Fadenwürmer
Die Fadenwürmer (Nematoda), auch Nematoden (altgriechisch νῆμα nema, deutsch ‚Faden‘) oder Älchen genannt, sind ein sehr artenreicher Stamm des Tierreichs. Bislang wurden mehr als 20.000 verschiedene Arten beschrieben, Schätzungen gehen aber von insgesamt 100.000 bis 10 Millionen Arten aus.[1] Überdies sind die Nematoden wahrscheinlich die individuenreichste Gruppe unter den vielzelligen Tieren: Einer Schätzung[2] zufolge stellen sie etwa 80 % aller vielzelligen Tiere. Es handelt sich zumeist um relativ kleine, weiße bis farblose, fädige Würmchen, die in feuchten Medien leben. Die Mehrzahl der Arten ernährt sich von Mikroben, es gibt aber auch räuberische und zahlreiche parasitische Arten, die Pflanzen, Tiere und den Menschen befallen.
| Fadenwürmer | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
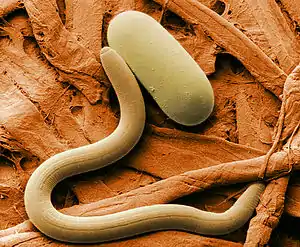
Heterodera glycines mit einem Ei, ein Parasit der Sojabohne | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Nematoda | ||||||||||||
| Rudolphi, 1808 | ||||||||||||
| Klassen | ||||||||||||
|
Fadenwürmer haben sich erfolgreich an nahezu jedes terrestrische und aquatische Ökosystem angepasst einschließlich extremer Lebensräume wie tieferer Bereiche der obersten Erdkruste und der Polarregionen. In vielen Lebensräumen stellen sie oft sowohl hinsichtlich der Individuenanzahl als auch der Artenvielfalt die größte Gruppe in der Metazoenfauna.
Anatomie
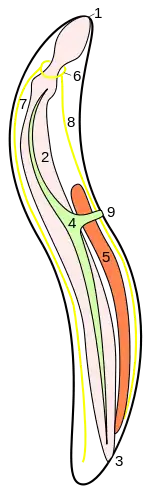
1 Mundöffnung
2 Darm
3 Kloake
4 Exkretionsorgan
5 Hoden
6 Circumpharyngealer Ring des Nervensystems
7 Dorsaler Hauptnervenstrang
8 Ventraler Hauptnervenstrang
9 Exkretionspore
Körperbau
Nematoden sind triploblastische Urmünder (Protostomia). Sie haben eine typisch wurmförmige Gestalt, sind lang und im Querschnitt rund. Eine Segmentierung fehlt. Die Körperhöhle ist ein enges Pseudocoel, wie auch bei mehreren anderen Wirbellosen-Tierstämmen mit überwiegend wurmartigen und sehr kleinwüchsigen Vertretern.
Das Vorderende eines Fadenwurmes enthält eine große muskulöse Mundöffnung mit anschließendem Pharynx (Rachen). Die Anatomie von Mund und Pharynx ist in vielfältiger Weise an die Art der Nahrungsaufnahme angepasst. Der schlauchförmige Körper enthält die Verdauungs- und Fortpflanzungsorgane. Der Anus liegt kurz vor dem Hinterende.
Der größte Fadenwurm ist das in der Pottwal-Plazenta lebende Placentonema gigantissima. Weibchen erreichen eine Länge von bis zu 8,40 m und einen Durchmesser von 2,5 cm, die Männchen werden nur 4 m lang bei einem Durchmesser von 0,9 cm. Diese Art gehört zur Klasse der Secernentea (Unterklasse Spiruria, Familie Tetrameridae).
Epidermis und Cuticula
Die Epidermis (Hautzellschicht) eines Fadenwurmes ist bemerkenswert, weil sie nicht wie bei anderen Tieren aus einzelnen Zellen besteht, sondern aus einer Masse des zellulären Materials, die nicht durch Membranen in einzelne Zellen unterteilt ist und mehrere Zellkerne besitzt. Solche Bildungen werden als Syncytium bezeichnet.
Die Epidermis sondert eine wesentlich dickere, mehrlagige Cuticula ab, die die Nematoden vor Austrocknung oder anderen ungünstigen Umweltbedingungen schützt, bei parasitischen Arten auch vor den Verdauungssäften des Wirtes. Neben den in heißen Quellen lebenden Arten wurden auch Arten gefunden, deren Cuticula ihnen ermöglicht, pH-Werte von 2,5 (z. B. Zitronensaft, Essig) bis 11,5 (z. B. Haushalts-Ammoniak) auszuhalten (siehe dazu auch folgende pH-Wert-Skala), oder solche, die mehrere Stunden in flüssigem Helium (ca. −272 °C bis −268 °C) am Leben bleiben. Das Vorhandensein einer „steifen“ Cuticula in Verbindung mit der Längsmuskulatur (Nematoden haben fast keine Ringmuskeln) erlaubt ihnen nur eine schlängelnde Fortbewegung.
Die Cuticula besteht bei Fadenwürmern aus bis zu vier Schichten:
- Die innere Faserschicht besteht aus diagonal miteinander in entgegengesetzter Richtung laufenden Fasern. Diese Schicht trägt am meisten zur Festigkeit und Elastizität der Cuticula bei.
- Die Matrixschicht hat eine weniger definierte Struktur.
- Die Kortikalschicht besteht aus Kollagen.
- Die äußere Epicuticula ist lipidhaltig und wird bei einigen Gattungen zusätzlich von einer Lipidschicht bedeckt.
Muskeln
Fadenwürmer besitzen wie die Rundwürmer zur Fortbewegung ausschließlich Längsmuskeln, die sich von Kopf bis Schwanz erstrecken. Die Muskelzellen bestehen aus drei Teilen:
- Dem Monocyton: Ein nicht zusammenziehbarer Teil, der die Zellkerne, die Mitochondrien und den Golgi-Apparat enthält.
- Einem zusammenziehbaren Teil, der die Actin- und Myosinfasern enthält.
- Dem Prozess, einem nicht zusammenziehbaren Teil der Muskelzellen, der Verbindungen mit anderen Muskelzellen oder Nerven eingehen kann.

Die Fadenwurmmuskeln liegen wie ein Schlauch unterhalb der Haut. Diese Einheit aus verschiedenen Geweben wird als Hautmuskelschlauch bezeichnet. Die starke Cuticula und der hohe Innendruck der Pseudocoelflüssigkeit, der zwischen 70 und 210 mmHg liegt, stellen ein sogenanntes Hydroskelett dar. Zusammen mit den Längsmuskeln als Antagonisten kann sich der Fadenwurm schlängelnd fortbewegen oder einen Teil in die Höhe strecken.
Daneben gibt es Ringmuskeln, jedoch nur an Mund und After.
Nervensystem
Das Nervensystem der Fadenwürmer ist sehr einfach aufgebaut. Es besteht aus einem circumpharyngealen bzw. circumoesophagealen Ring, von dem ein dorsaler und ein ventraler Hauptstrang nach hinten ziehen. Es ist in der Lage, einfache und verschiedene Reize wahrzunehmen und zu verarbeiten. Die Längsnerven sind direkt mit den Muskelzellen und dem Cytoplasma im Kontakt und erstrecken sich durch den ganzen Körper. Bemerkenswert ist dabei die Tatsache, dass anders als bei anderen Tieren, bei denen sich die Nervenzellen zu den Muskeln hin ausbreiten, sich die Muskelzellen des Fadenwurms selbst zu den Nervenbahnen ausbreiten.
Fortpflanzungsorgane
Die Fortpflanzungsorgane bestehen bei den Weibchen aus einer Vulva in der Körpermitte, wobei die Verlagerung vom Körperende nach vorne eine taxontypische Apomorphie darstellt.
Bei den Männchen ist eine Kloake am Körperende ausgebildet, die Samenleiter, Rektum und den Spicularapparat umfasst. Letzterer stellt das taxontypische Begattungsorgan dar und besteht aus paarigen, verhärteten, hakenförmigen Spicula (Singular: Spiculum), die in einer Tasche der Cuticula liegen und durch eine Führungsstruktur (Gubernaculum) in der Bewegung geleitet werden. Selten ist durch Reduktion oder Fusion nur ein Spiculum vorhanden. Die Spicula dienen nicht zum Samentransport, sondern verhaken das Männchen beim Begattungsakt in der Vulva des Weibchens, so dass die Spermien direkt aus dem Samenleiter übertragen werden können.[3][4]
Physiologie
Ernährung
Die Nahrung ist unterschiedlich und reicht bei freilebenden Arten von Bakterien und Algen über Pilze, Aas und Fäkalien bis hin zu räuberisch erbeuteten Tieren. Am Mund befinden sich oft kleine Fortsätze, die zur Nahrungsaufnahme oder zum Tasten benutzt werden. Dort wird die Nahrung hineingezogen und durch starke Muskeln zerquetscht. Die Nahrung gelangt dann von dort in einen einfachen langen Darmraum, wo sie bearbeitet und verdaut wird. Nematoden besitzen kein Gefäßsystem, mit dem sie die Nahrungsbestandteile im Körper verteilen könnten. Stattdessen werden die Nährstoffe im Darmraum verarbeitet und von dort direkt durch die Wände zu den Körperzellen geleitet, wo sie gebraucht werden.
Im Darm können auch endosymbiotische Mikroorganismen (Bakterien und Pilze) vorkommen, die bei der Aufspaltung von bestimmten Nahrungsbestandteilen benötigt werden, z. B. für den Abbau von Cellulose. Daneben wurde bei wenigen Arten wie Bursaphelenchus xylophilus und der in Käfern lebenden Pristionchus pacificus endogene Cellulasen nachgewiesen.[5] Für den Ursprung ihrer Cellulasegene wurde ein horizontaler Gentransfer, ausgehend von ihren Endosymbionten, verlautbart.[6][7]
Atmung
Die Sauerstoffaufnahme funktioniert ähnlich der Verdauung. Da die Nematoden keine Atmungsorgane und kein Gefäßsystem besitzen, wird der Sauerstoff durch die Haut aufgenommen und diffundiert direkt zu den Körperzellen.
Fortpflanzung
Die Fortpflanzung erfolgt sexuell, meist mit zwei getrennten Geschlechtern. Die Männchen sind typischerweise kleiner als die Weibchen und haben oft einen charakteristisch gebogenen Schwanz. Allerdings sind auch selbstbefruchtende Hermaphroditen, wie zum Beispiel Caenorhabditis elegans, keine Seltenheit.[8] Parasitische Arten haben oft einen recht komplizierten Lebens- und Fortpflanzungszyklus mit Generationswechsel, der mit Wirtswechsel oder Organwechsel im Wirt einhergehen kann. Hierbei können die Mikrofilarien, d. h. die Larven des Wurms, von Stechmücken aufgenommen werden und in der nächsten Entwicklungsstufe an andere Endwirte weitergegeben werden.[9]
Häutung
Die Nematoden häuten sich und werden deshalb, sowie aufgrund von RNA-Untersuchungen innerhalb der Urmünder (Protostomia) zu den Häutungstieren (Ecdysozoa) gerechnet. Bei freilebenden Arten erfolgt die Entwicklung meist direkt mit vier Häutungen im Verlauf des Wachstums.
Lebensräume
Nematoden kommen fast überall vor, im Meer, im Süßwasser und in terrestrischen Ökosystemen. Sie gelten allgemein als „häufig und omnipräsent“[10] und sind oft mit mehr Arten und Individuen in einem Ökosystem vertreten als alle anderen Gruppen vielzelliger Tiere (Metazoa). Sehr bedeutend sind sie in den Böden, wo sie mehrere der unteren trophischen Niveaus belegen. Des Weiteren haben Nematoden auch extreme Lebensräume besiedelt. Die Art Halicephalobus mephisto wurde in Südafrika in Kluftwässern in Tiefen von bis zu 3,6 Kilometern gefunden und ist damit das am tiefsten in der Erdkruste lebende vielzellige Tier.[11] Andere bevölkern die Böden der McMurdo Dry Valleys in der Antarktis, wo sie extrem widrige Bedingungen, unter denen durch das Zusammenspiel von extremer Kälte, Salinität und Trockenheit faktisch kein Bodenwasser verfügbar ist, durch Eintreten in ein Ruhestadium (Anhydrobiose) überleben können.[12][13] Es gibt auch eine erhebliche Anzahl parasitischer Arten, sowohl in Pflanzen (siehe etwa Rübenälchen) als auch in Tieren, einschließlich des Menschen. Zu den Nematoden, die den Menschen parasitieren und in seinem Darm leben, gehören zum Beispiel der Spulwurm (Ascaris lumbricoides), der Peitschenwurm (Trichuris trichiura), der Medinawurm (Dracunculus medinensis), der Madenwurm (Enterobius vermicularis) und der Zwergfadenwurm (Strongyloides stercoralis), wohingegen die Filarien Wuchereria bancrofti, Brugia malayi und Loa loa in den Lymphgefäßen bzw. im Unterhautfettgewebe leben.
Die meisten freilebenden Nematoden sind mikroskopisch klein und gehören zur Meiofauna. Lediglich einige Tierparasiten können deutlich größer werden.
Infektionswege und -strategien parasitischer Arten
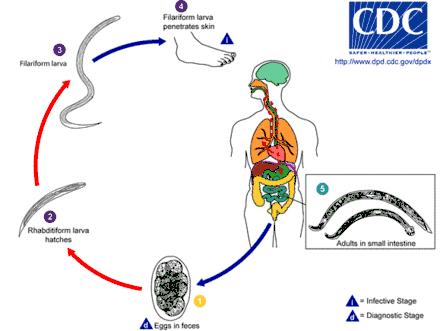
Die Infektion bzw. Infestation von als Endwirt dienenden Säugetieren, einschließlich des Menschen, geschieht z. B. durch den Verzehr von rohem Fleisch, in dem sich bereits Larven (z. B. Trichinen) befinden, oder infolge der Aufnahme von Wurmeiern durch das Fressen von Kot (beispielsweise bei Hunden). Auch aufgrund mangelnder Hygiene fäkal (mit Wurmeiern) verunreinigte Lebensmittel (Düngung von Nahrungspflanzen mit Gülle, kein Händewaschen nach Stuhlgang) können bei der Übertragung eine Rolle spielen. Bei mehreren Arten kann die Infektion aber auch durch aktives Eindringen von (filariformen) Larven durch die Haut geschehen (Hakenwürmer, z. B. Ancylostoma duodenale oder Necator americanus). Vertreter der Filarien wie Onchocerca volvulus, der Erreger der Flussblindheit, verbreiten sich in der Regel durch Insektenstiche.
Einige parasitische Nematodenarten sind im Hinblick auf ihren Lebens- und Fortpflanzungszyklus, wie auch parasitische Vertreter anderer Großgruppen von Wirbellosen oder Einzellern (vgl. u. a. → Verhaltensänderungen durch Toxoplasmose), in der Lage, das Verhalten und teils auch das äußere Erscheinungsbild ihrer Wirte zu manipulieren. Ein besonders komplexes Beispiel bietet Myrmeconema neotropicum.[14] Dieser Vertreter bewirkt, dass sich der mit seinen Eiern gefüllte Gaster der als Zwischenwirt dienenden Ameisenart Cephalotus atratus von schwarz zu kräftig rot verfärbt. Die betroffenen Ameisen, die im Vergleich zu ihren nicht-infizierten Artgenossen bei Bedrängung faktisch kein aggressives Abwehrverhalten zeigen und keine Alarmpheromone produzieren, recken permanent den roten Hinterleib in die Höhe. Dies soll wahrscheinlich fruchtfressende, als Endwirt dienende Vögel anlocken und zur Aufnahme des sich leicht vom Rest des Körpers ablösbaren Gasters animieren.[15]
Der Kiefernholznematode Bursaphelenchus xylophilus, ein in Deutschland und der Schweiz meldepflichtiger Quarantäneschaderreger, nutzt für seine Verbreitung Bockkäfer der Gattung Monochamus („Handwerkerböcke“) als Transportwirt.[16][17]
Taxonomie
Die Nematoden wurden ursprünglich von Nathan Cobb im Jahr 1919 als Stamm Nemata eingeführt, später als Klasse Nematoda in einem nicht mehr gültigen Stamm Aschelminthes klassifiziert. Hier werden die Fadenwürmer als eigener Stamm geführt. Die vorliegende Einteilung ist an die englische Wikipedia angelehnt und unterscheidet drei Klassen, fünf Unterklassen und 30 Ordnungen.
- Klasse Adenophorea
- Unterklasse Chromadoria
- Ordnung Areaolaimida
- Ordnung Ascaridida (Nicht in Wikispecies.)
- Ordnung Chromadorida
- Ordnung Desmodorida
- Ordnung Desmoscolecida
- Ordnung Monhysterida
- Ordnung Plectida
- Ordnung Rhabditida. Ascaris lumbricoides, Spulwurm, menschlicher Darmparasit.[18] Strongyloides stercoralis, Zwergfadenwurm, Erreger der tropischen Anguillulosis. Caenorhabditis elegans ist ein Modellorganismus der Molekularbiologen.
- Ordnung Spirurida. Onchocerca volvulus, Erreger der Flussblindheit[18] Wuchereria bancrofti, befällt die Lymphgefäße des Menschen. Übertragung durch Mücken. Loa loa, Wanderfilarie oder Augenwurm, Erreger der Loiasis oder Kamerunbeule[19]
- Ordnung Strongylida. Ancylostoma duodenale und Necator americanus sind Erreger der Grubenwurm- oder Hakenwurmkrankheit. Heterorhabditis[18]
- Unterklasse Chromadoria
- Klasse Enoplea (In Wikispecies als Ordnung.)
- Unterklasse Dorylaimia
- Ordnung Dioctophymatida (Nicht in Wikispecies.)
- Ordnung Dorylaimida
- Ordnung Isolaimida (Nicht in Wikispecies.)
- Ordnung Marimermithida (Nicht in Wikispecies.)
- Ordnung Mermithida (Nicht in Wikispecies.)
- Ordnung Mononchida (Nicht in Wikispecies.)
- Ordnung Muspiceida (Nicht in Wikispecies.)
- Ordnung Trichinellida (Nicht in Wikispecies.)
- Unterklasse Enoplia
- Ordnung Enoplida
- Ordnung Triplonchida. Trichuris trichiura, Peitschenwurm, Darmparasit des Menschen[20] Trichinella, Trichinen, Warmblüterparasiten, die durch Verzehr von ungegartem Fleisch auch den Menschen übertragen werden und das Krankheitsbild der Trichinose auslösen.[21]
- Unterklasse Dorylaimia
incerta sedis
- Ordnung Benthimermithida
- Ordnung Rhaptothyreida
- Klasse Secernentea
- Ordnung Camallanida (Nicht in Wikispecies.)
- Ordnung Drilonematida (Nicht in Wikispecies.)
- Ordnung Oxyurida. Enterobius vermicularis, Madenwurm, menschlicher Darmparasit[18]
- Ordnung Rhigonematida
- Ordnung Spirurida (Nicht in Wikispecies.)
- Unterklasse Diplogasteria
- Ordnung Diplogasterida (Nicht in Wikispecies.)
- Unterklasse Tylenchia
- Ordnung Aphelenchida (Nicht in Wikispecies.)
- Ordnung Tylenchida. Heterodera schachtii, Rübenälchen, Pflanzenparasit
Aus der englischen Wikipedia wurden Klassen, Unterklassen und Ordnungen übernommen. Diese stimmen nicht immer mit der Liste der Nematoden-Ordnungen in Wikispecies über ein.
Fossile Belege

Nematoden besitzen keine über geologische Zeiträume hinweg gut erhaltungsfähigen und relativ leicht in Sedimentgestein identifizierbare Körperteile. Deshalb ist ihr Fossilbericht im Verhältnis zu ihrer weiten rezenten Verbreitung und Vielfalt sehr lückenhaft und geringumfänglich und beschränkt sich auf ganz bestimmte Erhaltungsformen oder sedimentäre Fazies.[22][23]
Die meisten und am besten erhaltenen Nematoden-Fossilien wurden in Bernstein der Kreidezeit und des Tertiärs gefunden, wobei die ältesten dieser Exemplare aus der Unterkreide des Libanon kommen.[22][24][25] Die ältesten direkten Fossilbelege (Körperfossilien) entstammen dem berühmten Rhynie Chert des Unterdevons von Schottland und sind zugleich auch die ältesten direkten Belege für parasitische Nematoden. Sie bestehen aus Eiern, Juvenilstadien und ausgewachsenen Individuen, die im substomatären Hohlraum („Atemhöhle“) der frühen Landpflanze Aglaophyton major entdeckt und 2008 unter dem Namen Palaeonema phyticum beschrieben wurden.[22] Als Spurenfossilien freilebender mariner, infaunaler Nematoden gedeutete Sedimentstrukturen wurden aus Tonsteinen des Unterordoviziums der Hubei-Provinz Chinas beschrieben.[23]
Im Kolyma-Tiefland in Sibirien wurden Nematoden gefunden, die über etwa 42.000 bzw. 32.000 Jahre hinweg im Permafrostboden konserviert waren. Trotz der Tatsache, dass sie seit dem Jungpleistozän eingefroren waren, sollen zwei Exemplare dieser Würmer, die als Panagrolaimus aff. detritophagus und Plectus aff. parvus identifiziert wurden, erfolgreich wiederbelebt worden sein.[26]
Nematodenbekämpfung
Viele Nematodenarten sind Schädlinge in der Landwirtschaft und im Gartenbau, da sie durch ihr Eindringen in die Wurzelsysteme den Pflanzenstoffwechsel stark beeinträchtigen können. Gegen einen Nematodenbefall kommen verschiedene chemische Substanzen, die sogenannten Nematizide, sowie alternativ auch biologische Bekämpfungsmethoden, wie die Bepflanzung der befallenen Ackerflächen mit speziellen Nutzpflanzen (zum Beispiel resistenter Ölrettich, Tagetes und Senf), sowie thermische Verfahren, wie das Dämpfen (Bodendesinfektion) mit Heißdampf zur Bodenentseuchung zum Einsatz.
Nutzung und Forschung
Die Art Caenorhabditis elegans ist aufgrund ihrer einfachen Haltung und der Zellkonstanz (Eutelie) zu einem beliebten Versuchstier der Genetiker geworden und fungiert als Modellorganismus. Der Nematode Pristionchus pacificus wurde als Satellitenorganismus zu Caenorhabditis elegans etabliert. Durch den Vergleich dieser beiden Arten kann erforscht werden, wie sich Entwicklungsprozesse – der Übergang vom Ei zum erwachsenen Organismus – im Laufe der Evolution verändern. Außerdem werden Nematoden vermehrt als Nützlinge gegen Schnecken, Dickmaulrüssler und andere Pflanzenschädlinge verwendet.
Literatur
- Richard A. Sikora, Ralf-Peter Schuster: Handbuch der Phytonematologie. Berichte aus der Agrarwissenschaft. Shaker, Aachen 2000, 91 S., ISBN 3-8265-6978-4
- Johannes Hallmann: Biologische Bekämpfung pflanzenparasitärer Nematoden mit antagonistischen Bakterien. Mitteilungen aus der Biologischen Bundesanstalt für Land- und Forstwirtschaft Berlin-Dahlem, Heft 392. Dissertation. Biologische Bundesanstalt für Land- und Forstwirtschaft, Berlin und Braunschweig 2003, 128 S.
- Asmus Dowe: Räuberische Pilze und andere pilzliche Nematodenfeinde. 2., neu bearbeitete Auflage. (= Die Neue Brehm-Bücherei. Band 449). A. Ziemsen Verlag, Wittenberg Lutherstadt 1987, ISBN 3-7403-0042-6, 156 S.
- Susanne L. Kerstan: Der Befall von Fischen aus dem Wattenmeer und dem Nordatlantik 1988–1990 mit Nematodenlarven und eine Bibliographie über parasitische Nematoden in Fischen und Seesäugern. Berichte aus dem Institut für Meereskunde an der Christian-Albrechts-Universität Kiel, Nr. 219. Dissertation. Institut für Meereskunde, Abteilung Fischereibiologie, Kiel 1992, 205 S., doi:10.3289/ifm ber 219
- Andreas Overhoff: Einfluss von Bewirtschaftungssystem und Bodenbearbeitung auf die Populationsdichte von Nematoden. Mit besonderer Berücksichtigung antagonistischer Wirkung von Regenwürmern und nematophagen Pilzen. Dissertation. Wissenschaftlicher Fachverlag, Gießen 1990, ISBN 3-925834-87-7, 198 S.
- Jörn Alphei: Die freilebenden Nematoden von Buchenwäldern mit unterschiedlicher Humusform. Struktur der Gemeinschaften und Funktion in der Rhizosphäre der Krautvegetation. Berichte des Forschungszentrums Waldökosysteme. Reihe A, Band 125. Dissertation. Forschungszentrum Waldökosysteme der Universität Göttingen, Göttingen 1995, 165 S.
- Katrin Goralczyk: Küstendünen als Lebensraum für Nematoden. Forschen und Wissen – Umweltwissenschaft. Dissertation. GCA-Verlag, Herdecke 2002, ISBN 3-89863-095-1.
Einzelnachweise
- George O. Poinar, Jr.: Phylum Nemata. S. 169–180 in: James H. Thorp, D. Christopher Rogers (Hrsg.): Keys to Nearctic Fauna. Thorp and Covich’s Freshwater Invertebrates – Volume II. Fourth Edition. Academic Press/Elsevier, 2016, ISBN 978-0-12-385028-7
- T. Bongers, H. Ferris: Nematode community structure as a bioindicator in environmental monitoring. Trends Ecol Evol, Vol. 14, Issue 6, Juni 1999, S. 224–228, doi:10.1016/S0169-5347(98)01583-3
- Estelle V. Balian: Freshwater Animal Diversity Assessment. Springer, 2008, ISBN 978-1-4020-8258-0, S. 68–69.
- Peter Ax: Multicellular Animals: Order in Nature – System Made by Man: 3. Springer, Berlin 2003, ISBN 978-3-540-00146-1, S. 19–20.
- Käfer-Parasit mit ungewöhnlichen Genen: Genom des Fadenwurms Pristionchus pacificus entschlüsselt. g-o.de. 22. September 2008. Abgerufen am 1. Juli 2012.
- John T. Jones, Cleber Furlanetto, Taisei Kikuchi: Horizontal gene transfer from bacteria and fungi as a driving force in the evolution of plant parasitism in nematodes. Nematology. Bd. 7, Nr. 5, 2005, S. 641–646, doi:10.1163/156854105775142919.
- Werner E. Mayer, Lisa N. Schuster, Gabi Bartelmes, Christoph Dieterich, Ralf J. Sommer: Horizontal gene transfer of microbial cellulases into nematode genomes is associated with functional assimilation and gene turnover. BMC Evolutionary Biology. Bd. 11, Nr. 1, 2011, S. 13, doi:10.1186/1471-2148-11-13.
- Bei Steinernema longicaudum entwickelt das Männchen erst dann einen reifen Geschlechtsapparat und Spermien, wenn ein Weibchen in der Nähe ist, vgl. L. Ebssa, I. Dix, C. Griffin in Current Biology, Bd. 18, Heft 21, Seiten R997–R998
- Filarien. In: Lexikon der Biologie. Spektrum Akademischer Verlag, 1999, abgerufen am 25. April 2021.
- Gregor W. Yeates, Howard Ferris, Tom Moens, Wim H. Van der Putten: The Role of Nematodes in Ecosystems. S. 1–44 in: Michale J. Wilson, Thomas Kakouli-Duarte (Hrsg.): Nematodes as environmental indicators. CAB International, 2009, ISBN 978-1-84593-385-2
- G. Borgonie, A. García-Moyano, D. Litthauer, W. Bert, A. Bester, E. van Heerden, C. Möller, M. Erasmus, T. C. Onstott: Nematoda from the terrestrial deep subsurface of South Africa. In. Nature. Bd. 474, Nr. 7349, S. 79–82, doi:10.1038/nature09974; siehe dazu auch Dave Mosher: New „Devil Worm“ Is Deepest-Living Animal. In: National Geographic News, 2. Juni 2011
- Amy M. Treonis, Diana H. Wall, Ross A. Virginia: The use of anhydrobiosis by soil nematodes in the Antarctic Dry Valleys. In: Functional Ecology. Bd. 14, Nr. 4, 2000, S. 460–467, doi:10.1046/j.1365-2435.2000.00442.x.
- Amy M. Treonis, Diana H. Wall: Soil nematodes and desiccation survival in the extreme arid environment of the Antarctic Dry Valleys. In: Integrative and Comparative Biology. Bd. 45, Nr. 5, 2005, S. 741–750, doi:10.1093/icb/45.5.741.
- George. O. Poinar, Jr., Stephen P. Yanoviak: Myrmeconema neotropicum n. g., n. sp., a new tetradonematid nematode parasitising South American populations of Cephalotes atratus (Hymenoptera: Formicidae), with the discovery of an apparent parasite-induced host morph. In: Systematic Parasitology. Bd. 69, Nr. 2, 2008, S. 145–153, doi:10.1007/s11230-007-9125-3.
- Stephen P. Yanoviak, Michael E. Kaspari, Robert Dudley, George O. Poinar, Jr.: Parasite-Induced Fruit Mimicry in a Tropical Canopy Ant. In: American Naturalist. Bd. 171, Nr. 4, 2008, S. 536–544, doi:10.1086/528968.
- Ute Schönfeld: Der Kiefernholznematode (Bursaphelenchus xylophilus). Pflanzenschutzinformation: Pflanzengesundheitskontrolle 54/2015. Pflanzenschutzdienst des Landes Brandenburg, Landesamt für Ländliche Entwicklung, Landwirtschaft und Flurneuordnung, Frankfurt (Oder) 2015 (PDF 452 kB)
- Therese Plüss, Simone Prospero, Thomas Röthlisberger, Bea Schwarzwälder, Christiane Lellig (Red.): Leitfaden zum Umgang mit dem Kiefernholznematoden (Bursaphelenchus xylophilus). Eidgenössischer Pflanzenschutzdienst, Bundesamt für Umwelt, Bundesamt für Landwirtschaft, Bern 2015 (PDF 1,44 MB)
- Sebastian Suerbaum, Gerd-Dieter Burchard, Stefan H. E. Kaufmann, Thomas F. Schulz, Springer-Verlag GmbH: Medizinische Mikrobiologie und Infektiologie. 9., völlig überarb. u. erw. Auflage 2020. Berlin 2020, ISBN 978-3-662-61384-9.
- Charles Whittaker, Martin Walker, Sébastien D. S. Pion, Cédric B. Chesnais, Michel Boussinesq: The Population Biology and Transmission Dynamics of Loa loa. In: Trends in Parasitology. Band 34, Nr. 4, April 2018, ISSN 1471-5007, S. 335–350, doi:10.1016/j.pt.2017.12.003, PMID 29331268 (nih.gov [abgerufen am 23. Februar 2021]).
- Walter Alexander: American Psychiatric Association. In: Pharmacy and Therapeutics. Band 33, Nr. 6, Juni 2008, ISSN 1052-1372, S. 364–367, PMID 19561800, PMC 2683614 (freier Volltext).
- Edoardo Pozio, K. Darwin Murrell: Systematics and epidemiology of trichinella. In: Advances in Parasitology. Band 63, 2006, ISSN 0065-308X, S. 367–439, doi:10.1016/S0065-308X(06)63005-4, PMID 17134656 (nih.gov [abgerufen am 23. Februar 2021]).
- George O. Poinar, jr.: The Geological Record of Parasitic Nematode Evolution. S. 53-92 in: Kenneth De Baets, D. Timothy J. Littlewood (Hrsg.): Fossil Parasites. Advances in Parasitology, Bd. 90, Elsevier, 2015, doi:10.1016/bs.apar.2015.03.002
- Andrzej Baliński, Yuanlin Sun, Jerzy Dzik: Traces of marine nematodes from 470 million years old Early Ordovician rocks in China. In: Nematology. Bd. 15, Nr. 5, 2013, S. 567–474, doi:10.1163/15685411-00002702.
- George O. Poinar, jr., Aftim Acra, Fadi Acra: Earlist fossil nematode (Mermithidae) in cretaceous Lebanese amber. In: Fundamental and Applied Nematology. Bd. 17, Nr. 5, 1994, S. 475–477.
- für einen Überblick über die Funde speziell im eozänen baltischen Bernstein siehe Wolfgang Weitschat, Wilfried Wichard: Atlas der Pflanzen und Tiere im Baltischen Bernstein. Pfeil, München 1998, ISBN 978-3-931516-45-1, S. 54 ff.
- A. V. Shatilovich, A. V. Tchesunov, T. V. Neretina, I. P. Grabarnik, S. V. Gubin, T. A. Vishnivetskaya, T. C. Onstott, E. M. Rivkina: Viable Nematodes from Late Pleistocene Permafrost of the Kolyma River Lowland. In: Doklady Biological Sciences. Bd. 480, Nr. 1, 2018, S. 100–102, doi:10.1134/S0012496618030079.
Weblinks
- Nematode.net – Nematoden-Systematik
- Plant and Insect Parasitic Nematodes (engl.) – Seite der Universität Nebraska (Abgerufen am 20. Oktober 2009)
- Pristionchus pacificus die Wikionchus-Seite des Max-Planck-Instituts für Entwicklungsbiologie
- ProGemüse – Deutsch-Niederländisches EU-Projekt zur Nematodenproblematik im Gemüseanbau
- Max-Planck-Institut für Entwicklungsbiologie