Tenreks
Die Tenreks oder Tanreks (Tenrecidae) sind eine auf Madagaskar lebende Säugetierfamilie. Die Gruppe umfasst vor allem igel- und spitzmausähnliche Vertreter. Die nicht allzu großen Säugetiere sind hauptsächlich nachtaktiv, leben weitgehend einzelgängerisch und ernähren sich von Wirbellosen. Sie haben sich an unterschiedliche Lebensweisen angepasst. So gibt es unter den Tenreks grabende, bodenbewohnende, baumkletternde und im Wasser lebende Formen. Dadurch nutzen die Tiere verschiedene ökologische Nischen, die aufgrund fehlender Konkurrenten unter den Säugetieren auf Madagaskar unbesetzt geblieben waren. Diese Vielfältigkeit reduziert gleichzeitig die innerartliche Konkurrenz und führte zu mehr als 30 Arten, die sich aufgrund ihrer abweichenden Lebensweise auch im Körperbau teils deutlich unterscheiden. Insgesamt stellen die Tenreks so ein Paradebeispiel adaptiver Radiation dar. Darüber hinaus weisen die einzelnen Tenrekarten mitunter Eigenschaften auf, die auch für andere höhere Säugetiere eher ungewöhnlich sind. Hierzu gehören die stark schwankende Körpertemperatur und das Eintreten eines Torpors unter tropischen Bedingungen, das Vermögen einiger Igeltenreks, mit den Stacheln ihres Fellkleids zu stridulieren, sowie die teils extrem kurze Generationenfolge und die vor allem beim Großen Tenrek belegte hohe Anzahl an Neugeborenen.
| Tenreks | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Kleiner Igeltenrek (Echinops telfairi) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Tenrecidae | ||||||||||||
| Gray, 1821 |
Die erste Erwähnung von Tenreks durch westliche Naturforscher und Reisende erfolgte bereits in der Mitte des 17. Jahrhunderts. Rund 100 Jahre später waren schon mehrere Formen bekannt, bei denen es sich aber durchweg um stachelhaarige Tenreks handelte. Anfänglich wurden die Tenreks daher mit den europäischen Igeln in Beziehung gesetzt. Die Etablierung als eigenständige Gruppe setzte sich im Übergang vom 18. zum 19. Jahrhundert durch. Das charakteristische Erscheinungsbild und die teils insektenfressende Ernährungsweise waren Ursache, dass die Tenreks im Laufe des 19. und des 20. Jahrhunderts in die große, in sich aber nicht einheitliche Gruppe der Insektenfresser (Lipotyphla) eingeordnet wurden. Zum Ende des 20. Jahrhunderts zeigten molekulargenetische Untersuchungen jedoch, dass die Tenreks und die mit ihnan am nächsten Verwandten Otterspitzmäuse (Potamogalidae) zusammen mit den Goldmullen (Chrysochloridae) eine eigene Säugetiergruppe bilden: die Tenrekartigen (Afrosoricida). Diese Gruppe steht wiederum anderen originär afrikanischen Säugetieren nahe, mit denen sie unter der Bezeichnung Afrotheria zusammengefasst werden.
Der Ursprung der Tenreks ist bisher unbekannt. Auf Madagaskar liegen keine Fossilien vor, die älter als das Holozän datieren. Vom kontinentalen Afrika sind ebenfalls nur wenige Reste belegt, die ältesten stammen aus dem Eozän Südwestafrikas. Weitere Funde kamen im östlichen Afrika zu Tage und datieren in das Untere Miozän. Wahrscheinlich entwickelte sich die Gruppe auf dem Kontinent, die Vorfahren der madagassischen Tenreks gelangten dann vor etwa vierzig bis zwanzig Millionen Jahren auf die Insel. Es ist aber in der Fachwelt bisher umstritten, ob Madagaskar in einer oder in mehreren Besiedlungswellen erreicht wurde. Die heutigen Tenreks sind sowohl an die feuchten Regenwälder des östlichen als auch an die trockeneren Landschaften des westlichen Inselteils angepasst. Fast ein Fünftel der heutigen Arten sind in ihrem Bestand mehr oder weniger bedroht.
Merkmale
Habitus
_(44120142915)_2.jpg.webp)

Die Tenreks bilden eine vielgestaltige Gruppe kleinerer Säugetiere. Die Kopf-Rumpf-Länge variiert beträchtlich, sie erreicht bei den kleinsten Arten wie dem Zwergkleintenrek (Microgale parvula) oder dem Gnomkleintenrek (Microgale pusilla) 4,7 bis 6,4 cm, bei der größten Art, dem Großen Tenrek beträgt sie 26,5 bis 39,0 cm. Das Gewicht schwankt dementsprechend von 2,6 bis 2400 g. Die meisten Arten weisen keinen sonderlichen Geschlechtsdimorphismus auf.[1] Es lassen sich äußerlich zwei Formentypen unterscheiden. Die kleineren Arten wie die Reistenreks (Oryzorictinae) und Erdtenreks (Geogalinae) haben ein spitzmaus-, mitunter auch otterartiges Erscheinungsbild mit weichem Fell und langgezogenem Kopf. Die Schwanzlänge ist bei diesen variabel, es kommen Arten vor, deren Schwanz nur ein Drittel der Körperlänge ausmacht, bei anderen wiederum übertrifft er den restlichen Körper um mehr als das Doppelte an Länge. Die zweite Formengruppe, die Igeltenreks (Tenrecinae) zeichnet sich durch ein igelartiges Äußeres aus. Bei ihnen bildet der Schwanz nur einen kurzen Stummel, ihr Fell besteht aus Borsten und Stacheln. Allgemein variiert die Färbung des Fells der Tenreks von grau über braun bis schwarz, wobei die Unterseite meist heller gefärbt ist. Eine auffällige Musterung des Fells ist nur bei den Streifentenreks (Hemicentetes) ausgebildet. Die Hände und Füße enden bei den Tenreks in jeweils fünf Zehen, eine Ausnahme bildet hier der Vierzehen-Reiswühler (Oryzorictes tetradactylus), der nur vierstrahlige Hände besitzt. Die Beine zeigen keinen auffallenden Längenunterschied, der Daumen und die Großzehe sind nicht opponierbar. Der Kopf wirkt vergleichsweise groß und kann wie beim Großen Tenrek gut ein Drittel der Körperlänge ausmachen. Die Schnauze ist allgemein lang und beweglich. Die Ohrlänge variiert je nach ökologischer Anpassung der Tiere zwischen vergleichsweise groß bis klein, die Augen bleiben meist klein. Im Gesicht treten auffällig lange Vibrissen auf.[2][3][4]
Schädel- und Gebissmerkmale
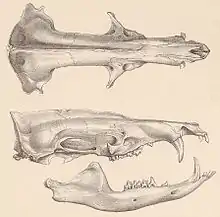
Der Schädel der Tenreks ist insgesamt langgestreckt und schmal sowie flach. Er besitzt ein lang ausgezogenes, häufig zylindrisch geformtes und vorn mehr oder weniger spitz zulaufendes Rostrum. Der Hirnschädel dagegen ist breiter, die größte Breite tritt hier meist an den Gelenkgruben für den Unterkiefer auf. Vor allem bei den Igeltenreks übertrifft das Rostrum den hinteren Schädelabschnitt deutlich an Länge und kann bis zu 60 % und mehr der Schädellänge einnehmen. Bei den Reistenreks ist das Rostrum dagegen kürzer und macht rund die Hälfte der Schädellänge aus. Auffälligstes Merkmal stellt der nicht geschlossene Jochbogen dar, was bei zahlreichen insektenfressenden Säugetieren vorkommt. Der Bereich hinter den Augen zieht häufig deutlich ein und ist langgestreckt. In der generellen Morphologie variieren die Schädel der Tenreks nur wenig,[5] im Detail weisen sie eine hohe Vielfalt auf, die ihre teils unterschiedlichen Anpassungen widerspiegeln. So kann das Rostrum sehr schmal sein wie bei den Streifentenreks oder breiter wie beim Großen Tenrek beziehungsweise beim Großen Igeltenrek. Auch ist ein Scheitelkamm unterschiedlich ausgebildet und kommt etwa beim Großen Tenrek vor, fehlt aber bei den Streifentenreks. Ebenso ist der Hirnschädel im unterschiedlichen Maße gewölbt oder abgeflacht und unter Umständen mit weiteren Knochenwülsten als Muskelansatzstellen ausgestattet. Im Embryonalstadium gleichen die Schädel aller Tenreks weitgehend denen der Kleintenreks (Microgale).[6][7]
Das Gebiss setzt sich aus 32 bis 40 Zähnen zusammen, die Zahnformel lautet: . Als gemeinsames Merkmal aller Tenreks gilt das Fehlen des ersten Prämolaren.[8] Im Bezug auf die vorderen Zähne unterscheiden sich die einzelnen Arten und Gattungen deutlich voneinander. Prinzipiell sind die Schneidezähne mit einer spitzen Zahnkrone ausgestattet, das Auftreten einzelner zusätzlicher Höckerchen vor und hinter der Hauptspitze variiert zwischen den einzelnen Arten. Der Eckzahn kann den Schneidezähnen gleichen, also incisiviform und sehr klein sein wie etwa bei den Kleintenreks oder den Streifentenrek oder aber wie bei den Reiswühlern und dem Großen Tenrek eher einem typischen Caninus (caniniform) entsprechen, bei letzterem überragt er die Schneidezähne. Auch die Anordnung der vorderen Zähne bis einschließlich der Prämolaren zeigt starke Unterschiede, da bei vielen Arten jeweils unterschiedlich ausgeprägte Diastemata (Zahnzwischenräume) vorkommen. Im Bau der Prämolaren und der Molaren weichen die Tenreks weniger deutlich voneinander ab. Sie werden ebenfalls durch spitze Höckerchen geprägt, von denen die Molaren jeweils drei tragen (tritubercular oder tricuspid). Die drei Haupthöcker umfassen, bezogen auf die Oberkieferzähne, den Paraconus, den Metaconus und den Protoconus. Paraconus und Metaconus liegen eng beieinander, der Protoconus ist in seiner Größe reduziert, nicht so stark wie bei den Goldmullen, aber stärker als bei den Otterspitzmäusen. Zudem besteht eine V-förmige Scherleiste (Ectoloph) auf der Kauoberfläche. Aufgrund dieser Merkmale können die Molaren der Tenreks als typisch zalambdodont aufgefasst werden. Teilweise besteht zungenseitig ein Cingulum, ein Zahnschmelzwulst.[6][9]

Skelettmerkmale
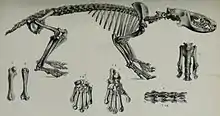
Wie bei allen Afrotheria zeichnet sich die Wirbelsäule der Tenreks durch eine erhöhte Anzahl der Rückenwirbel (Brust- und Lendenwirbel) aus. Sie besteht aus 7 Hals-, 15 bis 19 Brust, 4 bis 7 Lenden- 2 bis 3 Kreuzbein- und 8 bis 52 Schwanzwirbeln. Dabei besitzen die Igeltenreks die kürzesten Schwänze mit nur 8 bis 10 Wirbeln, was in einem sehr kurzen Schwanzstummel resultiert. Der Kleine Langschwanz-Kleintenrek (Microgale longicaudata) verfügt dagegen neben dem Langschwanzschuppentier über die höchste Anzahl an Schwanzwirbeln unter den heutigen Säugetieren, sein Schwanz erreicht häufig mehr als das Doppelte der Länge des restlichen Körpers.[10][6]
Generell zeigen die Tenreks einen übereinstimmenden Skelettbau, allen gemeinsam ist das Vorhandensein eines Schlüsselbeins und das im unteren Bereich miteinander verwachsene Schien- und Wadenbein. Zudem wird das obere Sprunggelenk wie bei vielen anderen Höheren Säugetieren nur durch Schien- und Sprungbein gebildet. Allerdings ist das obere Sprunggelenk im Bezug auf die Stellung der Knöchel bei den Igeltenreks etwas anders konfiguriert als bei den Reistenreks. Dies führt dazu, dass erstere ihre Füße in Ruhestellung nach außen rotiert halten, während sie bei letzteren parallel zur Körperachse orientiert sind. Darüber hinaus spiegeln sich die teils recht unterschiedlichen Lebensweisen der Tenreks durch verschiedene Variationen im Bau des Bewegungsapparates wider. Diese sind vor allen an den Vordergliedmaßen sichtbar, während die hinteren weniger deutlich betroffen sind. So tendiert das Schulterblatt bei grabenden Vertretern wie den Reiswühlern (Oryzorictes) oder den Streifentenreks (Hemicentetes) zu einer langschmalen Gestalt, bei kletternden Formen, etwa dem Kleinen Igeltenrek (Echinops) ist dieses dagegen kurz und breit. Zudem haben bodenwühlende Tenreks einen kurzen und breiten Oberarmknochen mit einem eher seitlich gepressten Gelenkkopf, dessen Form so rotierende Bewegungen einschränkt. Das untere Gelenkende (Ellenbogengelenk) verbreitet sich seitlich stark, was mit einem ebenfalls deutlich ausgezogenen Olecranon, dem oberen Gelenkfortsatz der Elle korrespondiert. Hier setzt hauptsächlich die Unterarmmuskulatur an. Bei baumbewohnenden Arten ist der Oberarmknochen dagegen eher lang und schmal, der Gelenkkopf mit seiner runden Form für weite rotierende Bewegungen ausgelegt und die Gelenke des Ellenbogens nicht so ganz auffällig gestreckt. Das Hand- und Fußskelett kann durch kurze und breite Knochen wie bei grabenden Arten oder durch längere und schmalere wie bei kletternden gekennzeichnet sein. Erstere besitzen auch einzelne Verwachsungen von Handwurzelknochen, was die Beweglichkeit der Hand mindert und stabilisiert. Außerdem ist der Mittelstrahl häufig verlängert.[11][12][13][14]
Weichteilanatomie
Die Tenreks besitzen in der vorderen Schnauze fünf Muskeltypen, die am Jochbein und der Voraugenregion ansetzen und mithilfe von Sehnen bis in die Nasenspitze reichen. Sie ermöglichen den Tieren, ihre langgestreckte Nase auf vielfältige Weise zu bewegen. Parallel zu den Anpassungen des Skelettes an die unterschiedlichen Lebensweisen kam es auch zu Veränderungen in der Muskulatur. Während bodenbewohnende Tiere eine relativ ursprüngliche Anordnung der Muskulatur bewahrten, entwickelte sich beim wasserbewohnenden Wassertenrek und bei dem grabenden Streifentenrek vor allem der Musculus semimembranosus des Hinterbeins enorm, dieser unterstützt das Knie bei der Beugung des Beines und sorgt somit für eine kraftvolle Bewegung im Wasser oder beim Graben. Ähnliches erfolgte auch an den Armen, wo es bei grabenden Tieren wie den Streifentenreks und den Reiswühlern zu Vergrößerung einiger Muskeln wie dem Musculus teres major und dem Musculus triceps brachii kam. Gleichzeitig bestehen zwischen den beiden Gattungen Unterschiede in der Arm- und Schultermuskulatur, etwa beim Größenverhältnis vom Musculus triceps brachii zum Musculus supraspinatus. Wahrscheinlich erfolgten die Anpassungen an die verschiedenen Lebensweise innerhalb der Unterfamilien der Tenreks unabhängig voneinander, was zu leicht variierenden Bewegungsweisen führte.[16][17]
Die Zunge wurde bisher nur beim Kleinen Igeltenrek genauer untersucht. Sie ist rund 2,0 cm lang und 0,5 bis 0,7 cm breit. Auf der Oberfläche sind vier verschiedenen Papillenarten ausgebildet: konische und fadenförmige mechanische sowie pilzförmige und eingedellte Geschmackspapillen. Die fadenförmigen Papillen besitzen im vorderen Abschnitt der Zunge lediglich einen spitzen Fortsatz. Die Anzahl nimmt nach hinten zur Zungenwurzel zu, so dass die Papillen hier kronenartig erscheinen. Die pilzförmigen Geschmackspapillen verteilen sich auf je vier Reihen an den Rändern der Zungenoberseite, während drei eingedellte Geschmackspapillen an der Zungenwurzel auftreten und ein Dreieck formen. Speicheldrüsen durchsetzen die gesamte Zungenoberfläche mit Ausnahme der Zungenspitze Daneben treten noch Drüsen mit serös-schleimiger beziehungsweise rein seröser Ausscheidefunktion auf.[18]
Ein auffälliges Merkmal der Tenreks ist die Kloake, eine gemeinsame Austrittsöffnung für Geschlechts-, Verdauungs- und Ausscheidungsorgane. Die Kloake hat eine Schüsselform, der Eingang des Verdauungstraktes wird über einen Schließmuskel reguliert.[19] Im Verdauungstrakt fehlt der Blinddarm.[20] Die Hoden liegen bei vielen Arten in der Bauchhöhle, bei den Kleintenreks und den Reiswühlern sind sie in das Becken vorverlagert. Die Größe der paarigen Hoden variiert bei den Reistenreks zwischen 0,07 und 0,56 g, was 0,8 bis 2 % der Körpermasse entspricht. Der Penis ist sehr dünn und ausgesprochen lang, er erreicht beim Kleinen Langschwanz-Kleintenrek gut 40 % und beim Großen Tenrek gut 70 % der Kopf-Rumpf-Länge, Penisstacheln sind nicht ausgebildet. Die Gebärmutter der Weibchen ist stets zweihörnig (Uterus bicornis), auffallend erscheint der Utero-Vagina-Kanal, der einem langen und dünnen sowie spiralartig gewundenen Schlauch gleicht und so der Form des Penis angepasst ist.[21][22] Im Unterschied zu den Otterspitzmäusen haben die Tenreks eine haemochoriale Plazenta.[23][24]
Das Gehirn zählt zu den am einfachsten gebauten aller Höheren Säugetiere. Das Gewicht beträgt 420 bis 2490 mg, was durchschnittlich etwa der Gehirngröße bei den Spitzmäusen entspricht, aber deutlich weniger als bei den etwa gleich großen Rüsselspringern ist.[25] Der Enzephalisationsquotient variiert zwischen 0,3 und 1,0. Er ist bei den kleinen weichhaarigen Tenreks deutlich höher als bei den größeren Igeltenreks.[26] Neocortex und Striatum sind sehr klein, der Riechkolben hingegen vergrößert.[27][28][29]
Verbreitung und Lebensraum
Die Tenreks sind heute endemisch auf Madagaskar verbreitet. Lediglich der Große Tenrek kommt auch auf den Maskarenen, Seychellen und dem Archipel der Komoren vor, er ist dort aber wohl in allen Fällen durch den Menschen eingebracht worden. Die Tiere bewohnen auf Madagaskar eine Vielzahl von Lebensräumen, diese schließen die feuchten Regenwälder des östlichen und zentralen Inselteils, die trockenen Wälder des westlichen und die Dornbuschsavannen des südwestlichen ein. Die meisten, vor allem kleineren Vertreter kommen in enger umgrenzten Lebensräumen vor. Es lassen sich drei Ökotypen unterscheiden:[30]
- Bewohner feuchter Landschaften: etwa 74 % aller Tenrekarten, vor allem ein Großteil der Vertreter der Kleintenreks, zudem die Vertreter der Gattung Nesogale, die Reistenreks und die Streifentenreks;
- Bewohner trockener Landschaften: etwa 19 % aller Tenrekarten, so einige Vertreter der Kleintenreks (Jenkins-Kleintenrek, Nasolo-Kleintenrek, Kurzschwanz-Kleintenrek, Grandidier-Kleintenrek), der Erdtenrek und der Kleine Igeltenrek;
- Bewohner beider Landschaftstypen: prinzipiell kommen nur der Große Igeltenrek und der Große Tenrek auf der gesamten Insel Madagaskar vor.
Die Tenreks sind sowohl auf Meeresspiegelhöhe als auch in hohen Gebirgslagen bis 2500 m anzutreffen. In vielen Habitaten tritt mehr als eine Art auf. So wurden am Tsaratanana-Massiv im Norden bis zu 18 Arten nachgewiesen,[31] während es im Waldgebiet von Tsinjoarivo immerhin noch 16 Arten sind.[32] Das häufige gemeinsames Auftreten spricht für eine starke ökologische und trophische Differenzierung innerhalb der Familie.[4][2][10]
Lebensweise
Territorial- und Sozialverhalten
_in_a_hollow_treetrunk%252C_Ankarafantsika_National_Park%252C_Madagascar.jpg.webp)


Die Lebensweise ist nur bei den wenigsten Arten gut erforscht, dies gilt vor allem für die Vertreter der Igeltenreks. Bei den kleineren Formen liegen mit Ausnahme des Erdtenreks und der Angehörigen der Gattung Nesogale nur spärliche Informationen vor. Generell zeigen die Tenreks ein variantenreiches Spektrum an Anpassungen an die unterschiedlichsten Lebensweisen, die sich wohl aufgrund der ursprünglichen Abwesenheit von ökologischen Konkurrenten wie Spitzmäusen, Igeln oder Maulwürfen herausgebildet haben. Es können verschiedene Grundtypen unterschieden werden: Dazu gehören die unterirdisch grabenden Formen wie die Streifentenreks, die Reiswühler, der Erdtenrek oder einige kurzschwänzige Kleintenreks. In der Regel sind die Tiere mit kurzen Ohren, kleinen Augen und langen, breiten Krallen an die bodenwühlende Lebensweise angepasst, Des Weiteren gibt es baumkletternde Formen wie den Kleinen Igeltenrek, die Vertreter der Gattung Nesogale oder zahlreiche langschwänzige Kleintenreks. Alle kletternden Formen verfügen als Anpassung über kurze, stark gebogene Krallen. Intermediär dazwischen stehen die generell bodenbewohnenden Formen, so der Große Tenrek, der Große Igeltenrek oder Kleintenreks mit mittellangem Schwanz. Als einzige wasserbewohnende Form tritt der Wassertenrek auf, der Schwimmhäute ausgebildet hat. Es gibt allerdings häufig Überschneidungen zwischen den einzelnen Grundtypen. So legt der Wassertenrek auch unterirdische Baue an, zudem können zahlreiche bodenlebende Kleintenreks und der Große Igeltenrek ebenfalls klettern.[33][4][2]
Die Tenreks sind generell Einzelgänger, vorübergehende Paarbildungen kommen weitgehend nur während der Fortpflanzungsphase vor. Eine Ausnahme bildet der Eigentliche Streifentenrek, der in der Paarungszeit in Kolonien aus mehreren Generationen lebt. Die Aktivitätszeiten der Tenreks beschränken sich überwiegend auf die Nachtstunden, tagsüber ziehen sie sich in selbstgegrabene Erdbaue, Baumhöhlen oder andere Unterschlüpfe zurück. Auffallend ist bei den besser untersuchten Igeltenreks eine bimodale Verteilung der Aktivität, die die Mitternachtsstunden zumeist auslässt.[3][4] Die Tiere bewegen sich am Boden hauptsächlich im Kreuzgang fort. Kletternde Tenreks sind eher langsam, die stark gebogenen Krallen werden dabei wie Steigeisen eingesetzt, bei den langschwänzigen Kleintenreks fungiert der lange Schwanz teilweise als Greiforgan.[7] Die Orientierung erfolgt weitgehend über den Geruchssinn. In unbekanntem oder unübersichtlichen Gelände können die Tenreks aber auch einfache Echoortungsrufe ausstoßen. Diese werden durch Zungenklicks in einem Frequenzbereich von 5 bis 17 kHz erzeugt. Bekannt ist dies von fast allen Igeltenreks und vom Dobson-Kleintenrek.[34] Eine besondere Kommunikationsform tritt bei einigen Igeltenreks auf. Diese können mithilfe ihrer Stacheln rasselnde Geräusche erzeugen, was als Stridulation bezeichnet wird. Bei den Streifentenreks ist dafür am hinteren Rücken ein spezielles Stridulationsorgan ausgebildet, das aus besonders geformten Stacheln besteht. Dieses kommt auch beim Großen Tenrek vor, allerdings nur bei Jungtieren. Auch der Kleine Igeltenrek striduliert gelegentlich, bei ihm sind die dafür verantwortlichen Stacheln regelmäßig über dem Rücken verteilt.[21][34][4][35]
Das Revierverhalten der Tenreks ist kaum erforscht. Nach Untersuchungen im Nationalpark Ankarafantsika nutzt der Große Igeltenrek dort vergleichsweise große Aktionsräume, die bis zu 13,7 ha einnehmen können.[36] Bei Begegnungen zwischen Artgenossen beschnuppern sich die Tiere an verschiedenen Körperteilen. Aggressives Verhalten führt bei den Igeltenreks zum Aufrichten der Stacheln, vor allem der im Nackenbereich, und zu typisch stoßenden Kopfbewegungen, teilweise verbunden mit Beißattacken. Bei Bedrohungen wird häufig das Maul aufgerissen, der Kleine und der Große Igeltenrek können sich bei Gefahr zu einer Kugel zusammenrollen. Über Sekretmarkierungen ist wenig bekannt, in der Regel werden die Eingänge der Baue mit Kot gekennzeichnet. In besonderen Stresssituationen tritt bei Tenreks ein weißliches Sekret an den Augen aus. Hierbei handelt es sich offensichtlich um eine Körperreaktion, die dem menschlichen „Augentränen“ gleicht.[37][38][4]
Ernährung und Energiehaushalt
Entsprechend ihren vielfältigen Lebensweisen ist auch die Ernährungsweise der Tenreks relativ variantenreich. In erster Linie besteht die Hauptnahrung aus Wirbellosen, vor allem Insekten und Regenwürmer. Der Wassertenrek nimmt zusätzlich auch Krebstiere zu sich.[39] Die Streifentenreks dagegen sind fast ausschließlich auf Regenwürmer spezialisiert.[3] Für einige Arten ist auch belegt, dass sie kleine Wirbeltiere erlegen oder Aas fressen. In geringem Ausmaß wird zusätzlich pflanzliches Material wie Früchte verzehrt. Bei zahlreichen der kleineren Vertreter aus der Gruppe der Reistenreks ist die genaue Ernährungsweise nur unzureichend erforscht. Hier verweisen aber Isotopenanalysen auf ein Überwiegen von tierischer Nahrung, deren Zusammensetzung je nach bevorzugter Lebensweise variiert.[32] Bei der Nahrungssuche wird überwiegend die Nase eingesetzt. Die Tiere nehmen ihre Nahrung mit dem Maul auf, eine Manipulation mit den Vorderfüßen findet kaum statt.[3][4][2]
Die meisten Tenreks können ihre Körpertemperatur nicht im gleichen Ausmaß regeln wie ein Großteil der anderen Höheren Säugetiere. Ausnahmen bilden hier die Kleintenreks, deren Körpertemperatur offensichtlich stabiler ist, allerdings sind bisher zu wenige Arten untersucht. Bei den übrigen Tenreks passt sich die Körpertemperatur im Ruhezustand weitgehend der Außentemperatur an und ist deshalb Schwankungen unterworfen. Während der Aktivitätsphase nachts stellen sich häufig über mehrere Stunden stabilere Zustände ein, zusätzlich schwankt die Körpertemperatur bei Weibchen in der Reproduktionsphase weniger stark. Die stabileren Zustände gehen in vielen Fällen mit einer erhöhten Stoffwechselrate einher, die ansonsten weitaus geringer ist, als von vergleichbar großen Säugetieren zu erwarten wäre.[4][40][41][42][43] Die Tenreks werden daher bisweilen als „protoendotherm“ oder „basoendotherm“ angesehen. Für mehrere Arten ist das Einsetzen eines Torpors (Starrezustand) bei ungünstigen äußeren Bedingungen bekannt. Derartige Starrephasen können kurzfristig während des Südsommers von November bis Mai auftreten,[44][45] werden aber ausgedehnter während des Südwinters von Juni bis Oktober. Teilweise verbringen die Tiere diese trockene und verhältnismäßig nahrungsarme Zeit durchgehend im Torpor, für den Großen Igeltenrek ist eine Dauer von bis zu sieben Monaten belegt,[46] für den Großen Tenrek sogar bis zu neun.[47] Neben einer so allgemein energiesparenden Lebensweise unter tropischen Bedingungen liegt der Vorteil derartig langer Ruheperioden wahrscheinlich darin, dass die Tiere so außerhalb der Reproduktionszeit kaum von Fressfeinden erbeutet werden können. Dies wiederum resultiert in einer höheren Lebenserwartung. Das in Bezug auf ihren Energiehaushalt und Metabolismus plastische Verhalten der Tenreks kann als Ergebnis der Entwicklung auf einer eher isolierten Insel mit wenig schwankenden Umweltverhältnissen und einer überschaubaren Anzahl an homoiothermen Konkurrenten aufgefasst werden. Nahe verwandte Gruppen auf dem kontinentalen Afrika, die unter extremeren Bedingungen leben, verfügen nicht über so stark variierende Körperfunktionen.[47][48] Die jeweils eigenständige Erhöhung der Körpertemperatur nach einer täglichen Ruhephase oder dem Torpor wird möglicherweise durch Braunes Fettgewebe unterstützt. Zumindest beim Kleinen Igeltenrek konnte dies in kleineren Mengen am Nacken und in größeren an den Fortpflanzungsorganen nachgewiesen werden.[49]
Fortpflanzung
Die Fortpflanzung ist nur bei den Igeltenreks, dem Erdtenrek und den Vertretern der Gattung Nesogale vergleichsweise gut untersucht. Bei einigen Arten wie beim Großen Tenrek tritt eine Polyovulation auf, bei der sich mehrere Eizellen im Follikel ablagern. Bemerkenswert ist, dass die Follikel keinen Graafschen Follikel bilden, wie Untersuchungen beim Großen Tenrek, beim Kleinen Igeltenrek und bei den Streifentenreks nahelegen. Vielmehr quellen die Granulosazellen auf, so dass sowohl die Theca als auch das Follikelepithel saftig wird. Die Befruchtung der Eizelle erfolgt noch im Follikel, wofür die Spermien die Theca durchdringen. Erst danach kommt es zur Ovulation. Dabei dringt die befruchtete Eizelle langsam Richtung Follikeloberfläche. Ausgelöst wird der Vorgang durch den Quelldruck der Granulosa. In Bezug auf den Charakter des Follikels, der darin stattfindenden Befruchtung und des erst danach erfolgenden Eisprungs weichen die Tenreks deutlich von den anderen Höheren Säugetieren ab.[50][51][52]

Soweit bekannt beschränkt sich die Reproduktionsphase auf den Südsommer und ist so saisonal abhängig. Sie fällt damit in die feuchte und nahrungsreiche Jahreszeit. Das Werberitual der Männchen besteht aus dem Beschnuppern verschiedener Körperteile des Weibchens.[4][2] Die Tragzeit liegt meist zwischen 50 und 70 Tagen, sie variiert wenig zwischen den Arten, kann aber innerhalb einzelner Arten beträchtlich schwanken. Demnach beeinflussen einzelne Torporphasen die Dauer der Tragzeit wie beim Erdtenrek oder beim Großen Tenrek,[40][53] ebenso wie die jeweilige Witterung. Beim Kleinen Igeltenrek wiederum werden die Tragzeiten kürzer, je später die Trächtigkeit in der Fortpflanzungsperiode eintritt.[54] Insgesamt ist die Frage der Einflussfaktoren bisher wenig geklärt. Die Anzahl der Zitzenpaare der Weibchen variiert zwischen drei und vier bei den Kleintenreks bis zu 14 beim Großen Tenrek. Dementsprechend liegen auch die Wurfgrößen zwischen einem und bis zu 32 beim Großen Tenrek, letzteres ist der größte Wert aller Säugetiere. Generell haben die weichhaarigen kleineren Tenreks kleinere Wurfgrößen als die stacheligen Igeltenreks.[46]
Neugeborene Tenreks kommen als Nesthocker zur Welt und sind blind sowie nackt, die Individualentwicklung verläuft vergleichsweise schnell. Auffällig sind Unterschiede bei einzelnen Phasen der Individualentwicklung zwischen den stacheligen Igeltenreks und den weichhaarigen kleineren Arten. So öffnen sich die Augen bei den Igeltenreks bereits nach neun bis 14 Tagen, während dieser Prozess bei den kleineren Formen bis zu vier Wochen andauert. In der Regel sind die Jungen nach einem bis zwei Monaten ausgewachsen. Die insgesamt kurze Entwicklungsphase und die Befähigung einiger Arten, noch während der Aufzucht erneut trächtig zu werden bedingt, dass Weibchen mehrere Würfe während einer Fortpflanzungsphase haben können.[38][4][46] Den bisherigen Untersuchungen zufolge ist bei den Tenreks weder die Tragzeit noch die Individualentwicklung allgemein abhängig vom Körpergewicht eines Individuums/einer Art, was von den Gegebenheiten bei anderen Höheren Säugetieren abweicht. Dies lässt vermuten, dass einige Elemente in der Biologie der Familie eher phylogenetisch und nicht konstitutionell gesteuert werden.[55][40][41][2]
Systematik
Äußere Systematik
Innere Systematik der Afrotheria nach Heritage et al. 2021[56]
|
Die Tenreks sind eine Familie aus der Ordnung der Tenrekartigen (Afrosoricida), die außerdem noch die Otterspitzmäuse (Potamogalidae) und die Goldmulle (Chrysochloridae) enthält. Die nächsten Verwandten der Tenreks bilden dabei die Otterspitzmäuse, die enge Beziehung zueinander wird durch das übergeordnete Taxon der Tenrecomorpha innerhalb der Afrosoricida ausgedrückt.[30] Die Tenrekartigen wiederum gehören zur Überordnung der Afrotheria, einer der vier Hauptlinien innerhalb der Höheren Säugetiere. Die Afrotheria stellen eine Verwandtschaftsgemeinschaft aus überwiegend in Afrika heimischen oder von dort stammenden Tiergruppen dar. Ihre Zusammengehörigkeit beruht vor allem auf molekulargenetischen Untersuchungen weniger auf anatomischen Gemeinsamkeiten. Innerhalb der Afrotheria bestehen zwei größere Linien, einerseits die Paenungulata und andererseits die Afroinsectiphilia. Erstere schließen die heutigen Elefanten, die Schliefer und die Seekühe ein, die Gruppe galt schon länger als eine gemeinsame Abstammungseinheit. Die letztere Linie vereint neben den Tenrekartigen auch die Rüsselspringer und das Erdferkel. Die genaueren Verwandtschaftsverhältnisse der drei letztgenannten Gruppen sind in Diskussion. Einige genetische Untersuchungen befürworten eine Schwestergruppenbeziehung der Afrosoricida zu den Rüsselspringern, beide Gruppen werden dann als Afroinsectivora zusammengefasst.[57][58][59] Andere wiederum sehen die Tenrekartigen in einer Schwestergruppenposition zu allen anderen Afrotheria, während die Rüsselspringer eine engere Beziehung zum Erdferkel formen.[60] Laut den molekulargenetischen Untersuchungen zufolge bildeten sich die Afrotheria in der Oberkreide vor 90,4 bis 80,9 Millionen Jahren heraus, ihre Aufspaltung in die beiden Hauptgruppen schloss sich etwa 15 Millionen Jahre später an. Die Afrosoricida traten dann kurz vor der Kreide-Tertiär-Grenze vor etwa 68 Millionen Jahren in Erscheinung.[59] Rund zehn Millionen später trennten sich die Goldmulle von der gemeinsamen Linie mit den Tenreks und den Otterspitzmäusen ab.[30]
Ebenfalls in ein näheres Verwandtschaftsverhältnis zu den Tenreks gehört das ausgestorbene Plesiorycteropus. Ursprünglich wurde eine engere Beziehung zum Erdferkel angedacht, was sich auch in dem deutschen Trivialnamen „Madagassisches Erdferkel“ ausdrückt.[61] Spätere anatomische Analysen zeigten jedoch keine unmittelbare Nähe zum Erdferkel, vielmehr erwiesen sich zahlreiche als übereinstimmend angesehene Merkmale zwischen beiden als Anpassung an eine grabende Lebensweise. Basierend darauf lagerte Ross D. E. MacPhee Plesiorycteropus in die eigene Ordnung der Bibymalagasia aus.[62] Genetische Analysen aus dem Jahr 2013 erbrachten nun, dass Plesiorycteropus den Tenreks nahesteht, die Tiere wären demnach als giant tenrecs („Riesentenreks“) anzusehen.[63]
Innere Systematik
Innere Systematik der Tenreks nach Everson et al. 2016[30]
|
Die Familie der Tenreks wird heute in drei Unterfamilien geteilt. Dabei bilden die Igeltenreks (Tenrecinae) die Schwestergruppe der übrigen Tenreks (Reis-- und Erdtenreks beziehungsweise Oryzorictinae und Geogalinae). Erstere unterscheiden sich durch ihr stacheliges, an Igel erinnerndes Fell und ihren kurzen Stummelschwanz von letzteren beiden, die wiederum durch ein weichhaariges Fell, ein spitzmausartiges Äußeres und einen langen Schwanz gekennzeichnet sind. Die Trennung der beiden Linien fand laut molekulargenetischen Untersuchungen im Oberen Eozän vor etwa 35,6 Millionen Jahren statt. Die äußerlich ähnlichen Angehörigen der Oryzorictinae und der Geogalinae zeigen Abweichungen in der Gebissstruktur, da der Erdtenrek ein gegenüber den Reistenreks deutlich reduziertes Gebiss aufweist. Die beiden Unterfamilien spalteten sich im Unteren Oligozän vor rund 30,1 Millionen Jahren voneinander ab.[30][4]
Teilweise wurden die Otterspitzmäuse als vierte Unterfamilie der Tenreks angesehen.[64][65] Eine molekulargenetische Studie aus dem Jahr 2016 stuft sie allerdings wieder als eigenständige Familie ein, die die Schwestergruppe der Tenreks bildet. Beide Familien teilen zahlreiche Merkmale, voran das zalambdodonte Gebiss und den fehlenden Jochbogen. Allerdings liegen auch bedeutende Unterschiede vor. So verfügen die Otterspitzmäuse gegenüber den Tenreks nicht über ein Schlüsselbein, des Weiteren sind die zweiten und dritten Zehen des Hinterfußes miteinander verwachsen und der Eckzahn besitzt zwei Wurzeln. Die Trennung der beiden Familien vollzog sich bereits im Unteren Eozän vor rund 48 Millionen Jahren.[30]
Eine Debatte bestand in der Frage, ob die madagassischen Tenreks eine monophyletische Einheit bilden. So ergaben anatomische Untersuchungen von Robert J. Asher aus dem Jahr 1999 einen polyphyletischen Ursprung der madagassischen Tenreks im Bezug auf die Stellung der Otterspitzmäuse und der Goldmulle. Je nach Gewichtung der Merkmale zeichnete sich einerseits eine stärkere Einheit der Otterspitzmäuse mit dem Wassertenrek sowie den anderen weichhaarigen Tenreks und der Goldmulle mit den Igeltenreks heraus, andererseits aber auch eine tiefe Einbettung der Otterspitzmäuse in die Tenreks unter Einbehaltung der engen Verwandtschaft ersterer zum Wassertenrek.[66] Kurz darauf publizierte molekulargenetische Analysen widersprachen dem und ließen vermuten, dass die Tenreks zusammen mit den Otterspitzmäusen eine Klade bilden und die Goldmulle als Schwestergruppe fungieren (die Analysen schlossen damals aber nur eine begrenzte Anzahl an Tenrekarten ein).[67][68] Nachfolgende genetische Studien unter Einbeziehung weiterer Arten betätigten dieses Bild und führen zu der Ansicht, dass alle heute auf Madagaskar vorkommenden Tenreks auf einen gemeinsamen Vorfahren zurückgehen.[69][30]
Überblick über die rezenten und fossilen Gattungen der Tenreks
Die rezenten Vertreter der Familie der Tenreks gliedern sich folgendermaßen:[30][70]
- Familie Tenrecidae Gray, 1821 (Tenreks)
- Unterfamilie Tenrecinae Gray, 1821 (Igeltenreks oder Borstenigel)
- Tenrec Lacépède, 1799 (Großer Tenrek, 1 Art)
- Hemicentetes Mivart, 1871 (Streifentenreks, 2 Arten)
- Setifer Froriep, 1806 (Großer Igeltenrek, 1 Art)
- Echinops Martin, 1838 (Kleiner Igeltenrek, 1 Art)
- Unterfamilie Oryzorictinae Dobson, 1882 (Reistenreks)
- Oryzorictes A. Grandidier, 1870 (Reiswühler, 2 Arten)
- Microgale Thomas, 1882 (Kleintenreks, 21 Arten)
- Nesogale Thomas, 1918 (2 Arten),
- Unterfamilie Geogalinae Trouessart, 1879 (Erdtenreks)
- Geogale Milne-Edwards & A. Grandidier, 1872 (Erdtenrek, 1 Art)
Ursprünglich gehörte mit Limnogale noch eine weitere Gattung zu den Reistenreks, die den Wassertenrek (Microgale mergulus) enthielt und monotypisch war. Bereits Anfang des 21. Jahrhunderts vorgenommene molekulargenetische Untersuchungen der Tenreks legten eine enge Bindung an die Kleintenreks nahe,[71][72] einige Studien sprachen auch für eine tiefe Einbettung des Wassertenreks in die Gattung Microgale.[73][58] Letzteres wurde durch eine neuere genetische Studie aus dem Jahr 2016, die alle bekannten Vertreter der Tenreks berücksichtigte, bestätigt, so dass der Wassertenrek in die Kleintenreks eingegliedert wurde.[30]
Fossil können folgende Formen noch unterschieden werden:[74][75][76]
- Sperrgale Pickford, 2015
- Arenagale Pickford, 2015
- Protenrec Butler & Hopwood, 1957
- Erythrozootes Butler & Hopwood, 1957
- Parageogale Butler, 1984
- Promicrogale Pickford, 2018
Im Jahr 1987 wurde anhand einzelner Unterkieferreste aus dem Unteren Miozän von Songhor im westlichen Kenia die Gattung Ndamathaia beschrieben, die die Autoren als einen riesenhaften Tenrek einstuften.[77] Eine phylogenetische Studie aus dem Jahr 2000 kommt dagegen zu dem Schluss, dass es sich hierbei um Überreste einer Manguste handelt, die zur ausgestorbenen Gattung Kelba gehört.[78]
Forschungsgeschichte
Entdeckung


Die ersten Hinweise auf die Tenreks in der westlichen Wissenschaft gehen auf die Mitte des 17. Jahrhunderts zurück und stammen von Étienne de Flacourt (1607–1660). Der Naturforscher und ehemalige Gouverneur von Madagaskar veröffentlichte im Jahr 1658 mit dem Werk Histoire de la grande isle Madagascar seinen Reisebericht über die Insel (zuzüglich seines Aufenthaltes im kontinentalen Afrika), in dem er auch die Naturgeschichte behandelte. In dem mehrfach aufgelegten Buch beschrieb Flacourt ein igelartiges Tier, das das halbe Jahr schlafen und sich dabei in den Boden eingraben würde. Zudem nähme es während dieser Zeit auch keine Nahrung auf. Flacourt bezeichnete das Tier mit „Tendrac“, gab aber als weiteren lokalen Namen „Sora“ an. Außerdem fügte er eine Zeichnung bei, die aus heutiger Sicht sehr grob gehalten ist.[79] Es dauerte dann gut ein Jahrhundert, bis sich Georges-Louis Leclerc de Buffon (1707–1788) mit den madagassischen Tieren beschäftigte. Buffon gab seit 1749 sein sehr umfassendes Werk Histoire naturelle, générale et particulière heraus, das zahlreiche Bände umfasst. Im zwölften Band aus dem Jahr 1764 beschrieb er den „Tendrac“ genauer. Außerdem stellte er ihm den „Tanrec“ zur Seite, der Buffons Aussagen zufolge größer ist als der „Tendrac“. Mit zwei beigehefteten Zeichnungen verbildlichte Buffon die Tiere, als Vorlage dafür und für die Beschreibungen dienten ihm Individuen aus dem Cabinet du roi, dem Vorgänger des Muséum national d’histoire naturelle in Paris. Diese waren über einem Händler aus Lyon in die Sammlung des königlichen Kabinetts gelangt.[80] Etwa 13 Jahre später veröffentlichte Johann Christian von Schreber (1739–1810) sein Werk Die Säugthiere in Abbildungen nach der Natur mit Beschreibungen. In diesem führte er den „Tendrac“ und den „Tanrec“ in deutscher Sprache ein, auch er fügte zwei Abbildungen der Tiere hinzu, die in ihrer Ausführung stark an die Zeichnungen aus Buffons Vorlage angelehnt sind. Zusätzlich vergab Schreber noch die wissenschaftlichen Artbezeichnungen Erinaceus setosus für den „Tendrac“ und Erinaceus ecaudatus für den „Tanrec“.[81] Dadurch steht der „Tendrac“ mit dem Großen Igeltenrek (heute Setifer setosus) und der „Tanrec“ mit dem Großen Tenrek (heute Tenrec ecaudatus) in Verbindung. Buffon hatte in zwei Zusatzbänden seiner Histoire naturelle, die 1776 und 1789 veröffentlicht wurden, noch zwei gestreifte Tenreks abgebildet und mit le jeune tanrec beziehungsweise mit le Tandrac angesprochen, diese werden heute aber als Individuen des Eigentlichen Streifentenreks und des Schwarzkopftenreks angesehen.[82][83]
Von den madagassischen Igeln zu den Tenreks
Das äußere Erscheinungsbild der bis zum Ende des 18. Jahrhunderts bekannten Tenrekvertreter – es handelte sich ausschließlich um solche der Igeltenreks – verleitete nahezu alle Forscher der damaligen Zeit, die Tiere als igelartig zu interpretieren und den Igeln zuzuweisen. Durch Schrebers Etablierung der wissenschaftlichen Namen für den Großen Tenrek und den Großen Igeltenrek wurden diese somit direkt zur Gattung Erinaceus und damit zu den europäischen Igeln (genauer zu den heutigen Kleinohrigeln) geordnet. Eine formale Trennung der Tenreks von der Gattung Erinaceus führte erstmals Georges Cuvier im Jahr 1798 durch, er gab aber keinen gültigen Gattungsnamen an.[84] Dies wird erst Bernard Germain Lacépède (1756–1825) ein Jahr darauf zugebilligt, der in einer taxonomischen Übersicht den Namen Tenrec angibt,[85] weitere Gattungsnamen sollten später folgen. Trotzdem verblieben die Tenreks vorerst innerhalb der Gruppe der Igel, etwa bei Étienne Geoffroy Saint-Hilaire 1803.[86]
Tenrecidae versus Centetidae

Der von Lacépède 1799 benannte wissenschaftliche Gattungsname Tenrec bildet die Grundlage für den heute gültigen Namen der Familie Tenrecidae. Dieser stammt wiederum von John Edward Gray (1800–1875) aus dem Jahr 1821. Gray formulierte die kurze Beschreibung der Familie folgendermaßen: Teeth, grinders, acutely tubercular; cutting, small, equal, six above, four below, canine long; body spinous; tail none. („Zähne, Mahlzähne, strikt tuberkular; schneidend, klein, gleichgroß, sechs oben, vier unter; Eckzahn lang, Körper stachelig; kein Schwanz.“).[87] Sowohl die Bezeichnung Tenrec für die Gattung als auch der Name Tenrecidae für die Familie wurden im Laufe des 19. Jahrhunderts selten gebraucht. Bereits 12 Jahre nach Lacépède etablierte Johann Karl Wilhelm Illiger den Gattungsnamen Centetes für den Großen Tenrek. Dieser entstammt der griechischen Sprache (κεντητήϛ kentetes) und leitet sich von κεντητος (kentetos) für „gestickt“ oder κεντεω (kenteo) für „stechen“ her.[88] Centetes stand wiederum Pate für den Familiennamen der Centetidae. Diesen hatte Andrew Murray im Jahr 1866 eingeführt, er vereinte darin alle damals bekannten Tenrekarten (Murray nannte Centetes, Ericulus und Echinogale; diese werden heute unter Tenrec, Setifer und Echinops geführt).[89] Murrays wissenschaftliche Bezeichnung für die Familie ging aber der von Charles Lucien Jules Laurent Bonaparte im Jahr 1838 geprägte Begriff Centetina voraus. Bonaparte führte Centetina zwar als Untergruppe innerhalb der Igel, trennte aber dadurch die Tenreks von den eigentlichen Igeln ab.[90] Die Bezeichnung Centetidae blieb über den Rest des 19. Jahrhunderts weit verbreitet und wurde auch in der ersten Hälfte des 20. Jahrhunderts verwendet. In den Jahren 1931 und 1945 wies George Gaylord Simpson in seinen Klassifikationsschemata der Säugetiere darauf hin, dass Tenrec der gegenüber Centetes ältere Gattungsname ist, auch wurde Tenrecidae vor Centetidae benannt. Dadurch sind die beiden später vergebenen Namen ungültig und nur als Synonyme aufzufassen.[91][92] Trotzdem blieben Centetes und Centetidae teilweise noch bis in die 1960er Jahre im Gebrauch.[21]
Die Tenreks als Insektenfresser
Die im ausgehenden 18. und im beginnenden 19. Jahrhundert angenommene nahe Verwandtschaft der Igel und der Tenreks führte dazu, dass sie auch in der übergeordneten Systematik zusammengeführt wurden. In seinem Werk von 1811 hatte Illiger die Gruppe der Subterranea („Unterirdische“) aufgestellt und darin sowohl die Igel und Tenreks als auch die Goldmulle, die Maulwürfe und die Spitzmäuse vereint,[88] er schuf damit ein heute nicht mehr anerkanntes Taxon, das alle die Formen vereinte, die später unter dem Begriff der „Insektenfresser“ geführt werden sollten. Erstmals informell wurden die Insektenfresser (Les Insectivores) von Cuvier 1817 vorgestellt, er unterteilte diese in zwei Gruppen: eine solche mit langen inneren Schneidezähnen, in die er die Goldmulle, Spitzmäuse und Igel stellte, und eine zweite mit großen Eckzähnen, etwa bei den Maulwürfen und den Tenreks.[93] Das Konzept wurde vier Jahre später von Thomas Edward Bowdich mit der Einführung des wissenschaftlichen Namens Insectivora übernommen, was die Anerkennung der Insektenfresser als Klassifizierungsgruppe zur Folge hatte.[94]
Im Verlauf des 19. und einem Großteil des 20. Jahrhunderts wurde das Konzept der Insektenfresser kaum in Frage gestellt. Neue Erkenntnisse bildeten die Grundlage für eine immer feinere Gliederung der Gruppe. Wilhelm Peters arbeitete im Jahr 1863 zwei namenlose Gruppen heraus, wobei ihm die weitaus größere durch das Fehlen eines Blinddarms auffiel, eine kleinere dagegen ein solches Organ besaß. Letztere umfasst die Rüsselspringer und die Spitzhörnchen, erstere den Rest der Insectivora einschließlich der Tenreks und der Goldmulle.[95] Später verlieh Ernst Haeckel im zweiten Band seines Werkes Generelle Morphologie der Organismen den beiden Gruppen die offiziellen Namen Lipotyphla (ohne Blinddarm) und Menotyphla (mit Blinddarm).[96] Einen gänzlich anderen Ansatz verfolgte Theodore Gill in den 1880er Jahren. Er unterschied die Insektenfresser nach der Ausprägung des Kaumusters der Mahlzähne. Demnach bestanden zwei Typen, einerseits die Zalambdodonta, die eine V-förmige Zahnschmelzleiste auf den Backenzähnen besitzen (von griechisch ζα (za-) für eine Übersteigerung und dem griechischen Buchstaben Λ (Lambda)), andererseits die Dilambdodonta mit einer W-förmigen Leiste (von griechisch δι (di „zwei“) und dem griechischen Buchstaben Λ (Lambda)). Die Tenreks gehörten nach Auffassung von Gill somit zu den zalambdodonten Insektenfressern, eine Position die sie mit den Goldmullen und den Schlitzrüsslern teilten. Alle anderen Insektenfresser formten demnach die dilambdodonte Gruppe. Damit erkannte Gill auch erstmals eine nähere Verwandtschaft der Tenreks mit den Goldmullen.[97]
Einen weiteren Gliederungsversuch unternahm William King Gregory im Jahr 1910 in seinem Werk The orders of Mammals, dabei stützte er sich teilweise auf die Arbeiten von Haeckel und Gill und kombinierte diese. So gliederte Gregory die Menotyphla aus den Insektenfressern aus, den Rest der Gruppe führte er unter der Bezeichnung Lipotyphla. Diese unterteilte er wiederum und behielt die Tenreks und Goldmulle innerhalb der Zalambdodonta nach Gill, während er die verbliebenen Dilambdodonta in die beiden Großgruppen Erinaceomorpha (Igel) und Soricomorpha (Spitzmäuse und Maulwürfe) aufteilte.[98] George Gaylord Simpson untergliederte in seiner 1945 erschienenen generellen Taxonomie der Säugetiere die Insektenfresser in fünf rezente Überfamilien, die Tenreks bildeten dabei die Tenrecoidea, die Goldmulle die Chrysochloroidea, Beide stellte er in relative Nähe zueinander.[92] Die engere Beziehung der Goldmulle und der Tenreks wurde auch später häufig thematisiert, so beispielsweise von Percy M. Butler im Jahr 1956. Butler übernahm teilweise das Systematikschema der Insektenfresser von Gregory, er unterschied aber nur die beiden Großgruppen der Erinaceomorpha und der Soricomorpha, die Tenreks und die Goldmulle ordnete er zu den Soricomorpha.[99] In den 1960er Jahren gliederte Leigh Van Valen die Zalambdodonta, also die Tenreks, Goldmulle und Schlitzrüssler, zwischenzeitlich aus den Insektenfressern aus und stellte sie in die von ihm geschaffene Ordnung der Deltatheridia. Darin gruppierte van Valen die Zalambdodonta den sogenannten Hyaenodonta gegenüber. Letztere bestanden aus mehreren ausgestorbenen Familien, darunter die Hyaenodontidae und die Oxyaenidae; in einer eher klassischen Sicht gehören diese zu der in sich nicht geschlossenen Gruppe der „Creodonta“ („Urraubtiere“).[100] Die Ansicht wurde aber weitgehend nicht übernommen. Nachfolgende Autoren wie Malcolm C. McKenna in den 1970er Jahren[101] und Michael J. Novacek in den 1980er Jahren[102] behielten die zalambdodonten Formen zumeist bei den Insektenfressern und dort bei den Soricomorpha. Im Bezug auf die Tenreks wiederholte McKenna diese Auffassung zusammen mit Susan K. Bell noch einmal in seinem Überblickswerk zur Systematik der Säugetiere in den 1990er Jahren, die Goldmulle hatte er hier aber aus den Soricomorpha ausgeschlossen und in eine eigene Ordnung platziert.[64]
Neue Einsichten
Erst im Übergang von 20. zum 21. Jahrhundert setzten sich mit der Entwicklung neuer Untersuchungsmethoden auch neue Erkenntnisse durch. Im Jahr 1998 veröffentlichte ein Arbeitsteam und Michael J. Stanhope erstmals molekulargenetische Untersuchungen zu den Insektenfressern. Dabei stellte sich heraus, dass die Insektenfresser keine in sich geschlossene Einheit bilden. Demnach stellen die Maulwürfe, Igel und Spitzmäuse eine gemeinsame Gruppe dar, die eine nähere Beziehung zu den Paar- und Unpaarhufern sowie zu den Schuppen-, Raub- und Fledertieren besitzen (die Gruppe erhielt später die Bezeichnung Laurasiatheria[103]). Die Tenreks dagegen formen eine Einheit mit den Goldmullen, was schon aus anatomischen Gründen vorher bekannt war, zusätzlich auch mit den Rüsselspringern, dem Erdferkel, den Elefanten, den Schliefern und den Seekühen. Da es sich hier hauptsächlich um endemisch in Afrika auftretende Tiergruppen handelt, gaben ihnen Stanhope und Kollegen die Bezeichnung Afrotheria. Den afrikanischen Zweig der ursprünglichen Insektenfresser, bestehend aus den Tenreks und Goldmullen, verwiesen sie gleichzeitig in eine eigenständige Ordnung innerhalb der Afrotheria, die Tenrekartigen (Afrosoricida; eigentlich übersetzt „afrikanische Spitzmausartige“).[104] Drei Jahre später wurden dann die Tenrekartigen, die Rüsselspringer und das Erdferkel in einem übergeordneten Taxon namens Afroinsectiphilia zusammengefasst.[57] In der Folgezeit konnten weitere Analysen das Ergebnis stützen, unter anderem durch die Isolierung eines spezifischen Retroposons, des sogenannten AfroSINEs, das alle Vertreter der Afrotheria gemein haben.[105][106] Demnach sind die Ähnlichkeiten der Tenreks und Goldmulle zu anderen Vertretern der Insektenfresser lediglich als Konvergenz aufzufassen.[104]
Stammesgeschichte
Zur Herkunft der madagassischen Tenreks
Die ältesten Fossilfunde der Tenreks stammen aus dem Eozän aus Afrika, von Madagaskar liegen nur subfossile Reste aus dem Holozän vor. Der Ursprung der Tenreks ist somit höchstwahrscheinlich in Afrika zu suchen. Madagaskar trennte sich von Afrika bereits vor rund 160 Millionen Jahren und besteht seit wenigstens 80 bis 90 Millionen Jahren als eigenständige Insel. Sie ist heute durch die etwa 400 km breite Straße von Mosambik vom Kontinent getrennt.[107] Gemäß den molekulargenetischen Daten gehen alle madagassischen Tenreks auf eine Ursprungsform zurück. Die Familie entstand der molekularen Uhr zufolge vor rund 48,3 Millionen Jahren, die Auffächerung der Tenreks setzte vor rund 35,6 Millionen Jahren ein. Etwa zwischen diesen beiden Eckpunkten kann die Besiedlung Madagaskars durch frühe Formen der Tenreks angenommen werden. Wie das geschah, ist unklar. Vorgeschlagen wurde die Möglichkeit einer zeitweiligen Landbrücke durch die Straße von Mosambik, eventuell entlang der Davie Fracture Zone, einer Störungszone vor der westlichen Küste der Insel. Die Landbrücke könnte infolge stärkerer seismischer Aktivität vom Eozän bis zum Unteren Miozän bestanden haben. Eine weitere Möglichkeit wäre das Überqueren der Wasserfläche auf Treibholz.[108][67][71][30]
Unklar ist die Anzahl der Einwanderungen der Tenreks nach Madagaskar. Ursprünglich wurde von mehreren Besiedlungswellen ausgegangen, was weitgehend auf die unklaren Verwandtschaftsverhältnisse der Tenreks untereinander im ausgehenden 20. Jahrhundert zurückzuführen ist.[66] Aus heutiger Sicht sind die rezenten Tenreks monophyletisch, womit eine einzelne Besiedlung ausreichend wäre. Problematisch sind aber die fossilen Formen aus dem frühen Miozän Afrikas, die aus anatomischer Sicht eine nähere Verwandtschaft zum Erdtenrek zeigen. Sollte dies zutreffen, ist die biogeographische Entwicklungsgeschichte der Tenreks komplizierter als eben dargestellt. Dann muss möglicherweise wenigstens eine zweite Wanderungsbewegung (vom Festland nach Madagaskar oder umgekehrt) angenommen werden. Diese Frage kann aber noch nicht beantwortet werden.[67][71][69]
Nach der Besiedlung Madagaskars vollzogen die Tenreks eine schnelle Diversifizierung. Ausgehend von den heutigen Vertretern war die Artbildungsrate in den feuchten Tropenwäldern des östlichen Inselteils besonders hoch. Anpassungen an die trockeneren Verhältnisse im Westen erfolgten mehrfach unabhängig sowohl in der Gruppe der Igeltenreks als auch unter den weichhaarigen Formen.[30] Im Zuge der Besiedlung Madagaskars durch die Tenreks kam es offenbar zu einer Reduktion oder einer „Verwaschung“ geschlechtsspezifischer Größenmerkmale, die bei kontinentalen Vertretern der Afrotheria teils stärker ausgeprägt sind. Die Ursachen hierfür – ob auf eine spezifische Gründerpopulation (Gründereffekt) oder auf Anpassungen an die Bedingungen einer isolierten Landmasse (ökologisch gesteuerte evolutionäre Entwicklung) zurückführbar – sind bisher ungeklärt. Ein ähnliches Phänomen liegt auch bei den Lemuren Madagaskars vor.[1] Lediglich die Igeltenreks nahmen im Laufe ihrer Entwicklungsgeschichte spürbar an Größe zu. Wohl verbunden damit und durch ihre stärker exponierte Lebensweise in den eher offeneren Landschaften entwickelte sich bei ihnen das stachelige Haarkleid. Die deutlich kleineren und weitgehend verborgen agierenden weichhaarigen Tenreks behielten hingegen ihre ursprüngliche Fellbedeckung.[26]
Eozän
Die bisher ältesten bekannten Fossilien von Tenreks kamen im nördlichen Sperrgebiet in Namibia zum Vorschein. Es handelt sich um zahlreiche Schädel- und Kieferreste sowie Teile des Bewegungsapparates aus der Fundstelle Eocliff, einer Travertinlagerstätte, die sich im Oberen Eozän an der Austrittstelle einer kalkhaltigen Quelle gebildet hatte. Die Funde repräsentieren zwei Gattungen, einerseits Sperrgale, andererseits Arenagale. Bei beiden Formen handelt es sich um kleine Vertreter der Tenreks, etwa von der Größe des heutigen Erdtenreks. Sie werden durch eine primitivere Bezahnung charakterisiert, bei der das zalambdodonte Kauflächenmuster durch den deutlich vom Paraconus abgetrennten Metaconus der Oberkiefermolaren noch nicht ganz so deutlich ausgeprägt ist. Der Bewegungsapparat lässt annehmen, dass die Gliedmaßen sehr mobil eingesetzt werden konnten, Hinweise auf eine grabende Lebensweise gibt es dadurch noch nicht.[75][109] Ebenfalls aus dem Sperrgebiet liegt ein Unterkieferast von Nanogale vor. Er kam in den Kalksteinlagen der Fundstelle Black Crow zu Tage, die etwas älter als das Eocliff sind. Morphologisch lässt sich Nanogale durch die Gestaltung des aufsteigenden Astes am Unterkiefer nicht direkt mit den Tenreks verbinden, sondern steht im weiteren Beziehungsumfeld, den Tenrecomorpha. Mit einer Länge von nur 8,6 mm ist der Unterkiefer aber vergleichbar zu dem der kleinsten bekannten, nicht zu den Fledertieren gehörenden Säugetiere, wie etwa zu verschiedenen Vertretern der Dickschwanzspitzmäuse.[110]
Oligozän
Im weiteren Verlauf des Eozän und im Unteren Oligozän sind keine direkten Angehörigen der Tenreks nachgewiesen, allerdings können einige nahe verwandte Formen belegt werden. Von Bedeutung hier ist die Fossillagerstätte Fayyum im nördlichen Ägypten. Aus der Qasr-el-Sagha-Formation und damit noch aus dem obersten Abschnitt des Eozän stammt Dilambdogale, das im Jahr 2010 beschrieben wurde. Weitere Formen wurden mit Widanelfarasia, Jawharia und Qatranilestes aus der Gebel-Qatrani-Formation geborgen und gehören dem Oligozän an. Alle vier Vertreter sind weitgehend nur durch Unter- und Oberkieferfragmente bekannt, in den wenigsten Fällen liegt auch noch postcraniales Skelettmaterial vor. Diese relativ urtümlichen Vertreter besaßen an den oberen Molaren ebenfalls einen getrennten Para- und Metaconus, zusätzlich war auf dem vordersten Mahlzahn ein W-förmiges Schmelzleistenmuster ausgebildet, was an die dilambdodonte Kauflächenstruktur der Insektenfresser erinnert. Die hinteren Molaren wiesen dagegen ein zalambdodontes Muster auf, womit die Formen eine Übergangsstellung zu den typischen Afrosoricida einnehmen.[111][112][113][74]
Miozän
Erst im folgenden Miozän sind Tenreks wieder eindeutig belegt. Der Großteil der bisher entdeckten Fossilreste stammt aus Ostafrika. Sie wurden unter anderem in der Legetet-Formation, in der Hiwegi-Formation und in den Kapurtay-Agglomeraten gefunden, deren Alter etwa 18 bis 20 Millionen Jahre beträgt. Insgesamt sind in den einzelnen Aufschlüssen drei Gattungen überliefert. Parageogale, belegt über einen Teilschädel und mehrere Gebissfragmente, stellt möglicherweise einen nahen Verwandten des Erdtenreks dar. Er unterscheidet sich von der rezenten Form durch seine geringfügig größeren Ausmaße und durch sein noch stärker reduziertes Gebiss. Im Schädelbau zeigt er dagegen einige urtümliche Merkmale. Erythrozootes wurde anhand eines Teilschädels beschrieben, zusätzlich sind noch mehrere Gebiss- und Kieferfragmente bekannt. Die Form war größer als die Kleintenreks. Auffälligste Kennzeichen sind die sehr großen vorderen oberen Schneidezähne, ein ausgesprochen gut entwickelter Protoconus an den Oberkiefermolaren und ein wenig reduziertes Talonid an den Mahlzähnen des Unterkiefers. In diesen Merkmalen stimmt Erythrozootes mit der dritten Fossilgattung, Protenrec, überein, allerdings war letztere deutlich kleiner und besaß neben einem umfangreicheren Gebiss verschiedene abweichende Schädelmerkmale, etwa den vergleichsweise längeren Kanal des Foramen infraorbitale oder das sehr weit nach vorn verlagerte Foramen lacrimale. Von Protenrec sind bisher mehrere Unterkieferreste und ein vorderer Schädelteil dokumentiert worden, herausragend ist die Fundstelle Songhor im westlichen Kenia.[114][115][8] Zudem kamen von der Gattung auch einzelne Unterkiefer- und Schädelteile in Arrisdrift sowie in Elisabethfeld und Langental im Sperrgebiet von Namibia zu Tage. Die Tiere waren im Durchschnitt etwas kleiner als jene aus Ostafrika. Die Fundstellen datieren in das Untere und in das beginnende Mittlere Miozän.[116][117] Im Gegensatz zu Parageogale sind die verwandtschaftlichen Verhältnisse für Erythrozootes und Protenrec schwieriger zu bestimmen; sie werden mitunter in die eigene Unterfamilie der Protenrecinae gestellt, die aber möglicherweise ebenfalls im nahen Verhältnis zum Erdtenrek steht.[8][69][74] Bereits aus dem Unteren Miozän ist von Elisabethfeld in der gleichen Region ein bisher einzelner Unterkiefer von Promicrogale aufgedeckt worden. Die Gattung weist Beziehungen zu den Kleintenreks auf.[76]
Plio- und Pleistozän
Weder aus dem Pliozän noch aus dem Pleistozän sind bisher Funde von Tenreks in Afrika oder auf Madagaskar überliefert.[74]
Holozän
Der früheste Fossilnachweis der Tenreks auf Madagaskar datiert in das Holozän, wobei die meisten Funde jünger als 2000 Jahre sind. Knochenreste von Vertretern der Familie fanden sich in mehreren Höhlenfundstellen. Zu den bedeutendsten gehören die Anjohibe-Höhle[118] im nordwestlichen, die Ankilitelo-Höhle[119] im südwestlichen und die Andrahomana-Höhle[120] im südöstlichen Teil der Insel. Alle drei zeichnen sich durch einen außerordentlichen Fossilreichtum an Säugetieren aus: so sind Fledertiere, Nagetiere, Raubtiere oder Primaten vertreten. Die Tenreks sind mit rund einem Dutzend Arten repräsentiert, die sowohl zu den Igeltenreks als auch zu den Erdtenreks und zu den Reistenreks verwiesen werden können. Darunter befinden sich einige Arten wie der Major-Langschwanz-Kleintenrek oder der Nasolo-Kleintenrek, deren gegenwärtiges Verbreitungsgebiet deutlich eingeschränkt ist, so dass sie in der entsprechenden Fundregion nicht mehr vorkommen. Außerdem konnte mit Microgale macpheei auch ein ausgestorbener Angehöriger nachgewiesen werden; er ist bisher nur von der Andrahomana-Höhle dokumentiert.[121] Neben den eigentlichen Tenreks trat im Verlauf des Holozäns zudem die nah verwandte Form Plesiorycteropus mit wenigstens zwei Arten auf. Sie ist von wenigstens einem Dutzend Fundstellen bekannt, die sich vorwiegend auf das zentrale und westliche Madagaskar verteilen.[62] Das Aussterben einzelner Angehöriger der Tenreks beziehungsweise naher verwandter Formen, aber auch der Rückzug mancher Arten aus dem ehemaligen Verbreitungsgebiet hat verschiedene Gründe. Diese stehen einerseits mit der zunehmenden Austrocknung des westlichen Inselteils in den letzten zwei bis drei Tausend Jahren, andererseits mit der Ankunft des Menschen in diesem Zeitraum in Zusammenhang.[122]
Bedrohung und Schutz
Die größte Bedrohung für den Bestand der einzelnen Tenrekarten wie auch für viele andere waldbewohnende Tierarten Madagaskars ist die fortschreitende Zerstörung ihres Lebensraums. Die IUCN listet sechs Arten als „stark gefährdet“ (endangered) oder „gefährdet“ (vulnerable), dabei handelt es sich ausschließlich um Vertreter der Kleintenreks mit deutlich begrenztem Verbreitungsgebiet. Eine weitere Art aus der Gattung der Reiswühler wird aufgrund fehlender Informationen in der Kategorie „unzureichende Datenlage“ (data deficient) eingestuft. Die Igeltenreks hingegen sind weit verbreitet und haben sich als Kulturfolger an die Nähe des Menschen gewöhnt. Sie gelten daher gegenwärtig als nicht bedroht. Allerdings sind sie aufgrund ihrer Größe wiederum stärker der Jagd durch den Menschen ausgesetzt. Ihr Fleisch wird meist als Nahrungsressource verwendet. So gut wie alle Tenrekarten kommen in Naturschutzgebieten vor.[123]
Literatur
- J. F. Eisenberg und Edwin Gould: The Tenrecs: A Study in Mammalian Behavior and Evolution. Smithsonian Institution Press, 1970, S. 1–138
- Nick Garbutt: Mammals of Madagascar. A complete guide. Yale University Press, 2007, S. 1–304 (S. 32–56)
- Konrad Herter: Über die Borstenigel von Madagaskar (Tenrecinae). Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin N. F. 2, 1962, S. 5–37
- Paulina D. Jenkins: Tenrecidae (Tenrecs and Shrew tenrecs). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos. Lynx Edicions, Barcelona 2018, S. 134–172 ISBN 978-84-16728-08-4
- Ronald M. Nowak: Walker's Mammals of the World. The Johns Hopkins University Press, Baltimore 1999, ISBN 0-8018-5789-9.
- Link E. Olson: Tenrecs. Current Biology 23 (1), 2013, S. R5–R8
Einzelnachweise
- Peter M. Kappeler, Charles L. Nunn, Alexander Q. Vining und Steven M. Goodman: Evolutionary dynamics of sexual size dimorphism in nonvolant mammals following their independent colonization of Madagascar. Scientific Reports 9, 2019, S. 1454 doi:10.1038/s41598-018-36246-x
- Nick Garbutt: Mammals of Madagascar. A complete guide. Yale University Press, 2007, S. 1–304 (S. 32–56)
- Konrad Herter: Über die Borstenigel von Madagaskar (Tenrecinae). Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin N. F. 2, 1962, S. 5–37
- J. F. Eisenberg und Edwin Gould: The Tenrecs: A Study in Mammalian Behavior and Evolution. Smithsonian Institution Press, 1970, S. 1–138
- Sive Finlay and Natalie Cooper: Morphological diversity in tenrecs (Afrosoricida, Tenrecidae): comparing tenrec skull diversity to their closest relatives. PeerJ 3, 2015, S. e927 doi:10.7717/peerj.927
- Wilhelm Leche: Zur Entwicklungsgeschichte des Zahnsystems der Säugetiere, zugleich ein Beitrag zur Stammesgeschichte dieser Tiergruppe. Zweiter Teil: Phylogenie. Zweites Heft: Familien der Centetidae, Solenodontidae und Chrysochloridae. Zoologica 20, 1906/1908, S. 1–157 ()
- Steven M. Goodman und Paulina D. Jenkins: The Insectivores of the Réserve Spéciale d’Anjanaharibe-Sud, Madagascar. Fieldiana Zoology 90, 1998, S. 139–161
- Percy M. Butler: Macroscelidea, Insectivora and Chiroptera from the Miocene of East Africa. Palaeovertebrata 14 (3), 1984, S. 117–200
- Robert J. Asher und Marcelo R. Sánchez-Villagra: Locking Yourself Out: Diversity Among Dentally Zalambdodont Therian Mammals. Journal of Mammalian Evolution. 12 (1/2), 2005, S. 265–282
- Link E. Olson: Tenrecs. Current Biology 23 (1), 2013, S. R5–R8
- Justine A. Salton und Frederick S. Szalay: The Tarsal Complex of Afro-Malagasy Tenrecoidea: A Search for Phylogenetically Meaningful Characters. Journal of Mammalian Evolution 11 (2), 2004, S. 73–104
- Justine A. Salton und Eric J. Sargis: Evolutionary morphology of the Tenrecoidea (Mammalia) carpal complex. Biological Journal of the Linnean Society, 93, 2008, S. 267–288
- Justine A. Salton und Eric J. Sargis: Evolutionary Morphology of the Tenrecoidea (Mammalia) Forelimb Skeleton. In: E.J. Sargis und M. Dagosto (Hrsg.): Mammalian Evolutionary Morphology: A Tribute to Frederick S. Szalay, Springer Science, 2008, S. 51–71
- Justine A. Salton und Eric J. Sargis: Evolutionary Morphology of the Tenrecoidea (Mammalia) Hindlimb Skeleton. Journal of Morphology 270, 2009, S. 367–387
- Hideko Endo, Takahiro Yonezawa, Felix Rakotondraparany, Motoki Sasaki und Masami Hasegawa: The adaptional strategies of the hindlimb muscles in the Tenrecidae species including the aquatic web-footed tenrec (Limnogale mergulus). Annals of Anatomy 188, 2006, S. 383–390
- H. Endo, M. Oishi, T. Yonezawa, F. Rakotondraparany und M. Hasegawa: The Semifossorial Function of the Forelimb in the Common Rice Tenrec (Oryzorictes hova) and the Streaked Tenrec (Hemicentetes hemispinosus). Anatomie, Histology, Embryology 36, 2007, S. 413–418
- Petr Cizek, Pavla Hamouzova, Karolina Goździewska‑Harłajczuk, Joanna Klećkowska‑Nawrot und Pavel Kvapil: Microscopic structure of the tongue in the lesser hedgehog tenrec (Echinops telfairi, Afrosoricida) and its relation to phylogenesis. Anatomical Science International 95, 2020, S. 313–322, doi:10.1007/s12565-019-00522-1
- B. Riedelsheimer, Pia Unterberger, H. Künzle und U. Welsch: Histological study of the cloacal region and associated structures in the hedgehog tenrec Echinops telfairi. Mammalian Biology 72, 2007, S. 330–341
- Peter Langer: The digestive tract and life history of small mammals. Mammal Review 32 (2), 2002, S. 107–131
- J. J. Petter und A. Petter-Rousseaux: Notes biologiques sur les Centetinae. La Terre et la Vie 17 (1), 1963, S. 66–80
- J. Michael Bedford, Orin B. Mock und Steven M. Goodman: Novelties of conception in insectivorous mammals (Lipotyphla), particularly shrews. Biological Review 79, 2004, S. 1–19
- A. C. Enders, T. N. Blankenship, S. M. Goodman, V. Soarimalala und A. M. Carter: Placental Diversity in Malagasy Tenrecs: Placentation in Shrew Tenrecs (Microgale spp.), The Mole-Like Rice Tenrec (Oryzorictes hova) and The Web-Footed Tenrec (Limnogale mergulus). Placenta 28, 2007, S. 748–759
- A. M. Carter, T. N. Blankenship, H.Künzle und A. C. Enders: Structure of the Definitive Placenta of the Tenrec, Echinops telfairi. Placenta 25, 2004, S. 218–232
- Jason A. Kaufman, Gregory H. Turner, Patricia A. Holroyd, Francesco Rovero und Ari Grossman: Brain Volume of the Newly-Discovered Species Rhynchocyon udzungwensis (Mammalia: Afrotheria: Macroscelidea): Implications for Encephalization in Sengis. PlosOne 8 (3), 2013, S. e58667 doi:10.1371/journal.pone.0058667
- Theodore Stankowich und Colin Stensrud: Small but spiny: the evolution of antipredator defenses in Madagascar tenrecs. Journal of Mammalogy 100 (1), 2019, S. 13–20, doi:10.1093/jmammal/gyz003
- Leah Krubitzer, Heinz Künzle und Jon Kaas: Organization of Sensory Cortex in a Madagascan Insectivore, the Tenrec (Echinops telfairi). Journal of Comparative Neurology 379, 1997, S. 399–414
- Susanne Radtke-Schuller und Heinz Künzle: Olfactory bulb and retrobulbar regions in the hedgehog tenrec: Organizations and interconnections. Journal of Comparative Neurology 423 (4), 2000, S. 687–705.
- Heinz Künzle: Thalamo-striatal projections in the hedgehog tenrec. Brain Research 1100, 2006, S. 78–92
- Kathryn M. Everson, Voahangy Soarimalala, Steven M. Goodman und Link E. Olson: Multiple loci and complete taxonomic sampling resolve the phylogeny and biogeographic history of tenrecs (Mammalia: Tenrecidae) and reveal higher speciation rates in Madagascar’s humid forests. Systematic Biology 65 (5), 2016, S. 890–909 doi: 10.1093/sysbio/syw034
- Claudette Patricia Maminirina, Steven M. Goodman und Christopher J. Raxworthy: Les microammifères (Mammalia, Rodentia, Afrosoricida et Soricomorpha) du massif du Tsaratanana et biogéographie des forêts de montagne de Madagascar. Zoosystema 30 (3), 2008, S. 695–721
- Melanie Dammhahn, Voahangy Soarimalala und Steven M. Goodman: Trophic Niche Differentiation and Microhabitat Utilization in a Species-rich Montane Forest Small Mammal Community of Eastern Madagascar. Biotropica 45 (1), 2013, S. 111–118
- R. D. E. MacPhee: The Shrew Tenrecs of Madagascar: Systematic Revision and Holocene Distribution of Microgale (Tenrecidae, Insectivora). American Museum Novitates 2889, 1987, S. 1–45
- Edwin Gould: Evidence for echolocation in the Tenrecidae of Madagaskar. Proceedings of the American Philosophical Society 109 (6), 1965, S. 352–360
- Olga V. Zherebtsova: Morphofunctional interpretation of the quills stridulating in tenrecs (Lipotyphla, Tenrecidae). Russian Journal of Theriology 5 (1), 2006, S. 1–11
- Danielle L. Levesque, D. Rakotondravony und Barry G. Lovegrove: Home range and shelter site selection in the greater hedgehog tenrec in the dry deciduous forest of Western Madagascar. Journal of Zoology 287, 2012, S. 161–168
- Walter Poduschka: Augendrüsensekretion bei den Tenreciden Setifer setosus (Froriep 1806), Echinops telfairi (Martin 1838); Microgale dobsoni (Thomas 1918) und M. talazaci (Thomas 1918). Zeitschrift für Tierpsychologie 35, 1974, S. 303–319
- Edwin Gould und John F. Eisenberg: Notes on the biology of the Tenrecidae. Journal of Mammalogy 47 (4), 1966, S. 660–686
- Jonathan Benstead, Kevin H. Barnes und Catherine M. Pringle: Diet, acvtivity patterns, foraging movements and response to deforestation of the aquatic tenrec Limnogale mergulus (Lipotyphla: Tenrecidae) in eastern Madagascar. Journal of Zoology 254, 2001, S. 119–129
- Peter J. Stephenson und Paul A. Racey: Reproductive energetics of the Tenrecidae (Mammalia: Insectivora). I. Large-eared tenrec, Geogale aurita. Physiological Zoology 66 (5), 1993, S. 643–663
- Peter J. Stephenson und Paul A. Racey: Reproductive energetics of the Tenrecidae (Mammalia: Insectivora). II. The shrew-tenrecs, Microgale spp. Physiological Zoology 66 (5), 1993, S. 664–685
- P. J. Stephenson, J. R. Speakman und P. A. Racey: Field metabolic rate in two species of shrew-tenrec, Microgale dobsoni and Microgale talazaci. Comparative Biochemistry and Physiology 107A (2), 1994, S. 283–287
- Paul A. Racey und Peter J. Stephenson: Reproductive and energetic differentiation of the Tenrecidae of Madagascar. In: W.R Lourenço (Hrsg.): Biogéographie de Madagascar. Paris, 1996, S. 307–319
- Danielle L. Levesque, Kerileigh D. Lobban und·Barry G. Lovegrove: Effects of reproductive status and high ambient temperatures on the body temperature of a free‑ranging basoendotherm. Journal of Comparative Physiology B, 2014 doi:10.1007/s00360-014-0858-4
- Barry G. Lovegrove, Cindy Canale, Danielle Levesque, Gerhard Fluch, Milada Řeháková-Petrů und Thomas Ruf: Are Tropical Small Mammals Physiologically Vulnerable to Arrhenius Effects and Climate Change? Physiological and Biochemical Zoology 87 (1), 2014, S. 30–45
- Danielle Danielle L. Levesque, O. M.A. Lovasoa, S. N. Rakotoharimalala und Barry G. Lovegrove: High mortality and annual fecundity in a free-ranging basal placental mammal, Setifer setosus (Tenrecidae: Afrosoricida). Journal of Zoology 291, 2013, S. 205–212
- Barry G. Lovegrove, Kerileigh D. Lobban und Danielle L. Levesque: Mammal survival at the Cretaceous-Palaeogene boundary: metabolic homeostasis in prolonged tropical hibernation in tenrecs. Proceedings of the Royal Society B 281, 2014, S. 20141304 doi:10.1098/rspb.2014.1304
- Michael D. Treat, Lori Scholer, Brandon Barrett, Artur Khachatryan, Austin J. McKenna, Tabitha Reyes, Alhan Rezazadeh, Charles F. Ronkon, Dan Samora, Jeremy F. Santamaria, Claudia Silva Rubio, Evan Sutherland, Jeffrey Richardson, John R. B. Lighton und Frank van Breukelen: Extreme physiological plasticity in a hibernating basoendothermic mammal, Tenrec ecaudatus. Journal of Experimental Biology 221, 2018, S. jeb185900 doi:10.1242/jeb.185900
- Rebecca Oelkrug, Nadja Goetze, Cornelia Exner, Yang Lee, Goutham K. Ganjam, Maria Kutschke, Saskia Müller, Sigrid Stöhr, Matthias H. Tschöp, Paul G. Crichton, Gerhard Heldmaier, Martin Jastroch und Carola W. Meyer: Brown fat in a protoendothermic mammal fuels eutherian evolution. Nature Communication 4, 2013, S. 2140 doi:10.1038/ncomms3140
- H. Bluntschli: Über die Frühentwicklung eines Centetinen (Hemicentetes semispinosus Cuv.). Revue Suisse de Zoologie 44, 1937, S, 271–282 ()
- Fritz Strauß: Die Befruchtung und der Vorgang der Ovulation bei Ericulus aus der Familie der Centetiden. Biomorphosis 1, 1938, S. 281–312
- Martin Nicoll und P. A. Racey: Follicular development, ovulation, fertilization and fetal development in tenrecs (Tenrec ecaudatus). Journal of Reproduction and Fertility 74 (1), 1985, S. 47–55
- Martin Nicoll: The common tenrec, Tenrec ecaudatus. Afrotherian Conservation 7, 2009, S. 2–3
- H. Künzle, C. Poulsen Nautrup und F. Schwarzenberger: High inter-individual variation in the gestation length of the hedgehog tenrec, Echinops telfairi (Afrotheria). Animal Reproduction Science 97, 2007, S. 364–374
- Walter Poduschka: Hyperthelie, Wurfgröße und Trächtigkeitsdauer bei der Unterfamilie Tenrecinae Cabrera, 1925 (Mammalia: Insectivora: Tenrecidae), nebst Bemerkungen zur Längsstreifung der Gattung Hemicentetes. Contributions to Zoology 66 (2), 1996, S. 119–128
- Steven Heritage, Erik R. Seiffert und Matthew R. Borths: Recommended fossil calibrators for time-scaled molecular phylogenies of Afrotheria. Afrotherian Conservation 17, 2021, S. 9–13
- Peter J. Waddell, Hirohisa Kishino und Rissa Ota: A Phylogenetic Foundation for Comparative Mammalian Genomics. Genome Informatics 12, 2001, S. 141–154
- Matjaž Kuntner, Laura J. May-Collado und Ingi Agnarsson: Phylogeny and conservation priorities of afrotherian mammals (Afrotheria, Mammalia). Zoologica Scripta 40 (1), 2011, S. 1–15
- Robert W. Meredith, Jan E. Janečka, John Gatesy, Oliver A. Ryder, Colleen A. Fisher, Emma C. Teeling, Alisha Goodbla, Eduardo Eizirik, Taiz L. L. Simão, Tanja Stadler, Daniel L. Rabosky, Rodney L. Honeycutt, John J. Flynn, Colleen M. Ingram, Cynthia Steiner, Tiffani L. Williams, Terence J. Robinson, Angela Burk-Herrick, Michael Westerman, Nadia A. Ayoub, Mark S. Springer und William J. Murphy: Impacts of the Cretaceous Terrestrial Revolution and KPg Extinction on Mammal Diversification. Science 334, 2011, S. 521–524
- Erik R Seiffert: A new estimate of afrotherian phylogeny based on simultaneous analysis of genomic, morphological, and fossil evidence. BMC Evolutionary Biology 7, 2007, S. 224 doi:10.1186/1471-2148-7-224
- Bryan Patterson: The fossil aardvarks (Mammalia: Tubulidentata). Bulletin of the Museum of Comparative Zoology 147, 1975, S. :185–237
- Ross D. E. MacPhee: Morphology, adaptations, and relationships of Plesiorycteropus, and a diagnosis of a new order of eutherian mammals. Bulletin of the American Museum of Natural History 220, 1994, S. 1–214
- Michael Buckley: A Molecular Phylogeny of Plesiorycteropus Reassigns the Extinct Mammalian Order ‘Bibymalagasia’. PlosOne 8 (3), 2013, S. e59614 doi:10.1371/journal.pone.0059614
- Malcolm C. McKenna und Susan K. Bell: Classification of mammals above the species level. Columbia University Press, New York, 1997, S. 1–631 (S. 293–295)
- Don E. Wilson und DeeAnn M. Reeder: Mammal Species of the World. A taxonomic and geographic Reference. Johns Hopkins University Press, 2005 ()
- Robert J. Asher: A morphological basis for assessing the phylogeny of the „Tenrecoidea“ (Mammalia, Lipotyphla). Cladistics 15, 1999, S. 231–252
- Christophe J. Douady, Francois Catzeflis, Diana J. Kao, Mark S. Springer und Michael J. Stanhope: Molecular Evidence for the Monophyly of Tenrecidae (Mammalia) and the Timing of the Colonization of Madagascar by Malagasy Tenrecs. Molecular Phylogenetics and Evolution 22 (3), 2002, S. 357–363
- Christophe J. Douady, Francois Catzeflis, Mark S. Springer und Michael J. Stanhope: Molecular evidence for the monophyly of Tenrecidae: a reply to Asher. Molecular Phylogenetics and Evolution 26, 2003, S. 331–332
- Robert J. Asher und Robert Hofreiter: Tenrec Phylogeny and the Noninvasive Extraction of Nuclear DNA. Systematic Biology 55 (2), 2006, S. 181–194
- Paulina D. Jenkins: Tenrecidae (Tenrecs and Shrew tenrecs). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos. Lynx Edicions, Barcelona 2018, S. 134–172 ISBN 978-84-16728-08-4
- Céline Poux, Ole Madesen, Elisabeth Marquard, David R. Vieites, Wilfried W. de Jong und Miguel Vences: Asynchronous Colonization of Madagascar by the Four Endemic Clades of Primates, Tenrecs, Carnivores, and Rodents as Inferred from Nuclear Genes. Systematic Biology 54 (5), 2005, S. 719–730
- Céline Poux, Ole Madesen, Julian Glos, Wilfried W. de Jong und Miguel Vences: Molecular phylogenetic and divergence times of Malagasy tenrecs: Influence of data partitioning and taxon sampling on dating analyses. BMC Evolutionary Biology, 2008, S. 102
- Link E. Olson und Steven M. Goodman: Phylogeny and biogeography of tenrecs. In: Steven M. Goodman und Jonathan P. Benstead (Hrsg.): The natural history of Madagascar. University of Chicago Press, 2003, S. 1235–1242
- Robert J. Asher: Tenrecoidea. In: Lars Werdelin und William Joseph Sanders (Hrsg.): Cenozoic Mammals of Africa. University of California Press, Berkeley, London, New York, 2010, S. 99–106
- Martin Pickford: Late Eocene Potamogalidae and Tenrecidae (Mammalia) from the Sperrgebiet, Namibia. Communications of the Geological Survey of Namibia 16, 2015, S. 114–152
- Martin Pickford: Tenrecoid mandible from Elisabethfeld (Early Miocene) Namibia. Communications of the Geological Survey of Namibia 18, 2018, S. 87–92
- Louis L. Jacobs, William Anyonge und John C. Barry: A giant tenrecid from the Miocene of Kenya. Journal of Mammalogy 68 (1), 1987, S. 10–16
- Jorge Morales, Martin Pickford, Manuel Salesa und Dolores Soria The systematic status of Kelba, Savage, 1965, Kenyalutra, Schmidt-Kittler, 1987 and Ndamathaia, Jacobs et al., 1987, (Viverridae, Mammalia) and a review of Early Miocene mongoose-like carnivores of Africa. Annales de Paléontologie 86 (4), 2000, S. 243–251
- Étienne de Flacourt: Histoire de la grande isle Madagascar. Paris, 1661, S. 1–471 (S. 152) ()
- Georges-Louis Leclerc de Buffon: Histoire naturelle, générale et particulière., Tome Douzième. Paris, 1764, S. 440–441, S. 450 und Tafeln 56 und 57 (), Supplément, Tome troisième. Paris, 1776, Tafel 37 () und Supplément, Tome Septième. Paris, 1789, Tafel 76 ()
- Johann Christian Daniel Schreber: Die Säugthiere in Abbildungen nach der Natur mit Beschreibungen. Dritter Teil. Erlangen, 1777, S. 584 () und Tafelband 1 Tafel 164–166 ()
- Fredericus Anna Jentink: On the hedgehogs from Madagascar. Notes from the Royal Zoological Museum at Leyden 1879, S. 137–151 ()
- Christopher D. Marshall und John F. Eisenberg: Hemicentetes semispinosus. Mammalian Species 541, 1996, S. 1–4
- Georges Cuvier: Tableau elementaire de l’histoire naturelle des animaux. Paris, 1798, S. 1–710 ( 108) ()
- Bernard Germain Lacépède: Tableau des divisions, sous-divisoins, ordres et genres des mammifères. Paris, 1799; Nachdruck in den Memoirs de l'Institut national des sciences et arts 3, 1801, S. 489–519 (S. 492) ()
- Étienne Geoffroy Saint-Hilaire: Catalogue des mammifères du Muséum National d'Histoire Naturelle. Paris, 1803, S. 1–272 (S. 71) ()
- John Edward Gray: On the natural arrangement of vertebrose animals. London Medical Repository 15, 1821, S. 297–310 (301) ()
- Johann Karl Wilhelm Illiger: Prodromus systematis mammalium et avium additis terminis zoographicis utriudque classis. Berlin, 1811, S. 1–301 (S. 124) ()
- Andrew Murray: The geographic distribution of mammals. London, 1866, S. 1–420 (S. XIV) ()
- Charles Lucien Jules Laurent Bonaparte: Synopsis vertebratorum systematis. Nuovi Annali delle Scienze Naturali 2, 1838, S. 105–133 ()
- George Gaylord Simpson: A new classification of mammals. Bulletin of the American Museum of Natural History 59 (5), 1931, S. 259–293
- George Gaylord Simpson: The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History 85, 1945, S. 1–350 (S. 48–49 und 175–179)
- Georges Cuvier: Le regne animal distribué d'apres son organisation pour servir de base a l'histoire naturelle des animaux et d'introduction à l'anatomie comparée. Paris, 1817, S. 1–540 (S. 131–138) ()
- Thomas Edward Bowdich: An analysis of the natural classifications of Mammalia for the use of students and travellers. Paris, 1821, S. 1–115 (S. 31–33) ()
- Wilhelm Peters: Über die Säugethirgattung Selenodon. Abhandlungen der Königlichen Akademie der Wissenschaften zu Berlin 1863, S. 1–22 ()
- Ernst Haeckel: Generelle Morphologie der Organismen. Band 2. Berlin, 1866, S. 1–462 (S. CLX)()
- Theodore Gill: Insectivora. In: John Sterling Kingsley (Hrsg.): Standard natural history. Volume 5. Boston, 1885, S. 1–535 (S. 134–158) ()
- William King Gregory: The orders of mammals. Bulletin of the American Museum of Natural History 27, 1910, S. 1–524 (S. 464–465)
- Percy M. Butler: The skull of Ictops and the classification of the Insectivora. Proceedings of the Zoological Society of London 126, 1956, S. 453–481
- Leigh van Valen: New Paleocene Insectivores and Insectivore classification. Bulletin of the American Museum of Natural History 135, 1967, S. 217–284
- Malcolm C. McKenna: Toward a phylogenetic classification of the Mammalia. In W. Patrick Luckett und Frederick S. Szalay (Hrsg.): Phylogeny of the primates: a multidisciplinary approach. New York, London, 1975, S. 21–46
- Michael J. Novacek: The skull of leptictid insectivorans and the higher-level classification of eutherian mammals. Bulletin of the American Museum of Natural History 183, 1986, S. 1–111
- Peter J. Waddell, Norihiro Okada und >Masami Hasegawa: Towards Resolving the Interordinal Relationships of Placental Mammals. Systematic Biology 48 (1), 1999, S. 1–5
- Michael J. Stanhope, Victor G. Waddell, Ole Madsen, Wilfried de Jong, S. Blair Hedges, Gregory C. Cleven, Diana Kao und Mark S. Springer: Molecular evidence for multiple origins of Insectivora and for a new order of endemic African insectivore mammals. PNAS 95, 1998, S. 9967–9972 ()
- Masato Nikaido, Hidenori Nishihara, Yukio Hukumoto und Norihiro Okada: Ancient SINEs from African Endemic Mammals. Molecular Biology and Evolution 20 (4), 2003, S. 522–527
- Hidenori Nishihara, Yoko Satta, Masato Nikaido, J. G. M. Thewissen, Michael J. Stanhope und Norihiro Okada: A Retroposon Analysis of Afrotherian Phylogeny. Molecular Biology and Evolution 22 (9), 2005, S. 1823–1833
- Anne D. Yoder und Michael D. Nowak: Has Vicariance or Dispersal Been the Predominant Biogeographic Force in Madagascar? Only Time Will Tell. Annual Review of Ecology, Evolution, and Systematics 37 (1), 2006, S. 405–431 ()
- Robert A. McCall: Implication of recent geological investigations of the Mozambique Channel for the mammalian colonization of Madagaskar. Proceedings of the Royal Society of London 264, 1997, S. 663–665
- Robert J. Asher: Recent additions to the fossil record of tenrecs and golden moles. Afrotherian Conservation 15, 2019, S. 4–13
- Martin Pickford: Tiny Tenrecomorpha (Mammalia) from the Eocene of Black Crow, Namibia. Communications of the Geological Survey of Namibia 21, 2019, S. 15–25
- Erik R. Seiffert und Elwyn L. Simons: Widanelfarasia, a diminutive placental from the late Eocene of Egypt. PNAS 97 (6), 2000, S. 2646–2651
- Erik R. Seiffert, Elwyn L. Simons, Timothy M. Ryan, Thomas M. Bown und Yousry Attia: New remains of Eocene and Oligocene Afrosoricida (Afrotheria) from Egypt, with implications for the origin(s) of afrosoricid zalambdodonty. Journal of Vertebrate Paleontology 27 (4), 2007, S. 963–972
- Erik R. Seiffert: The oldest and youngest records of afrosoricid placentals from the Fayum Depression of northern Egypt. Acta Palaeontologica Polonica 55 (4), 2010, S. 599–616
- Percy M. Butler und A. Tindell Hopwood: Insectivora and Chiroptera from the Miocene rocks of Kenya Colony. Fossil Mammals of Africa 13, 1957, S. 1–35 ()
- Percy M. Butler: Insectivora and Chiroptera. In: V. J. Maglio und H. B. S. Cooke (Hrsg.): Evolulion of African Mammals. Harvard University Press, 1978, S. 56–68
- Pierre Mein und Martin Pickford: Insectivora from Arrisdrift, a basal Middle Miocene locality in southern Namibia. Memoir of the Geological Survey of Namibia 19, 2003, S. 143–146
- Pierre Mein und Martin Pickford: Early Miocene Insectivores from the Northern Sperrgebiet, Namibia. Memoir of the Geological Survey of Namibia 20, 2008, S. 169–183
- Karen E. Samonds, Sara N. Parent, Kathleen M. Muldoon, Brooke E. Crowley und Laurie R. Godfrey: Rock matrix surrounding subfossil lemur skull yields diverse collection of mammalian subfossils: Implications for reconstructing Madagascar’s paleoenvironments. Malagasy Nature 4, 2010, S. 1–16
- Kathleen M. Muldoon, Donald D. de Blieux, Elwyn L. Simons und Prithijit S. Chatrath: The Subfossil Occurrence and Paleoecological Significance of Small Mammals at Ankilitelo Cave, Southwestern Madagascar. Journal of Mammalogy 90 (5), 2009, S. 1111–1131
- D. A. Burney, N. Vasey, L. R. Godfrey, Ramilisonina, W. L. Jungers, M. Ramarolahy und L. Raharivony: New Findings at Andrahomana Cave, Southeastern Madagascar. Journal of Cave and Karst Studies 70 (1), 2008, S. 13–24
- Steven M. Goodman, Natalie Vasey und David A. Burney: Description of a new species of subfossil shrew tenrec (Afrosoricida: Tenrecidae:Microgale) from cave deposits in southeastern Madagascar. Proceedings of the Biological Society of Washington 120 (4), 2007, S. 367–376
- Steven M. Goodman und William L. Jungers: Extinct Madagaskar. Picturing the island's past. University of Chicago Press, 2014, S. 1–206.
- IUCN: The IUCN Red List of Threatened Species. Version 2015. (), abgerufen am 24. Juni 2016