Megatheriidae
Die Megatheriidae sind eine heute ausgestorbene Familie der Faultiere mit mittelgroßen bis sehr großen Vertretern. Sie traten erstmals im Unteren Miozän vor etwa 18 Millionen Jahren im südlichen Teil Südamerikas, in Patagonien auf. Diese frühen und vergleichsweise kleinen Formen werden meist der Unterfamilie Planopinae zugewiesen, die morphologisch etwas abweichen von den späteren Angehörigen. Die im Pleistozän verbreiteten und heute bekanntesten Gattungen waren Eremotherium und Megatherium, sie gehören zu der jüngeren Linie der Megatheriinae, die erstmals im Mittleren Miozän nachweisbar ist. Vertreter beider Gattungen erreichten teils riesige Ausmaße von bis zu 6 m Länge und 4 bis 6 t Körpergewicht. Sie stellen damit die größten Faultiere dar, die je gelebt haben und die größten endemischen Landsäugetiere Südamerikas. Megatherium war dabei vor allem in der Pamparegion und in den Anden verbreitet, Eremotherium bewohnte tropische Tiefländer und gelangte als einzige Form der Megatherien auch nach Nordamerika. Die letzten Vertreter der Megatheriidae starben im Übergang vom Oberen Pleistozän zum Unteren Holozäns aus.
| Megatheriidae | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelettrekonstruktion von Megatherium im Natural History Museum von London | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Unteres Miozän bis frühes Holozän | ||||||||||||
| 18 Mio. Jahre bis 11.000 Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Megatheriidae | ||||||||||||
| Gray, 1821 | ||||||||||||
Vor allem die späten Megatherien gehören zu den am besten untersuchten Faultieren überhaupt, über die frühen Gattungen liegen aber aufgrund des spärlicheren Fossilmaterials weniger Informationen vor. Die Megatherien waren aufgrund ihres enormen Körpergewichts reine Bodenbewohner, die sich überwiegend vierfüßig fortbewegten, spätere Formen konnten sich auch auf den Hinterbeinen aufrichten, um im Geäst von Bäumen nach Nahrung zu suchen. Diese bestand zumeist aus weicher Pflanzennahrung, die mit den charakteristischen Zähnen zerbissen wurde. Die Erforschung der Megatherien reicht bis in das ausgehende 18. Jahrhundert zurück und begann mit einem Skelettfund in Luján in der Pamparegion Argentiniens. Nur knapp 30 Jahre später, 1821, stellte John Edward Gray den Familiennamen Megatheriidae auf.
Merkmale
Allgemein
Die Megatheriidae umfassen die größten Vertreter der Faultiere. Frühe Formen aus dem Unteren Miozän wie Prepotherium, die in die Gruppe Planopinae gehören, waren mit etwa 123 kg noch relativ klein,[1] spätere, vor allem jene der Unterfamilie Megatheriinae jedoch außerordentlich groß. So erreichte der frühe Megatheriine Pyramiodontherium ein Körpergewicht von 2,5 bis 3,6 t,[2][3] die bekannten Vertreter des Pleistozäns wie Eremotherium und Megatherium brachten etwa 4 bis 6 t auf die Waage.[4][5] Beide waren aber recht variabel in der Größe der einzelnen Arten und umfassen auch kleinere Formen, die nur etwa 1 bis 1,5 t wogen.[6] Die Megatherien zeichneten sich als überwiegend bodenbewohnende Säugetiere durch kräftige und im Vergleich zu den heutigen Baumfaultieren kürzere Gliedmaßen aus. Ebenso verfügten sie wie alle ausgestorbenen Faultiere über längere Schwänze als die heutigen Arten. Der Schwanz war bei den frühen Megatherien lang und robust, bei den späteren vergleichsweise kürzer, aber extrem kräftig.[7][8]
Schädel- und Gebissmerkmale
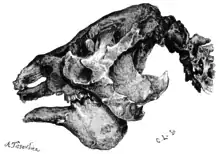
Der Schädel der kleinen Formen wie Prepoplanops erreichte, sofern er vollständig bekannt ist, etwa 25 cm Länge,[9] die größten Schädel wiesen Eremotherium und Megatherium auf, die zwischen 65 und 86 cm lang wurden.[10] In der Aufsicht war der Schädel meist langschmal bis röhrenförmig gestaltet. Markantestes Merkmal stellte der am vorderen Teil des Jochbogens ausgebildete Knochenfortsatz dar, der nach unten reichte und als Ansatzstelle der Kaumuskulatur diente. In der Regel ist der Jochbogen bei den Faultieren nicht geschlossen ausgebildet, nur bei den größten Vertretern der Megatheriidae zeigte dieser sich sekundär wieder verwachsen. Der Unterkiefer war in der Regel massiv und besaß an der Unterkante des Knochenkörpers als besonderes Kennzeichen eine nach unten gerichtete, buckelartige Ausstülpung. Verglichen mit der Größe des Unterkiefers ist diese Ausstülpung tiefer als bei anderen Faultieren, wobei die Tiefe im Laufe der Entwicklung der Megatherien zunahm. Sie diente zur Aufnahme der Alveolen der ebenfalls zunehmend hochkroniger werdenden (hypsodonten) Zähne.[11][12]
Das Gebiss wich wie bei allen Nebengelenktieren markant von dem der meisten Höheren Säugetiere ab. Die entwickelten Megatherien zeichneten sich im Gegensatz zu den heute lebenden und den überwiegenden, ausgestorbenen Faultieren durch vollständig einförmige (homodonte) Zähne aus, die ein molarenartiges Aussehen besaßen und in einer geschlossenen Reihe standen. Nur bei den frühesten Vertretern waren die jeweils vordersten Zähne eckzahnförmig (caniniform) umgestaltet, ähnlich den verwandten Megalonychidae. Sie standen bei diesen auch mit einem mehr oder weniger kurzen Abstand zu den hinteren Backenzähnen.[13] Die molarenartigen Zähne wiesen einen bilophodonten Aufbau mit zwei spitzen Querleisten auf der Kauoberfläche auf und hatten einen rechteckigen bis quadratischen Umriss.[12] Den Zähnen fehlte analog zu den anderen Nebengelenktieren der Zahnschmelz, die härteste Substanz des Zahns. Sie bestanden weitgehend aus Zahnbein, wobei dessen härteste Variante, das Orthodentin, nur 16 bis 19 % eines Zahnes einnahm, was in etwa nur der Hälfte dessen bei den Mylodontidae oder Nothrotheriidae entspricht. Erklärt wird diese Ausdünnung des Orthodentins gegenüber anderen Faultierlinien aus dem besonderen Zahnbau der Megatheriidae. Dadurch unterlagen die meist sehr hohen und im Querschnitt spitzen Querleisten auf den Kauoberflächen einem ständigen Selbstschärfungsprozess beim Zerkauen der Nahrung und stumpften so nicht ab. Dementsprechend bestanden die tiefen Kerben zwischen den Leisten aus dem weicheren Vasodentin.[14][15]
Körperskelett

.jpg.webp)
Auch im Körperskelett gab es einzelne Modifikationen. Neben den für alle Nebengelenktiere typischen xenarthrischen Gelenken (Nebengelenke oder Xenarthrale), die an den Querfortsätzen der hinteren Brustwirbel und der Lendenwirbel auftraten und zur festeren Verankerung der Wirbelsäule beitragen sowie seitliche Ausscherungen verhindern,[16] sind vor allem im Bewegungsapparat Besonderheiten zu finden. Der Oberarmknochen bildete einen röhrenförmigen Knochen mit einem ausladenden unteren Ende.[17] So fehlte bei den Megatherien am massiven und vorn und hinten abgeplatteten Oberschenkelknochen der Dritte Trochanter – eine markante Knochenerhebung am Schaft als Muskelansatzstelle –, der ansonsten ein typisches Merkmal der Nebengelenktiere ist. Ausnahmen stellen hier die frühesten Formen dar, bei denen dieser noch als leichte Erhebung vorkam.[9] Schien- und Wadenbein tendierten dazu, fest zu verwachsen, im Gegensatz zu anderen Faultiergruppen mit einer deutlichen Trennung beider Knochen. Auffallende Abweichungen finden sich weiterhin im Fußskelett. Der Hinterfuß war stärker rotiert als bei anderen Bodenfaultieren, so dass das Körpergewicht auf dem äußersten, fünften Strahl und dem Fersenbein ruhte (pedolateral). Besonders deutlich ist dies bei Eremotherium und Megatherium der Fall, doch ist die Drehung des Fußes mehr oder weniger bei allen entwickelten Megatherien vorhanden. Die Rotation zum pedolateralen Fuß ist mit Ausnahme der Megalonychidae bei allen Bodenfaultieren bekannt und entwickelte sich mit unterschiedlichen Varianten innerhalb der einzelnen Linien unabhängig (Konvergente Evolution), wobei diese Drehung hauptsächlich im Bereich der Fußwurzel stattfand und etwa das Sprungbein und teilweise die Mittelfußknochen stark überformte. Durch die Drehung wurde der Fuß zusätzlich gekrümmt, wobei die Krümmung nicht so umfassend und hoch ausfiel wie bei den verwandten Nothrotheriidae. Dadurch setzte das Fersenbein bei den Megatherien mit der vollen Länge auf, was von den Nothrotherien abweicht, bei denen nur das hinterste Ende des Calcaneums den Boden berührte.[18] Insgesamt besaß der Hinterfuß eine hoch modifizierte Gestaltung und wies bei allen Megatherien mit bekanntem Fußskelett einen ähnlichen Aufbau auf. So bestand er nur aus den drei äußeren Strahlen (III bis V), wobei der letzte Strahl zudem eine deutlich abgeplattete Form besaß. Von den drei Strahlen wies wiederum bei den späten Megatherien nur der Mittelstrahl (Strahl III) drei Phalangen auf (von denen die ersten beiden miteinander verwachsen waren). Das Vorkommen der Endphalanx am dritten Fußstrahl zeigt an, dass hier eine Klaue ausgebildet war. Die beiden äußeren (Strahlen IV und V) verfügten nur über zwei Zehenglieder, so dass hier demzufolge keine Krallen bestanden. Eine Ausnahme bildet Pyramiodontherium, bei dem am vierten Zeh noch eine kleine, stark reduzierte dritte Phalanx vorkam. Möglicherweise ist dies ein Hinweis darauf, dass frühe Vertreter der Megatherien ebenfalls über dieses Merkmal verfügten.[19][20][9] Der Vorderfuß ist nur bei Eremotherium und Megatherium wirklich gut überliefert. Er besaß bei Eremotherium bei frühen Formen fünf (I bis V) beziehungsweise bei späteren drei Strahlen (III bis V), wobei Klauen nur an den Strahlen I bis IV oder entsprechend III und IV vorhanden waren. Die Hand von Megatherium wies eine zwischen diesen beiden Varianten intermediäre Form auf mit vier Strahlen (II bis V) und Krallen an den Strahlen II bis IV.[21][22] Eine weitere Besonderheit findet sich in dem Verwachsen des innersten Mittelhandknochens (teilweise auch der beiden innersten) und einigen Elementen der Handwurzel, etwa dem Großen Vieleckbein, zu einer Einheit, die als Metacarpal-Carpal-Komplex (MCC) bezeichnet wird.[23]
Verbreitung
Die Megatherien entstanden im Unteren Miozän im südlichen Teil von Südamerika und waren weitgehend auf diesen Kontinent beschränkt. Nur im Pliozän und Pleistozän erreichte Eremotherium als einziger Vertreter in Verbindung mit dem Großen Amerikanischen Faunenaustausch auch Nordamerika.[24] Die sehr frühen Formen aus der Gruppe der Planopinae sind weitgehend aus Patagonien bekannt, einige wenige Zähne aus La Venta im heutigen Kolumbien lassen aber auch eine Präsenz im Norden vermuten. Die jüngeren Vertreter, die Megatheriinae, kamen dann über weite Teile Südamerikas vor, einen Schwerpunkt der Verbreitung bildeten jedoch die Regionen der Pampa und das nördlich anschließende Mesopotamia. Die Ausbreitung der Megatherien im Mittleren Miozän von Süd nach Nord könnte über einen im westlichen Amazonasgebiet bestehenden Korridor erfolgt sein, wie es auch für einige Südamerikanische Huftiere angenommen wird.[25][26] Möglich ist aber auch eine noch weiter westlich gelegene Route über das Gebiet der heutigen Anden, deren rapide Auffaltung erst im Oberen Miozän in einem Zeitraum zwischen 10,3 und 6,8 Millionen Jahren stattfand.[27] Ihre größte Verbreitung fanden die Megatherien dann im Pleistozän. Dabei bewohnte Eremotherium die tropisch geprägten Tieflandgebiete in Süd- und Nordamerika. Sein Verwandter Megatherium hingegen kam in den temperierten Regionen der Pampa und in höheren Gebirgslagen der Anden vor, wo er bis in eine Höhe von 4500 m nachgewiesen ist.[28] Bemerkenswert ist weiterhin das Auftreten der Megatherien an der pazifischen Küste, die im nördlichen Chile und in Peru durch teilweise extrem trockenes Wüstenklima charakterisiert ist, was im Falle der Atacama seit wenigstens 14 Millionen Jahren anhält.[27] Hier wird davon ausgegangen, dass im Pleistozän in den Küstengebieten Oasen in Form von Lagunen, Sumpfgebieten und Grasländern entlang der Küstenzone bestanden, welche Lebensraum für Säugetiere boten. Dies konnte unter anderem für die Region um Sacaco und Ocucaje für das späte Mittelpleistozän bestätigt werden. Als Ausbreitungsrouten sind hier die Täler der Anden wahrscheinlich, die nicht nur von den Megatherien, sondern auch von den Rüsseltieren Südamerikas genutzt wurden.[6]
Paläobiologie

Die Megatherien gehören zu den am besten untersuchten, fossilen Faultieren überhaupt. In der Regel beschränkt sich das Wissen aber auf die beiden terminalen Formen Megatherium und Eremotherium, die älteren Gattungen sind weit weniger umfassend untersucht. Aufgrund des teils gewaltigen Gewichts sind alle Vertreter der Megatherien als terrestrische, also bodenbewohnende Tiere anzusehen, im Gegensatz zu den heutigen baumlebenden Faultieren. Dies gilt auch für die frühesten Formen des Unteren Miozän. Diese waren mit ihren 100 bis 200 kg Körpergewicht[1] wohl zu schwer, um Bäume dauerhaft zu erklimmen. Zudem besaßen frühe Megatherien wie Prepotherium zwar ähnlich anderen frühen Faultieren eine massive und kurze Elle, die aber etwa im Bau und Ausrichtung des Olecranons, dem oberen Gelenkfortsatz, abweicht. Daraus resultierend konnten sie nicht so große Kraft und Hebelwirkung im Unterarm über den Ellenbogen entwickeln, wie es etwa beim Klettern oder auch Graben erforderlich ist. Andere frühe Faultiere dieser Zeit lebten jedoch durchaus in den Bäumen, bewegten sich in diesen aufrecht kletternd ähnlich den heutigen Ameisenbären fort und nicht nach unten hängend wie die heutigen Baumfaultiere, was aus der Struktur der Gliedmaßen geschlossen werden kann.[29][30] Bei den moderneren Megatherien war das Olecranon sehr kurz, weiterhin wies der Oberarmknochen im Vergleich zu anderen Bodenfaultieren einen langen und schlanken Bau auf mit einem massiven unteren Gelenk, wie es schon bei Pyramiodontherium aus dem Oberen Miozän nachgewiesen ist. Beides spricht gegen eine Funktion der Vorderbeine als Grabwerkzeuge. Vielmehr wurden diese neben innerartlichen Rivalenkämpfen zum Austeilen von Schlägen wohl auch zur Nahrungsaufnahme eingesetzt, indem sie sich auf die Hinterbeine aufrichteten und mit den Armen Äste und Zweige heranzogen.[2] Der Bau der Hinterbeine zeigt für alle Megatherien eine recht einheitliche Gestaltung. Hierbei ist vor allem auffallend, das sich der Oberschenkelknochen und das Schienbein in ihrer Größe weniger stark unterscheiden als etwa bei den Megalonychidae oder den Mylodontidae mit ihrem langen Femur und der kürzeren Tibia. Längere untere Gliedmaßen werden meist mit einer agileren Fortbewegung in Verbindung gebracht, wie es etwa bei den flinken Pferden und Hirschen im Gegensatz zu den schwerfälligen Nashörnern der Fall ist. Dies wird auch für die Megatherien im Bezug auf die Mylodonten angenommen.[2] Über die Fortbewegung der Megatherien geben auch Ichnofossilien Auskunft. Eine gut 59 m lange Spur konnte in der Río-Negro-Formation des Oberen Miozän im nördlichen Patagonien aufgedeckt werden. Aufgrund der Größe der Trittsiegel (durchschnittliche Länge 66 cm beziehungsweise Breite 32 cm) lässt sich hier Pyramiodontherium als Verursacher annehmen. Aus Pehuén-Có in der argentinischen Provinz Buenos Aires ist eine weitere, 35 m lange Fährte bekannt. Sie datiert ins ausgehende Pleistozän steht der Größe der Trittsiegel zufolge (durchschnittliche Länge 88 cm beziehungsweise Breite 48 cm) mit Megatherium in Verbindung. Die erstgenannte Spur weist auf einen dauerhaft vierfüßigen Gang hin, für letzterer wurde teilweise auch eine zweifüßige Fortbewegung in Betracht gezogen.[3][31][32] Es ist aber aufgrund des Skelettbaus davon auszugehen, dass weder Megatherium noch andere späte Megatherien dauerhaft biped waren.[33]
Extrem hochkronige (hypsodonte) Backenzähne sind normalerweise ein Indikator für eine stärker auf abrasive Grasnahrung spezialisierte Ernährungsweise. Der fehlende Zahnschmelz der Zähne erlaubt keine detaillierten Untersuchungen zu Abnutzungsspuren. Der Aufbau der querstehenden Leisten mit einem im Querschnitt dreieckigen Aufbau und scharfer oberer Kante lässt eher eine Ernährung von weicher Pflanzenkost annehmen, die lediglich im Maul zerquetscht und nicht zerkaut wurde.[34] Dafür sprechen neben dem Bau des Kauapparates selbst auch Funde von Koprolithen von Megatherium, die derartiges Pflanzenmaterial enthielten.[12][35] Eine eher blattfressende Lebensweise wird auch für die frühen Megatherien angenommen. Dabei ist es wahrscheinlich, dass ihre sehr bewegliche Oberlippe zum Einsatz kam. Dies wird einerseits aus dem zahnlosen vorderen Bereich des Maules geschlossen,[29] andererseits konnte dies für Megatherium auch anhand des Schädels rekonstruiert werden.[36]
Systematik
Äußere und Innere Systematik
Innere Systematik der Faultiere nach Presslee et al. 2019 (basierend auf Proteinanalysen)[37]
Die Unterteilung der Megalocnoidea erfolgt nach Delsuc et al. 2019[38] |
Innere Systematik der Faultiere nach Varela et al. 2019 (basierend auf skelettanatomischen Merkmalen)[39]
|
Die Megatheriidae bilden eine Familie mittelgroßer bis sehr großer Vertreter aus der Unterordnung der Faultiere (Folivora). Die Faultiere wiederum formen zusammen mit den Ameisenbären (Vermilingua) die Ordnung der Zahnarmen (Pilosa), die innerhalb der Überordnung der Nebengelenktiere (Xenarthra) steht, welche zudem die Ordnung der Gepanzerten Nebengelenktiere mit den heutigen Gürteltieren (Dasypoda) einschließt. Die Nebengelenktiere bilden eine der vier großen Hauptlinien der Höheren Säugetiere (Eutheria), ihre hauptsächliche Gemeinsamkeit ist das Auftreten von xenarthrischen Gelenken (Nebengelenke oder Xenarthrale) an den Querfortsätzen der hinteren Brustwirbel und der Lendenwirbel.[16] Innerhalb der Faultiere stellen die Megatheriidae zusammen mit den Nothrotheriidae und den Megalonychidae eine engere Verwandtschaftsgruppe dar, die als Überfamilie Megatherioidea bezeichnet wird. Unter Berufung auf skelettanatomische Merkmale können die Megatherioidea als eine der zwei großen Faultierlinien betrachtet werden. Die andere wird durch die Mylodontoidea mit den Mylodontidae, den Scelidotheriidae und den Orophodontidae als Mitglieder angezeigt (unter Umständen haben die beiden letztgenannten auch nur den Status als Unterfamilie der Mylodontidae inne[39]).[40] Basierend auf molekulargenetischen Studien und Proteinanalysen lassen sich mit dem Megalocnoidea eine dritte Linie hinzufügen, die aus den Faultieren der Westindischen Inseln besteht. Den letztgenannten Untersuchungen zufolge schließen die Megatherioidea mit den Dreifinger-Faultieren (Bradypus) eine der beiden heute noch bestehenden Faultiergattungen ein. Die zweite Gattung, die Zweifinger-Faultiere (Choloepus), sind den Mylodontoidea zuzuweisen.[38][37] Zumeist gelten innerhalb der Megatherioidea die Nothrotherien als die nächsten Verwandten der Megatherien.[40] Es wird aber auch eine nähere Beziehung der Nothrotherien mit den Megalonychidae diskutiert.[41] Die vorliegenden genetischen und proteinbasierten Analysen konnten diese unterschiedlichen Auffassungen bisher nicht eindeutig klären.[38][37]
Innerhalb der Megatherien sind zwei Unterfamilien zu unterscheiden. Die stammesgeschichtlich ältere Gruppe wird durch die Planopinae aus dem Unteren und Mittleren Miozän repräsentiert. Diese besaßen noch einen caniniformen vorderen Zahn, der durch ein kleines Diastema von den hinteren molarenartigen Zähnen getrennt war. Die modernen Megatheriinae, die ab dem Mittleren Miozän nachweisbar sind, hingegen hatten vollständig homodonte Backenzähne in einer geschlossenen Reihe.[9] Ursprünglich wurden in die Megatheriidae auch noch die Unterfamilien der Nothrotheriinae und der Schismotheriinae gestellt.[42] Aufgrund von Schädeluntersuchungen gelten die Nothrotheriidae, in die unter anderem Nothrotherium, Nothrotheriops und das semiaquatisch lebende Thalassocnus gestellt werden, aber als eigenständige Familie, die die Schwestergruppe der Megatheriidae formt. Die Vertreter der Schismotheriinae wie Schismotherium, Hapalops oder Analcimorphus gelten nur noch als basale Formen der Megatherioidea, deren Verwandtschaft zueinander und zu anderen Angehörigen der Überfamilie unklar ist.[40]
Überblick über die Gattungen der Megatheriidae
Innere Systematik der Megatheriidae nach Varela et al. 2019[39]
|
Es werden heute zwei Unterfamilien mit mehr als einem Dutzend Gattungen unterschieden:[42][13][43][9][44]
- Familie Megatheriidae Gray, 1821
- Unterfamilie Planopinae Ameghino, 1887 (auch Planopsinae Winge, 1923)
- Planops Ameghino, 1887
- Prepotherium Ameghino, 1891
- Prepoplanops Carlini, Brandoni & Molin, 2013
- Unterfamilie Megatheriinae Gray, 1821
- Megathericulus Ameghino, 1904
- Anisodontherium Brandoni & De Iuliis , 2007
- Eomegatherium Kraglievich, 1926
- Plesiomegatherium Roth, 1911
- Megatheriops C. Ameghino, 1921
- Sibotherium Rincón, Valerio & Laurito, 2020
- Pyramiodontherium Rovereto, 1914
- Promegatherium Ameghino, 1883
- Urumaquia Carlini, Brandoni & Sánchez, 2006
- Proeremotherium Carlini, Brandoni & Sánchez, 2006
- Pliomegatherium Kraglievich, 1930
- Megatheridium Cabrera, 1928
- Megatherium Cuvier, 1796 (Syn.: Essonodontherium, Hebetotherium, Neoracanthus, Ocnobates, Oracanthus, Paramegatherium, Perezfontanatherium ?)
- Eremotherium Spillmann, 1948 (Syn.: Perezfontanatherium ?, Pseuderemotherium, Schaubia, Schaubitherium)
Gegenüber dieser klassischen Ansicht fasst eine phylogenetische Studie aus dem Jahr 2017 die Thalassocninae, die ursprünglich zu den Nothrotherien gestellt wurden und mit Thalassocnus den einzigen bekannten semi-aquatisch lebenden Vertreter der Faultiere enthalten, als dritte Unterfamilie der Megatherien auf. In dieser bilden sie Schwestergruppe der Megatheriinae.[45] Innerhalb der Planopinae gelten die Gattungen Paraplanops, Prepotheriops und Proprepotherium als problematisch. Sie wurden alle von Florentino Ameghino um die Wende des 19. zum 20. Jahrhunderts eingeführt, ihr Typusmaterial ist meist stark fragmentiert oder sehr schlecht erhalten. Deshalb werden gegenwärtig die Gattungsbezeichnungen als nomina dubia geführt.[9] Ebenfalls problematisch ist Ocnopus aus der Gruppe der Megatheriinae, eingeführt 1875 von Johannes Theodor Reinhardt, dessen Holotyp zwei Zähne darstellen, die aber Megatherium zugehören. Weiteres, zur Gattung gestelltes postcraniales Skelettmaterial dagegen wurde teilweise den Nothrotheriidae[42] oder Megalonychidae zugewiesen. Nach jüngeren Untersuchungen gilt die Gattung als synonym zu Valgipes aus der Familie der Scelidotheriidae, nahen Verwandten der Mylodontidae.[46] Das 1976 beschriebene Perezfontanatherium aus Uruguay wird weiterhin als identisch zu Megatherium oder Eremotherium angesehen.[19] Möglicherweise muss auch Diabolotherium zu den Megatheriinae gestellt werden. Die Gattung wurde zuvor als Mitglied der Megatherioidea ohne genauere Zuweisung aufgefasst.[39]
Stammesgeschichte
Entwicklungstendenzen
Neben der allgemeinen Körpergrößenzunahme und der stärkeren Eindrehung des Fußes hin zur pedolateralen Form lassen sich vor allem Veränderungen im Zahnbau erkennen. Die stammesgeschichtlich älteren Planopinae wiesen noch ein Gebiss auf, das ihren Verwandten, den Megalonychidae, ähnelte. Der jeweils vorderste Zahn war caniniform, besaß also eine eckzahnartige Gestalt, und stand in einem gewissen Abstand vor den hinteren Zähnen, der im Laufe der Entwicklung immer kürzer wurde. Weiterhin waren die hinteren, molarenartigen Zähne noch oval in ihrer Gestalt sowie vorn und hinten verschmälert, zudem wiesen sie neben den beiden querstehenden Leisten zusätzliche Höckerchen auf der Kauoberfläche auf. Die Megatheriinae hatten dagegen eine geschlossene Zahnreihe, der vorderste Zahn war ebenfalls molarenartig geformt, so dass das gesamte Gebiss einen homodonten Aufbau besaß, während Höckerchen nicht mehr auftraten. Frühe Megatheriinen waren noch durch vorn und hinten verschmälerte und dadurch eher rechteckig bis rautenförmig wirkende Zähne charakterisiert, die späteren zeichneten sich durch typisch quadratisch gestaltete aus.[9][27][47]
Miozän
Erstmals sind Megatherien im Unteren Miozän nachgewiesen. Entstanden sind sie vermutlich, ähnlich zahlreichen anderen Gruppen der Nebengelenktiere im südlichen Südamerika, wo sie auch ihr erstes Auftreten haben. Die bedeutendsten Funde aus dieser frühen Zeit stammen aus der Santa-Cruz-Formation im südlichen Teil Patagoniens und datieren zwischen 18 und 16 Millionen Jahren. Hier sind unter anderem die Gattungen Planops und Prepotherium nachgewiesen, die zu den am besten dokumentierten Vertretern der frühen Megatherien gehören. Von ersterem stammt mehrere, mitunter vollständige Schädel und ein Teilskelett,[48] von letzterem ein schädelloses Teilskelett und ein Unterkiefer. Sie gehören der Unterfamilie der Planopinae an und stellten gegenüber den riesigen späteren Vertretern der Familie noch relativ kleine Formen dar mit einem Gewicht von 123 kg für Prepotherium, ein bisher noch nicht genauer bestimmter Angehöriger der Megatheriidae könnte schon bis zu 200 kg gewogen haben.[1] Andere hier häufig vorkommende Faultiere wie Hapalops können dagegen nur in eine nähere Verwandtschaft mit den Megatherien gestellt werden, ohne diesen direkt anzugehören.[40] Außerhalb dieser Gesteinseinheit ist weiterhin Prepoplanops aus der Cerro-Boleadoras-Formation südlich des Lago Buenos Aires im südlichen Argentinien überliefert. Dieses wurde anhand eines vollständigen Schädels mit Unterkiefer und einigen Beinknochen beschrieben und steht zeitlich den Funden der südlicheren Santa-Cruz-Formation nahe. Die Planopinae sind mit Planops noch bis in das Mittlere Miozän nachweisbar, wo Funde aus dem westlichen Teil der argentinischen Provinz Neuquén stammen.[9] Zähne einer bisher nicht genau bestimmten Gattung mit Affinitäten zu Planops aus der 14 Millionen Jahre alten Fossillagerstätte La Venta am Mittellauf des Río Magdalena in Kolumbien sprechen möglicherweise auch für das Auftreten der Planopinae weit im Norden des südamerikanischen Kontinents.[27]
Im Mittleren Miozän sind dann erstmals auch die Megatheriinae anzutreffen. Diese gehören mit ihren späteren Vertretern zu den am besten untersuchten Faultiergruppen, die frühen Formen sind allerdings meist fragmentiert überliefert. Die früheste bekannte Gattung ist das kleine Megathericulus, von dem zahlreiche Reste aus Patagonien stammen, so unter anderem aus 11 bis 12 Millionen alten Ablagerungen der Río-Mayo-Formation nahe Cerro Guenguel und Arroyo Pedregosa, beide im westlichsten Argentinien am Fuß der Anden gelegen.[49] Allgemein wird für die Megatheriinae ebenfalls ein Ursprung im südlichen Südamerika angenommen, doch zeigt ein zahnloser Unterkiefer von Megathericulus vom Río Sepa, einem Nebenfluss des Río Urubamba im peruanischen Amazonasgebiet, dass die Gruppe schon im Mittleren Miozän weit im Norden verbreitet war. Der Fund gehört zu einer reichhaltigen Faunengemeinschaft, die als Fitzcarrald-Lokalfauna bezeichnet wird und auf ein Alter von 10 bis 17 Millionen Jahre datiert. Die Ablagerungen, in denen die Fossilien eingebettet sind, gehen auf ein ehemals großflächiges Feuchtgebiet aus Sümpfen, Seen und Flüssen zurück, die nach Norden in die Karibik entwässerten, das als Pebas megawetland den Vorläufer des heutigen Amazonasregenwaldes darstellt.[27][26] Da zudem im nördlich angrenzenden Fundgebiet von La Venta postcraniale Skelettteile eines noch nicht näher beschriebenen Megatheriinen entdeckt wurden, ist ein Ursprung der moderneren Vertreter weiter im Norden auch nicht ausgeschlossen.[50][12]
Aus dem Oberen Miozän ist im südlichen Südamerika weiterhin Anisodontherium nachgewiesen, ein mittelgroßer Vertreter, der zuerst anhand eines zerdrückten Schädels mit Unterkiefer aus der Arroyo-Chasicó-Formation in der argentinischen Provinz Buenos Aires beschrieben worden war.[43] Weiterer Funde, die Unterkiefer- und Handknochenreste umfassen, liegen aus der Saladillo-Formation im nordwestlichen Argentinien vor. Dabei gehört Anisodontherium mit seinen schmalen Backenzähnen noch zu den ursprünglicheren Megatheriinen.[47] Ein anderer bedeutender Vertreter dieser Zeit ist das schon zwischen 2,5 und 3,6 t schwere Pyramiodontherium, das unter anderem neben einem schädellosen Teilskelett aus der Toro-Negro-Formation[2][3] oder einem für miozäne Megatherien bisher außergewöhnlich vollständigen Fußskelett aus Andalhuala, beide im nordwestlichen Argentinien gelegen,[20] auch einige Beinelemente aus dem Conglomerado osífero umfasst. Der Conglomerado osífero stellt den unteren Abschnitt der Ituzaingó-Formation dar, die an den Uferbänken des Unterlaufs des Río Paraná nahe der Stadt Paraná im Nordosten Argentiniens aufgeschlossen ist.[51] Die dort vorgefundene reichhaltige Faunengemeinschaft spiegelt auch die Diversität der Megatheriinen jener Zeit wider, da von den wenigstens ein halbes Dutzend bekannten Gattungen des Obermiozäns allein vier hier vorkommen: neben Pyramiodontherium zudem Pliomegatherium, Promegatherium und Eomegatherium.[52][53] Pliomegatherium umfasst dabei mehrere Unterkieferfragmente und Fußknochen.[54] Im nördlichen Südamerika sind ebenfalls seit Beginn des 21. Jahrhunderts mehrere Gattungen der Megatheriinen nachgewiesen, was vermuten lässt, dass die taxonomische Vielfalt hier ebenfalls recht hoch war, jedoch durch die im Gegensatz zum Süden des Kontinentes begrenzte Anzahl an Fundstellen gegenwärtig noch limitiert erscheint.[12] Als besonders divers erwies sich die obermiozäne Urumaco-Formation im Nordwesten von Venezuela, in der wenigstens 20 verschiedene Arten der Nebengelenktiere nachgewiesen wurden,[55] darunter auch zahlreiche Gliedmaßenknochen, die zum großen Urumaquia gestellt werden. Aus der Codore-Formation des Pliozäns, ebenfalls im nordwestlichen Venezuela, wiederum liegt ein vollständiger, über 41 cm langer Schädel vor, der Proeremotherium repräsentiert.[13][56]
Bereits im ausgehenden Oberen Miozän vor rund 5,8 Millionen Jahren tritt mit Sibotherium ein Angehöriger der Megatherien im südlichen Nordamerika auf. Dies geschah noch vor dem eigentlichen Einsetzen des Großen Amerikanischen Faunenaustausches, der im Mittleren Pliozän vor rund 3,5 Millionen Jahren mit der Schließung des Isthmus von Panama und der Entstehung einer Landbrücke zwischen den beiden amerikanischen Kontinenten begann. Nachgewiesen wurde die Gattung mit einigen Zähnen, Unterkieferresten und Beinknochen in der Curré-Formation in Costa Rica.[44]
Plio- und Pleistozän

Im Pliozän sind dann auch erstmals die beiden größten Vertreter der Megatheriidae fassbar: Megatherium und Eremotherium. Die frühesten Funde von Megatherium stammen aus dem Hochland des Altiplano im westlichen Bolivien und sind zwischen 5,4 und 2,8 Millionen Jahre alt. Sie umfassen einige Unterkiefer- und Schädelreste sowie postcraniale Elemente, unter anderem einen 39 cm langen Oberschenkelknochen, und verweisen so auf einen kleinen Angehörigen der Gattung, der nur rund 1 bis 1,7 t wog.[57] Die geographische Entstehung von Megatherium wird diskutiert, die Gattung blieb auf Südamerika beschränkt und bewohnte die temperierten südlichen Regionen der Pampagebiete und die Hochlagen der Anden.[6] Der älteste Nachweis von Eremotherium stammt mit mehreren Teilskeletten aus dem Oberpliozän von Florida. Die aufgefundenen Individuen sind etwas graziler als spätere Vertreter von Eremotherium, erreichen aber schon beinahe deren Größe. Der Ursprung von Eremotherium wird teilweise in den tropischen Flachlandgebieten von Südamerika gesucht, wo die Gattung auch ihre Hauptverbreitung hat,[21] eine Entstehung weiter im Norden ist aber auch nicht ausgeschlossen.[56] Der Nachweis von Eremotherium in Nordamerika zeigt zudem weiterhin, dass es auch den Megatherien neben zahlreichen anderen originär südamerikanischen Tiergruppen gelang, im Zuge des Großen Amerikanischen Faunenaustausches Südamerika zu verlassen.[24] Im Oberen Pleistozän sind dann sowohl von Eremotherium als auch von Megatherium die größten Formen überliefert, die jeweils zwischen 4 und 6 t wogen und etwa 6 m lang wurden. Eremotherium verschwand bereits um 11.300 vor heute mit dem Ausklingen der letzten Kaltzeit,[58] Megatherium hielt sich noch bis in das Untere Holozän vor etwa 8000 Jahren, eventuell auch nur bis vor rund 11.000 Jahren, wie Funde aus Campo Laborde in Argentinien zeigen.[59][60][61]
Forschungsgeschichte


Der nachweislich erste Fund eines Vertreters der Megatherien wurde Ende der 1780er Jahre nahe der Ortschaft Luján in der Pamparegion des heutigen Argentinien entdeckt. Es handelte sich dabei um ein nahezu vollständiges Skelett eines riesigen Tieres, das nach Madrid in Spanien gebracht wurde und dort als erste Skelettrekonstruktion eines ausgestorbenen Landwirbeltiers im Jahr 1793 aufgebaut wurde. Der französische Anatom Georges Cuvier untersuchte das Skelett und führte auch die noch heute gültige wissenschaftliche Bezeichnung Megatherium americanum ein, seine Erstbeschreibung erschien im Jahr 1796. Weiteres aufschlussreiches Fundmaterial kam erst rund 40 Jahre nach der Entdeckung des Skelettes zum Vorschein. Bedeutung hat hierbei die Reise von Charles Darwin mit der HMS Beagle, auf der er 1832 in Punta Alta nahe Bahía Blanca, ebenfalls in der Pamparegion, zahlreiche Knochen von Megatherium fand. Das gesamte, während der Reise geborgene Fossilmaterial wurde nach London verschifft, wo es Richard Owen ab 1836 detaillierten Analysen unterzog. Nach einem ersten Bericht 1840 veröffentlichte Owen zwischen 1851 und 1860 mehrere Publikationen zu Megatherium, die maßgeblich das Bild der großen Bodenfaultiere prägten.[5]
Bedeutende weitere Funde kamen dann wieder zum Ende des 19. Jahrhunderts ans Licht. Zwischen 1887 und 1893 besuchte Carlos Ameghino mehrmals die frühmiozänen Ablagerungen der Santa-Cruz-Formation im Süden von Patagonien. Dabei sammelte er unzähliges Fundmaterial, das überwiegend von seinem Bruder Florentino Ameghino vorgestellt wurde.[62] Aus diesen Sammlungen gingen unter anderem die Erstbeschreibungen von Planops im Jahr 1887 und von Prepotherium vier Jahre später hervor, womit erstmals auch Vertreter der stammesgeschichtlich älteren Linie der Planopinae entdeckt worden waren.[9]
Bereits 1821 hatte John Edward Gray einen Namen für die Familie eingeführt, den er allerdings mit Megatheriadae bezeichnete. Dieser basierte auf Cuviers Megatherium, dem damals einzigen bekannten Vertreter. Der Begriff Megatherium selbst setzt sich aus den griechischen Wörtern μέγας (mégas „groß“) und θηρίον (thērion „Tier“) zusammen und wurde von Cuvier aufgrund der immensen Größe des Ursprungsfundes gewählt. Gray beschrieb die Merkmale der Familie kurz mit Face short: nose with a short trunk: teeth, grinders, cylindrical: claws compressed: body hairy („Gesicht kurz: Nase mit kurzem Rüssel: Zähne, Mahlzähne, zylindrisch: Krallen verschmälert: Körper behaart“).[63] Der Name wurde erst 1843 von Richard Owen in die richtige Version Megatheriidae korrigiert.[64] Trotz falscher Schreibweise gilt Gray vielen Wissenschaftlern als Erstautor des Familiennamens.[42]
Einzelnachweise
- Néstor Toledo, Guillermo Hernán Cassini, Sergio F. Vizcaíno und M. Susana Bargo: Mass estimation of Santacrucian sloths from the Early Miocene Santa Cruz Formation of Patagonia, Argentina. Acta Palaeontologica Polonica 59 (2), 2014, S. 267–280
- Gerardo De Iuliis, Guillermo H. Ré und Sergio F. Vizcaíno: The Toro Negro megatheriine (Mammalia, Xenarthra): A new species of Pyramiodontherium and a review of Plesiomegatherium. Journal of Vertebrate Paleontology 24 (1), 2004, S. 214–227
- Ricardo N. Melchor, Mariano Perez, M. Cristina Cardonatto und Aldo M. UmazanoLate: Miocene ground sloth footprints and their paleoenvironment: Megatherichnum oportoirevisited. Palaeogeography, Palaeoclimatology, Palaeoecology 439, 2015, S. 126–143, doi:10.1016/j.palaeo.2015.02.010
- Carlos Castor Cartelle: Preguiças terrícolas, essas desconhecidas. Anales Ciência Hoje 27, 2000, S. 19–25
- Sergio F. Vizcaíno, M. Susanna Bargo und Richard A. Fariña: Form, function, and paleobiology in xenarthrans. In: Sergio F. Vizcaíno und W. J. Loughry (Hrsg.): The Biology of the Xenarthra. University Press of Florida, 2008, S. 86–99
- François Pujos: Paleogeographic distribution and anatomical adaptions in Peruvian megatheriine ground sloths (Xenarthra, Megatherioidea). In: Sergio F. Vizcaíno und W. J. Loughry (Hrsg.): The Biology of the Xenarthra. University Press of Florida, 2008, S. 56–63
- M. Susana Bargo, Sergio F. Vizcaíno, Fernando M. Archuby und R. Ernesto Blanco: Limb bone proportions, strength and digging in some Lujanian (Late Pleistocene-Early Holocene) mylodontid ground sloths (Mammalia, Xenarthra). Journal of Vertebrate Paleontology 20 (3), 2000, S. 601–610
- Giuseppe Tito: New remains of Eremotherium laurillardi (Lund, 1842) (Megatheriidae, Xenarthra) from the coastal region of Ecuador. Journal of South American Earth Sciences 26, 2008, S. 424–434
- Alfredo A. Carlini, Diego Brandoni und Carlos N. Dal Molin: A new genus and species of Planopinae (Xenarthra: Tardigrada) from the Miocene of Santa Cruz Province, Argentinia. Zootaxa 3694 (6), 2013, S. 565–578
- Virginia L. Naples: Cranial osteology and function in the tree sloths. Bradypus and Choloepus. American Museum Novitates 2739, 1982, S. 1–21
- H. Gregory McDonald: Xenarthran skeletal anatomy: primitive or derived? Senckenbergiana biologica 83, 2003, S. 5–17
- H. Gregory McDonald und Gerardo de Iuliis: Fossil history of sloths. In: Sergio F. Vizcaíno und W. J. Loughry (Hrsg.): The Biology of the Xenarthra. University Press of Florida, 2008, S. 39–55
- Alfredo A. Carlini, Diego Brandoni und Rodolfo Sánchez: First Megatheriines (Xenarthra, Phyllophaga, Megatheriidae) from the Urumaco (Late Miocene) and Codore (Pliocene) Formations, Estado Falcón, Venezuela. Journal of Systematic Palaeontology 4 (3), 2006, S. 269–278
- Sergio F. Vizcaíno: The teeth of the “toothless”: novelties and key innovations in the evolution of xenarthrans (Mammalia, Xenarthra). Paleobiology 35 (3), 2009; S. 343–366
- Daniela C. Kalthoff: Microstructure of Dental Hard Tissues in Fossil and Recent Xenarthrans (Mammalia: Folivora and Cingulata). Journal of Morphology 272, 2011, S. 641–661
- Kenneth D. Rose: The beginning of the age of mammals. Johns Hopkins University Press, Baltimore, 2006, S. 1–431 (S. 200–204)
- Gerardo De Iuliis: Toward the morphofunctional understanding of the humerus of Megatheriinae: The identity and homology of some diaphyseal humeral features (Mammalia, Xenarthra, Megatheriidae). Senckenbergiana biologica 83, 2003, S. 69–78
- H. Gregory McDonald: Evolution of the Pedolateral Foot in Ground Sloths: Patterns of Change in the Astragalus. Journal of Mammalian Evolution 19, 2012, S. 209–215
- François Pujos und Rodolfo Salas: A new species of the genus Megatherium (Mammalia: Xenarthra: Megatheriidae) from the Pleistocene of Sacaco and Tres Ventanas. Palaeontology 47 (3), 2004, S. 579–604
- Diego Brandoni, Alfredo A. Carlini, Francois Pujos und Gustavo J. Scillato-Yané: The pes of Pyramiodontherium bergi (Moreno & Mercerat, 1891) (Mammalia, Xenarthra, Phyllophaga): The most complete pes of a Tertiary Megatheriinae. Geodiversitas 26 (4), 2004, S. 643–659
- Gerardo De Iuliis und Cástor Cartelle: A new giant megatheriine ground sloth (Mammalia: Xenarthra: Megatheriidae) from the late Blancan to early Irvingtonian of Florida. Zoological Journa1o the Linnean Society 127, 1999, S. 495–515
- Giuseppe Tito und Gerardo De Iuliis: Morphofunctional aspects and paleobiology of the manus in the giant ground sloth Eremotherium Spillmann 1948 (Mammalia, Xenarthra, Megatheriidae). Senckenbergiana biologica 83 (1), 2003, S. 79–94
- Gerardo De Iuliis und Cástor Cartelle: The medial carpal and metacarpal elements of Eremotherium and Megather-ium (Xenarthra: Mammalia). Journal of Vertebrate Paleontology 14, 1994, S. 525–533
- H. Gregory McDonald: Paleoecology of extinct Xenarthrans and the Great American Biotic Interchange. Bulletin of the Florida Museum of Natural History 45 (4), 2005, S. 313–333
- Darin A. Croft: The Middle Miocene (Laventan) Quebrada Honda Fauna, Southern Bolivia and a description of its notoungulates. Palaeontology 50 (1), 2007, S. 277–303
- Julia V. Tejada-Lara, Rodolfo Salas-Gismondi, François Pujos, Patrice Baby, Mouloud Benammi, Stéphane Brusset, Dario De Franceschi, Nicolas Espurt, Mario Urbina und Pierre-Olivier Antoine: Life in proto-Amazonia: Middle Miocene mammals from the Fitzcarrald Arch (Peruvian Amazonia). Palaeontology 58 (2), 2015, S. 341–378
- François Pujos, Rodolfo Salas-Gismondi, Guillaume Baby, Patrice Baby, Cyrille Goillot, Julia Tejada und Pierre-OlivereAntoine: Implications of the presence of Megathericulus (Xenarthra: Tardigrada: Megatheriidae) in the Laventan of Peruvian Amazonia. Journal of Systematik Palaeontology 11 (7–8), 2013, S. 973–991
- François Pujos und Rodolfo Salas: A systematic reassessment and paleogeographic review of fossil Xenarthra from Peru. Bulletin de l'Institut Français d'Etudes Andines, 33, 2004, S. 331–378
- M. Susana Bargo, Néstor Toledo und Sergio F. Vizcaíno: Paleobiology of Santacrucian sloths and anteaters (Cenarthra, Pilosa). In: Sergio F. Vizcaíno, Richard F. Kay und M. Susana Bargo (Hrsg.): Early Miocene paleobiology in Patagonia: High-latitude paleocommunities of the Santa Cruz Formation. Cambridge University Press, New York, 2012, S. 216–242
- Néstor Toledo, M. Susana Bargo und Sergio F. Vizcaíno: Muscular Reconstruction and Functional Morphology of the Forelimb of Early Miocene Sloths (Xenarthra, Folivora) of Patagonia. The Anatomical Record 296, 2013, S. 305–325
- Adrià Casinos: Bipedalism and quadrupedalism in Megatherium: an attempt at biomechanical reconstruction. Lethaia 29, 1996, S. 87–96
- R. Ernesto Blanco und Ada Czerwonogora: The gait of Megatherium Cuvier 1796 (Mammalia, Xenarthra, Megatheriidae). Senckenbergiana biologica 83 (1), 2003, S. 61–68
- Richard A. Fariña, Sergio F. Vizcaíno und Gerardo De Iuliis: Megafauna. Giant beasts of Pleistocene South America. Indiana University Press, 2013, S. 1–436 (S. 254–256) ISBN 978-0-253-00230-3
- M. Susana Bargo, Gerardo De Iuliis und Sergio F. Vizcaíno: Hypsodonty in Pleistocene ground sloths. Acta Palaeontologica Polonica 51 (1), 2006, S. 53–61
- M. Susana Bargo: The ground sloth Megatherium americanum: Skull shape, bite forces, and diet. Acta Palaeontologica Polonica 46 (2), 2001, S. 173–192
- M. Susana M. Bargo, Néstor Toledo und Sergio F. Vizcaíno: Muzzle of South American Pleistocene Ground Sloths (Xenarthra, Tardigrada). Journal of Morphology 267, 2006, S. 248–263
- Samantha Presslee, Graham J. Slater, François Pujos, Analía M. Forasiepi, Roman Fischer, Kelly Molloy, Meaghan Mackie, Jesper V. Olsen, Alejandro Kramarz, Matías Taglioretti, Fernando Scaglia, Maximiliano Lezcano, José Luis Lanata, John Southon, Robert Feranec, Jonathan Bloch, Adam Hajduk, Fabiana M. Martin, Rodolfo Salas Gismondi, Marcelo Reguero, Christian de Muizon, Alex Greenwood, Brian T. Chait, Kirsty Penkman, Matthew Collins und Ross D. E. MacPhee: Palaeoproteomics resolves sloth relationships. Nature Ecology & Evolution 3, 2019, S. 1121–1130, doi:10.1038/s41559-019-0909-z
- Frédéric Delsuc, Melanie Kuch, Gillian C. Gibb, Emil Karpinski, Dirk Hackenberger, Paul Szpak, Jorge G. Martínez, Jim I. Mead, H. Gregory McDonald, Ross D.E. MacPhee, Guillaume Billet, Lionel Hautier und Hendrik N. Poinar: Ancient mitogenomes reveal the evolutionary history and biogeography of sloths. Current Biology 29 (12), 2019, S. 2031–2042, doi:10.1016/j.cub.2019.05.043
- Luciano Varela, P. Sebastián Tambusso, H. Gregory McDonald und Richard A. Fariña: Phylogeny, Macroevolutionary Trends and Historical Biogeography of Sloths: Insights From a Bayesian Morphological Clock Analysis. Systematic Biology 68 (2), 2019, S. 204–218
- Timothy J. Gaudrin: Phylogenetic relationships among sloths (Mammalia, Xenarthra, Tardigrada): the craniodental evidence. Zoological Journal of the Linnean Society 140, 2004, S. 255–305
- Christian de Muizon, H. Gregory McDonald, Rodolfo Salas und Mario Urbina: The youngest species of the sloth Thalassocnus and a reassessment of the relationships of the sloths (Mammalia: Xenarthra). Journal of Vertebrate Paleontology 24 (2), 2004, S. 387–397
- Malcolm C. McKenna und Susan K. Bell: Classification of mammals above the species level. Columbia University Press, New York, 1997, S. 1–631 (S. 96–99)
- Diego Brandoni und Gerardo De Iuliis: A new genus for the Megatheriinae (Xenarthra, Phyllophaga, Megatheriidae) from the Arroyo Chasicó Formation (Upper Miocene) of Buenos Aires Province, Argentina. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 244 (1), 2007, S. 53–64
- Ascanio D. Rincón, Ana L. Valerio und César Laurito: First fossil record of a Megatheriidae-Megatheriinae in the Early Hemphillian (Late Miocene) from San Gerardo de Limoncito, Curré Formation, Costa Rica. Revista Geológica de América Central 62, 2020, doi:10.15517/rgac.v62i0.41278
- Eli Amson, Christian de Muizon und Timothy J. Gaudin: A reappraisal of the phylogeny of the Megatheria (Mammalia: Tardigrada), with an emphasis on the relationships of the Thalassocninae, the marine sloths. Zoological Journal of the Linnean Society 179 (1), 2017, S. 217–236, doi:10.1111/zoj.12450
- Gerardo De Iuliis, François Pujos und Cástor Cartelle: A new ground sloth (Mammalia: Xenarthra) from the Quaternary of Brazil. Comptes Rendus Palevol 8, 2009, S. 705–715
- Diego Brandoni, Jaime E. Powell und Osvaldo E. González: Anisodontherium from the Late Miocene of North-Western Argentina. Acta Palaeontologica Polonica 57 (2), 2012, S. 241–249
- Robert Hoffstetter: Description d´un squelete de Planops (Gravigrade du Miocène de Patagonie). Mammalia 25, 1961, S. 57–96
- Gerardo De Iuliis, Diego Brandoni und Gustavo J. Scillato-Yané: New Remains of Megathericulus patagonicusAmeghino, 1904 (Xenarthra, Megatheriidae): Information on Primitive Features of Megatheriine. Journal of Vertebrate Paleontology 28 (1), 2008, S. 181–196
- H. Gregory McDonald: Xenarthrans: Pilosans. In: Richard F. Kay, Richard H. Madden, Richard L. Cifelli und John J. Flynn (Hrsg.): Vertebrate Paleontology in the Neotropics. The Miocene Fauna of La Venta, Colombia. Smithsonian Institution Press, Washington, 1997, S. 233–245
- Diego Brandoni und Alfredo A. Carlini: On the presence of Pyramiodontherium (Mammalia, Xenarthra, Megatheriidae) in the Late Miocene of Northeastern Argentina and its biogeographical implications. Revista Italiana di Paleontologia e Stratigrafia 115 (1), 2009, S. 111–123
- Diego Brandoni: Los Megatheriinae (Xenarthra, Tardigrada) de la Formación Ituzaingó (Mioceno Superior-Plioceno) de la provincia de Entre Ríos. In: F. G. Aceñolaza (Hrsg.): Temas de la Biodiversidad del Litoral fluvial argentino II, Miscelánea. 14, 2005, S. 27–36
- Diego Brandoni: Los Tardigrada (Mammalia, Xenarthra) del Mioceno Tardío de Entre Ríos, Argentina. In: Diego. Brandoni und J. I. Noriega (Hrsg.): El Neógeno de la Mesopotamia argentina. Asociación Paleontológica Argentina, Publicación Especial 14, 2013, S. 135–144
- Diego Brandoni: A review of Pliomegatherium Kraglievich, 1930 (Xenarthra: Phyllophaga: Megatheriidae). Jahrbuch für Geologie und Paläontologie, Monatshefte 4, 2006, S. 212–224
- Marcelo R. Sánchez-Villagra, Juan D. Carrillo, Alfredo A. Carlini und Carlos Jaramillo: New fossil mammals from the Northern Neotropics (Urumaco, Venezuela, Castilletes, Colombia) and their significance fort he latitudinal gradient in diversity and The Great American Biotic Interchange. In: Erin Maxwell und Jessica Miller-Camp (Hrsg.): 74th Annual Meeting of the Society of Vertebrate Paleontology November 5.8, 2014, Berlin, Abstracts and Papers. Berlin, 2014, S. 220
- Alfredo A. Carlini, Diego Brandoni und Rodolfo Sánchez: Additions to the knowledge of Urumaquia robusta (Xenarthra, Phyllophaga, Megatheriidae) from the Urumaco Formation (Late Miocene), Estado Falcón, Venezuela. Paläontologische Zeitschrift 82 (2), 2008, S. 153–162
- Pierre-Antoine Saint-André und Gerardo De Iuliis: The smallest and most ancient representative of the genus Megatherium Cuvier, 1796 (Xenarthra, Tardigrada, Megatheriidae), from the Pliocene of the Bolivian Altiplano. Geodiversitas 23 (4), 2001. S. 625–645
- Dilce de Fátima Rossetti, Peter Mann de Toledo, Heloísa Maria Moraes-Santos und Antônio Emídio de Araújo Santos, Jr.: Reconstructing habitats in central Amazonia using megafauna, sedimentology, radiocarbon, and isotope analyses. Quaternary Research 61, 2004, S. 289–300
- Gustavo G. Politis, Clara Scabuzzo und Robert H. Tykot: An Approach to Pre-Hispanic Diets in the Pampas during the Early/ Middle Holocene. International Journal of Osteoarchaeology 19, 2009, S. 266–280
- Marína A. Gutiérrez und Gustavo A. Martínez: Trends in the faunal human exploitation during the Late Pleistocene and Early Holocene in the Pampean region (Argentina). Quaternary International 191, 2008, S. 53–68
- Gustavo G. Politis, Pablo G. Messineo, Thomas W. Stafford Jr. und Emily L. Lindsey: Campo Laborde: A Late Pleistocene giant ground sloth kill and butchering site in the Pampas. Science Advances 5, 2019, S. eaau4546, doi:10.1126/sciadv.aau4546
- Larry G. Marshall: Fossil localities for Santacrucian (early Miocene) mammals, Santa Cruz Province, southern Patagonia, Argentina. Journal of Paleontology 50, 1976, S. 1129–1142
- John Edward Gray: On the natural arrangement of vertebrose animals. London Medical Repository 15, 1821, S. 297–310 (306) (PDF)
- George Gaylord Simpson: The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History 85, 1945, S. 1–350 (S. 71)