: Reproductive parameters and placentation in anteaters and sloths. In: Sergio F. Vizcaíno und W. J. Loughry (Hrsg.): The Biology of the Xenarthra. University Press of Florida, Gainesville, 2008, S. 160–171
- Allen C. Enders: Placentation in armadillos, with emphasis on development of the placenta in polyembryonic species. In: Sergio F. Vizcaíno und W. J. Loughry (Hrsg.): The Biology of the Xenarthra. University Press of Florida, Gainesville, 2008, S. 172–180
- Agustín Manuel Abba und Mariella Superina: The 2009/2010 Armadillo Red List Assessment. In: Edentata 11 (2), 2010, S. 135–184
- Sergio F. Vizcaíno und María S. Bargo: The masticatory apparatus of the armadillo Eutatus (Mammalia, Cingulata) and some allied genera: paleobiology and evolution. In: Paleobiology 24 (3), 1998, S. 371–383
- Barry Gordon Lovegrove: The Zoogeography of Mammalian Basal Metabolic Rate. In: The American Naturalist 156 (2), 2000, S. 201–219
- Brian K. McNab: Physiological convergence among ant-eating and termite-eating mammals. In: Journal of Zoology 203 (4), 1984, S. 485–510
- Brian K. McNab: Energetics, population biology, and distribution of Xenarthrans, living and extinct. In: G. Gene Montgomery (Hrsg.): The evolution and ecology of armadillos, sloths, and vermilinguas. Smithonian Institution Press, 1985, S. 219–232
- D. F. Boggs, P. B. Frappell und D. L. Kilgore Jr.: Ventilatory, cardiovascular and metabolic responses to hypoxia and hypercapnia in the armadillo. In: Respiration Physiology 113 (2), 1998, S. 101–109
- Dennis A. Meritt Jr: Xenarthrans of the Paraguayan Chaco. In: Sergio F. Vizcaíno SF und W. J. Loughry (Hrsg.): The Biology of the Xenarthra. University Press of Florida, 2008, S. 294–299
- Brian K. McNab: Energetics and the limits to a temperate distribution of armadillos. In: Journal of Mammalogy 61 (4), 1980, S. 606–627
- G. J. Tattersall und V. Cadena: Insights into animal temperature adaptations revealed through thermal imaging. In: The Imaging Science Journal 58, 2010, S. 263–268
- Barry Gordon Lovegrove: The evolution of body armor in mammals: Plantigrade constraints of large body size. In: Evolution 55 (7), 2001, S. 1464–1473
- Richard D. Peppler: Reproductive biology of the nine-banded armadillo. In: Sergio F. Vizcaíno und W. J. Loughry (Hrsg.): The Biology of the Xenarthra. University Press of Florida, Gainesville, 2008, S. 151–159
- Thiago F. Rodrigues, Aline M. B. Mantellatto, Mariella Superina und Adriano G. Chiarello: Ecosystem services provided by armadillos. In: Mammal Review 95 (1), 2020, S. 1–21, doi:10.1111/brv.12551
- Guillaume Billet, Lionel Hautier, Christian de Muizon und Xavier Valentin: Oldest cingulate skulls provide congruence between morphological and molecular scenarios of armadillo evolution. In: Proceedings of the Royal Society B 278, 2011, S. 2791–2797
- Frédéric Delsuc, Sergio F. Vizcaíno und Emmanuel J. P. Douzery: Influence of Tertiary paleoenvironmental changes on the diversification of South American mammals: a relaxed molecular clock study within xenarthrans. In: BMC Evolutionary Biology 4 (11), 2004, S. 1–13
- Maureen A. O’Leary, Jonathan I. Bloch, John J. Flynn, Timothy J. Gaudin, Andres Giallombardo, Norberto P. Giannini, Suzann L. Goldberg, Brian P. Kraatz, Zhe-Xi Luo, Jin Meng, Xijun Ni, Michael J. Novacek, Fernando A. Perini, Zachary S. Randall, Guillermo W. Rougier, Eric J. Sargis, Mary T. Silcox, Nancy B. Simmons, Michelle Spaulding, Paúl M. Velazco, Marcelo Weksler, John R. Wible und Andrea L. Cirranello: The Placental Mammal Ancestor and the Post–K-Pg Radiation of Placentals. In: Science 339, 2013, S. 662–667
- Kenneth D. Rose: The beginning of the age of mammals. Johns Hopkins University Press, Baltimore, 2006, S. 1–431 (S. 200–204)
- George Gaylord Simpson: The Principles of Classification and a Classification of Mammals. In: Bulletin of the American Museum of Natural History 85, 1945, S. 1–350 (S. 75 und 194–195)
- Timothy J. Gaudin und John R. Wible: The Phylogeny of Living and Extinct Armadillos (Mammalia, Xenarthra, Cingulata): A Craniodental Analysis. In: M. T. Carrano, T. J. Gaudin, R. W. Blob und J. R. Wible (Hrsg.): Amniote Paleobiology. Chicago/London: University of Chicago Press, 2006, S. 153–198
- Leopold Héctor Soibelzon, Martín Zamorano und Gustavo Juan Scillato-Yané: Un Glyptodontidae de gran tamaño en el Holoceno Temprano de la Region Pampeana, Argentina. In: Revista Brasileira de Paleontologia 15 (1), 2012, S. 105–112
- Frédéric Delsuc, Gillian C. Gibb, Melanie Kuch, Guillaume Billet, Lionel Hautier, John Southon, Jean-Marie Rouillard, Juan Carlos Fernicola, Sergio F. Vizcaíno, Ross D. E. MacPhee und Hendrik N. Poinar: The phylogenetic affinities of the extinct glyptodonts. In: Current Biology 26, 2016, S. R155–R156, doi:10.1016/j.cub.2016.01.039
- Kieren J. Mitchell, Agustin Scanferla, Esteban Soibelzon, Ricardo Bonini, Javier Ochoa und Alan Cooper: Ancient DNA from the extinct South American giant glyptodont Doedicurus sp. (Xenarthra: Glyptodontidae) reveals that glyptodonts evolved from Eocene armadillos. In: Molecular Ecology 25 (14), 2016, S. 3499–3508, doi:10.1111/mec.13695
- Juan C. Fernicola, Andrés Rinderknecht, Washington Jones, Sergio F. Vizcaíno und Kleberson Porpino: A new species of Neoglyptatelus(Mammalia, Xenarthra, Cingulata) from the late Miocene of Uruguay provides new insights on the evolution of the dorsal armor in cingulates. In: Ameghiniana 55, 2018, S. 233–252
- Gillian C. Gibb, Fabien L. Condamine, Melanie Kuch, Jacob Enk, Nadia Moraes-Barros, Mariella Superina, Hendrik N. Poinar und Frédéric Delsuc: Shotgun Mitogenomics Provides a Reference Phylogenetic Framework and Timescale for Living Xenarthrans. In: Molecular Biology and Evolution 33 (3), 2015, S. 621–642
- Frédéric Delsuc, Michael J. Stanhope und Emmanuel J. P. Douzery: Molecular systematics of armadillos (Xenarthra, Dasypodidae): contribution of maximum likelihood and Bayesian analyses of mitochondrial and nuclear genes. In: Molecular Phylogenetics and Evolution 28, 2003, S. 261–275
- Maren Möller-Krull, Frédéric Delsuc, Gennady Churakov, Claudia Marker, Mariella Superina, Jürgen Brosius, Emmanuel J. P. Douzery und Jürgen Schmitz: Retroposed Elements and Their Flanking Regions Resolve the Evolutionary History of Xenarthran Mammals (Armadillos, Anteaters and Sloths). In: Molecular Biology and Evolution 24, 2007, S. 2573–2582
- Frédéric Delsuc, Mariella Superina, Marie-Ka Tilak, Emmanuel J. P. Douzery und Alexandre Hassanin: Molecular phylogenetics unveils the ancient evolutionary origins of the enigmatic fairy armadillos. In: Molecular Phylogenetics and Evolution 62, 2012, 673–680
- Laureano Raúl González Ruiz und Gustavo Juan Scillato-Yané: A new Stegotheriini (Mammalia, Xenarthra, Dasypodidae) from the “Notohippidian” (early Miocene) of Patagonia, Argentina. In: Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 252 (1), 2009, S. 81–90
- Martín Ricardo Ciancio, Alfredo A. Carlini, Kenneth E. Campbell und Gustavo J. Scillato-Yané: New Palaeogene cingulates (Mammalia, Xenarthra) from Santa Rosa, Perú and their importance in the context of South American faunas In: Journal of Systematik Palaeontology 11 (5-6), 2013, S. 727–741 doi:10.1080/14772019.2012.704949
- Darin Andrew Croft, Federico Anaya, David Auerbach, Carmala Garzione und Bruce J. MacFadden: New Data on Miocene Neotropical Provinciality from Cerdas, Bolivia. In: Journal of Mammalian Evolution 16 (3), 2009, S. 175–198
- Gustavo Juan Scillato-Yané, Cecilia Mariana Krmpotic und Graciela Irene Esteban: The species of genus Chasicotatus Scillato-Yané (Eutatini, Dasypodidae). In: Revista Mexicana de Ciencias Geológicas 27 (1), 2010, S. 43–55
- Daniel Barasoain, Rodrigo L. Tomassini, Alfredo E. Zurita, Claudia I. Montalvo und Mariella Superina: A new fairy armadillo (Cingulata, Chlamyphorinae) from the upper Miocene of Argentina: first fossil record of the most enigmatic Xenarthra. In: Journal of Vertebrate Paleontology 39 (5), 2019, S. e1716778, doi:10.1080/02724634.2019.1716778; Daniel Barasoain, Rodrigo L. Tomassini, Alfredo E. Zurita, Claudia I. Montalvo und Mariella Superina: Chlamydophractus, new name for Chlamyphractus Barasoain et al., 2020 (Xenarthra, Chlamyphorinae), non Chlamyphractus Castellanos, 1940 (Xenarthra, Glyptodontidae). In: Journal of Vertebrate Paleontology, 2020, S. e1774890, doi:10.1080/02724634.2020.1774890 (Namenskorrektur)
- C. M. Krmpotic, A. Carlini und G. J. Scillato-Yané: The species of Eutatus (Mammalia, Xenarthra): Assessment, morphology and climate. In: Quaternary International 210, 2009, S. 66–75
- Malcolm C. McKenna und Susan K. Bell: Classification of mammals above the species level. Columbia University Press, New York, 1997, S. 1–631 (S. 82–86)
- M. C. Castro: Sistemática y evolución de los armadillos Dasypodini (Xenarthra, Cingulata, Dasypodidae). In: Revista del Museo de La Plata, Sección Paleontología 15 (73), 2015, S. 1–50
- Daniel Barasoain, Laureano R. González Ruiz, Rodrigo L. Tomassini, Alfredo Zurita, Víctor H. Contreras und Claudia I. Montalvo: First phylogenetic analysis of the Miocene armadillo Vetelia reveals novel affinities with Tolypeutinae. In: Acta Palaeontologica Polonica 66 (3), 2021, S. s031–s046, doi:10.4202/app.00829.2020
- Alfredo A. Carlini, Sergio F. Vizcaíno und Gustavo J. Scillato-Yané: Armored Xenarthrans: A unique taxonomic and ecologic assemblage. In: Richard F. Kay, Richard H. Madden, Richard L. Cifelli und John J. Flynn (Hrsg.): Vertebrate Paleontology in the Neotropics. The Miocene Fauna of La Venta, Colombia. Smithsonian Institution Press, Washington, 1997, S. 213–226
- Lílian Paglarelli Bergqvist, Érika Aparecida Leite Abrantes und Leonardo dos Santos Avilla: The Xenarthra (Mammalia) of São José de Itaboraí Basin (upper Paleocene, Itaboraian), Rio de Janeiro, Brazil. In: Geodiversitas 26 (2), 2004, S. 323–337
- Alfredo A. Carlini, Martín R. Ciancio und Gustavo J. Scillato-Yané: Middle Eocene – Early Miocene Dasypodidae (Xenarthra) of Southern South America: faunal succession at Gran Barranca – biostratigraphy and paleoecology. In: Richard H. Madden, Alfredo A. Carlini, Maria Guiomar Vucetich und Richard F. Kay (Hrsg.): The Paleontology of Gran Barranca. Evolution and environmental change through the Middle Cenozoic of Patagonia. Cambridge University Press, 2010, S. 106–129
- Claudia M. Herrera, Jaime E. Powell und Cecilia del Papa: Un nuevo Dasypodidae (Mammalia, Xenarthra) de la Formación Casa Grande (Eoceno) de la Provincia de Jujuy, Argentina. In: Ameghiniana 49 (2), 2012, S. 267–271
- Mariela C. Castro, Ana Maria Ribeiro, Jorge Ferigolo und Max C. Langer: Redescription of Dasypus punctatus Lund, 1840 and considerations on the genus Propraopus Ameghino, 1881 (Xenarthra, Cingulata). In: Journal of Vertebrate Paleontology 33 (2), 2013, S. 434–447
- Mariela C. Castro, Alfredo A. Carlini, Rodolfo Sánchez und Marcelo R. Sánchez-Villagra: A new Dasypodini armadillo (Xenarthra: Cingulata) from San Gregorio Formation, Pliocene of Venezuela: affinities and biogeographic interpretations. In: Naturwissenschaften 101, 2014, S. 77–86
- Martín R. Ciancio, Claudia Herrera, Alejandro Aramayo, Patricio Payrola und María J. Babot: Diversity of cingulate xenarthrans in the middle-late Eocene of Northwestern Argentina. In: Acta Palaeontologica Polonica 61 (3), 2016, S. 575–590, doi:10.4202/app.00208.2015
- Claudia M. R. Herrera, Jaime E. Powell, Graciela I. Esteban und Cecilia del Papa: A New Eocene Dasypodid with Caniniforms (Mammalia, Xenarthra, Cingulata) from Northwest Argentina. In: Journal of Mammalian Evolution 24 (3), 2017, S. 275–288, doi:10.1007/s10914-016-9345-x
- Fernando A. Sedor, Édison V. Oliveira, David D. Silva, Luiz A. Fernandes, Renata F. Cunha, Ana M. Ribeiro und Eliseu V. Dias: A New South American Paleogene Land Mammal Fauna, Guabirotuba Formation (Southern Brazil). In: Journal of Mammalian Evolution 24, 2017, S. 39–55
- Daniel Barasoain, Laureano González-Ruiz, Alfredo Zurita und Carlos Villarroel: Oldest new Dasypodini (Xenarthra, Cingulata) provides new trails about armadillos evolutionary history. In: Historical Biology: An International Journal of Paleobiology, 2021, doi:10.1080/08912963.2021.1917566
- Juan Carlos Fernicola, Ana N. Zimicz, Laura Chornogubsky, Mihai Ducea, Laura E. Cruz, Mariano Bond, Michelle Arnal, Magalí Cárdenas und Mercedes Fernández: The Early Eocene Climatic Optimum at the Lower Section of the Lumbrera Formation (Ypresian, Salta Province, Northwestern Argentina): Origin and Early Diversification of the Cingulata. In: Journal of Mammalian Evolution 28 (3), 2021, S. 621–633, doi:10.1007/s10914-021-09545-w
- Juan Carlos Fernicola, Ana Natalia Zimicz, Laura Chornogubsky, Laura Edith Cruz, Mariano Bond, Michelle Arnal, Magalí Cárdenas und Mercedes Fernández: New assemblage of cingulates from the Quebrada de Los Colorados Formation (middle Eocene) at Los Cardones National Park (Salta Province, Argentina) and the Casamayoran SALMA problem at the Northwestern Argentina. In: Journal of South American Earth Sciences 111, 2021, S. 103476, doi:10.1016/j.jsames.2021.103476
- J. M. Anderson und K. Benirschke: The armadillo, Dasypus novemcinctus, in experimental biology. In: Laboratory Animal Care 16, 1966, S. 202–216
- Pierre Belon: Les observations de plusieurs singularites & choses memorables, trouvees en Grece, Asie, Judee, Égypte, Arabie, & autres pays etranges, redigees en trois livres. Revues derechef, & augmentees de figures, avec une nouvelle table de toutes les matieres traitees en iceux. Paris, 1553, S. 1–212 (S. 211) ()
- Pierre Belon: Les Observations De Plusieurs Singularites et Choses Memorables Trouvée En Grèce, Asie, Judée, Egypte, Arabie et Autres Pays Estrangèrs, Redigées Trois Livres. Paris, 1588, S. 1–468 (S. 467) ()
- Georges-Louis Leclerc de Buffon: Histoire naturelle, générale et particulière., Tome Dixième. Paris, 1763, S. 200–268 ()
- Wilhelm Pape: Handwörterbuch der griechischen Sprache 1. Braunschweig, 1880, S. 524 ()
- Oldfield Thomas: The mammals of the tenth edition of Linnaeus; an attempt to fix the types of the genera and the exact bases and localities of the species. In: Proceedings of the Zoological Society of London 1911, S. 120–158
- Karen McBee und Robert J. Baker: Dasypus novemcinctus. In: Mammalian Species 162, 1982, S. 1–9
- Carl von Linné: Systema Naturae. 10. Auflage, Stockholm, 1758, 1–824 (S. 50–51) ()
- John Edward Gray: On the natural arrangement of vertebrose animals. London Medical Repository 15, 1821, S. 297–310 (305) (; PDF; 898 kB)
- Friedrich August Quenstedt: Handbuch der Petrefaktenkunde. Tübingen, 1885, S. 1–1239 (S. 62) ()
- Rudolf Hoernes: Manuel de paléontologie. Paris, 1886, S. 1–741 (S. 662) ()
- Rudolf Hoernes: Elemente der Paläontologie. Paris, 1884, S. 1–594 (S. 527) ()
- Bruce J. Shockey: New Early Diverging Cingulate (Xenarthra: Peltephilidae) from the Late Oligocene of Bolivia and Considerations Regarding the Origin of Crown Xenarthra. In: Bulletin of the Peabody Museum of Natural History 58 (2), 2017, S. 371–396
- Martín Ricardo Ciancio. Los armadillos (Dasypodidae, Xenarthra) del Cenozoico temprano-medio de Argentina: Aspectos evolutivos, bioestratigráficos y biogeográficos. In: Federico L. Agnolin, Gabriel L. Lio, Federico Brissón Egli, Nicolas R. Chimento und Fernando E. Novas (Hrsg.): Historia evolutiva y paleobiográfica de los vertebrados de América del Sur. Buenos Aires, 2016, S, 231–247
- Martín R. Ciancio, Emma C. Vieytes und Alfredo A. Carlini: When xenarthrans had enamel: insights on the evolution of their hypsodonty and paleontological support for independent evolution in armadillos. In: Naturwissenschaften 101, 2014, S. 715–725
- Édison Vicente Oliveira und Lílian Paglarelli Bergqvist: A new Paleogene armadillo (Mammalia, Dasypodoidea) from the Itaboraí basin, Brazil. In: Associatión Paleontológica Argentina, Publicatión Especial 5, 1998, S. 35–40
- Richard L. Cifelli: Eutherian Tarsals from the Late Paleocene of Brazil. In: American Museum Novitates 2761, 1983, S. 1–31
- Lílian P. Bergqvist, Paulo Victor Luiz G. C. Pereira, Alessandra S. Machado, Mariela C. de Castro, Luiza B. Melki und Ricardo T. Lopez: Osteoderm microstructure of Riostegotherium yanei, the oldest Xenarthra. In: Anais da Academia Brasileira de Ciências 91 (Suppl. 2), 2019, S. e20181290, doi:10.1590/0001-3765201920181290
- Marcelo F. Tejedor, Francisco J. Goin, Javier N. Gelfo, Guillermo López, Mariano Bond, Alfredo A. Carlini, Gustave J. Scillato-Yane, Michael O. Woodburne, Laura Chornogubsky, Eugenio Aragón, Marcelo A. Reguero, Nicholas J. Czapkewski, Sergio Vincon, Gabriel M. Martin und Martin R. Ciancio: New Early Eocene Mammalian Fauna from Western Patagonia, Argentina. In: American Museum Novitates 3638, 2009, S. 1–43
- Fernando A. Sedor, Tabata D. F. Klimeck, Eliseu V. Dias, Edison V. Oliveira, Martín R Ciancio,Kimberlym T. P. Vieira, Luiz A.Fernandes und Rodolfo J. Angulo: The Eocene armadillo Utaetus buccatus (EUPHRACTINAE) in the guabirotuba formation (Curitiba basin) and carapace morphological implications. In: Journal of South American Earth Sciences, 2022, S. 103694, doi:10.1016/j.jsames.2021.103694
- Claudia M. R. Herrera, Graciela I. Esteban, Martín R. Ciancio und Cecilia Del Papa: New specimen of Pucatherium parvum (Xenarthra, Dasypodidae), a singular dasypodid of the Paleogene (Eocene) of northwest Argentina: importance in the early evolution of armadillos. In: Journal of Vertebrate Paleontology 39 (4), 2019, S. 1670669, doi:10.1080/02724634.2019.1670669
- François Pujos, Martin R. Ciancio, Analía M. Forasiepi, Michel Pujos, Adriana M. Candela, Bárbara Vera, Marcelo A. Reguero, Ana María Combina und Esperanza Cerdeño: The late Oligocene xenarthran fauna of Quebrada Fiera (Mendoza, Argentina) and its implications for sloth origins and the diversity of Palaeogene cingulates. In: Papers in Paleontology 7 (3), 2021, S. 1613–1656, doi:10.1002/spp2.1356
- Sergio F. Vizcaíno, Juan C. Fernicola und M. Susana Bargo: Paleobiology of Santacrucian glyptodonts and armadillos (Xenarthra, Cingulata). In: Sergio F. Vizcaíno, Richard F. Kay und M. Susana Bargo (Hrsg.): Early Miocene paleobiology in Patagonia: High-latitude paleocommunities of the Santa Cruz Formation. Cambridge University Press, New York, 2012, S. 194–215
- Sergio F. Vizcaíno, M. Susana Bargo, Richard F. Kay, Richard A. Fariña, Mariana Di Giacomo, Jonathan M. G. Perry, Francisco J. Prevosti, Néstor Toledo, Guillermo H. Cassini und Juan C. Fernicola: A baseline paleoecological study for the Santa Cruz Formation (late–early Miocene) at the Atlantic coast of Patagonia, Argentina. In: Palaeogeography, Palaeoclimatology, Palaeoecology 292, 2010, S. 507–519
- John J. Flynn, Michael J. Novacek, Holly E. Dodson, Daniel Frassinetti, Malcolm C. McKenna, Mark A. Norell, Karen E. Sears, Carl C. Swisher III und André R. Wyss: A new fossil mammal assemblage from the southern Chilean Andes: implications for geology, geochronology, and tectonics. In: Journal of South American Earth Sciences 15, 2002, S. 285–302
- Darin A. Croft, John J. Flynn und André R. Wyss: A new basal glyptodontid and other Xenarthra of the Early Miocene Chucal fauna, Northern Chile. In: Journal of Vertebrate Paleontology 27 (4), 2007, S. 781–797
- Daniel Barasoain, Víctor H. Contreras, Rodrigo L. Tomassini und Alfredo E. Zurita: A new pygmy armadillo (Cingulata, Euphractinae) from the late Miocene of Andean Argentina reveals an unexpected evolutionary history of the singular Prozaedyus lineage. In: Journal of South American Earth Sciences 100, 2020, S. 102589, doi:10.1016/j.jsames.2020.102589
- Gustavo Juan Scillato-Yané, Flávio Góis, Alfredo E. Zurita, Alfredo A. Carlini, Laureano R. González Ruiz, Cecilia M. Krmpotic, Cristian Oliva und Martín Zamorano: Los Cingulata (Mammalia, Xenarthra) del “Conglomerado osífero” (Mioceno Tardío) de la Formación Ituzaingó de Entre Ríos, Argentina. In: Diego. Brandoni und J. I. Noriega (Hrsg.): El Neógeno de la Mesopotamia argentina. Asociación Paleontológica Argentina, Publicación Especial 14, 2013, S. 118–134
- Juan J. Urrutia, Claudia I. Montalvo und Gustavo J. Scillato-Yané: Dasypodidae (Xenarthra, Cingulata) de la Formación Cerro Azul (Mioceno tardío) de la provincia de La Pampa, Argentina. In: Ameghiniana 45 (2), 2008, S. 289–302
- Diego Brandoni, Gabriela I. Schmidt, Adriana M. Candela, Jorge I. Noriega, Ernesto Brunetto und Lucas E. Fiorélli: Mammals from the Salicas Formation (Late Miocene), La Rioja Province, Northwestern Argentina: Paleobiogeography, age, and paleoenvironments. Ameghiniana 49 (3), 2012, S. 375–387
- Gustavo Juan Scillato-Yané, Cecilia Krmpotic und Graciela Irene Esteban: The species of the genus Chasicotatus Scillato-Yané (Eutatini, Dasypodidae). In: Revista Mexicana de Ciencias Geológicas 27 (1), 2010, S. 43–55
- Rodrigo L. Tomassini, Claudia I. Montalvo und María C. Ezquiaga: The oldest record of flea/armadillos interaction as example of bioerosion on osteoderms from the late Miocene of the Argentine Pampas. In: International Journal of Paleopathology 15, 2016, S. 65–68, doi:10.1016/j.ijpp.2016.08.004
- Paulina E. Nabel, Alberto Cione und Eduardo P. Tonni: Environmental changes in the Pampean area of Argentina at the Matuyama–Brunhes (C1r–C1n) Chrons boundary. In: Palaeogeography, Palaeoclimatology, Palaeoecology 162, 2000, S. 403–412
- E. Soibelzon, A. A. Carlini, E. P. Tonni und L. H. Soibelzon: Chaetophractus vellerosus (Mammalia: Dasypodidae) in the Ensenadan (Early-Middle Pleistocene) of the Southeastern Pampean Region (Argentina) – Paleozoogeographical and Paleoclimatic Aspects. In: Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 12, 2006, S. 734–748
- Sebastián Poljak, Viviana Confalonieri, Mariana Fasanella, Magalí Gabrielli und Marta Susana Lizarralde: Phylogeography of the armadillo Chaetophractus villosus (Dasypodidae Xenarthra): Post-glacial range expansion from Pampas to Patagonia (Argentina). In: Molecular Phylogenetics and Evolution 55 (1), S. 38–46
- H. Gregory McDonald: Paleoecology of extinct Xenarthans and the Great American Biotic Interchange. In: Bulletin of the Florida Museum of Natural History 45 (4), 2005, S. 313–333
- Ascanio D. Rincón, Richard S. White und H. Gregory Mcdonald: Late Pleistocene Cingulates (Mammalia: Xenarthra) from Mene De Inciarte Tar Pits, Sierra De Perijá, Western Venezuela. In: Journal of Vertebrate Paleontology 28 (1), 2008, S. 197–207
- Herbert Hof, Rüdiger Dörries und Robert L. Müller: Mikrobiologie. Georg Thieme Verlag Stuttgart, 2000, S. 1–640, S. 342 ff., ISBN 3-13-125311-8
- Richard W. Truman, Pushpendra Singh, Rahul Sharma, Philippe Busso, Jacques Rougemont, Alberto Paniz-Mondolfi, Adamandia Kapopoulou, Sylvain Brisse, David M. Scollard, Thomas P. Gillis und Stewart T. Cole: Probable Zoonotic Leprosy in the Southern United States. In: The New England Journal of Medicine 364 (17), 2011, S. 1626–1633
- Felipe P. Melo, José A. Siqueira, Bráulio A. Santos, Orione Álvares-da-Silva, Gerardo Ceballos und Enrico Bernard: Football and Biodiversity Conservation: FIFA and Brazil Can Still Hit a Green Goal. In: Biotropica 46 (3), 2014, S. 257–259
- Enrico Bernard und Felipe P. L. Melo: Fuleco™ revisited: Football, conservation and lessons learned from the 2014 FIFA World Cup. In: Biotropica 51 (4), 2019, S. 473–476
Weblinks
- Armadillo Online – englischsprachige Seite, u. a. mit Artbeschreibungen und Fotos
- Katharina Hübel: Das Gürteltier – Gepanzerter Sonderling und Superheld des Tierreichs Bayern 2 Radiowissen, Ausstrahlung am 15. Januar 2021 (Podcast)