Fossillagerstätte Geiseltal
Die Fossillagerstätte Geiseltal befindet sich im ehemaligen Braunkohlerevier des Geiseltales südlich der Stadt Halle in Sachsen-Anhalt. Sie ist eine bedeutende Fundstelle heute ausgestorbener Pflanzen und Tiere aus der Zeit des Mittleren Eozäns vor 48 bis 41 Millionen Jahren. Im Geiseltal wurde nachweislich seit 1698 erstmals Kohle gefördert, die ersten Fossilien kamen aber erst Anfang des 20. Jahrhunderts eher zufällig zu Tage. Planmäßige wissenschaftliche Ausgrabungen begannen 1925 seitens der Martin-Luther-Universität Halle-Wittenberg. Unterbrochen durch den Zweiten Weltkrieg, können die Untersuchungen in zwei Forschungsphasen untergliedert werden. Aufgrund der zunehmenden Auskohlung der Rohstofflager kamen die Ausgrabungen Mitte der 1980er Jahre allmählich zum Erliegen und endeten endgültig zu Beginn des dritten Jahrtausends.


Die Braunkohle des Geiseltales wird in vier Hauptflöze untergliedert, fossilführend waren hauptsächlich die drei unteren. Der Schwerpunkt der Fossilverteilung liegt im südlichen und zentralen Geiseltal. Die Funde umfassen Reste von Pflanzen und Tieren. Eine Besonderheit stellen dabei nahezu vollständige Funde dar mit Blättern, Früchten und Stämmen, aber auch Skeletten von Wirbeltieren und Resten von Insekten. Zu den bekanntesten Funden gehört der eines vollständigen Skelettes des Urpferdchens Propalaeotherium aus dem Jahr 1933. Hinzu kommen Wirbeltiere wie Paarhufer, Kleinsäugetiere wie Insektenfresser und Fledermäuse sowie Vögel, Krokodile, Schildkröten, Schlangen, Amphibien und Fische. Bei zahlreichen Wirbeltieren wurden auch Reste des Weichteilgewebes gefunden. Insgesamt sind mehr als 80 Einzelfundstellen mit mehr als 50.000 Fundobjekten bekannt, darunter 36 mit einer nennenswerten Anzahl von Wirbeltieren. Aufgrund der guten und umfangreichen Fossilerhaltung gilt das Geiseltal als Konservat- und Konzentratlagerstätte.
Die Besonderheit der Geiseltalfossilien, vor allem der Wirbeltiere und speziell der Säugetiere, liegt in der einmaligen Erhaltung in Braunkohle, was in Mitteleuropa sonst nicht vorkommt. Für die Entwicklung der Säugetiere stellt das Geiseltal eine wichtige Fundstelle dar, da dort über einen Zeitraum von mehreren Millionen Jahren die Entwicklung einzelner Gruppen beobachtet werden kann. Dadurch gilt die Säugetierfauna als Referenz für das Geiseltalium, einen Abschnitt aus der Stratigraphie der europäischen Landsäugetiere von 47 bis 43 Millionen Jahren. Die umfangreichen Funde aus der Tier- und Pflanzenwelt, aber auch die zahlreichen geologischen Daten ermöglichen eine recht genaue Landschaftsrekonstruktion. Demzufolge bestand zur Zeit der Braunkohlebildung ein mehrstöckiger Niederungswald in Küstennähe, der mit Bächen, Teichen und Mooren durchsetzt war. Der Wald unterlag dem Einfluss von subtropischem Klima und war Heimat einer artenreichen Tierwelt. Der gesamte Fossilbestand des Geiseltales steht unter nationalem Schutz.
Geographische Lage
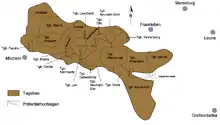
Das Geiseltal, ein Randbereich des Mitteldeutschen Braunkohlereviers, liegt etwa 20 km südlich von Halle (Saale) und rund 10 km südwestlich von Merseburg in Sachsen-Anhalt. Es erstreckt sich auf einer Länge von 15 km von Westnordwest nach Ostsüdost und auf einer Breite von 0,5 bis 5 km. Im Norden grenzt es an die flache Merseburger Buntsandsteinplatte, im Süden an das Müchelner Muschelkalkplateau der Querfurt-Freyburger Mulde. Durchflossen wird das Geiseltal vom namengebenden Fluss Geisel, der in St. Micheln bei Mücheln aus einer der größten Spring- oder Überfallquellen Mitteldeutschlands entspringt und nach 19 km in Merseburg über die Gotthardtsteiche in die Saale entwässert. Sein Einzugsgebiet beträgt etwa 35 km².[1]
Ursprünglich war das Gebiet des Geiseltales relativ eben, was durch die sich westlich der Saale ausbreitenden Buntsandsteine zu erklären ist. Allgemein lagen die Höhen im östlichen Geiseltal bei rund 100 Meter über dem Meeresspiegel, nach Westen hin stiegen sie auf 150 Meter und mehr an. Durch die intensive Tätigkeit der Braunkohletagebaue wurde die Landschaft vor allem in den letzten 150 Jahren aber gravierend verändert und führte neben der Zerstörung mehrerer Ortschaften auch zur Überprägung eines rund 90 km² großen Gebietes mit mehrfacher Verlegung des Flusslaufes der Geisel nach Süden. Infolge der Renaturierung der später aufgelassenen Tagebaue entstanden größere Seen, die gegenwärtig das gesamte Geiseltal bestimmen. Bereits in der zweiten Hälfte des 20. Jahrhunderts wurden im östlichen und südöstlichen Geiseltal der Südfeldsee und der Runstedter See geschaffen, während im westlichen Teil der Geiseltalsee, einer der größten künstlichen Seen Mitteleuropas, erst 2011 fertig geflutet werden konnte. Das westliche und östliche Geiseltal werden heute durch einen bis zu 140 m hohen Kippendamm getrennt, der neben dem heutigen Flusslauf der Geisel auch wichtige Verkehrswege wie Straßen und Schienen trägt.[2]
Geologie
Geologischer Untergrund
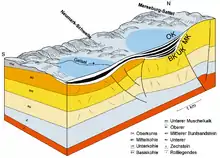
Die Geologie des Geiseltales wurde seit Beginn des 20. Jahrhunderts bei Vorfelduntersuchungen zur Ergründung der Lagerungsverhältnisse der Braunkohle mit Tiefbohrungen gut untersucht. Diese hatten in einzelnen Abschnitten Abstände von nur 100 Metern. Der geologische Untergrund besteht weitgehend aus Ablagerungen des Rotliegenden und des Zechsteins aus der geologischen Periode des Perm vor rund 300 bis 240 Millionen Jahren. Diesen sind die Sedimente des Buntsandsteins (vor 251 bis 243 Millionen Jahren) aufgelagert, wobei überwiegend der Untere und Mittlere Buntsandstein vorkommen. Im südlichen Teil des Geiseltales zum Müchelner Muschelkalkplateau hin sind aber auch Reste des Oberen Buntsandsteins vorhanden, dem in den Rändern des Tales die Ablagerungen des Muschelkalkes aufliegen. Der folgende Hiatus umfasst den Keuper und die Kreide aus einem Zeitraum von etwa 140 Millionen Jahren.[3][4]
Das heutige Geiseltal ist mehrfach in sich gegliedert. Die Neumark-Hauptschwelle, eine Erhebung des Buntsandsteins, teilt es in etwa zwei gleich große Bereiche, das westliche und das östliche Geiseltal. Neben- und untergeordnete Becken sind der Elise-Kessel, der Elisabeth-Kessel, der Wernsdorfer Kessel und andere, prätertiäre Schwellen sind die Kayna-Schwelle und die Ostschwelle.[5] Geologisch stellt das Geiseltal eine Senkungsstruktur dar, deren Entstehung noch nicht vollständig geklärt ist, die aber durch mehrere Prozesse gesteuert wurde. Möglicherweise führte im Paläogen die Auslaugung des Zechsteinsalzes verbunden mit der chemischen Verwitterung des Muschelkalkes (Subrosion) zu Masseverlusten im Untergrund, was eine Absenkung der auflagernden Schichten und die Beckenbildung zur Folge hatte. Vor allem die Salzbewegung (Halokinese) stand dabei im Zusammenhang mit tektonischen Kräften im Untergrund, resultierend aus der Plattentektonik, die auch die Auffaltung der Alpen während der Kreide/Tertiär-Wende vor etwa 65 Millionen Jahren hervorriefen. Dadurch kam es zu einer Erhöhung des Druckes auf das ursprünglich flach gelagerte Zechsteinsalinar, das daraufhin seitlich abfloss. Im nördlichen Bereich des Geiseltals wurden mit seismischen Messungen tektonische Störungen ermittelt, die sogenannte Geiseltal-Nordrand-Störung, die teilweise eine Sprunghöhe von bis zu 200 m in den oberen Schichtlagen (Buntsandstein) erreicht.[3][4][6]
Paläogene Ablagerungen
Die gesamten Ablagerungen des Paläogens erreichen im Geiseltal eine Mächtigkeit von bis zu rund 200 m, beschränken sich aber weitgehend auf das Eozän (vor 56 bis 34 Millionen Jahren). Eingeleitet werden diese tertiären Sedimente durch Tone und Schluffe sowie teils gröberklastisches Material. Die Mächtigkeit der überwiegend fein geschichteten bis blättrigen Braunkohle liegt bei etwa 30 bis 80 m, erreicht stellenweise jedoch auch über 120 m.[7] Generell wird die Geiseltal-Braunkohle in vier Hauptflöze unterteilt, die Unterkohle, Mittelkohle (differenziert in Untere und Obere Mittelkohle) und Oberkohle, weist aber im nördlichen Geiseltal noch lokal eine Basiskohle auf. Bis auf die Basiskohle werden alle Flöze in weitere Flözabschnitte untergliedert. Die einzelnen Flöze erreichen eine Mächtigkeit von 10 bis teilweise 60 m. Unterbrochen werden sie von sandig-schluffigen Sedimentabschnitten (die sogenannten Hauptmittel), die Leithorizonte darstellen und zur Unterscheidung der Flöze dienen. Im südlichen Geiseltal im Tagebau Cecilie bestand allerdings ein teilweise lückenloser Übergang von der Mittelkohle zur Oberkohle. Die Ausdehnung der einzelnen Kohleflöze von Nord nach Süd ist unterschiedlich ausgeprägt und an die halokinetischen und subrosiven Absenkungen des Untergrundes gebunden, sie setzt sich aber generell mit der zunehmenden stratigraphischen Höhenlage der Flöze weiter nach Süden fort. Nur die Oberkohle ist im Geiseltal flächendeckend ausgebildet. Die sehr reiche eozäne Fossilgemeinschaft stammt vor allem aus der Unterkohle und der Mittelkohle.[8][3][9]
Auflagernde Schichten
Das Deckgebirge besteht im Geiseltal aus bis zu 50 m mächtigen Ablagerungen des Mittelpleistozäns und Jungpleistozäns (vor 0,78 bis 0,012 Millionen Jahren). Die ältesten, von der Erosion verschonten Schichten wurden als Grundmoräne durch das Inlandeis der Elster-Kaltzeit (vor 400.000 bis 335.000 Jahren) abgelagert. In der Holstein-Warmzeit (vor 335.000 bis 320.000 Jahren) verlagerte die Unstrut ihren Lauf bis in das Geiseltal und schüttete die sogenannte Körbisdorfer Terrasse auf. Das Inlandeis der Saale-Kaltzeit (vor 320.000 bis vor 128.000 Jahren) hat das Geiseltal nur beim ersten Vorstoß, in Mitteldeutschland als „Zeitz-Phase“[10] und in Norddeutschland als „Drenthe-I-Phase“ oder „Haupt-Drenthe“ bezeichnet, überfahren.[11][12]
Der Rückzug des Inlandgletschers der Saale-Kaltzeit führte insbesondere im Nordostteil des Geiseltales, im ehemaligen Tagebaurandfeld Neumark-Nord, durch Mollisoldiapirismus[13][6][7] zur Bildung von abflusslosen und meist wassererfüllten Senken, in denen überwiegend limnische Sedimente abgelagert wurden. Die einzelnen Seebecken standen zwischen 1986 und 2008 im Fokus interdisziplinärer wissenschaftlicher Untersuchungen, deren Ergebnisse in einer Vielzahl von Publikationen veröffentlicht wurden. Das größte, rund 600 m lange und 400 m breite Becken Neumark-Nord 1 barg Reste einer reichen Großsäugerfauna mit teils vollständigen Skeletten unter anderem vom Europäischen Waldelefanten, Auerochsen, Wisent, Damhirsch und von verschiedenen Nashörnern (Waldnashorn, Steppennashorn), aber auch vom Höhlenlöwen und der Tüpfelhyäne. Die Großsäugerfauna des kleineren Beckens Neumark-Nord 2 ähnelte prinzipiell der des größeren, war aber bis auf wenige Ausnahmen kleinstückig und disartikuliert. In den Uferbereichen beider Becken wurde eine Großzahl an Feuersteinartefakten des mittelpaläolithischen Menschen entdeckt.[11][12][14] Über die Altersstellung besteht noch kein Einvernehmen. Nach verschiedenen Pollenanalysen zeigen die warmzeitlichen Sedimente der beiden Becken eine für die Eem-Warmzeit (vor 128.000 bis 115.000 Jahren) typische Sukzession.[15][16] Für das Becken Neumark-Nord 2 wird eine eemzeitliche Einstufung durch weitere Analysen, etwa Paläomagnetik und radiometrische Altersdatierungen, gestützt.[17][18] Dagegen könnten die warmzeitlichen Sedimente des Beckens Neumark-Nord 1 auch in einer „intrasaalezeitlichen“ Warmzeit abgelagert worden sein. Hinweise dafür liefern die botanischen Reste, so die Makroflora mit den extrem reichhaltigen Relikten eines durch den Tataren-Ahorn geprägten Steppen-Eichen-Mischwaldes, oder einzelnen Leitfossilien der Kleinsäuger, unter anderem der Zwergwaldmaus Apodemus maastrichtensis.[19][20] Ebenso sprechen sowohl die Ostrakodenfauna als auch weitere Befunde nicht für ein eemwarmzeitliches Alter.[21][22]
Das gesamte Schichtpaket wird diskordant von Bachschotter der Geisel und vom mehrere Meter mächtigen Löss der letzten Kaltzeit (Weichsel-Kaltzeit; vor 115.000 bis vor 11.600 Jahren) überlagert, der den Schwarzerdeboden trägt. In den Löss sind verschiedene Eiskeilhorizonte eingebettet, die auf die besonders kalten Phasen der letzten Kaltzeit verweisen. Der späte Abschnitt der Weichsel-Kaltzeit ist am südlichen Rand des Geiseltales umfassender aufgeschlossen (ehemaliger Tagebau Mücheln). Hier ließen sich in einer kleinen Senke vier Warm-Kaltphasen-Zyklen nachweisen, von denen die beiden oberen mit dem Bölling-Interstadial (im klassischen Sinn) und der Älteren Dryaszeit sowie dem Alleröd-Interstadial und der Jüngeren Dryaszeit korrelierbar sind. In der jüngsten wärmeren Folge (Alleröd-Interstadial) dicht unterhalb des rezenten Bodenhorizontes hat sich noch eine dünne Lage des Laacher-See-Tuffs als Relikt einer Vulkaneruption vor rund 13.000 Jahren erhalten.[23][24][11][12]
Fossilfundstellen
Funderhaltung und Fundverteilung
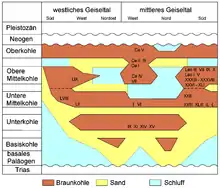


Während Pflanzen und teils auch Wirbellose in allen Bereichen der Kohleflöze gefunden wurden, waren Wirbeltiere an bestimmte, enger begrenzte Fundstellen gebunden. Insgesamt sind mehr als 80 Fundstellen bekannt, wobei 36 einen nennenswerten Bestand an Wirbeltierresten aufweisen. Diese befinden sich überwiegend in der Unter- und der Mittelkohle, mit rund doppelt so vielen Fundstellen in der Mittelkohle wie in der Unterkohle. Die Oberkohle enthielt nur eine relevante Fundstelle und war sonst weitgehend fossilfrei, was teilweise auf die diagenetischen Veränderungen während des Pleistozäns unter Einwirkung periglazialer Bedingungen zurückzuführen ist.[9] In der Basiskohle konnte keine Fossilisation nachgewiesen werden. Die räumliche Verteilung der Fossilfundstellen von Wirbeltieren spiegelt die Voraussetzung für die Funderhaltung wider, die im gesamten Geiseltal nicht im gleichen Maße gegeben war. Dadurch konzentriert sich der größere Teil der Wirbeltierfundstellen, vor allem derjenigen mit teils vollständigen Skeletten oder Weichteilen, im zentralen und südlichen Geiseltal, hauptsächlich nahe der Neumark-Hauptschwelle und im östlich anschließenden Wernsdorfer Kessel. Die überwiegende Anzahl der Fundstellen liegt deswegen in den Tagebauen Cecilie, Leo, Geiselröhlitz und Pfännerhall und verteilt sich auf eine Fläche von rund 20 km².[25] Nach Norden und Westen und teilweise nach Osten nimmt die Güte der Fossilerhaltung stark ab.[26][27]
Wirbeltierfunde aus Braunkohlen sind weltweit sehr selten und wurden außerhalb des Geiseltals in Mitteleuropa bisher nicht beobachtet. Die gute Fossilerhaltung im Geiseltal hat mehrere Gründe. Hauptsächlich verantwortlich sind kalkhaltige Wässer aus den angrenzenden triassischen Gesteinen, hier überwiegend Muschelkalk, die aus südlicher oder südwestlicher Richtung aus dem Bereich der Querfurt-Freyburger Mulde während der Bildung der Braunkohle als zirkulierende Grundwässer eindrangen. Diese neutralisierten weitgehend die zersetzenden Eigenschaften der Huminsäure der Niedermoore im damaligen Geiseltal, waren aber, anhand des Fundbildes erkennbar, nur lokal wirksam. In untergeordnetem Ausmaß sorgte auch Kieselsäure für einen guten Erhalt der Fossilien. Einen weiteren Einfluss auf die Erhaltung der Tierkadaver hatten die meist in der Mittelkohle recht häufig nachgewiesenen Überschwemmungen der ehemaligen Landoberfläche, die zu einer raschen Bedeckung der Kadaver mit Sedimenten führten.[28] Sowohl die Fossilreste als auch die Braunkohle sind im frischen Zustand zu 50 % mit Wasser gesättigt und lösen sich während des Austrocknens sehr schnell auf, indem sie zerblättern und anschließend zu Staub zerfallen. Aus diesem Grunde wurde bereits zu Beginn der 1930er-Jahre für die gezielte Bergung der Fossilien speziell im Geiseltal die Lackfilmmethode entwickelt.[29][3]
Fundstellentypen
Neben Einzel- und Streufunden sowie gelegentlichen Funden aus Bohrkernen können insgesamt drei verschiedene Fundstellentypen der Wirbeltiere unterschieden werden, die zum Teil aber auch in verschiedenen Kombinationen vorkommen:[30][31][3]
- Einsturztrichter
- Hierbei handelt es sich um kreisförmige, teils dolinenartige Vertiefungen mit symmetrischem Aufbau. Entstanden sind sie syngenetisch durch Auslaugung von Gips im unterlagernden Buntsandstein (Subrosion). In der Regel sind die Einsturztrichter an ihren randlichen Schichtstörungen und den daraus resultierenden tektonischen Verschiebungen erkennbar. Es lassen sich zwei Trichtertypen unterscheiden: der eigentliche Einsturztrichter mit Durchmessern von 12 bis 18 m und der Setzungstrichter, der kleiner und flacher ist und Durchmesser von 3 bis 8 m erreicht. Die Eintiefungen waren bei ihrer Entstehung weitgehend mit Wasser gefüllt und bildeten kleine Teiche und Tümpel. Dabei bildeten die oft steilen Randsäume natürliche Fallen für Wirbeltiere, wodurch eine natürliche Grabgemeinschaft entstand. Durch die mittels schneller Sedimentauflage am Teichboden entstandenen Faulschlämme und die dort vorherrschenden anaeroben Bedingungen bildeten sich die Fossilien.
- Leichenfelder
- Diese sind ausgedehntere Bereiche mit Fossilfunden, die sich über 80 bis 100 m erstrecken. Meist befinden sich die Fossilien in großflächigen Vertiefungen und sind in der Regel von einer 20 bis 30 cm dünnen Kohleschicht überdeckt. Sie sind Reste der ehemaligen, durch Überschwemmungen überfluteten Landoberfläche und Moorgebiete und vor allem im zentralen Geiseltal konzentriert.
- Bachläufe
- Den Leichenfeldern ähnelten die Bachläufe, die zum Teil vom westlichen Muschelkalkgebiet kommend das Geiseltal durchzogen und in einem der lokalen Becken mündeten. Es sind rinnenartige Eintiefungen in die Kohle, in denen sich meist kreuzgeschichtete Quarzsande abgelagert hatten. Nur in den tieferen Beckenbereichen wurden auch tonige Sedimente gefunden, was auf eine Verringerung der Fließkraft der Bäche schließen lässt. Die Fossilfunde sind auf die ehemaligen Uferbereiche konzentriert. Die meisten Tiere kamen dort wie bei den Einsturztrichtern an den teils steilen Hängen zu Tode, wurden aber auch Opfer zahlreicher dort lebender Beutegreifer.
Einsturztrichter und Leichenfelder kommen am häufigsten vor, die Bachläufe sind eher selten und wurden erst Mitte der 1950er-Jahre entdeckt. Der Erhaltungszustand der Wirbeltierfossilien in den Leichenfeldern und in den Bachläufen ist ähnlich und lässt gewisse Umlagerungen durch Wasserbewegung verbunden mit Skelettzerfall erkennen. In den Einsturztrichtern kamen vollständige Skelettreste nur in den zentralen und tiefsten Stellen vor, wo die Kadaver vollständig mit Wasser bedeckt waren. Zu den Rändern der Trichter hin sind auch deutliche Disartikulationen der Skelette zu beobachten.[32][30] Die bedeutenden Funde des fossilisierten Weichteilgewebes stammen nur aus den Einsturztrichtern und den Leichenfeldern.[33]
Funde
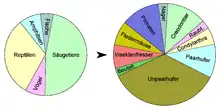
Das Fossilmaterial aus der Braunkohle des Geiseltales ist sehr reichhaltig und umfasst Reste von Pflanzen und Tieren. Das floristische Material liegt in Form von Mikro- und Makroresten vor. Die Fauna ist durch Wirbellose und Wirbeltiere vertreten, von Letzteren gibt es auch eine größere Anzahl von vollständigen Skeletten. Hervorzuheben ist die gute Konservierung von Weichteilgewebe, das fossil nur sehr schlecht erhalten bleibt. Trotz der guten Erhaltungsbedingungen ist ein größerer Teil der Fossilien nicht autochthon abgelagert, sondern wurde während der Fossilisation transportiert, verursacht durch das Fließen von Wasser. Dies trifft überwiegend für die Funde aus den Leichenfeldern und den Bachläufen zu. Die Lagerungsverhältnisse der Fossilien sind allgemein als parautochthon bis teilweise allochthon anzusehen. Die genaue Anzahl der Funde ist unklar, der Bestand des ehemaligen Geiseltalmuseums der Universität Halle umfasst rund 50.000 Fundobjekte, davon zum größten Teil Wirbeltiere. Während der intensiven Grabungsphase der 1960er-Jahre wuchs der Bestand um jährlich mehr als 5830 Objekte. Eine Analyse von mehr als 10.000 Wirbeltierresten ergab mit 5000 Stücken fast zur Hälfte Säugetiere, während Reptilien mit 2000 Funden am zweithäufigsten vertreten sind. Die wissenschaftliche Auswertung des Gesamtbestandes an Fossilien hält nach wie vor an, zahlreiche fossile Lebewesen erhielten anhand des Fundmaterials des Geiseltales ihre Erstbeschreibung. Allein die Wirbeltiere umfassen mehr als 120 Taxa.[3]
Flora


Unter den recht häufigen Pflanzenresten sind neben Pollen und Sporen als Mikroflora, die allein mehr als 100 stratigraphisch relevante Taxa einnehmen,[34] vor allem die Makroreste von Bedeutung. Diese umfassen Blätter, Zweige, Äste, Rinde und Stämme sowie Früchte, Samen und Blütenstände. Allein anhand der Früchte und Samen lassen sich wenigstens 18 Familien nachweisen. Insgesamt sind mehr als zwei Dutzend Familien mit rund 40 Gattungen bekannt. Neben Algen, Moosen, Urfarnen und Farnen – diese teilweise in konzentrierten Anreicherungen von unter anderem Rippen- und Kletterfarnen[35] – kommen überwiegend Reste der höher entwickelten Samenpflanzen vor. Die Nacktsamer sind mit Palmfarnen, teilweise mit ganzen Wedeln, und Koniferen mit Zapfen und Zweigen vertreten. Letztere umfassen unter anderem Kiefern und Mammutbäume, die teilweise gehäuft auftreten. Letztere werden in die Zypressengewächse eingeordnet, von denen des Weiteren auch die ausgestorbene Gattung Doliostrobus überliefert ist. Wesentlich umfangreicher sind die Reste der Bedecktsamer. Hier sind die Zweikeimblättrigen mit rund 20 Familien die größte Gruppe. Dazu gehören Lorbeergewächse und Buchengewächse. Letztere bilden teilweise auch dichte Blattlagen, wie etwa bei der Gattung Dryophyllum. Gagelstrauchgewächse, überwiegend Farnmyrte, sind sehr häufig. In hoher Anzahl belegte Pflanzenreste stammen von Lindengewächsen, Icacinaceen und Myrtengewächsen, von denen Rhodomyrtophyllum dominant ist. Charakteristisch sind auch die langschmalen Blätter von Apocynophyllum aus der Gruppe der Hundsgiftgewächse. Dieser Pflanzenfamilie werden auch mehrere bis zu 44 cm lange Rindenreste mit dem daran anhaftenden, sogenannten „Affenhaar“, fossilisierte Milchsaftröhren, zugewiesen.[36] Einkeimblättrige haben eine geringere Vielfalt. Unter diesen kommen hauptsächlich die Palmengewächse in einer hohen Anzahl vor und sind unter anderem mit Sabal und Phoenicites vertreten. Die häufig überlieferten Palmenstämme können aber taxonomisch nicht genau zugeordnet werden.[37] Auch weitere Vertreter der Einkeimblättrigen wurden beschrieben, beispielsweise aus der Gruppe der Liliengewächse und der Schraubenbaumgewächse.[38]
Wirbellose

Wirbellose wurden erstmals 1913 erwähnt[39] und sind sehr zahlreich. Mollusken, zu denen Muscheln und Schnecken gehören und die teilweise in Massen auftraten, sind mit rund 20 Gattungen vertreten und eignen sich hervorragend für die Rekonstruktion von Kleinbiotopen.[40] Gleiches gilt auch für die Ostracoden, die Muschelkrebse mit 10 Gattungen.[41] Weitere Funde von Gliederfüßern werden den Zehnfußkrebsen zugewiesen. Diese sind aber mit nur 15 Exemplaren äußerst selten und meist nur mit Resten der Panzerung erhalten.[42] Die häufigsten Funde von Wirbellosen sind Insekten. Hier dominieren die Käfer und darunter mit 28 % die Prachtkäfer. Die Gattung Psiloptera ist mit acht Arten nachgewiesen, aber auch Buprestis und Anthaxia sind mit mehreren Arten vertreten. Zu den Bockkäfern, erkennbar an den außerordentlich langen Fühlern, wird Xyleoconites gerechnet, während Eocallidium innerhalb der Gruppe der Schwarzkäfer steht.[43] Daneben kommen einige weitere Käferfamilien vor, darunter die Blatthornkäfer, die Hakenkäfer und die Blattkäfer. Da die Insekten fast ausschließlich mit dem Rücken erhalten sind, ist zwar die teils prächtige Farbgebung der Tiere überliefert, die genaue Artzuweisung kann dadurch aber nicht in allen Fällen mit Sicherheit erfolgen.[44] Auch Fluginsekten wie Eintagsfliegen und Libellen konnten nachgewiesen werden, Kleinlibellen allerdings nur über winzige, lediglich wenige Millimeter große Eigelege. Diese sind in Doppelreihen auf dem Blatt eines Bedecktsamers deponiert und ähneln denen, die heute von Teichjungfern hinterlassen werden („Lestiden“-Typ).[45][46][47] In die Gruppe der Fluginsekten gehören auch Fächerflügler, wobei eine nur 140 μm lange und 90 μm breite Larve von Pseudococcites aus den Ausscheidungen eines Käfers einen der weltweit wenigen Belege dieses Entwicklungsstadiums der Insektengruppe bildet.[48] In sehr geringer Anzahl treten Spinnen und Tausendfüßer auf. Der Fund eines Saitenwurms der Gattung Gordius, der an Insekten parasitiert, stellt eine Besonderheit dar, da derartige Parasiten fossil nur äußerst selten vorkommen.[49][50]
Fische und Amphibien

Die Fischfauna ist mit rund 2000 Fundobjekten, darunter vielen vollständigen Skelettindividuen sehr umfangreich, umfasst aber insgesamt nur 5 Familien mit ebenso vielen Gattungen. Am häufigsten sind die Knochenfische, vertreten durch Thaumaturus und Palaeoesox. Ersterer gehört in die Familie der Lachsartigen, letzterer zu den Hechtartigen.[51] Ebenfalls ein Knochenfisch ist Anthracoperca, ein Verwandter der Barsche. Mit Cyclurus kommt auch ein Vertreter der Knochenganoiden vor.[52][53]
Unter den Amphibien sind überwiegend die Frösche und Molche von Bedeutung, die jeweils mit 200 bis 300 Exemplaren vorkommen, zuzüglich zahlreicher Einzelknochen. Die artikulierten Skelette der Frösche sind alle sehr fragil überliefert. Recht häufig ist Eopelobates aus der Familie der Europäischen Schaufelfußkröten, zu der auch die heutige Knoblauchkröte gehört. Wie diese lebte Eopelobates weitgehend terrestrisch. Dagegen war der ebenfalls häufige Palaeobatinopsis ein wasserbewohnendes Tier aus der Familie der Palaeobatrachidae, einer heute ausgestorbenen Froschgruppe. Weiteres Fossilmaterial kann wohl zu einem nahe verwandten Vertreter gehören.[54] Von den Molchen ist Palaeoproteus besonders zahlreich. Es handelt sich um einen amphibisch lebenden Olm, der bis zu 25 cm lang wurde und sehr kurze Gliedmaßen besaß. Seltener tritt Tylototriton auf. Diese als Krokodilmolche bezeichnete Gattung kommt noch in Ost- und Südostasien vor und ihre Vertreter gehören zu den Echten Salamandern.[55]
Reptilien und Vögel



Reptilien gehören zu den häufigsten Fossilfunden und können fast einem Dutzend Familien mit rund 20 Gattungen zugewiesen werden. Bedingt durch das feuchte Milieu sind zahlreiche Schildkröten überliefert, vor allem deren Panzerreste, die durch die Sedimentlast meist flachgedrückt sind. Alle aus dem Geiseltal bekannten Schildkröten können zu den Halsberger-Schildkröten gerechnet werden und umfassen rund 500 Exemplare. Dazu gehört unter anderem die Gattung Geiselemys aus der Gruppe der Altwelt-Sumpfschildkröten mit einem rund 18 cm langen gut verknöcherten Panzer. Ähnliche Ausmaße erreichte Borkenia, welche früher zur Gattung Chrysemys und damit in eine verwandtschaftliche Nähe mit den Zierschildkröten gestellt wurde. Am anderen Ende der Größenordnung steht Geochelone (auch als Barnesia bezeichnet), eine Landschildkröte, die einen durchschnittlich 50 bis 60 cm, möglicherweise auch bis zu 120 cm langen Panzer besaß.[56][57] Eine weitere, eher kleinere Form ist Hummelemys,[58] ihre genaue systematische Position ist aber umstritten.[57] Zu den Weichschildkröten zählt Palaeoamyda, deren Panzer rund 30 cm Länge erreichte. Das Tier war gut an ein Leben im Süßwasser angepasst.[59][60]

Schuppenkriechtiere sind mit fast 300 vollständigen Exemplaren zuzüglich zahlreicher Einzelfunde nachgewiesen. Dabei war Eolacerta ein recht großes Tier, das 60 cm lang wurde und mit mehreren vollständigen Skeletten und zusätzlich Häutungsresten überliefert ist. Es wurde ursprünglich zu den Echten Eidechsen gerechnet, einige besondere anatomische Merkmale führten jedoch zur Aufstellung der eigenständigen Familie der Eolacertidae, welche wiederum die Schwestergruppe der Echten Eidechsen bildet.[61][62] Zu den Leguanen ist Geiseltaliellus zu stellen, eine leicht gebaute Echse mit extrem langem Schwanz und feiner Hautbeschuppung, die wahrscheinlich ein Baumbewohner war. Aufgrund der kurzen Vordergliedmaßen wird vermutet, dass sich Geiseltaliellus wie die heutigen Basilisken gelegentlich nur auf den Hinterbeinen fortbewegen konnte.[63] Ophisauriscus kann wiederum den Schleichen zugeordnet werden und ist mit den Glasschleichen verwandt. Die schlangenähnlichen Tiere wiesen noch rudimentäre Vorder- und Hintergliedmaßen auf. Mehr als 20 Funde wurden dokumentiert, deren Erhaltungszustand von nahezu vollständigen Skeletten bis zu disartikulierten Einzelfunden sowie Resten der Hautpanzerung reichen. Die einzelnen Knochenplättchen des einstigen Schuppenkleids unterscheiden sich in Gestalt und Ornamentierung markant von den heutigen Formen.[64] Nur selten treten Verwandte der heutigen Warane auf. Ein schlecht erhaltenes Teilskelett von Eosaniwa besitzt einen 19 cm langen Schädel, damit war diese Echse eine der größten im Geiseltal.[65] Gut 60 vollständige Exemplare sind zu den Schlangen zu zählen, die unter anderem mit zwei Gattungen der Riesenschlangen aus der Familie der Boas überliefert sind. Dabei wurde Paleryx bis zu 2,3 m lang, bei einem weitgehend vollständigen Exemplar konnten insgesamt 243 Wirbel erkannt werden.[66] Des Weiteren kommt Palaeopython vor, ebenso wie einige Rollschlangen.[67][68]


Ebenfalls in großer Anzahl sind Krokodile mit rund 120 vollständigen Skeletten belegt, hinzu kommen noch unzählige isolierte Knochen und Zähne. Die Alligatoren vertritt dabei Allognathosuchus, das nur etwa 0,8 m lang wurde und eine recht schmale Schnauze besaß. Diplocynodon zählt zu den häufigsten Krokodilformen im Geiseltal und wird in die ausgestorbene Gruppe der Diplocynodontidae eingeordnet. Es lebte in kleineren Gewässern und erreichte gut 1,3 m Länge. Ein besonderer Fund umfasst ein nahezu vollständiges Skelett mit Knochenpanzer und fünf Eiern in unmittelbarer Nähe; es handelt sich um einen der weltweit seltenen Belege für Brutpflege bei fossilen Krokodilen.[69] Mit gut 1,7 m Länge etwas größer und ebenfalls zahlreich ist Boverisuchus, ein zu den Echten Krokodilen gehörender Vertreter. Aufgrund verhältnismäßig langer Gliedmaßen und hufartig verbreiteter Endglieder der Zehen wird für dieses Krokodil eine eher terrestrische Lebensweise angenommen. Den größten Beutegreifer im Geiseltal stellte Asiatosuchus mit rund 3 m Körperlänge dar. Dieser ähnelte ökologisch dem heutigen Nilkrokodil und bevorzugte offene Gewässer. Er ist mit mehreren vollständigen Skeletten nachgewiesen. Extrem selten mit nur einigen Unterkieferfragmenten trat dagegen Bergisuchus in Erscheinung, ein möglicherweise ebenfalls eher landbewohnendes Tier, das mit 1,5 m Länge von kleinwüchsiger Gestalt war. Im Gegensatz zu den anderen Formen repräsentiert Bergisuchus einen urtümlichen und entfernteren Krokodilsverwandten aus der Gruppe der Sebecosuchia.[70][71] Detaillierte Untersuchungen ergaben, dass alle Krokodile im Geiseltal unterschiedliche ökologische Nischen besetzten.[72][73] Daneben liegen Fragmente von Krokodileiern[74] und zahlreiche Gastrolithen (Magensteine) vor, die auch ohne Fossilerhaltung die Anwesenheit von Krokodilen belegen.[3]


Eher selten sind die Überreste von Vögeln, die mit rund einem halben Dutzend Familien und gut doppelt so vielen Gattungen nachgewiesen wurden. Meist sind aber nur Extremitätenreste überliefert. Einen relativ kleinen Vertreter der Urkiefervögel repräsentiert der über die hinteren Extremitäten und zusätzlich über ein Teilskelett nachgewiesene Palaeotis. Er wurde ursprünglich als verwandt mit den Trappen angesehen,[75] später aber als in der Ahnenreihe der heutigen Strauße stehend aufgefasst.[76] Neben den eindeutigen Merkmalen, die Palaeotis mit den Urkiefervögeln vereint, erinnern einige Charakteristika der Hinterbeine eher an heutige Kraniche.[77] Mit bis zu 1,8 m Gesamthöhe deutlich größer war Gastornis (auch Diatryma genannt). Von diesem großen bodenlaufenden Vogel liegen über drei Dutzend Fundobjekte vor, so mehrere Beinknochen, aber auch Schädelreste und Teile der Flügel, die die größte Kollektion aller Fundstellen innerhalb Europas darstellen.[78][79] Lange Zeit galt Gastornis als Fleischfresser, Isotopenuntersuchungen an einigen Knochen aus dem Geiseltal zeigten allerdings, dass der Vogel sich hauptsächlich vegetarisch ernährte.[80][81] Zu den echten fleischfressenden Vögeln wurde ursprünglich Eocathartes gezählt. Dieser sollte die Neuweltgeier im Geiseltal repräsentieren und ist anhand von Teilen des Körperskelettes als auch durch Federn beschrieben worden. Weiterhin hatte Geiseloceros als Angehöriger der Nashornvögel einen Status als Exot inne. Belegt war er über Beinknochen und zusammenhängende Flügelreste, an denen zusätzlich die blau schimmernden Schwungfedern erkennbar sind.[82] Beide Taxa werden heute zu Strigogyps aus der Familie der Ameghinornithidae gestellt, einer Gruppe größerer Laufvögel.[83] Über einen Oberarmknochen ist weiterhin Aegialornis, ein fossiler Segler, nachgewiesen.[84]
Säugetiere
Die sehr umfangreiche Säugetierfauna umfasst Reste aus rund zwei Dutzend Familien mit mehr als 50 Gattungen. Zu den urtümlichsten Vertretern gehören die Beuteltiere, die mit Peratherium und Amphiperatherium vertreten sind. Vor allem von Peratherium liegen mehrere Schädel und Unterkieferreste vor.[85] Beide Gattungen sind Ahnen des südamerikanischen Opossums. Taxonomisch schwer einzuordnen ist dagegen Microtarsioides, da zwar ein relativ vollständiges Skelett überliefert ist, das aber von einem juvenilen Tier stammt.[86]

Wesentlich umfangreicher und vielgestaltiger sind die Höheren Säugetiere. Einige wenige charakteristisch kräftig geformte Langknochen werden Eurotamandua zugewiesen.[86] Ursprünglich als Angehöriger der heutigen Ameisenbären angesehen,[87] stellt er aber nach weiteren Untersuchungen eine Basalform der Schuppentiere mit ähnlicher Lebensweise dar.[88] Zu den Kleinsäugetieren gehört Heterohyus, ein Apatemyide, der durch zwei verlängerte Finger und kreisbogenförmige Schneidezähne charakterisiert ist.[89] Die vergrößerten Vorderzähne dienten zum Benagen von Baumrinde, mit den langen Fingern bohrte er in Spalten und Rissen nach Insekten. In seiner Lebensweise ähnelte Heterohyus so dem heutigen Fingertier von Madagaskar. Mit einem Teilskelett und einem Unterkiefer ist Leptictidium nachgewiesen. Dieses räuberische Tier bewegte sich auf den Hinterbeinen springend fort, was die basale Stellung der Leptictida anzeigt, da dies bei heutigen Insektenfressern nicht mehr vorkommt.[90] Das nur selten auftretende Buxolestes war dagegen ein gedrungenes, semiaquatisch lebendes Tier aus der Gruppe der Pantolestidae, dessen Äußeres und Lebensweise etwa den heutigen Fischottern entsprach.[91] In das unmittelbare Verwandtschaftsumfeld gehören die Paroxyclaenidae, von denen aber nur ein 12 cm langer Schädel von Vulpavoides und ein 5 cm langes Unterkieferfragment von Pugiodens dokumentiert sind. Die kurzschnauzigen Tiere waren vermutlich baumbewohnend. Ihre Reste wurden ursprünglich den „Creodonta“, später den „Condylarthra“ beigeordnet.[92][93][94][95] Ebenfalls sehr selten, aber mit mehreren, teils vollständig bezahnten Unterkiefern ist der Insektenfresser Saturninia aus der Gruppe der Nyctitheriidae vertreten.[96][86] Nagetiere sind selten und kamen mit nur wenigen Formen vor. Ailuravus war sehr groß, rund einen Meter lang, und erinnerte an heutige Hörnchen. Wenige Gebissreste stammen vom wiederum deutlich kleineren Masillamys, die nach der Grube Messel benannte Messelmaus mit einem sehr langen Schwanz und einer Gesamtlänge von 40 cm. Bedeutend sind auch die Reste der Fledermäuse, von denen 25 vollständige Skelette mit Teilen der Flügelhaut vorliegen. Sie können weitgehend den Gattungen Matthesia und Cecilionycteris zugeordnet werden. Sie gehören zur Familie der Palaeochiropterygidae, deren Vertreter aufgrund des Baus der Ohrknöchelchen bereits mit Echoortung von 30 bis 70 kHz jagten.[97]
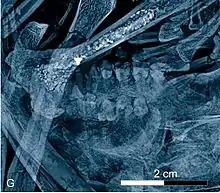
Bemerkenswerte Reste stammen auch von den Primaten. Hier sind vor allem die Adapidae wichtig, von denen mehrere Gattungen im Geiseltal auftraten und die möglicherweise Vorläufer der heutigen Lemuren waren. Mit mehreren Schädeln und Unterkiefern ist Europolemur überliefert. Eines der wenigen fast vollständigen Skelette weltweit eines adapiden Primaten liegt mit Godinotia vor. Dieser frühe Primat, der zur näheren Verwandtschaft des aus der Grube Messel bekannten Darwinius gehörte, besaß einen deutlich kurzen Gesichtsschädel und große Augenhöhlen, die auf eine nachtaktive Lebensweise hinweisen.[98][99][100] Sehr variantenreich mit mehreren Arten tritt Nannopithex auf, der aber zu einer moderneren Familie der Primaten gehört.[94]

Eher räuberisch lebende Tiere waren die heute ausgestorbenen „Creodonta“. Hierzu zählen vor allem die Hyaenodonta mit rund 50 Funden, darunter befinden sich überwiegend Unterkiefer, aber auch einige Schädel, Oberkieferfragmente und vereinzelte Reste des Bewegungsapparates. Dazu werden unter anderem Eurotherium, Matthodon, Prodissopsalis und Leonhardtina gestellt, die letzteren beiden waren am häufigsten und umfassen jeweils rund ein Dutzend Fossilien. Insgesamt waren alle im Geiseltal nachgewiesenen Creodontier relativ klein, so erreichte ein vollständiger Schädel von Eurotherium etwa 12 cm Länge.[101][102][103] Ein zerquetschter Schädel und ein einzelner hinterer oberer Prämolar vertreten die echten Raubtiere und wurden ursprünglich zu Miacis, heute aber zu Quercygale gezählt.[104][105]

Vergleichsweise umfangreich ist das Fossilmaterial der Paarhufer mit rund 170 Fundobjekten von etwa 60 Individuen. Die Paarhufer des Geiseltales zeichneten sich alle durch einen sehr altertümlichen Körperbau mit einem nach oben gewölbten Rückenverlauf, sehr langen Schwänzen und kurzen Vorder- sowie langen Hinterbeinen aus. Sie waren durchweg kleiner als die heutigen Verwandten und erreichten maximal Ferkelgröße. Einer der urtümlichsten Paarhufer war Diacodexis aus der Gruppe der Diacodexeidae, das allerdings nur mit wenigen Knochenresten überliefert ist. Etwas weiter entwickelt ist das durch einen zerdrückten Schädel nachgewiesene Eurodexis aus der nahe verwandten Gruppe der Dichobunidae.[106] Zu seiner näheren Verwandtschaft gehören Messelobunodon und Aumelasia, beide sind ebenfalls mit wenigen Funden belegt. Am häufigsten konnten bisher die Vertreter der etwas weiter entwickelten Choeropotamidae beobachtet werden, die in die nähere Verwandtschaft der Flusspferde zu stellen sind und endemisch im heutigen Europa verbreitet waren. Ein nahezu vollständiges Skelett liegt von Masillabune vor,[107] während Hallebune nur mit wenigen Gebissfragmenten vertreten ist.[108][109] Mehrere Skelettreste, darunter sieben fast vollständige, sind Amphirhagatherium zuzuweisen. Dieses rund 70 cm lange und etwa 4 kg schwere Tier war nicht nur der häufigste, sondern auch der größte Paarhufer des Geiseltales und ist weit über die Unter- und Mittelkohle verteilt.[110][111] Darüber hinaus sind noch wenige Reste anderer Paarhufer wie Haplobunodon oder Rhagatherium bekannt.[112]


Von den Unpaarhufern liegt das umfangreichste Fundmaterial vor. Auch diese besaßen einen altertümlichen Körperbau mit einem aufgewölbten Rücken und vier Zehen an den Vorder- und drei an den Hinterfüßen, was heute nur noch bei den Tapiren vorkommt. Über 310 Unter- und 120 Oberkieferreste einschließlich zweier vollständiger Skelette stammen von den Pferdeverwandten aus der Gruppe der Palaeotheriidae.[113] Hervorzuheben ist das bekannte Urpferdchen Propalaeotherium, das mit mehreren Arten vertreten ist. Ein vollständiges Skelett mit einem 56 cm langen Rumpf, einem 20 cm langen Schädel und einer Schulterhöhe von 40 cm wurde 1933 entdeckt. Andere vorkommende Gattungen sind Lophiotherium, Hallensia und Eurohippus. Von dem sehr seltenen Plagiolophus liegen nur wenige Zähne vor.[114][115][116] Mit fast 180 Individuen ebenfalls sehr häufig ist Lophiodon, ein Tapirverwandter, der mit einer Körperlänge von 2,5 m und einer Schulterhöhe von 1 m den größten Vertreter der Säugetiere aus dem Geiseltal repräsentiert. Allein auf einer Fläche von 131 m² wurden 110 Individuen entdeckt, wohl Reste eines Krokodilfraßplatzes. Fossilfunde von Lophiodon sind aus allen fossilführenden Flözen bekannt, wobei die Gattung dabei von unten nach oben eine bedeutende Körpergrößenzunahme durchlief.[117][118][119] Daneben erschien sein Verwandter Hyrachyus seltener und konnte mit etwa 75 Gebissresten und einigen Teilen des Körperskeletts nachgewiesen werden.[120] Hyrachyus war moderner gebaut als Lophiodon und steht an der Basis der Entwicklung der Nashörner und Tapire.[121]
Weichteile, Chemofossilien und Nahrungsreste

In der Braunkohle des Geiseltals haben sich auch Überreste der Weichteile erhalten, was äußerst selten vorkommt. Erstmals nachgewiesen wurden fossile Weichteile im Geiseltal 1934; sie gehören damit zu den ersten derartigen Funden aus dem Eozän überhaupt. Die Entdeckungen gelangen erst mit der Entwicklung der Lackfilmmethode. So konnte bei einzelnen Fischen unter anderem die feine Schuppenreliefierung erkannt werden, etwa bei Thaumaturus, ebenso bei Schuppenkriechtieren und seltener bei Krokodilen.[122] Bei zahlreichen Fröschen ließ sich die Haut untersuchen, deren einzelne Schichten, die Epidermis und Dermis, bis zu den einzelnen Zellen erkennbar sind.[33] An einigen Resten von Vögeln konnten Federn festgestellt werden, die zum Teil gefärbt waren;[123] von Fledermäusen ist die Flughaut überliefert. Bei Säugetieren, wie Amphirhagatherium und seltener Propalaeotherium, sind Teile des Haarkleids nachgewiesen.[124] Es gelang auch, Muskulatur, Knorpel und Ähnliches bis hin zu Blutzellen zu dokumentieren.[125]
Darüber hinaus ließ sich die ursprüngliche Färbung bestimmter Weichteile, vor allem der Haut, erkennen. So war der Fisch Palaeoesox am Körper zebragemustert und hatte einen dunkel gefärbten Rücken, während bei Fröschen eine grünliche Hauttönung beobachtet werden konnte. Bei Insekten sind die Farbschattierungen der Tiere erhalten, bei den Pflanzen hat das Chlorophyll überdauert.[126] Zu solchen Chemofossilien gehört auch das oft in großen Mengen vorkommende „Affenhaar“. Hierbei handelt es sich um faserige, durch die Lagerung in der Braunkohle flachgedrückte Gebilde von hellbrauner Farbe, die durch natürliche Schwefelvulkanisiation überliefert blieben. Sie stellen fossilisierte Milchsaftröhren gummiproduzierender Gehölze aus der Familie der Hundsgiftgewächse dar, von denen Blätter der Gattung Apocynophyllum aus dem Geiseltal vorliegen. Nach der rezenten Gattung Couma wird das „Affenhaar“ als Formtaxon aber zu Coumoxylon verwiesen.[127][36]
Weitere Funde sind vor allem die Nahrungsreste zahlreicher Tiere, die ebenfalls bereits früh entdeckt wurden, so 1935 beim Tapirverwandten Lophiodon als grünliche Pflanzenmasse.[128] Sie sind auch von anderen Säugetieren wie dem Urpferdchen Propalaeotherium[129] und dem Paarhufer Amphirhagatherium nachgewiesen,[110][111] darüber hinaus auch von dem straußenartigen Vogel Palaeotis. Wenigstens 100 Exemplare von Koprolithen und zusätzliche Fragmente wurden ebenfalls dokumentiert. Sie liegen zumeist in ei- oder wurstartiger Form vor mit Längen von wenigen Millimetern bis hin zu einem Dezimeter und enthalten teils Reste von Knochen, Krallen oder Pflanzen. Als Verursacher können verschiedene Reptilien und Säugetiere angenommen werden. Einzelne der versteinerten Kotreste enthalten auch Gastrolithen, die eventuell von Krokodilen stammen. Krokodile hinterließen verschiedentlich auch Bissmarken an Säugetierknochen und Schildkrötenpanzern.[130]
Datierung
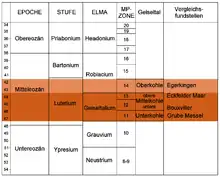
Bedeutend für die genaue Alterseinstufung der Braunkohle des Geiseltales sind die faunistischen Überreste, insbesondere der Säugetiere, was darüber hinaus einen Vergleich mit anderen Fundstellen und Aufschlüssen ermöglicht. Diese biostratigraphische Altersbestimmung erfolgt weitgehend über die Veränderungen der Zahnmorphologie bestimmter Säugetiergruppen, womit das zeitliche Auftreten ausgestorbener Arten und Gattungen ermittelt werden kann. Bereits die ersten, Anfang des 20. Jahrhunderts entdeckten Wirbeltierfunde, Zahnreste des Tapirverwandten Lophiodon, veranlassten die Einstufung der Braunkohle in die geologische Epoche des Mittleren Eozäns. Damit war auch das genauere Relativalter der Braunkohlen ermittelt, das vorher nur vermutet wurde.[131] Das Mittlere Eozän wird auf ein Alter von 47,8 bis 41,3 Millionen Jahren datiert und besteht aus der unteren Stufe Lutetium und der oberen Stufe Bartonium. Über das Vorkommen bestimmter Säugetierformen kann das genauere Alter der Geiseltaler Braunkohlen relativ genau ermittelt werden. Dadurch werden die reichhaltigen Funde der Unterkohle bis zur Oberen Mittelkohle in die Zone des Geiseltaliums gestellt, eine Stufe innerhalb der Stratigraphie der europäischen Landsäugetiere (European Land Mammal Ages, ELMA). Dem Geiseltalium dient die Geiseltalfauna als Referenz, zudem bildet es einen mittleren Abschnitt des Lutetium. Die nur wenigen Funde der Oberkohle werden in eine jüngere Zone, das Robiacium verwiesen. Der Beginn des Geiseltaliums geht dabei mit dem Auftreten früher Formen des Urpferdchens Propalaeotherium, aber auch seines Verwandten Lophiotherium oder des Primaten Europolemur einher, während das Ende durch das erste Auftreten von Primaten wie Adapis angezeigt wird.[26][132]
Absolute Datierungen mittels radiometrischer Messverfahren liegen aus dem Geiseltal nicht vor. Allerdings wurden solche in etwa gleich alten Fundstellen vorgenommen. So stammen Daten, ermittelt mit der Kalium-Argon-Datierung, aus der bedeutenden Grube Messel in Hessen, die aus biostratigraphischer Sicht dem Abschnitt der Unterkohle des Geiseltales entspricht, und ergaben ein Alter von 47,8 Millionen Jahren. Da der untersuchte Basalt aus einem Bereich unter den fossilführenden Seesedimenten der Grube Messel stammt, ist der Alterswert als Maximalalter anzusehen, die Funde dort sind also etwas jünger (terminus post quem).[133] Das Eckfelder Maar in Rheinland-Pfalz stellt ein biostratigraphisches Äquivalent zur oberen Mittelkohle dar. Dort mit Hilfe der gleichen Messmethodik gewonnene Daten aus Pyroklasten, ebenfalls unterhalb der Fundschicht, ergaben einen Wert von 44,3 Millionen Jahren. Dies ist ebenfalls als unterstes Alter anzusehen.[134] Dadurch ist die Alterseinstufung des Geiseltaliums, dem heute ein Alter von 47,4 bis 43,4 Millionen Jahren zugesprochen wird, und damit der Funde aus dem Geiseltal als recht sicher anzusehen.[132]
Landschaftsrekonstruktion
Die zahlreichen Pflanzen- und Tierreste, aber auch die hohe Anzahl an geologischen Daten ermöglichen eine recht genaue Rekonstruktion der ehemaligen Landschaftsverhältnisse. Demnach war das Geiseltal im Mittleren Eozän eine wasserreiche Moorlandschaft, die sich in Nord-Süd-Richtung auf einer Länge von vier bis fünf Kilometern erstreckte und die in Küstennähe an einer weit ins Landesinnere eindringenden Bucht lag. Dieser als Mitteldeutscher Ästuar bezeichnete Landeinschnitt, der anhand von Pollenfunden der Palmengattung Nypa aus der Mittelkohle zumindest zeitweilig den Gezeiten ausgesetzt war,[135] bildete vor allem im mittleren Abschnitt des Eozäns (genauer im Lutetium) zahlreiche Buchten sowie lokale Senkungen und Becken, die die Entstehung von Braunkohle förderten, so das Helmstedter Braunkohlerevier im Nordwesten oder die Braunkohlelager von Egeln und Edderitz weiter südlich. Dieses verzweigte System führte auch im Geiseltal zur Bildung mariner, brackiger bis hin zu fluviatilen und limnischen Ablagerungen (die Hauptmittel) als Hinterlassenschaften des Ästuars. Das damals subtropische Klima trug zur Entstehung von zahlreichen Mooren, Teichen und Tümpeln mit einem reichen Vegetationsbestand am Rande des Ästuars bei. Durch das Einsetzen von Inkohlung des abgestorbenen Pflanzenmaterials kam es in einem Zeitraum von sechs bis möglicherweise acht Millionen Jahren zur Ausbildung mächtiger Braunkohleflöze.[4] Begrenzt wurde das Gebiet im Süden von einer Muschelkalkhochfläche mit steilen, nach Norden abfallenden Hängen. Die Hochfläche selbst war zum Teil verkarstet. Durch die chemische Verwitterung entstanden Kalkwässer, die in die Moorniederung eindrangen und für die vorzügliche Erhaltung der Fossilien sorgten.[136]

Durchsetzt war das Moorgebiet von zahlreichen kleinen, im Durchmesser rund acht bis zehn Meter großen Tümpeln und Teichen, die durch Einsturztrichter angezeigt werden und der fossilen Tierwelt als Trinkstellen dienten. Im angrenzenden Uferbereich wuchsen Acrostichum-, Myricaceen- und Restionaceen-Pflanzengemeinschaften. In der weiteren Umgebung gab es Strauch- und Waldlandschaften aus Palmen-Kiefernwäldern, durchsetzt mit Mammutbäumen und anderen Nadel- und Laubhölzern, die kraut- und lichtreich waren und mehrere Wuchsebenen aufwiesen. Diese Wälder waren durchzogen von einem Netz mäandrierender Fließgewässer, die periodisch über die Ufer stiegen. Nach Süden in Richtung des Muschelkalkzuges gingen diese Wälder in einen dichten Urwald aus Lorbeergewächsen über. Die Höhenlage dürfte aufgrund der relativen Küstennähe damals nicht über 50 m über dem Meeresspiegel betragen haben. Das Klima war insgesamt subtropisch mit frostfreien und trockenen Wintern mit minimal 5 °C Durchschnittstemperatur und feuchten Sommern mit einer bis zu neun Monate langen Vegetationsperiode und durchschnittlich 25 °C im wärmsten Monat, was anhand der zahlreichen Früchte und Samen ermittelt wurde. Der jährliche Niederschlag dürfte bei rund 2070 mm gelegen haben. Durch die deutlichen Temperaturunterschiede während der Sommer- und Wintermonate unterlag die Landschaft einer jährlichen Periodik aus Regen- und Trockenzeit, was sich unter anderem auch durch Baumringe sowie durch eine feine Warvung der Braunkohle nachweisen lässt. Dadurch ist der Laubfall als wahrscheinlich anzusehen, so dass die Blätter eine lockere Streuschicht am Boden bildeten. Von der Unter- zur Mittelkohle konnte allerdings eine zunehmende Trockenheit des Klimas ermittelt werden.[137][72][111][138]
Diese vielgestaltige Landschaft war von zahlreichen Tieren bewohnt. Vor allem in den Wäldern lebten blatt- und fruchtfressende Säugetiere wie Lophiodon, Propalaeotherium und Amphirhagatherium. Die reichhaltige Insektenfauna war wiederum Ernährungsgrundlage zahlreicher weiterer Tiere, wie Frösche, Molche und Schuppenkriechtiere, aber auch Leptictiden, Fledermäuse und einige Primaten. Die Teiche und Tümpel beherbergten wasserbewohnende oder amphibisch lebende Tiere. Hervorzuheben sind die zahlreichen Krokodilformen, die wie heute ein deutlich wärmeres Klima bevorzugten und dadurch auch einen wichtigen Klimaanzeiger darstellen.[137][72] Die Reichhaltigkeit der Ökosystems und der lange Ablagerungszeitraum von rund 3 Millionen Jahren lässt darüber hinaus auch bestimmte Beziehungsgeflechte erkennen, die über reine Nahrungsketten hinausgehen. Dies lässt sich unter anderem an Lophiodon und Propalaeotherium erkennen, die die beiden häufigsten Säugetierformen repräsentieren und aufgrund der Nutzung der gleichen Nahrungsressourcen wohl eine besondere ökologische Beziehung zueinande hatten. Während aber Lophiodon im Laufe der Zeit an Körpergröße zunahm (durchschnittlich 124 kg in der Unterkohle und 223 kg in der Oberkohle), büßte Propalaeotherium hingegen an Gewicht ein (durchschnittlich 39 kg in der Unterkohle und 26 kg in der Oberen Mittelkohle). Möglicherweise ist dies ein Beispiel für die allmähliche Anpassung an unterschiedliche ökologische Nischen zur Vermeidung zu starker Konkurrenz untereinander.[138]
Vergleich mit regional und überregional bedeutenden Fundstellen
Aus der regionalen Nachbarschaft des Geiseltales sind vor allem die gleich alten Braunkohleaufschlüsse etwa aus der Leipziger Umgebung (das Weißelsterbecken) oder dem Helmstedter Raum von Bedeutung. Obwohl hier keine Vertebratenreste vorliegen, sind unzählige floristische Funde bekannt. So kommen in den Tagebauen bei Helmstedt Makroreste in Form von Zapfen von Koniferen und Palmenblättern vor, aus dem Tagebau Profen stammen wiederum zahlreiche Blattfunde von Zweikeimblättrigen.[37] Die allgemeine, anhand der Mikroflora ermittelte Vegetationsgeschichte entspricht weitgehend der des Geiseltales.[34] Gleich alte Wirbeltierfundstellen sind aus der näheren Region nicht bekannt. Mit den Funden von Walbeck im Nordwesten von Sachsen-Anhalt ist aber eine der umfangreichsten Säugetiergemeinschaften weltweit aus dem Mittleren Paläozän vor knapp 60 Millionen Jahren bekannt. Die mehrere Tausend Knochen- und Zahnreste umfassende Fauna unterscheidet sich von jener des Geiseltals durch die Dominanz urtümlicher Säugetiere aus den Gruppen der Procreodi, „Condylarthra“ oder Leptictida, während Vertreter modernerer, heute noch bestehender Linien wie den Primaten eher selten auftreten oder wie bei den Paarhufern und Unpaarhufern vollständig fehlen. Walbeck stellt somit ein bedeutendes Zeugnis aus der Frühphase der Radiation der Säugetiere kurz nach dem Aussterben der Dinosaurier dar.[139][140]
Im überregionalen Vergleich eine herausragende Bedeutung haben die Grube Messel in Hessen und das Eckfelder Maar in Rheinland-Pfalz. Beide stellen Fundstellen in ehemaligen Maaren dar, wobei Messel etwa zeitgleich zur Unterkohle des Geiseltales, Eckfeld aber zur Oberen Mittelkohle ist. Von beiden Fundstellen ist neben Pflanzen- und Invertebratenfunde eine hohe Anzahl von Wirbeltierresten bekannt, die in Messel gegenwärtig rund 130 Taxa umfasst, die Menge der Funde aus Eckefeld ist dagegen vergleichsweise geringer. Während die Zusammensetzung der Säugetierfauna generell Übereinstimmungen zeigt, gibt es aber im Einzelnen deutliche Unterschiede. So sind im Geiseltal und in Eckfeld etwa die Nagetiere und Fledermäuse[97] untervertreten, kommen in Messel aber häufiger und formenreicher vor. Dagegen zeichnet sich Messel durch eine geringere Anzahl an Primaten- und Paarhuferformen aus, die im Geiseltal und in Eckfeld wiederum gut und vielfältig belegt sind, wobei sich markante Abweichungen im Auftreten verschiedener Gattungen widerspiegeln. Ähnliche Differenzen können auch für einzelne Säugetiergattungen herausgearbeitet werden, etwa Lophiodon aus der Gruppe der Unpaarhufer, das im Geiseltal zu den dominierenden Formen gehört, in Messel aber nur mit einem Jungtier und einzelnen Zahnfunden, in Eckfeld mit einem Unterkiefer und ebenfalls wenigen Zahnresten nachgewiesen werden konnte. Bemerkenswert in Eckfeld ist das bisherige Fehlen altertümlicher Säugetiergruppen wie den „Creodonta“ und „Condylarthra“, die zumindest in Messel und im Geiseltal in geringem Umfang auftreten.[3][141][142] Darüber hinaus lassen sich auch in anderen Fundgruppen Unterschiede aufzeigen. So sind die Prachtkäfer mit mehr als einem Viertel aller Käferfunde verhältnismäßig häufig im Geiseltal überliefert, in Messel dagegen mit weniger als einem Zehntel eher rar, allerdings zeigt sich hier eine allgemein höherer Formenreichtum an Käfern insgesamt. Dass dabei unter anderem nur wenige Formen der Prachtkäfer an beiden Fundstellen gleichzeitig vorkommen, weist auf bedeutende ökologische Unterschiede hin, die sich auch in der teils abweichenden Vielfalt der verschiedensten Pflanzen- und Tiergruppen niederschlägt und aufzeigt, dass die einzelnen Fossillagerstätten nur jeweils einen Ausschnitt aus der damaligen reichhaltigen Landschaft repräsentieren.[44]
Forschungsgeschichte
Wissenschaftliche Ausgrabungen

Obwohl der Abbau von Braunkohle im Geiseltal nachweislich bis in das Jahr 1698 zurückreicht,[2][143] wurden die ersten Fossilien relativ spät entdeckt. Der früheste Fossilfund datiert in das Jahr 1908 und umfasst Reste des Tapirverwandten Lophiodon, die eher zufällig von einem Steiger im Tagebau Cecilie aufgesammelt wurden. Weitere Funde wurden im Jahr 1912 geborgen, die mehrere Gebissreste dreier Individuen einschließen und zur selben Gattung gehören. Im darauffolgenden Jahr konnten erstmals Reste von Schildkröten in einer pflanzenführenden Kohleschicht beobachtet werden, wobei aber ein Teil der Funde nach der Freilegung zerfiel. Einige verbliebene Fossilien wurden der Geologischen Landesanstalt in Berlin übergeben.[131][39]
Weitere Schildkrötenfunde im Jahr 1925 führten dann zu systematischen wissenschaftlichen Ausgrabungen, die der Geologe Johannes Walther (1860–1937) initiierte und welche die Universität Halle koordinierte. Die Leitung vor Ort übernahm der gebürtige Engländer Ben Barnes. Er untersuchte vor allem verschiedene Bereiche im Tagebau Cecilie. Ziel war es dabei, nicht nur qualitativ vollständige Stücke zu erfassen, sondern auch quantitativ einen Überblick über den Fossilgehalt der Braunkohlen zu gewinnen, wodurch auch kleinere oder schlecht erhaltene Fundobjekte dokumentiert wurden. Barnes Ausgrabungen deckten unter anderem Reste von Schlangen und Schildkröten sowie Zähne und Knochen verschiedener Huftiere auf. Die Arbeiten resultierten in einer ersten umfassenderen wissenschaftlichen Publikation im Jahr 1927.[144][145] Nach dem Ausscheiden Walthers aus dem Lehrbetrieb führte diese Johannes Weigelt (1890–1948) weiter. Zum Zweck der besseren Haltbarmachung der Fossilien entwickelte Ehrhard Voigt (1905–2004) Anfang der 1930er-Jahre die Lackfilmmethode.[29] Ein erster Höhepunkt war 1933 mit der Entdeckung eines vollständigen, 74 cm langen Skelettes des Urpferdchens Propalaeotherium im Tagebau Cecilie erreicht, das im Zentrum eines Einsturztrichters lag und aus dem weiterhin zahlreiche Krokodilreste und teils vollständige Eidechsenskelette stammen.[32] Im gleichen Tagebau wurde nur ein Jahr später die einzige bedeutende Fundstelle der Oberkohle mit über 20 Individuen von Lophiodon untersucht, darunter auch ein nahezu vollständiges Skelett.[128] Im Tagebau Cecilie kamen die wissenschaftlichen Forschungen im Jahr 1935 aufgrund des Erschöpfens der Kohlevorräte zum Erliegen. Weitere Grabungen erfolgten daraufhin unter anderem im Tagebau Leonhardt, kamen aber 1938 zum Stillstand. Der Ausbruch des Zweiten Weltkrieges 1939 beendete diese erste wichtige Forschungsperiode endgültig.[25]
Im Jahr 1949 wurden die Grabungstätigkeiten wieder aufgenommen und im Folgenden von der Regierung der DDR im größeren Rahmen gefördert. Zudem konnten zahlreiche Wissenschaftler aus anderen Nationen in die Auswertung der Funde eingebunden werden. Durch die Ausweitung des Tagebaubetriebes, der in der Mitte des 20. Jahrhunderts seinen Höhepunkt fand, wurden zahlreiche weitere Fundstellen unter anderem in den Abbaufeldern Pfännerhall und Mücheln (welches die ehemaligen Bereiche Pauline, Elisabeth, Emma und Elise II des westlichen Geiseltales umfasste) erschlossen. Dabei entdeckten die Wissenschaftler mit der Fundstelle XIV eine der fossilreichsten und konnten dort mit den Bachläufen neben den bereits von Weigelt definierten Fundstellentypen einen neuen belegen. Die erhöhte Kohleförderung führte dazu, dass abbaubegleitend verstärkt Grabungsmaßnahmen erfolgten, sodass vor allem die 1960er- und 1970er-Jahre einen Höhepunkt der Erforschung des Geiseltales darstellten. Erst mit dem sich abzeichnenden Ausklingen der Kohlevorräte (das östliche und südöstliche Geiseltal war bereits Anfang der 1970er-Jahre erschöpft und die aufgelassenen Tagebaue teilweise geflutet worden) und der Verlagerung des Abbaus in den nördlicheren und westlicheren Teil des Geiseltales, wo die Fossilerhaltung weniger gut war, gingen auch die wissenschaftlichen Vorortuntersuchungen zurück und endeten vorläufig 1985. Während dieser Forschungsphase waren mehr als 55 neue Fundstellen entdeckt worden, darunter 23 mit Wirbeltierresten.[25][27]
Nach der politischen Wende in der DDR im Jahr 1989 änderte sich die Situation grundlegend. Nachdem ursprünglich die Förderung der Braunkohle im westlichen Geiseltal noch bis 1998 weitergeführt werden sollte, wurde diese jedoch aufgrund veränderter Marktbedingungen Mitte 1993 eingestellt und mit der Renaturierung des gesamten Tagebaugebietes begonnen. Dadurch kam es von Frühjahr bis Herbst 1992 zu den letzten Untersuchungen einer Fundstelle im südlichen Geiseltal, wobei hier auch Mitarbeiter des Forschungsinstitutes Senckenberg beteiligt waren. Im darauf folgenden Jahr wurde diese letzte Wirbeltierfundstelle von aufsteigendem Grundwasser überspült.[27] Zwischen den Jahren 2000 und 2003 fanden die letzten Geländeaktivitäten wiederum gemeinsam mit dem Forschungsinstitut Senckenberg im westlichen Geiseltal statt, die vorwiegend einen über 20 m mächtigen Kohleflöz betrafen, der unzähliges Pflanzenmaterial bis hin zu 25 m langen Baumstämmen enthielt. Aufgrund der chemischen Eigenschaften der Braunkohle in diesem Bereich des Geiseltales konnten allerdings keine Wirbeltierreste beobachtet werden.[47] Im Juni 2003 begann die planmäßige Flutung des westlichen Geiseltales zum Geiseltalsee, der im Frühjahr 2011 seine Endwasserhöhe erreicht hatte, wodurch einer der größten künstlichen Seen Mitteleuropas geschaffen wurde. Damit waren die wissenschaftlichen Tätigkeiten vor Ort endgültig beendet.[2]
Fundpräsentation

Bereits 1934 wurde nur neun Jahre nach Beginn der Grabungstätigkeiten aufgrund des immens angestiegenen Fundmaterials das Geiseltalmuseum in der im Auftrag Kardinal Albrechts II. in den Jahren 1531 bis 1537 errichteten Neuen Residenz nahe dem Halleschen Dom in der Saalestadt Halle eröffnet. Das Museum diente gleichzeitig als Sammlungsdepot und Ausstellung, wobei diese in der Allerheiligenkapelle auf einer Fläche von 267 m² eingerichtet wurde. Die anderen Räumlichkeiten der Neuen Residenz nutzte dagegen das Geologisch-Paläontologische Institut der Martin-Luther-Universität Halle-Wittenberg. Als Wappentier des Museums diente das Propalaeotherium zugewiesene Urpferdchenskelett aus dem Jahr 1933. Eine Unterbrechung der Ausstellung erfolgte kurz vor Ende des Zweiten Weltkrieges im Jahr 1945, als die Funde aus Sicherheitsgründen ausgelagert werden mussten.[146][3]
Im Jahr 1950 eröffnete die Ausstellung wieder unter dem Motto „Gang durch Mitteldeutschlands Erdgeschichte“ – erweitert um acht Räume und 1954 fachlich überarbeitet. Infolge einer Hochschulreform Ende der 1960er Jahre sollten Museum und Sammlung ausgelagert werden, was vom damaligen Direktor Horst Werner Matthes abgewendet werden konnte. Allerdings musste die 1950 eröffnete neue Ausstellung geschlossen und reduziert werden. Technische und finanzielle Engpässe führten in den folgenden Jahren dazu, dass weder in eine modernere Ausstellung noch in eine Reparatur der Gebäude der Neuen Residenz investiert werden konnte, wodurch unter anderem durch Regeneinbrüche oder Tauwetter die Notwendigkeit bestand, einzelne Fundobjekte vor Beschädigung zu bewahren. Erst nach 1989 mit der politischen Wende in der DDR änderte sich dies zum Positiven.[146] Allerdings blieb die Bausubstanz der Neuen Residenz weiter in schlechtem Zustand. Als das jetzige Institut für Geowissenschaften im Jahr 2004 in neue und modernere Räumlichkeiten am neuen Campus am Von-Seckendorf-Platz zog, verblieb nur das Geiseltalmuseum samt Sammlung zurück und feierte dort sein 70-jähriges Bestehen.[47]
Von Ende 2011 bis Mai 2018 war das Museum geschlossen und die Sammlung nicht öffentlich zugänglich. Die Wiedereröffnung erfolgte im Rahmen der Langen Nacht der Museen am 5. Mai 2018.[147][148] Im Frühjahr 2015 informierte eine Sonderausstellung mit dem Titel „Aus der Morgendämmerung: Pferdejagende Krokodile und Riesenvögel“ im Tschernyschewski-Haus der Leopoldina in Halle über die neuesten Ergebnisse aus der Erforschung der Fossillagerstätte Geiseltal.[149] Zwischen November 2017 und Mai 2018 waren einige bedeutende Fossilien des Geiseltals Bestandteil der Sonderausstellung „Klimagewalten – Treibende Kraft der Evolution“ am Landesmuseum für Vorgeschichte in Halle.[150] Im Jahr 2012 erhielt die gesamte Sammlung den Status als „national wertvolles Kulturgut“ und steht daher unter Schutz der Regierung.[145][151]
Literatur
- Alexander K. Hastings, Meinolf Hellmund: Aus der Morgendämmerung: Pferdejagende Krokodile und Riesenvögel. Neueste Forschungsergebnisse zur eozänen Welt Deutschlands vor ca. 45 Millionen Jahren. Halle (Saale), 2015, S. 1–120.
- Meinolf Hellmund: Exkursion: Ehemaliges Geiseltalrevier, südwestlich von Halle (Saale). Aus der Vita des eozänen Geiseltales. In: Jörg Erfurt, Lutz Christian Maul (Hrsg.): 34. Tagung des Arbeitskreises für Wirbeltierpaläontologie der Paläontologischen Gesellschaft 16. bis 18. März 2007 in Freyburg/Unstrut. In: Hallesches Jahrbuch für Geowissenschaften. Beiheft 23, 2007, S. 1–16.
- Günter Krumbiegel, Ludwig Rüffle, Hartmut Haubold: Das eozäne Geiseltal: ein mitteleuropäisches Braunkohlenvorkommen und seine Pflanzen- und Tierwelt. Ziemsen, Wittenberg 1983, S. 1–227, OCLC 9998655.
Einzelnachweise
- Krumbiegel, Rüffle, Haubold 1983, S. 7–14.
- Joachim Wirth, Rudolf Eichner, Andreas Schroeter: Revier Halle und Geiseltal. In: Gerhard H. Bachmann, Bodo-Carlo Ehling, Rudolf Eichner, Max Schwab (Hrsg.): Geologie von Sachsen-Anhalt. Stuttgart 2008, S. 491–493.
- Meinolf Hellmund: Exkursion: Ehemaliges Geiseltalrevier, südwestlich von Halle (Saale). Aus der Vita des eozänen Geiseltales. In: Jörg Erfurt, Lutz Christian Maul (Hrsg.): 34. Tagung des Arbeitskreises für Wirbeltierpaläontologie der Paläontologischen Gesellschaft 16. bis 18. März 2007 in Freyburg/Unstrut. (Hallesches Jahrbuch für Geowissenschaften BH 23), 2007, S. 1–16.
- Karl-Heinz Radzinski, Bodo-Carlo Ehling, Reinhard Kunert, Gerhard Beutler: Südöstliches Harzvorland. In: Gerhard H. Bachmann, Bodo-Carlo Ehling, Rudolf Eichner, Max Schwab (Hrsg.): Geologie von Sachsen-Anhalt. Stuttgart 2008, S. 458–471.
- Krumbiegel, Rüffle, Haubold 1983, S. 23.
- Matthias Thomae, Ivo Rappsilber: Zur Entstehung der Geiseltalsenke. In: Harald Meller (Hrsg.): Elefantenreich – Eine Fossilwelt in Europa. Halle/Saale 2010, S. 27–33.
- Matthias Thomae, Ivo Rappsilber: Beitrag zur Klärung der Lagerungsverhältnisse des Quartärs im Tagebau Neumark-Nord. In: Dietrich Mania u. a. (Hrsg.): Neumark-Nord: Ein interglaziales Ökosystem des mittelpaläolithischen Menschen. (Veröffentlichungen des Landesmuseums für Vorgeschichte, 62). Halle/Saale 2010, S. 71–80.
- Horst Blumenstengel, Wilfried Krutzsch: Tertiär. In: Gerhard H. Bachmann, Bodo-Carlo Ehling, Rudolf Eichner, Max Schwab (Hrsg.): Geologie von Sachsen-Anhalt. Stuttgart 2008, S. 267–292.
- Hartmut Haubold, Matthias Thomae: Stratigraphische Revision der Wirbeltierfundstellen des Geiseltales. In: Hallesches Jahrbuch für Geowissenschaften. 15, 1990, S. 3–20.
- Thomas Litt, Karl-Ernst Behre, Klaus-Dieter Meyer, Hans-Jürgen Stephan, Stefan Wansa: Stratigraphische Begriffe für das Quartär des norddeutschen Vereisungsgebietes. In: Eiszeitalter und Gegenwart (Quaternary Science Journal). 56, 2007, S. 7–65 ().
- Dietrich Mania: Zur Geschichte des Geiseltals im Quartär. In: Harald Meller (Hrsg.): Elefantenreich – Eine Fossilwelt in Europa. Halle/Saale 2010, S. 34–38.
- Dietrich Mania: Quartärforschung im Tagebau Neumark-Nord, Geiseltal (Sachsen-Anhalt) und ihre bisherigen Ergebnisse. In: Dietrich Mania u. a. (Hrsg.): Neumark-Nord: Ein interglaziales Ökosystem des mittelpaläolithischen Menschen. (Veröffentlichungen des Landesmuseums für Vorgeschichte, 62). Halle/Saale 2010, S. 11–69.
- Lothar Eißmann: Mollisoldiapirismus. In: Zeitschrift für angewandte Geologie. 24, 1978, S. 130–138.
- Thomas Laurat, Enrico Brühl: Zum Stand der archäologischen Untersuchungen im Tagebau Neumark-Nord, Ldkr. Merseburg-Querfurt (Sachsen-Anhalt) – Vorbericht zu den Ausgrabungen 2003–2005. In: Jahresschrift für mitteldeutsche Vorgeschichte. 90, 2006, S. 9–69 ().
- Thomas Litt: Zur stratigraphischen Einstufung von Neumark-Nord aufgrund neuer pollenanalytischer Befunde. In: Altenburger naturwissenschaftliche Forschungen. 7, 1994, S. 328–333.
- Jaqueline Strahl, Matthias R. Krbetschek, Joachim Luckert, Björn Machalett, Stefan Meng, Eric A. Oches, Ivo Rappsilber, Stefan Wansa, Ludwig Zöller: Geologie, Paläontologie und Geochronologie des Eem-Beckens Neumark-Nord 2 und Vergleich mit dem Becken Neumark-Nord 1 (Geiseltal, Sachsen-Anhalt). In: Eiszeitalter und Gegenwart (Quaternary Science Journal). 59 (1/2), 2010, S. 120–167.
- Mark J. Sier, Wil Roebroeks, Corrie C. Bakels, Mark J. Dekkers, Enrico Brühl, Dimitri De Loecker, Sabine Gaudzinski-Windheuser, Norbert Hesse, Adam Jagich, Lutz Kindler, Wim J. Kuijper, Thomas Laurat, Herman J. Mücher, Kirsty E. H. Penkman, Daniel Richter, Douwe J.J. van Hinsbergen: Direct terrestrial–marine correlation demonstrates surprisingly late onset of the last interglacial in central Europe. In: Quaternary Research. 75, 2011, S. 213–218.
- Daniel Richter, Matthias Krbetschek: Preliminary luminiscense dating results for two Middle Palaeolithic occupations at Neumark-Nord 2. In: Sabine Gaudzinski-Windheuser, Wil Roebroeks (Hrsg.): Multidisciplinary studies of the Middle Palaeolithic record from Neumark-Nord (Germany). (Veröffentlichungen des Landesmuseums für Vorgeschichte, 69), Halle/Saale 2014, S. 131–136.
- Wolf-Dieter Heinrich: Kleinsäugerreste aus den pleistozänen Beckenablagerungen von Neumark-Nord 1. In: Dietrich Mania u. a. (Hrsg.): Neumark-Nord: Ein interglaziales Ökosystem des mittelpaläolithischen Menschen. (Veröffentlichungen des Landesmuseums für Vorgeschichte, 62). Halle/Saale 2010, S. 327–337.
- Dieter-Hans Mai, Jeanette Hoffmann: Die Vegetation von Neumark-Nord - Eine Rekonstruktion anhand karpologischer Reste. In: H. Meller (Hrsg.): Elefantenreich - Eine Fossilwelt in Europa. Halle/Saale 2010, S. 140–149.
- Roland Fuhrmann: Die Ostrakodenfauna der Interglazialbecken von Neumark-Nord (Geiseltal, Sachsen-Anhalt) und ihre Aussage zur stratigraphischen Stellung. In: Mauritiana. 32, 2017, S. 40–105 ().
- Roland Fuhrmann: Warthe-Kaltzeit oder Warthe-Stadium – zur stratigraphischen Gliederung des jüngeren Quartärs. In: Mauritiana. 22, 2011, S. 77–93 ((PDF)).
- Dietrich Mania, Maria Seifert, Matthias Thomae: Spät- und Postglazial im Geiseltal (mittleres Elb-Saalegebiet). In: Eiszeitalter und Gegenwart. 43, 1993, S. 1–22.
- Tatjana Böttger, Achim Hiller, Frank Wolfgang Junge, Thomas Litt, Dietrich Mania, Norbert Scheele: Late glacial stable isotope record, radiocarbon stratigraphy, pollen and mollusc analyses from the Geiseltal area, Central Germany. In: Boreas. 27, 1998, S. 88–100.
- Hartmut Haubold: Wirbeltiergrabung- und forschung im Geiseltaleozän. In: Hallesches Jahrbuch für Geowissenschaften. B 17, 1995, S. 1–18.
- Hartmund Haubold: Die Referenzfauna des Geiseltalium, MP Levels 11 bis 13 (Mitteleozän, Lutetium). In: Palaeovertebrata. 19 (3), 1989, S. 81–93.
- Meinolf Hellmund: Letzte Grabungsaktivitäten im südwestlichen Geiseltal bei Halle (Sachsen-Anhalt, Deutschland) in den Jahren 1992 und 1993. In: Hercynia. N. F. 30, 1997, S. 163–176.
- Hans Gallwitz: Kalk, Kieselsäure und Schwefeleisen in der Braunkohle des Geiseltales und ihre Bedeutung für die Fossilisation. In: Paläontologische Zeitschrift. 29 (1/2), 1955, S. 33–37.
- Ehrhard Voigt: Die Übertragung fossiler Wirbeltierleichen auf Zellulose-Filme, eine neue Bergungsmethode für Wirbeltiere aus der Braunkohle. In: Paläontologische Zeitschrift. 15, 1933, S. 72–78.
- Günter Krumbiegel: Die Fossilfundstellen der mitteleozänen Braunkohle des Geiseltales. In: Wissenschaftliche Zeitschrift der Martin-Luther-Universität Halle-Wittenberg, Mathematisch-Naturwissenschaftliche Reihe. 11 (6), 1962, S. 745–762.
- Krumbiegel, Rüffle, Haubold 1983, S. 25–34.
- Johannes Weigelt: Die Geiseltalgrabungen des Jahres 1933 und die Biostratonomie der Fundschichten. In: Nova Acta Leopoldina. N F 1 (4/5), 1934, S. 552–600, hier: S. 578.
- Ehrhard Voigt: Die Erhaltung von Epithelzellen mit Zellkernen, von Chromatophoren und Corium in fossiler Froschhaut aus der mitteleozänen Braunkohle des Geiseltales. In: Nova Acta Leopoldina. N F 3 (14), 1935, S. 339–360.
- Wilfried Krutzsch: Die Mikroflora des Geiseltales. IV: Die stratigraphische Stellung des Geiseltalprofils im Eozän und die sporenstratigraphische Untergliederung des mittleren Eozän. In: Abhandlungen des Zentralen Geologischen Institutes, Paläontologische Abhandlungen. 26, 1976, S. 47–92, hier: S. 47 und 51–57.
- Volker Wilde: Ein Farnhorizont aus dem Mitteleozän des Geiseltales (Sachsen-Anhalt, Deutschland). In: Hallesches Jahrbuch für Geowissenschaften. BH 13, 2001, S. 69–75.
- Victoria E. McCoy, Arnoud Boom, Oliver Wings, Torsten Wappler, Conrad C. Labandeira, Carole T. Gee: Fossilization of the Eocene „monkeyhair“ laticifer tree from Geiseltal, Germany: A deeper undertsanding using micro-CT and pyrolysis GC/MS. In: Palaios. 36 (1), 2021, S. 1–14, doi:10.2110/palo.2020.052.
- Volker Wilde: Die Makroflora aus dem Mitteleozän des Geiseltalgebietes, kurze Übersicht und Vergleiche. In: Hallesches Jahrbuch für Geowissenschaften. BH 17, 1995, S. 121–138.
- E. Kahlert, L. Rüffle: Monokotylen in der mitteleozänen Braunkohle des Geiseltales bei Merseburg (Sachsen-Anhalt) und ihre ökologische Bedeutung. In: Feddes Repertorium. 119 (3–4), 2008, S. 217–224.
- W. Salzmann: Das Braunkohlenvorkommen im Geiseltal mit besonderer Berücksichtigung der Genesis. In: Archiv für Lagerstättenforschung. 17, 1914, S. 1–105, hier: S. 31–33.
- Günter Krumbiegel: Molluskenfunde (Gastropoda der Ordnung Pulmonata CUVIER 1795 in der mitteleozänen Braunkohle des Geiseltales und ihre fazielle und stratigraphische Bedeutung). In: Geologie. 11 (5), 1962, S. 557–603.
- Günter Krumbiegel: Ostracodenfunde im Tagebau Neumark-Süd (Geiseltal) In: Geologie. 11 (3), 1962, S. 334–353.
- Karl Beurlen: Crustaceenreste aus der Geiseltalbraunkohle. In: Nova Acta Leopoldina. N F 5, 1938, S. 361–368.
- Francesco Vitali: Systematic revision of the fossil cerambycids from Geiseltal (Coleoptera Cerambycidae). In: Entomapeiron Paleoentomology. 3 (1), 2008, S. 1–16.
- Thomas Hörneschmeyer, Gert Tröster, Sonja Weidmann: Die eozänen Käferfaunen des Geiseltales und der Grube Messel – ein Vergleich unter systematischen und paläoökologischen Gesichtspunkten. In: Hallesches Jahrbuch für Geowissenschaften. B 17, 1995, S. 107–119.
- Meinolf Hellmund, Winfried Hellmund: Erster Nachweis von Kleinlibellen-Eilogen (Insecta, Zygoptera, Lestidae) in der mitteleozänen Braunkohle des ehemaligen Tagebaus Mücheln, Baufeld Neumark-Nord (Geiseltal, Sachsen-Anhalt, Deutschland). In: Hallesches Jahrbuch für Geowissenschaften. BH 24, 2002, S. 47–55.
- Meinolf Hellmund, Winfried Hellmund: Neufunde und Ergänzungen zur Fortpflanzungsbiologie fossiler Kleinlibellen (Insecta, Odonata, Zygoptera). In: Stuttgarter Beiträge zur Naturkunde. Serie B 319, 2002, S. 1–26.
- Volker Wilde, Meinolf Hellmund: Neue Geländearbeiten im ehemaligen Braunkohlerevier Geiseltal. In: Natur und Museum. 136 (7/8), 2006, S. 162–173.
- Hans Pohl: The oldest fossil strepsipteran larva (Insecta: Strepsiptera) from the Geisel Valley, Germany (Eocene). In: Insect Systematics & Evolution. 40, 2009, S. 333–347.
- Ehrhard Voigt: Ein fossiler Saitenwurm (Gordius tenuifibrosus n. sp.) aus der eozänen Braunkohle des Geiseltales. In: Nova Acta Leopoldina. N F 5, 1938, S. 352–360.
- Krumbiegel, Rüffle, Haubold 1983, S. 53–90.
- Jean Gaudant: An attempt at the palaeontological history of the European mudminnows (Pisces, Teleostei, Umbridae). In: Neues Jahrbuch für Geologie und Paläontologie Abhandlungen. 263/2, 2012, S. 93–109.
- Norbert Micklich, Jean Gaudant: Anthracoperca siebergi VOIGT 1934 (Pisces, Perciformes) – Ergebnisse einer Neuuntersuchung der mitteleozänen Barsche des Geiseltales. In: Zeitschrift für geologische Wissenschaften. 17 (5), 1989, S. 503–521.
- Krumbiegel, Rüffle, Haubold 1983, S. 114–118.
- Michael Wuttke, Tomáš Přikryl, Viacheslav Yu. Ratnikov und Zdeněk Dvořák: Generic diversity and distributional dynamics of the Palaeobatrachidae (Amphibia: Anura). In: Palaeobiology Palaeoenvironment. 92, 2012, S. 367–395.
- Krumbiegel, Rüffle, Haubold 1983, S. 119–124.
- Stefanie Zimmermann-Rollius: Beiträge zur Schildkrötenfauna der mitteleozänen Braunkohle des Geiseltales. In: Hercynia. N F 4, 1967, S. 83–104.
- Hans-Volker Karl: Die Schildkröten aus den Mitteleozänen Sedimenten des Eckfelder Maares mit taxonomischen Notizen zu Testudo eocaenica Hummel, 1935 (Mitteleozän, Deutschland, Rheinland-Pfalz, Vulkaneifel). In: Mainzer naturwissenschaftliches Archiv. 55, 2018, S. 61–95.
- Sophie Hervet: A new genus of ‘Ptychogasteridae’ (Chelonii, Testudinoidea) from the Geiseltal (Lutetian of Germany). In: Comptes Rendus Palevol. 3, 2004, S. 125–132.
- Günter Krumbiegel: Trionychidenfunde (Flußschildkröten) aus der Gattung Trionyx GEOFFROY 1809 aus der eozänen Braunkohle des Geiseltales. In: Geologie. 12, 1963, S. 196–223.
- Edwin Cadena: Palaeoamyda messeliana nov. comb. (Testudines, Pan-Trionychidae) from the Eocene Messel Pit and Geiseltal localities, Germany, taxonomic and phylogenetic insights. In: PeerJ. 4, 2016, S. e2647 doi:10.7717/peerj.2647.
- Johannes Müller: Osteology and relationships of Eolacerta robusta, a lizard from the Middle Eocene of Germany (Reptilia, Squamata). In: Journal of Vertebrate Paleontology. 21 (2), 2001, S. 261–278.
- Andrej Čerňanský, Krister T. Smith: Eolacertidae: a new extinct clade of lizards from the Palaeogene; with comments on the origin of the dominant European reptile group – Lacertidae. In: Historical Biology. 30 (7), 2018, S. 994–1014.
- Krister T. Smith: Eocene Lizards of the Clade Geiseltaliellus from Messel and Geiseltal, Germany, and the Early Radiation of Iguanidae (Reptilia: Squamata). In: Bulletin of the Peabody Museum of Natural History. 50 (2), 2009, S. 219–306, hier: S. 256–257.
- Robert M. Sullivan, Thomas Keller, Jörg Habersetzer: Middle Eocene (Geiseltalian) anguid lizards from Geiseltal and Messel, Germany. I. Ophisauriscus quadrupes KUHN 1940. In: Courier Forschungsinstitut Senckenberg. 216, 1999, S. 97–129.
- Olivier Rieppel, Jack L. Conrad, Jessica A. Maisano: New morphological data for Eosaniwa koehni and revised phylogenetic analysis. In: Journal of Paleontology. 81 (4), 2007, S. 760–769.
- Johannes Weigelt: Die Wirbeltierausgrabungen im Geiseltal. In: Die Naturwissenschaften. 21, 1934, S. 321–325.
- Oskar Kuhn: Die Schlangen (Boidae) aus dem Mitteleozän des Geiseltales. In: Nova Acta Leopoldina. N F 7, 1939, S. 119–133.
- Krumbiegel, Rüffle, Haubold 1983, S. 142–145.
- Alexander K. Hastings und Meinolf Hellmund: Rare in situ preservation of adult crocodylian with eggs from the Middle Eocene of Geiseltal, Germany. In: Palaios. 30 (6), 2015, S. 446–461.
- Christopher A. Brochu: Phylogenetic relationships of Palaeogene ziphodont eusuchians and the status of Pristichampsus Gervais, 1853. In: Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 103, 2013, S. 521–550, hier: 524–527.
- Torsten Rossmann, Michael Rauhe, Francesco Ortega: Studies on Cenozoic crocodiles: 8. Bergisuchus dietrichbergi KUHN (Sebecosuchia: Bergisuchidae n. fam.) from the Middle Eocene of Germany, some new systematic and biological conclusions. In: Paläontologische Zeitschrift. 74 (3), 2000, S. 379–392.
- Michael Rauhe: Die Lebensweise und Ökologie der Geiseltal-Krokodilier – Abschied von traditionellen Lehrmeinungen. In: Hallesches Jahrbuch für Geowissenschaften. B 17 1995, S. 65–80.
- Alexander K. Hastings, Meinolf Hellmund: Evidence for prey preference partitioning in the middle Eocene high-diversity crocodylian assemblage of the Geiseltal-Fossillagerstätte, Germany utilizing skull shape analysis. In: Geological Magazine. 154 (1), S. 119–146 doi:10.1017/S0016756815001041.
- Rolf Kohring: Ultrastrukturen und Erhaltungsbedingungen fossiler Krokodil-Eierschalen aus dem Geiseltal (Mitteleozän). In: Hallesches Jahrbuch für Geowissenschaften. B 17, 1995, S. 93–105.
- Kálmán Lambrecht: Palaeotis Weigelti n. g. n. sp., eine fossile Trappe aus der mitteleozänen Braunkohle des Geiseltales. In: Jahrbuch des Halleschen Verbandes. N F 7, 1928, S. 20–29.
- Peter Houde, Hartmut Haubold: Palaeotis weigelti restudied: a small Middle Eocene ostrich (Aves: Struthioniformes). In: Palaeovertebrata. 17, 1987, S. 27–42.
- Gerald Mayr: Hindlimb morphology of Palaeotissuggests palaeognathous affinities of the Geranoididae and other “crane-like” birds from the Eocene of the Northern Hemisphere. In: Acta Palaeontologica Polonica. 64 (4), 2019, S. 669–678, doi:10.4202/app.00650.2019.
- Karl-Heinz Fischer: Der Riesenlaufvogel Diatryma aus der eozänen Braunkohle des Geiseltales. In: Hallesches Jahrbuch für Mitteldeutsche Erdgeschichte. 4, 1962, S. 26–33.
- Meinolf Hellmund: Reappraisal of the bone inventory of Gastornis geiselensis (Fischer, 1978) from the Eocene “Geiseltal Fossillagerstätte” (Saxony-Anhalt, Germany). In: Neues Jahrbuch für Geologie und Paläontologie Abhandlungen. 269 (2), 2013, S. 203–220.
- Max-Planck-Gesellschaft: Harmloser Terrorvogel. vom 30. August 2013, in: Forschung/Aktuelles, Webangebot der Max-Planck-Gesellschaft (), abgerufen 11. Dezember 2013.
- Alexander K. Hastings, Meinolf Hellmund: Aus der Morgendämmerung: Pferdejagende Krokodile und Riesenvögel. Neueste Forschungsergebnisse zur eozänen Welt Deutschlands vor ca. 45 Millionen Jahren. Halle (Saale), 2015, S. 1–120, hier: S. 66–75.
- Kálmán Lambrecht: Drei neue Vogelformen aus dem Lutétium des Geiseltales. In: Nova Acta Leopoldina. 3, 1935, S. 361–367.
- Gerald Mayr: Synonymy and actual affinities of the putative Middle Eocene „New World vulture“ Eocathartes LAMBRECHT, 1935 and „hornbill“ Geiseloceros LAMBRECHT, 1935 (Aves, Ameghinornithidae). In: Paläontologische Zeitschrift. 81 (4), 2007, S. 457–462.
- Dennis Stefan Peters: Erstnachweis eines Seglers aus dem Geiseltal (Aves: Apodiformes). In: Senckenbergiana lethaea. 76 (1/2), 1998, S. 211–212.
- Florian Heller: Neue Beuteltierreste aus der mitteleozänen Braunkohle des Geiseltales bei Halle (Saale). In: Nova Acta Leopoldina. N F 4, 1936, S. 311–316.
- Gerhard Storch, Hartmut Haubold: Additionals to the Geiseltal Mammalian faunas, Middle Eocene: Didelphidae, Nyctitheriidae. Myrmecophagidae. In: Palaeovertebrata. 19 (3), 1989, S. 95–114.
- Gerhard Storch: Eurotamandua joresi, ein Myrmecophagide aus dem Eozän der „Grube Messel“ bei Darmstadt (Mammalia, Xenarthra). In: Senckenbergiana lethaea. 61 (3/6), 1981, S. 247–289, hier: S. 253.
- Timothy J. Gaudin, Robert J. Emry, John R. Wible: The Phylogeny of Living and Extinct Pangolins (Mammalia, Pholidota) and Associated Taxa: A Morphology Based Analysis. In: Journal of Mammal Evolution. 16, 2009, S. 235–305, hier: S. 241 und 274.
- Florian Heller: Die Säugetierfauna der Mitteleozänen Braunkohle des Geiseltales bei Halle a. S. In: Jahrbuch des Halleschen Verbandes für die Erforschung der Mitteldeutschen Bodenschätze und ihrer Verwertung. 9, 1930, S. 13–41.
- Jerry J. Hooker: Origin and evolution of the Pseudorhyncocyonidae, a European Paleogene famaly of insectivorous placental mammals. In: Palaeontology. 56 (4), 2013, S. 807–835, hier: S. 818–820.
- Wighart von Koenigswald: Der erste Pantolestide (Proteutheria, Mammalia) aus dem Eozän des Geiseltales bei Halle. In: Zeitschrift für Geologische Wissenschaften. 11, 1983, S. 781–787.
- Horst Werner Matthes: Die Creodontier aus der mitteleozänen Braunkohle des Geiseltales. In: Hallesches Jahrbuch für Mitteldeutsche Erdgeschichte. 1, 1952, S. 201–240.
- Leigh Van Valen: Paroxyclaenidae, an Extinct Family of Eurasian Mammals. In: Journal of Mammalogy. 46 (3), 1965, S. 388–397.
- Krumbiegel, Rüffle, Haubold 1983, S. 160–165.
- Floréal Solé, Olivia Plateau, Kévin Le Verger, Alain Phélizon: New paroxyclaenid mammals from the early Eocene of the Paris Basin (France) shed light on the origin and evolution of these endemic European cimolestans. In: Journal of Systematic Palaeontology., 2019 doi:10.1080/14772019.2018.1551248.
- Bernhard Sigé, Gerhard Storch: Un nouveau Saturninia (Nyctitheriidae, Lipotyphla, Mammalia) de l’assise OK (Oberkohle, MP 14) du bassin lignitifère du Geiseltal (Eocène moyen supérieur d’Allemagne). In: Senckenbergiana lethaea. 81 (2), 2001, S. 343–346.
- Gerhard Storch: Kleinsäugetiere aus dem Geiseltal und Messel im Kontext alttertiärer Faunenentfaltungen. In: Hallesches Jahrbuch für Geowissenschaften. B 17, 1995, S. 59–64.
- Urs Thalmann, Hartmut Haubold, Robert Denis Martin: Pronycticebus neglectus – An almost complete adapid primate specimen from the Geiseltal (GDR). In: Palaeovertebrata. 19 (3), 1989, S. 115–130.
- Jens Lorenz Franzen: Der sechste Messel-Primate (Mammalia, Primates, Notharctidae, Cercamoniinae). In: Senckenbergiana lethaea. 80 (1), 2000, S. 289–303.
- Jens Lorenz Franzen, Philip D. Gingerich, Jörg Habersetzer, Jørn H. Hurum, Wighart von Koenigswald und B. Holly Smith: Complete Primate Skeleton from the Middle Eocene of Messel in Germany: Morphology and Paleobiology. In: PLoS ONE. 4 (5), 2009, S. e5723 doi:10.1371/journal.pone.0005723.
- Horst Werner Matthes: Ein neuer Creodontier: Prodissopsalis theriodis van Valen 1965 aus der eozänen Braunkohle des Geiseltales. In: Hallesches Jahrbuch für Mitteldeutsche Erdgeschichte. 8, 1967, S. 7–11.
- Brigitte Lange-Badré, Hellmund Haubold: Les créodontes (Mammifères) du gisement du Geiseltal (Eocène Moyen, RDA). In: Geobios. 23 (5), 1990, S. 607–637.
- Paul David Polly, Brigitte Lange-Badré: A new genus Eurotherium (Mammalia, Creodonta) in reference to taxonomic problems with some Eocene hyaenodontids of Eurasia. In: Comptes Rendus de l’Académie des Sciences Paris. Série 2 317, 1993, S. 991–996.
- Leigh Van Valen: Some Europaea Proviverrini (Mammalia, Deltatheridia). In: Palaeontology. 8 (4), 1965, S. 638–665, hier: S. 653–658.
- Krumbiegel, Rüffle, Haubold 1983, S. 165–172.
- Jörg Erfurt, Jean Sudre: Eurodexeinae, eine neue Unterfamilie der Artiodactyla (Mammalia) aus dem Unter- und Mitteleozän Europas. In: Palaeovertebrata. 25 (2–4), 1996, S. 371–390.
- Jörg Erfurt, Hartmut Haubold: Artiodactyla aus den eozänen Braunkohlen des Geiseltales bei Halle (DDR). In: Palaeovertebrata. 19 (1), 1989, S. 131–160, besonders S. 152–155.
- Jörg Erfurt, Jean Sudre: Un Haplobunodontidae nouveau, Hallebune krumbiegeli nov. gen. nov. sp. (Artiodactyla, Mammalia) dans L’Eocene Moyen du Geiseltal pres Halle (Sachsen-Anhalt, Allemagne). In: Palaeovertebrata. 24 (1–2), 1995, S. 84–99.
- Jerry J. Hooker, Katherine M. Thomas: New species of Amphirhagatherium (Choeropotamidae, Artiodactyla, Mammalia) from the late Eocene Headon Hill Formation of Southern England and phylogeny of endemic European Anthracotheroids. In: Palaeontology. 44(5), 2001, S. 827–853.
- Jörg Erfurt: Rekonstruktion des Skelettes und der Biologie von Anthracobunodon weigelti (Artiodactyla, Mammalia) aus dem Eozän des Geiseltales. In: Hallesches Jahrbuch für Geowissenschaften. Reihe B, Beiheft 12, 2000, S. 57–141, hier: S. 59 und 107.
- Jörg Erfurt, Hans Altner: Habitus-Rekonstruktion von Anthracobunodon weigelti (Artiodactyla, Mammalia) aus dem Eozän des Geiseltales. In: Jan Michal Burdukiewicz, Lutz Fiedler, Wolf-Dieter Heinrich, Antje Justus, Enrico Brühl (Hrsg.): Erkenntnisjäger. Festschrift für Dietrich Mania. (Veröffentlichungen des Landesmuseums für Vorgeschichte in Halle, 57). Halle/Saale 2003, S. 153–176.
- Jörg Erfurt: Taxonomie der eozänen Artiodactyla (Mammalia) des Geiseltales mit besonderer Berücksichtigung der Gattung Rhagatherium. In: Hallesches Jahrbuch für Geowissenschaften. B 17, 1995, S. 47–58.
- Horst Werner Matthes: Weitere eozäne Equiden aus dem Geiseltal. In: Zeitschrift für Geologische Wissenschaften. 7 (6), 1979, S. 783–805.
- Jens Lorenz Franzen, Hartmut Haubold: Revision der Equoidea aus den eozänen Braunkohlen des Geiseltales bei Halle (DDR). In: Palaeovertebrata. 16 (1), 1986, S. 1–34.
- Jens Lorenz Franzen: Eurohippus n. g., a new genus of horses from the Middle to Late Eocene of Europe. In: Senckenbergiana lethaea. 86 (1), 2006, S. 97–102.
- Meinolf Hellmund: Erstnachweis von Plagiolophus cartieri Stehlin (Palaeotheriidae, Perissodactyla) in der Unteren Mittelkohle (uMK, MP 12) des Geiseltales bei Halle (Sachsen-Anhalt, Deutschland). In: Neues Jahrbuch für Geologie und Paläontologie. 2000 (4), S. 205–216.
- Karl-Heinz-Fischer: Die tapiroiden Perissodactylen aus der eozänen Braunkohle des Geiseltales. In: Geologie. 45, 1964, S. 1–101, hier: S. 71.
- Karl-Heinz-Fischer: Neue Funde von Rhinocerolophiodon (n. gen.), Lophiodon, und Hyrachyus (Ceratomorpha, Perissodactyla, Mammalia) aus dem Eozän des Geiseltals bei Halle (DDR). 1. Teil: Rhinocerolophiodon. In: Zeitschrift für geologische Wissenschaft. 5, 1977, S. 909–919.
- Karl-Heinz-Fischer: Neue Funde von Rhinocerolophiodon (n. gen.), Lophiodon, und Hyrachyus (Ceratomorpha, Perissodactyla, Mammalia) aus dem Eozän des Geiseltals bei Halle (DDR). 2. Teil: Lophiodon. In: Zeitschrift für geologische Wissenschaft. 5, 1977, S. 1129–1152.
- Karl-Heinz-Fischer: Zur systematischen Stellung von Chasmotherium RÜTIMEYER 1862 (Mammalia, Perissodactyla). In: Berichte der deutschen Gesellschaft für geologische Wissenschaften. 12A (5), 1967, S. 595–600.
- Kerstin Hlawatsch, Jörg Erfurt: Zahnmorphologie und stratigraphische Verbreitung von Hyrachyus minimus (Perissodactyla, Mammalia) in den eozänen Geiseltalschichten. In: Jörg Erfurt, Lutz Christian Maul (Hrsg.): 34. Tagung des Arbeitskreises für Wirbeltierpaläontologie der Paläontologischen Gesellschaft 16. bis 18. März 2007 in Freyburg/Unstrut. (Hallesches Jahrbuch für Geowissenschaften, BH 23), 2007, S. 161–173.
- Erhard Voigt: Weichteile an Fischen, Amphibien und Reptilien aus der eozänen Braunkohle des Geiseltales. In: Nova Acta Leopoldina. N F 5, 1937, S. 115–142.
- Adolf Brachofen-Echt: Das Vorkommen von Federn in der eozänen Braunkohle des Geiseltales. In: Nova Acta Leopoldina. N F 4, 1936, S. 335–340.
- Erhard Voigt: Über das Haarkleid einiger Säugetieren aus der eozänen Braunkohle des Geiseltales. In: Nova Acta Leopoldina. N F 4, 1936, S. 317–334.
- Erhard Voigt: Weichteile an Säugetieren aus der eozänen Braunkohle des Geiseltales. In: Nova Acta Leopoldina. N F 4, 1936, S. 301–310.
- Krumbiegel, Rüffle, Haubold 1983, S. 189–193.
- Angelika Otto, Bernd R. T. Simoneit, Volker Wilde: Initial results on the biomarker composition and „Affenhaar“ from Middle Eocene lignites of the Geiseltal (Sachsen-Anhalt, Germany). In: Hallesches Jahrbuch für Geowissenschaften. BH 13, 2001, S. 57–68.
- Johannes Weigelt: Lophiodon in der oberen Kohle des Geiseltales. In: Nova Acta Leopoldina. N F 3 (14), 1935, S. 369–402, hier: S. 376 und 384–385.
- Volker Wilde, Meinolf Hellmund: First record of gut contents from a middle Eocene equid from the Geiseltal near Halle (Saale), Sachsen-Anhalt, Central Germany. In: Palaeobiodiversity and Palaeoenvironments. 90, 2010, S. 153–162.
- Daniel Falk, Victoria McCoy, Oliver Wings: Ichnofossils of the Eocene Fossillagerstätte Geiseltal (Saxony-Anhalt, Germany). In: Michael Buchwitz, Daniel Falk, Hendrik Klein, Dorothee Mertmann, Arila Perl, Oliver Wings (Hrsg.): 3rd International Conference of Continental Ichnology. Halle (Saale), Germany; 23rd - 29th September. Abstract Volume & Field Trip Guide. (Hallesches Jahrbuch für Geowissenschaften BH 46), 2019, S. 15–18.
- Henry Schroeder: Das Vorkommen der Gattung Lophiodon in der Braunkohle Sachsens. In: Centralblatt für Mineralogie, Geologie und Paläontologie. 11, 1913, S. 351.
- Jens Lorenz Franzen: Warum Geiseltalium? In: Courier des Forschungsinstitutes Senckenberg. 255, 2005, S. 77–79.
- Dieter F. Mertz, Paul R. Renne: A numerical age for the Messel fossil deposit (UNESCO World Heritage Site) derived from 40Ar/39Ardating on a basaltic rock fragment. In: Courier des Forschungsinstitutes Senckenberg. 255, 2005, S. 67–75.
- Dieter F. Mertz, Carl C. Swisher III, Jens Lorenz Franzen, Franz-Otto Neuffer, Herbert Lutz: Numerical dating of the Eckfeld maar fossil site, Eifel, Germany: calibration mark for the Eocene time scale. In: Naturwissenschaften. 8, 2000, S. 270–274.
- Horst Blumenstengel: Palynologische Untersuchungen tertiärer Ablagerungen aus dem ehemaligen Braunkohletagebau Mücheln-Westfeld (Geiseltal, Sachsen-Anhalt, Deutschland). In: Hallesches Jahrbuch für Geowissenschaften. BH 13, 2001, S. 31–39.
- Krumbiegel, Rüffle, Haubold 1983, S. 41–43.
- Krumbiegel, Rüffle, Haubold 1983, S. 34–37.
- Simon J. Ring, Hervé Bocherens, Oliver Wings, Márton Rab: Divergent mammalian body size in a stable Eocene greenhouse climate. In: Scientific Reports. 10, 2020, S. 3987, doi:10.1038/s41598-020-60379-7.
- Johannes Weigelt: Die Aufdeckung der bisher ältesten tertiären Säugetierfauna Deutschlands. In: Nova Acta Leopoldina. N F 7, 1939, S. 515–528.
- Kenneth D. Rose, Gerhard Storch, Katrin Krohmann: Small-mammal postcrania from the middle Paleocene of Walbeck, Germany. In: Paläontologische Zeitschrift. 89 (1), 2013, S. 95–124 doi:10.1007/s12542-013-0211-3.
- Kenneth D. Rose: The importance of Messel for interpreting Eocene Holarctic mammalian faunas. In: Palaeobiology Palaeoenvironment. 92, 2012, S. 631–647 doi:10.1007/s12549-012-0090-8.
- Herbert Frankenhäuser, Werner Löhnertz, Jens L. Franzen, Uwe Kaufluss, Martin Koziol Herbert Lutz, Dieter F. Mertz, Jens Mingram, Torsten Wappler, Volker Wilde: Das Eckfelder Maar in der Vulkaneifel – Fenster in einen küstenfernen Lebensraum vor 44 Millionen Jahren. In: Mainzer Naturwissenschaftliches Archiv. 47, 2009, S. 263–324, hier: S. 301–303.
- LMBV: Geiseltal. Mitteldeutsches Braunkohlenrevier 3. Senftenberg, 2009.
- Ben Barnes: Eine eozäne Wirbeltier-Fauna aus der Braunkohle des Geiseltales. In: Jahrbuch des Halleschen Verbandes für die Erforschung der mitteldeutschen Bodenschätze. 6, 1927, S. 5–24.
- Meinolf Hellmund: The Former Geiseltal Museum (1934-2011), the Eocene Geiseltal Fossilagerstätte (Germany) and the Scientific Meaning of Ben Barnes as a Pioneer of Systematic Quantitative Vertebrate Excavations in the Geiseltal Lignites. In: Anuário do Instituto de Geociências - UFRJ. 41 (1), 2018, S. 108–119.
- Hartmut Haubold: 60 Jahre Geiseltalmuseum an der Martin-Luther-Universität Halle-Wittenberg. In: Hallesches Jahrbuch für Geowissenschaften. B 17, 1995, S. 19–25.
- Martin-Luther-Universität Halle-Wittenberg: Geiseltalmuseum schließt – Fossilien ab 2015 im Naturkundlichen Universitätsmuseum. Pressemitteilung der Pressestelle der Martin-Luther-Universität Halle-Wittenberg, Nummer 237/2011 vom 21. Oktober 2011 (), zuletzt abgerufen am 25. März 2018.
- Friederike Stecklum: Alte Fossilien neu entdeckt. In: Scientia halensis. 1/2018, S. 32–34 ().
- Leopoldina: Aus der Morgendämmerung: Pferdejagende Krokodile und Riesenvögel. Ausstellung der Martin-Luther-Universität Halle-Wittenberg an der Leopoldina März bis Mai 2015 (), zuletzt abgerufen am 25. März 2018.
- Landesmuseum für Vorgeschichte Halle: Klimagewalten – Treibende Kraft der Evolution. Sonderausstellung des Landesmuseums für Vorgeschichte Halle November 2017 bis Mai 2018 (), zuletzt abgerufen 25. März 2018.
- Kulturgutschutz Deutschland: National wertvolles Kulturgut. Länderverzeichnisse national wertvollen Kulturgutes der Länder. Datenbank (), Nr. 14805, zuletzt abgerufen am 11. April 2019.
Weblinks
- Geiseltalsammlung an der Martin-Luther Universität Halle-Wittenberg
- Sonderausstellung: Aus der Morgendämmerung: Pferdejagende Krokodile und Riesenvögel