Heterohyus
Heterohyus ist eine ausgestorbene Gattung insektenfresserartiger Säugetiere, die vom Unteren bis zum Oberen Eozän vor 51 bis 34 Millionen Jahren im heutigen Europa verbreitet war. Es handelt sich um kleine Tiere mit charakteristisch vergrößerten Schneidezähnen und zwei verlängerten Fingern. Aufgrund der Merkmalskombination bestehen Ähnlichkeiten zum heutigen Fingertier von Madagaskar und den Streifenbeutlern von Neuguinea. Wie diese lebte Heterohyus wahrscheinlich in Bäumen und benagte mit den Vorderzähnen Rinde, während die langen Finger in Spalten und Hohlräumen nach Insekten bohrten. Fossilfunde von Heterohyus wurden in den zentralen und westlichen Bereichen des Kontinents gefunden. In der Regel liegen nur Schädel- und Gebissteile vor, einige gut erhaltene Skelette sind aus der Grube Messel belegt. Die Gattung erhielt bereits Mitte des 19. Jahrhunderts ihre wissenschaftliche Erstbeschreibung, ihre genaue systematische Stellung blieb aber lange Zeit ungeklärt. Heute wird sie zu den Apatotheria gestellt, die möglicherweise näher mit den Nagern und den Euarchonta verwandt sind. Innerhalb der Gattung werden mehrere Arten unterschieden.
| Heterohyus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelett von Heterohyus | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Eozän | ||||||||||||
| 50,7 bis 33,9 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Heterohyus | ||||||||||||
| Gervais, 1848 | ||||||||||||
Beschreibung
Heterohyus war ein kleiner Vertreter der Säugetiere. Einzelne vollständige Skelette von kleinen Angehörigen der Gattung weisen eine Kopf-Rumpf-Länge von 11,5 bis 13,6 cm und eine Schwanzlänge von 15 bis 17,4 cm auf. In der Körpergröße entsprechen diese etwa dem heutigen Siebenschläfer. Unter Verweis auf vorliegendes Schädelmaterial dürften die größten Formen von Heterohyus etwa fünfmal so groß gewesen sein.[1][2][3] Der Schädel selbst wurde bei den kleinen Heterohyus-Arten etwa 38 mm lang. Im Vergleich mit dem recht zarten Skelettbau war er massiv gestaltet. Durch den hohen und kurzen Mittelkieferknochen wirkte er außerdem eher stumpf. Die Orbita lag mit ihrem Vorderrand oberhalb des ersten Molaren, sie war nach hinten offen und besaß daher nur einen schwach ausgebildeten Processus supraorbitalis. Das Foramen infraorbitale befand sich im Vergleich zum vorderen Orbitarand etwas zurücksetzt oberhalb des ersten und zweiten Mahlzahns. Eine ähnliche Position nahm die Voraugengrube auch bei Apatemys, Carcinella und Sinclairella ein. Weitere zahlreiche Foramina öffneten sich im Schnauzenbereich, vor allem im Umfeld der Schneidezähne. Der Jochbogen war massiv und kräftig geschwungen. Unterhalb des vorderen Bogenansatzes saß eine kleine Grube, die auch bei anderen Apatemyiden wie Sinclairella vorkommt. Im hinteren Schädelbereich zeigten sich ebenfalls zahlreiche Foramina. Ähnliche Bildungen sind von Apatemys und Sinclairella berichtet worden. Auf dem Scheitelbein erhob sich ein kräftiger Scheitelkamm, wie er auch von Carcinella belegt ist. Bei Sinclairella hingegen kamen zwei parasagittale Leisten vor, während bei Stehlinella keines der beiden Merkmale auftrat.[1][2][4][5]
Der Unterkiefer wurde etwa 22 mm lang und am Kronenfortsatz 12,5 mm hoch. Die Symphyse im vorderen Bereich war nicht verwachsen. Nach vorn öffneten sich die beiden Unterkieferhälften in den Alveolen des massiven unteren Schneidezahns, dessen Zahnwurzel bis zum letzten Molaren reichte. Abweichend von den heutigen Nagetieren mit ebenfalls ktäftigen Schneidezähnen bestimmte dieser aber nicht die Kontur des Unterkiefers. Das Foramen mentale hatte seine Öffnung unterhalb des zweiten Molaren. Der aufsteigende Ast setzte vorn in einem Winkel von 130 bis 140 ° gegenüber der Kauebene an und war damit flacher als bei Sinclairella. Die Fossa masseterica zeichnete sich hier in ihrem vorderen Rand als scharf umgrenzt ab. Am hinteren Ende des Unterkiefers befand sich ein deutlicher Winkelfortsatz.[1][2]
Das Gebiss von Heteryus war in seiner Zahnanzahl reduziert, die Zahnformel lautete: . Gegenüber stammesgeschichtlich älteren Apatemyiden hatte sich die Zahl der Prämolaren verringert, es fehlen der erste und der dritte Vormahlzahn. Letzterer kam bei urtümlicheren Vertretern wie Labidolemur noch vor. Ein Eckzahn war bei den Apatemyiden generell nicht ausgebildet.[6][7][8] Die auffälligste Besonderheit im Gebiss von Heterohyus und anderer Apatemyiden findet sich in den massiven und großen Schneidezähnen. Die oberen beiden Zahnpaare riefen daher auch die hohe Gestaltung des Mittelkieferknochens hervor, der diese vollständig trug. Der innere obere wies eine halbkreisförmige Krümmung auf, er lief spitz zu und verfügte an der Zahnrückseite über einen kleinen zusätzlichen Höcker. Der äußere Schneidezahn war weniger stark gebogen. Im Unterkiefer stand der einzelne Schneidezahn schräg nach vorn (procumbent) und krümmte sich über seine gesamte Länge regelmäßig. Die oberen Schneidezähnen zeigten eine vollständige Bedeckung mit Zahnschmelz, der auf der Hinterseite aber deutlich ausdünnte, der untere Schneidezahn besaß nur auf der Vorderseite Zahnschmelz. Im oberen Gebiss trennte ein kurzes Diastema das vordere vom hinteren Gebiss. Zusätzliche Zahnlücken zwischen den Vormahlzähne, wie noch bei Labidolemur oder Apatemys zu beobachten, fehlten bei Heterohyus, so dass die hintere Zahnreihe geschlossen stand. Auffallend ist der vordere untere Prämolar, der stark vergrößert war sowie eine langgestreckte und seitlich verschmälerte Zahnkrone besaß, wodurch eine lange Schneidkante entstand. Trotz seiner Größe verfügte er nur über eine Wurzel. Der hintere untere Prämolar war dagegen klein und stiftartig, er erreichte nicht die Höhe der Kauebene. Grundsätzlich ragte seine Krone nicht über die des vorderen Prämolaren, wie es auch für Apatemys und Carcinella dokumentiert ist. Die oberen Mahlzähne zeichneten sich durch vier Haupthöcker aus (Para- und Metaconus auf der Wangen-, Proto- und Hypoconus auf der Zungenseite). Der Paraconus überragte den Metaconus an Höhe, neben diesen beiden Haupthöckern der Wangenseite erhob sich mit dem Parastyl noch ein kleinerer Nebenhöcker. In Bezug auf den Hypoconus auf der Zungenseite gab es Variationen innerhalb der Gattung Heterohyus, da einige Arten eine stärkere, andere eine schwächere Ausprägung zeigten. Ein Hypoconus war auch bei Apatemys, Labidolemur und Sinclairella vorhanden, fehlte jedoch bei Carcinella. Dadurch kann das vierhöckrige Kauflächenmuster der oberen Molaren als Zeichen für stammesgeschichtlich jüngere Formen bewertet werden. Die unteren Molaren wiesen ebenfalls vier Haupthöcker (Protoconid, Metaconid, Paraconid und Hypoconid) auf. Hier zeigten die einzelnen Vertreter stärkere Variationen bezüglich des Paraconids.[1][5][9]

Von der Wirbelsäule sind die ersten beiden Halswirbel, 13 Brust-, 6 Lenden- 2 Kreuzbein- und 34 bis 35 Schwanzwirbel überliefert. Der Schwanz war damit ausgesprochen lang und übertraf den restlichen Körper in seiner Ausdehnung. Die Größe der Schwanzwirbel nahm nach hinten kontinuierlich ab, so dass der Schwanz immer schlanker wurde. An den vordersten rund ein Dutzend befanden sich deutliche Querfortsätze. Die restlichen Schwanzwirbel wiesen eine zylindrische Form auf. Der Brustkorb bestand aus zwölf oder dreizehn Rippenpaaren. Die Knochen der vorderen Extremität waren relativ schlank und lang. Das Schulterblatt und das Brustbein verband ein relativ robustes Schlüsselbein. Der Oberarmknochen wurde rund 18 bis 21 mm lang und besaß keine auffallenden Muskelmarken, allerdings zog sich eine kräftige Aufrauhung über etwa ein Drittel des Knochenschaftes. Elle und Speiche waren nicht miteinander verwachsen. Der Radius übertraf den Humerus mit 21 mm nur geringfügig an Länge, die Ulna war mit 26 mm deutlich ausgedehnter. Letzterer Knochen besaß einen massigen oberen Gelenkfortsatz (Olecranon). Das Hinterbein war länger und kräftiger als das Vorderbein. Am Becken zeichnete sich eine lange Darmbeinschaufel ab. Der Oberschenkelknochen maß rund 27 mm in der Länge, er wies eine gerade und gestreckte Form auf. Das Schienbein wurde ähnlich lang. Möglicherweise war es an der Basis mit dem Wadenbein fusioniert. Die Schäfte der beiden Knochen standen aber vollständig frei voneinander.[1][2]
Die Hände und Füße waren jeweils fünfstrahlig aufgebaut. Bemerkenswert ist die Hand, die neben den vergrößerten Schneidezähnen die auffälligsten Eigentümlichkeiten der Apatemyiden bereitstellt. Hier zeigten die Mittelhandknochen und Fingerglieder am zweiten und dritten Strahl markante Verlängerungen. So erreichten der zweite und dritte Metacarpus eine Länge von 9,5 beziehungsweise 9,2 mm, während der am inneren Strahl nur 5,8, der am äußeren nur 4,5 mm lang wurde. Der Effekt der verlängerten mittleren Finger verstärkte sich noch durch die Streckung der einzelnen Fingerglieder. Die jeweils erste Phalanx wies am zweiten und dritten Finger eine Länge von 8,5 mm auf und war demnach fast genauso lang wie der entsprechende Mittelhandknochen. Die zweiten Phalangen maßen immerhin noch 5,5 bis 6,5 mm. Sie übertrafen damit die jeweiligen Fingerglieder der übrigen Strahlen um rund das Doppelte an Länge. Die Endphalangen an allen fünf Fingern waren dagegen mit 1,5 bis 2 mm vergleichsweise kurz. Sie liefen nach vorn spitz zu und zeigten seitliche Verschmälerungen. Außerdem erhob sich bei ihnen an der Basis jeweils ein kräftiger Sehnenhöcker. Als weitere Besonderheit des zweiten und dritten Fingerstrahls kann hervorgehoben werden, dass die einzelnen einander entsprechenden Knochenelemente zwar leicht abweichende Größen zueinander aufwiesen, beide Strahlen insgesamt aber eine gleiche Länge aufwiesen. Gegenüber der hochspezialisierten Hand war der Fuß relativ ursprünglich gebaut und zeigte keine Streckungen. Die einzelnen Knochen besaßen einen kräftigeren Bau als ihre Gegenstücke an der Hand. Die Strahlen II bis IV waren nahezu gleich lang, die Länge der Mittelfußknochen schwankte zwischen 10,3 und 11 mm, die jeweils ersten beiden Zehenglieder waren 5,5 beziehungsweise 3,7 mm lang. Der Außenstrahl erreichte nicht ganz die Dimension der drei mittleren Strahlen, der Innenstrahl war deutlich kürzer. Die Endphalangen der Zehen glichen denen der Hand.[1][2]
Fossilfunde
Überreste von Heterohyus verteilen sich über weite Bereiche Europas. Generell sind sie aber eher selten und beschränken sich zumeist auf einzelne Zahn- oder Gebissfunde, so dass die Tiere als rares Faunenelement aufgefasst werden können. Das früheste Auftreten markieren einige Backenzähne aus Cuis und Grauves in Frankreich, die in das Untere Eozän datieren.[10] Zu den jüngsten Nachweisen von Heterohyus zählen dagegen jene aus dem Übergang vom Mittleren zum Oberen Eozän etwa die Funde aus der Creechbarrow-Kalkstein-Formation im Hampshire-Becken in England oder aus den Phosphoritlagerstätten bei Quercy im südlichen Frankreich. Erstere Fundstelle umfasst mehr als ein Dutzend Zähne zuzüglich eines Unterkieferfragments,[11] letztere setzt sich ebenfalls aus Zahnmaterial unterschiedlichster Fundstellen zusammen, deren ältestes Fundmaterial bereits Ende des 19. Jahrhunderts vorgestellt worden war.[12][13][14][15] Aus etwa dem gleichen Zeithorizont sollten die zehn Zahnreste von Sossís im nordöstlichen Spanien stammen.[16] Mit einer Stellung im Oberen Eozän noch etwas jünger sind einzelne Zahnfunde aus der Headon-Hill-Formation gleichfalls im Hampshire-Becken gelegen.[17]
Ein bedeutender Fundkomplex liegt aus Egerkingen in der Schweiz vor, der verschiedenste Unter- und Oberkieferfragmente sowie Einzelzähne enthält und dem Mittleren Eozän angehört. Die Reste fanden bereits seit der zweiten Hälfte des 19. Jahrhunderts Beachtung.[18][19] Ähnliches kann zu Bouxwiller im nordostfranzösischen Département Bas-Rhin gesagt werden, welches das Typusmaterial der Gattung barg.[20] Einige wenige Exemplare, darunter ein linkes Unterkieferfragment mit erhaltenem Schneidezahn und der Reihe der hinteren Zähne vom dritten Prämolaren bis zum letzten Molaren, deren Länge gut 15 mm beträgt, kamen aus der Mittel- bis Oberkohle des Geiseltales in Sachsen-Anhalt zu Tage. Der früheste Fund wurde hier bei Forschungsgrabungen in den 1920er Jahren entdeckt.[21] Die bedeutendsten Funde sind bisher in der Grube Messel in Hessen geborgen worden. Von hier stammen wenigstens vier weitgehend vollständige Skelette, die allesamt in Seitenlage erhalten sind und teilweise noch die Konturen der ehemaligen Weichteilbedeckung erkennen lassen. Den ersten Fund tätigte ein Privatsammler bereits 1973, er wurde aber erst später als zu Heterohyus gehörig erkannt. Die drei weiteren Skelette kamen bei regulären Grabungen in den 1980er und 1990er Jahren zum Vorschein. Alle Skelette repräsentieren aufgrund nicht verwachsener Knochennähte Jungtiere.[22][1][2][3] Die gute Erhaltung der Skelette von Messel mit kaum vorhandener Disartikulation geht möglicherweise auf fetthärtende Prozesse während der Kadavereinbettung in ein feuchtes Milieu zurück.[23]
Paläobiologie
Biologische Funktionen

Die gut erhaltenen Skelette aus der Grube Messel erlauben zumindest für die kleineren Vertreter der Gattung Heterohyus einen Einblick in die Lebensweise. In Verbindung mit weiteren vorzüglich erhaltenen Resten verwandter Formen, etwa von Apatemys aus dem Fossil Butte Member der Green-River-Formation[4] oder von Labidolemur aus der Willwood-Formation,[24] beide im US-Bundesstaat Wyoming gelegen, ergibt sich so ein recht detaillierter Blick in die Biologie der Apatemyidae. Insgesamt handelt es sich bei Heterohyus um ein relativ grazil gebautes Tier, deren kleinere Angehörige die Größe des heutigen Siebenschläfers erreichten.[25] Für die Gattung wie für andere Mitglieder der Familie ist die Kombination aus den vergrößerten und vorstehenden Schneidezähnen sowie den verlängerten Fingergliedern auffallend. Eine vergleichbare Merkmalskombination ist heute beim Fingertier (Daubentonia), einer Primatenart auf Madagaskar, und bei den Streifenbeutlern (Dactylopsila) von Neuguinea ausgebildet, wodurch eine deutliche konvergente Entwicklung vorliegt. Bei beiden Gattungen benutzen die Tiere ihre langen Finger, um Insekten und deren Larven aus Spalten und Ritzen in der Baumrinde herauszubohren, während die Tiere mit den großen Incisiven die Hohlräume nagend erweitern. Eine vergleichbare Lebensweise kann für Heterohyus angenommen werden. So findet der nagende Einsatz der Schneidezähne Unterstützung in dem prinzipiell hohen Schädel und den ausladenden Jochbögen, wodurch ausreichend große Ansatzstellen für die Kaumuskulatur bestanden. Der nur einzelne Flächen der Schneidezähne bedeckende Zahnschmelz hatte den Effekt, dass sich die Zahnspitzen während des Gebrauchs immer wieder nachschärften. Abweichungen zum Fingertier und zu den Streifenbeutlern bestehen jedoch in der weiteren Gebissausprägung. Beim Fingertier folgt auf dem unteren Schneidezahn (ebenso wie auf den oberen) jeweils eine größere Zahnlücke, bei den Streifenbeutlern hingegen mehrere sehr kleine Vormahlzähne. Dagegen schließt bei Heterohyus im unteren Gebiss der große vordere Prämolar mit der langen Schneidkante direkt an den Schneidezahn an. Des Weiteren setzt sich das obere Vordergebiss bei den Streifenbeutlern aus drei Schneidezähnen zusammen, von denen nur der erste stark vergrößert ist, die anderen aber flach und niedrig sind. Diese beiden liegen bei Gebissschluss auf dem unteren großen Schneidezahn und werden von diesem flach geschliffen. Die bei den Streifenbeutlern dahinter anschließenden Zähne, ein Eckzahn und mehrere Prämolaren, stehen nicht im Kontakt mit dem unteren Schneidezahn. Dieser abweichende Gebissaufbau hatte möglicherweise Unterschiede im Detail der Ernährungsweise zur Folge. Eventuell ebenfalls in Verbindung mit der Nahrungsaufnahme beziehungsweise -beschaffung oder aber als Hinweis auf bewegliche Lippen können die zahlreichen Foramina im Schnauzenbereich gedeutet werden, die das Vorhandensein von Vibrissen anzeigen. Die stark verkürzten Endphalangen der Finger von Heterohyus ließen sich gut als Widerhaken beim Stochern in Baumlöchern oder zum Herausangeln von Insekten einsetzen. Als Hinweis darauf kann der robuste Sehnenhöcker an der Phalangenbasis herhalten. Auffallende Unterschiede ergeben sich allerdings aus der Lage der verlängerten Finger. Während beim Fingertier und bei den Streifenbeutlern jeweils der dritte und vierte Strahl gestreckt sind, weisen bei Heterohyus der zweite und dritte Strahl diese Merkmale auf. Die leicht abgewandelte Anatomie der beiden verlängerten Finger bei Heterohyus lassen sich dahingehend interpretieren, dass diese nicht gleichzeitig benutzt wurden oder aber eine leicht abgewandelte Funktion besaßen beziehungsweise eventuell gegeneinander wirkten. Ungeklärt bleibt bisher, ob die sehr großen Vertreter von Heterohyus eine vergleichbare Lebensweise verfolgten. Hier ist bisher zu wenig Fossilmaterial überliefert. Eine ähnliche Problematik liegt beim ausgestorbenen Verwandten des Fingertiers, dem im Vergleich extrem großen Daubentonia robusta vor. Das heutige Fingertier ist ebenfalls deutlich größer als die kleineren Angehörigen von Heterohyus und ernährt sich neben holzbohrenden Insekten auch von Samen und Früchten. Eventuell haben die großen Heterohyus-Formen zusätzliche andere Nahrungsquellen erschlossen.[26][1][2][25][3]

Einige Skelette aus Messel lassen durch bakterielle Nachzeichnungen (Bacteriographie) den ehemaligen Körperumriss und die Fellbedeckung erkennen. Die Nachzeichnungen sind eng am Skelett anliegend. Der gesamte Rumpf war wohl mit längeren Haaren bedeckt. Lediglich am Nacken traten dickere, möglicherweise borstenartige Haare auf. Auffällig ist der Schwanz, dessen Spitze bei einem Exemplar ein dickes Haarbüschel aufweist, das zwischen dem 27. (bauchseitig) und 32. (rückenseitig) Schwanzwirbel ansetzt und rund 15 % der Schwanzlänge einnimmt. Einzelne Haare sind rund 15 mm lang. Der restliche Schwanz war wohl eher nur spärlich mit Fell bedeckt, wie einige einzelne Haarabdrücke aufzeigen. Markante Schwanzbüschel sind von zahlreichen baumlebenden oder springenden Säugetieren belegt und dienen häufig als Steuerungsorgan bei der Fortbewegung. Hierzu gehören unter den Primaten etwa die Sulawesi-Koboldmakis oder der Senegal-Galago beziehungsweise unter den Spitzhörnchen das Federschwanz-Spitzhörnchen, die allesamt behände Baumspringer und -kletterer sind. Daneben zeigen auch einzelne Nagetiere wie der Südafrikanische Springhase oder die Pferdespringer als bodenbewohnende Springer beziehungsweise die Bilchschwänze oder Pinselstachler als Baumkletterer ein vergleichbares Schwanzende. Im Gegensatz zu zahlreichen der rezenten, sich springend fortbewegenden Säugetiere besitzt Heterohyus jedoch keine Anpassungen an eine solche Lebensweise, da die Hinterextremitäten nur mäßige Verlängerungen aufweisen. Jedoch impliziert die vermutete Ernährungsweise ein baumkletterndes Leben. Skelettanpassungen dafür lassen sich aber nur wenige finden. Ob der Schwanz beispielsweise als Greiforgan fungierte, ist derzeit unbekannt. Die hinteren Schwanzwirbel büßen nicht abrupt an Länge ein, was beispielsweise beim Greifschwanz der Primaten der Fall ist. Im Unterschied dazu verkürzen sich bei zahlreichen Beutelratten mit Greifschwanz die Schwanzwirbel nach hinten eher kontinuierlich. Dem gegenüber ermöglichte das gut entwickelte Schlüsselbein eine freie Rotation der Arme. Die kurzen und hohen Krallen des Hinterfußes wiederum waren gut für ein Anhaften in der Baumrinde geeignet. Die weiten Gelenkflächen an den Fußwurzelknochen bewirkten eine hohe Drehbarkeit des Fußes. Allerdings schränkten die am unteren Ende miteinander verwachsenen Knochen des Unterschenkels (Schien- und Wadenbein) diese Gelenkigkeit möglicherweise wieder zu einem gewissen Grad ein. Die parallel liegenden Strahlen des Fußes verweisen wiederum auf einen Sohlengang, bei dem der Fuß in ganzer Breite aufsetzte.[2][25][3]
Ökologische Implikationen
Die starke Spezialisierung von Heterohyus mit einer baumbewohnenden Lebensweise und einer Ernährung von holzbohrenden oder von in Holz lebenden Insekten macht es wahrscheinlich, dass die Gattung und möglicherweise auch andere Vertreter der Apatemyiden die ökologische Nische der heutigen Spechte einnahm. In Bezug auf die heutige Verbreitung des Fingertiers und der Streifenbeutler ist anzumerken, dass weder Madagaskar noch Neuguinea von den Spechten besiedelt wurden und diese somit auf den beiden Inseln durch die genannten Säugetiere ersetzt werden. Die Spechte entwickelten sich im Verlauf des Neogens, das Auftreten der Apatemyiden im Paläogen und das Fehlen fossiler Nachweise der Vögel aus der Grube Messel, dem Geiseltal und anderen Fundstätten jener Zeit unterstützt damit diese Ansicht.[26][1][25][3]
Systematik
Mögliches Verwandtschaftsverhältnis der Apatemyidae nach Silcox et al. 2010[24]
|
Heterohyus ist eine Gattung aus der ausgestorbenen Familie der Apatemyidae. Die Vertreter dieser Gruppe stellen überwiegend kleine, insektenfresserartige Säugetiere dar, welche im heutigen Nordamerika und in Europa fossil nachgewiesen sind. In Nordamerika erschienen sie erstmals im Paläozän, ihr letztes Auftreten hatten sie dort im Oligozän. Aus Europa liegen Nachweise vor allem aus dem Eozän vor, die Tiere verschwanden dann mit dem Einsetzen des Grande Coupure. Der Großteil des Fundmaterials der Apatemyidae beschränkt sich auf Zähne und Gebissreste. An diesen lassen sich als charakteristische Merkmale die auffallend vergrößerten und gebogenen Schneidezähne herausarbeiten, ebenso wie die relativ kleinen Backenzähne. Die verwandtschaftlichen Beziehungen der Apatemyidae waren lange Zeit ungeklärt und sind auch aus heutiger Sicht schwer zu beurteilen. Vor allem zu Beginn des 20. Jahrhunderts wurde häufig eine Einbindung in die Primaten oder Insektenfresser favorisiert.[27][28] Die besondere Stellung der Apatemyidae drückten William Berryman Scott und Glenn Lowell Jepsen im Jahr 1936 mit der Etablierung des höheren Taxons der Apatotheria aus, deren einziges Mitglied sie sind.[29] Später konnte auch anatomisch belegt werden, dass die Apatemyidae nicht zu den Primaten gehören.[30][31] Des Weiteren bestehen einzelne Übereinstimmungen zu den Insektenfressern, die aber hauptsächlich die rezenten Formen betreffen, etwa bei der Ausprägung der Schneidezähne und bei einzelnen Schädelmerkmalen, während zu den fossilen Vertretern größerer Abweichungen erkennbar sind. Nach phylogenetischen Untersuchungen aus dem Jahr 2010 lassen sich die Apatemyidae möglicherweise eher der Gruppe der Euarchontoglires zuordnen. Aufgrund der nur wenigen definierenden gemeinsamen anatomischen Merkmale der Euarchontoglires – diese wurden im Jahr 2001 anhand genetischer Gemeinsamkeiten definiert[32] – kann hierin sowohl die Möglichkeit einer engeren Bindung an die Glires (Nagetiere und Hasenartige) als auch an die Euarchonta (Primaten, Spitzhörnchen und Riesengleiter) nicht ausgeschlossen werden. Innerhalb der Apatemyidae bildet Heterohyus einen eher entwickelten Vertreter. Die nächsten Verwandten finden sich möglicherweise in der europäischen Form Carcinella aus dem Oberen Eozän sowie im nordamerikanischen Vertreter Sinclairella, welcher noch im spätesten Oligozän vorkam.[24]
Folgende Arten von Heterohyus sind anerkannt:[5]
- H. armatus Gervais, 1848
- H. europaeus (Rütimeyer, 1890)
- H. gracilis (Hans Georg Stehlin, 1916)
- H. heufelderi Heller, 1930
- H. morinionensis Hooker, 1986
- H. nanus Teilhard, 1922
- H. quercyi (Filhol, 1890)
- H. pygmaeus Sigé, 1990
- H. sudrei Sigé, 1975
Verschiedentlich werden innerhalb der Gattung Heterohyus einzelne Untergattungen abgetrennt, so etwa Gervaisyus, welche die Art H. pygmaeus einschließt, oder Chardinyus mit der Art H. nanus. Die Unterscheidung erfolgt anhand der Ausprägung bestimmter Merkmale an den Molaren, etwa das Vorhandensein oder Fehlen bestimmter Höcker wie dem Paraconid.[13][16]
Forschungsgeschichte

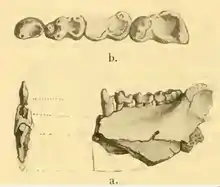
Die wissenschaftliche Erstbeschreibung von Heterohyus erfolgte im Jahr 1848 durch Paul Gervais in dessen Werk Zoologie et paléontologie françaises. Gervais stellte darin ein Unterkieferfragment aus Bouxwiller im nordostfranzösischen Département Bas-Rhin vor und nahm für Heterohyus eine Verwandtschaft mit den Wildschweinen sowie eine allesfressende Lebensweise an. Als Art wies er Heterohyus armatus aus.[20] Die Abbildung des Unterkiefers, die Gervais in seiner Erstbeschreibung publiziert hatte, wurde nur wenig später von Henri Marie Ducrotay de Blainville wiederverwendet. Allerdings verwies er ihn zu Arctomys, einem heutigen Synonym für die Gattungsbezeichnung Marmota der Murmeltiere.[33] In der zweiten Auflage seines Werkes Zoologie et paléontologie françaises aus dem Jahr 1859 lehnte Gervais diese Einschätzung de Blainvilles ab und verschob Heterohyus daher wieder zu den Schweinen. Gleichzeitig etablierte er mit den „Heteromyines“ eine eigenständige Tribus, die aber nicht anerkannt ist.[34] Zu einer völlig anderen Einschätzung kam Max Schlosser Ende der 1880er Jahre. Er schloss zwar Heterohyax in seiner sehr umfassenden, heute nicht anerkannten Gruppe der Pseudolemuridae ein, sah in der Form aber ein Artefakt, das sich aus mehreren anderen Gattungen verschiedener Primatenformen wie Adapis oder Caenopithecus und weiteren zusammensetzte.[35][6]
Im gleichen Zeitraum wie Schlosser, 1890, analysierte Henri Filhol Fossilreste aus den Phosphoritlagerstätten von Quercy im südlichen Frankreich und führte basierend auf Unterkieferresten eine Art namens Necrosorex quercyi ein. Er vermutete bei der sehr kleinen Form eine Verwandtschaft mit den Spitzmäusen, woraus sich auch die wissenschaftliche Gattungsbezeichnung erklärt.[12] Noch im gleichen Jahr benannte Ludwig Rütimeyer Calamodon europaeus unter Verwendung einzelner Gebissfragmente aus Egerkingen und ordnete diese sehr große Form den Taeniodonta zu, einer ausgestorbenen Gruppe insektenfresserartiger Säugetiere.[18] Hans Georg Stehlin erkannte dann 1916 bei seiner Aufarbeitung des Egerkingener Fundmaterials Ähnlichkeiten zu Heterohyus, stellte aber mit der Gattung Amphichiromys eine neue Form auf. Gleichzeitig etablierte er mit Heterochiromys und der Art Heterochiromys gracilis von der gleichen Fundstelle einen weiteren Vertreter, der deutlich kleiner ist als Amphichiromys europaeus. Für beide Gattungen und Arten sah Stehlin eine engere Bindung zum heutigen Fingertier und damit zur Gruppe der Primaten. In den gleichen Verwandtschaftskreis verschob er zudem Filhols Necrosorex, bei dem er ebenfalls die Übereinstimmungen mit Heterohyus hervorhob.[19] Alle drei Gattungen gelten heute als synonym zu Heterohyus.[6][36][37]
In der Folgezeit setzte sich die Gattungsbezeichnung Heterohyus zunehmend durch. Mit H. nanus beschrieb Teilhard de Chardin im Jahr 1922 die am häufigsten dokumentierte Form. Sie ist nicht nur aus Quercy bekannt, von wo das Belegmaterial stammt, sondern wurde unter anderem auch aus Spanien und England berichtet.[16][11] Zusätzlich werden ihr provisorisch alle in der Grube Messel aufgefundenen Skelette zugewiesen.[5] Die Art H. heufelderi erhielt im Jahr 1930 von Florian Heller basierend auf einem Unterkieferast aus dem Geiseltal ihre wissenschaftliche Beschreibung.[21] Einige Autoren stufen sie aber als identisch mit H. gracilis aus Egerkingen ein.[10][11] Die mittelgroße Form H. morinionensis basiert auf rund einem Dutzend Zähnen aus der Creechbarrow-Kalkstein-Formation und wurde 1986 von Jerry J. Hooker wissenschaftlich eingeführt. Sie ähnelt relativ stark H. quercyi.[11] Bernard Sigé benannte im Jahr 1975 H. sudrei aus Robiac in Frankreich. Fünfzehn Jahre später folgte mit H. pygmaeus eine weitere sehr kleine Form aus Quercy, deren Grundlage einige wenige Zähne bilden.[13]
Literatur
- Daniela C. Kalthoff, Wighart von Koenigswald und Cornelia Kurz: A new specimen of Heterohyus nanus (Apatemyidae, Mammalia) from the Eocene of Messel (Germany) with unusual soft-part preservation. Courier Forschungsinstitut Senckenberg 252, 2004, S. 1–12
- Wighart von Koenigswald, Gregg F. Gunnell, Thomas Lehmann, Kenneth D. Rose und Irina Ruf: Vier ursprüngliche, aber hoch spezialisierte Säugetiere. In: Stephan S. K. Schaal, Krister T. Smith und Jörg Habersetzer (Hrsg.): Messel – Ein fossiles Tropenökosystem. Senckenberg-Buch 79, Stuttgart, 2018, S. 223–233
Einzelnachweise
- Wighart von Koenigswald: Die Paläobiologie der Apatemyiden (Insectivora s. l.) und die Ausdeutung der Skelettfunde von Heterohyus nanus aus dem Miotteleozän von Messel bei Darmstadt. Palaeontographica Abteilung A, 210 (1–3), 1990, S. 41–77
- Daniela C. Kalthoff, Wighart von Koenigswald und Cornelia Kurz: A new specimen of Heterohyus nanus (Apatemyidae, Mammalia) from the Eocene of Messel (Germany) with unusual soft-part preservation. Courier Forschungsinstitut Senckenberg 252, 2004, S. 1–12
- Wighart von Koenigswald, Gregg F. Gunnell, Thomas Lehmann, Kenneth D. Rose und Irina Ruf: Vier ursprüngliche, aber hoch spezialisierte Säugetiere. In: Stephan S. K. Schaal, Krister T. Smith und Jörg Habersetzer (Hrsg.): Messel – ein fossiles Tropenökosystem. Senckenberg-Buch 79, Stuttgart, 2018, S. 223–233
- Wighart von Koenigswald, Kenneth D. Rose, Lance Grande und Robert D. Martin: First apatemyid skeleton from the Lower Eocene Fossil Butte Member, Wyoming (USA), compared to the European apatemyid from Messel, Germany. Palaeontographica Abteilung A 272, 2005, S. 149–169
- Wighart von Koenigswald, Irina Ruf und Philip D. Gingerich: Cranial morphology of a new apatemyid, Carcinella sigei n. gen. n. sp. (Mammalia, Apatotheria) from the Late Eocene of Southern France. Palaeontographica Abteilung A 288, 2009, S. 53–91
- Malcolm C. McKenna: Primitive Paleocene and Eocene Apatemyidae (Mammalia, Insectivora) and the Primate-Insectivore Boundary. American Museum Novitates 2160, 1963, S. 1–39
- Philip D. Gingerich und Kenneth D. Rose: Studies on the Paleocene and Early Eocene Apatemyidae (Mammalia, Insectivora). I. Dentition of Clarkforkian Labidolemur kayi. Contributions from the Museum of Paleontology, The University of Michigan 26 (4), 1982, S. 49–55
- Philip D. Gingerich: Studies on the Paleocene and Early Eocene Apatemyidae (Mammalia, Insectivora). II. Labidolemur and Apatemys from the Early Wasatchian of the Clark's Fork Basin, Wyoming. Contributions from the Museum of Paleontology, The University of Michigan 26 (4), 1982, S. 57–69
- Nicholas J. Czaplewski und Gary S. Morgan: A late-surviving apatemyid (Mammalia: Apatotheria) from the latest Oligocene of Florida, USA. PeerJ 3, 2015, S. e1509; doi: 10.7717/peerj.1509
- D. E. Russell, M. Godinot, P. Louis und D. E. Savage: Apatotheria (Mammalia) de l'Éocèn inférieur de France et de Belgique. Bulletin du Muséum national d'histore naturelle 4e, ser C:1, 1979, S. 203–243 ()
- J. J. Hooker: Mammals from the Bartonian (Middle/Late Eocene) of the Hampshire Basin, southern England. Bulletin of the British Museum (Natural History) 39 (4), 1986, S. 191–478 (S. 327–336)
- Henri Filhol: Description d'un nouveau genre d'insectivore. Bulletin de la Société philomathique de Paris 8 (1–2), 1890, S. 174–175 ()
- Jean Sudre, Bernard Sigé, Jean albert Remy, Bernard Marandat, Jean-Louis Hartenberger, Marc Dodinot und Jean Yves Crochet: Une faune du niveau d’Egerkingen (MP 14; Bartonien Inférieur) dans les phosphorites du Quercy (Sud de la France). Palaeovertebrata 20 (1), 1990, S. 1–32
- E. Maitre, M. Hugueney, J. G. Astruc, J.-Y. Crochet, G. Escarguel, M. Godinot, S. Legendre, B. Marandat, C. Mourer-Chauvierế, J.-C. Rage, J. A. Rếmy, R. Simon-Coinçon, J. Sudre, P. Valette und B. Sigế: Huit nouvelles faunes éocènes et oligocènes des phosphorites du Quercy. In: B. Sigế und T. Pelissie (Hrsg.): 30 Millions d'annees de biodiversite dynamique dans le paleokarst du Quercy. Strata 13, 2006, S. 113–127
- B. Sigế und M. Hugueney: Les micromammifères des gisements à phosphates du Quercy (SW France). In: B. Sigế und T. Pelissie (Hrsg.): 30 Millions d'annees de biodiversite dynamique dans le paleokarst du Quercy. Strata 13, 2006, S. 207–227
- Bernard Sigé: Les mammifères insectivores des nouvelles collections de Sossís et sites associés (Éocène Superieur, Espagne). Geobios30 (1), 1997, S. 91–113
- Katerina Vasileiadou, Jerry J. Hooker und Margaret E. Collinson: Paleocommunity reconstruction and accumulation of micromammalian remains (Late Eocene, Southern England). Palaios 24, 2009, S. 553–567
- Ludwig Rütimeyer: Übersicht der Eocänen Fauna von Egerkingen. Verhandlungen der Naturforschenden Gesellschaft in Basel 9, 1893, S. 331–362 ()
- Hans Georg Stehlin: Die Säugetiere des schweizerischen Eocaens. Critischer Catalog der Materialien. Abhandlungen der schweizerischen paläontologischen Gesellschaft 56, 1916, S. 1209–1556 (S. 1434–1508)
- Paul Gervais: Zoologie et paléontologie françaises (animaux vertébres). Paris, 1848–1852, Tome 1., S. 163 (), Tome 2., unnummeriert ()
- Florian Heller: Die Säugetierfauna der Mitteleozänen Braunkohle des Geiseltales bei Halle a. S. Jahrbuch des Halleschen Verbandes für die Erforschung der Mitteldeutschen Bodenschätze und ihrer Verwertung 9, 1930, S. 13–41
- Wighart von Koenigswald: Apatemyiden-Skelette aus dem Mitteleozän von Messel und ihre paläobiologische Aussage. Carolinea 45, 1987, S. 31–35
- Achim H. Schwermannm, M. Wuttke und J. A. Schultz: Virtopsy of the controlled decomposition of a dormouse Eliomys quercinus as a tool to analyse the taphonomy of Heterohyus nanus from Messel (Eocene, Germany). Palaeobiology and Palaeoenvironments 92, 2012, S. 29–43
- Mary T. Silcox, Jonathan I. Bloch, Doug M. Boyer und Peter Houde: Cranial anatomy of Paleocene and Eocene Labidolemur kayi (Mammalia: Apatotheria), and the relationships of the Apatemyidae to other mammals. Zoological Journal of the Linnean Society 160, 2010, S. 773–825
- Wighart von Koenigswald, Kenneth D. Rose, Lance Grande und Robert D. Martin: Die Lebensweise eozäner Säugetiere (Pantolestidae und Apatemyidae) aus Messel (Europa) im Vergleich zu neuen Skelettfunden aus dem Fossil Butte Member von Wyoming (Nordamerika). Geologisches Jahrbuch Hessen 132, 2005, S. 43–54
- Wighart von Koenigswald und H. P. Schierning: The ecological niche of an extinct group of mammals, the early Tertiary apatemyids. Nature 326, 1987, S. 595–597
- William Diller Matthew: The Carnivora and Insectivora of the Bridger Basin, Middle Eocene. Memoirs of the American Museum of Natural History 9 (4), 1909, S. 291–576 (S. 543–549)
- Glenn L. Jepsen: A revision of the American Apatemyidae and the description of a new genus, Sinclairella, from the White River Oligocene of South Dakota. Proceedings of the American Philosophical Society 74 (4), 1934, S. 287–305
- William Berryman Scott und Glenn Lowell Jepsen: The Mammalian Fauna of the White River Oligocene: Part I. Insectivora and Carnivora. Transactions of the American Philosophical Society, New Series 28 (1), 1936, S. 1–153
- Johannes Hürzeler: Über die europäischen Apatemyiden. Eclogae Geologicae Helvetiae 42, 1949, S. 485 ()
- Kenneth D. Rose: The beginning of the age of mammals. Johns Hopkins University Press, Baltimore, 2006, S. 1–431 (S. 103–105)
- William J. Murphy, Eduardo Eizirik, Stephen J. O‘Brien, Ole Madsen, Mark Scally, Christophe J. Douady, Emma Teeling, Oliver A. Ryder, Michael J. Stanhope, Wilfried W. de Jong und Mark S. Springer: Resolution of the Early Placental Mammal Radiation Using Bayesian Phylogenetics. Science 294, 2001, S. 2348–2351
- Henri Marie Ducrotay de Blainville: Ostéographie ou description iconographique comparée du squelette et du systeme dentaire des mammiferes recents et fossiles. Paris, 1839–1864, unnummeriert ()
- Paul Gervais: Zoologie et paléontologie françaises (animaux vertébres). Paris, 1859, S. 201–201 (), Atlas Tafel 35 ()
- Max Schlosser: Die Affen, Lemuren, Chiropteren, Insectivoren, Marsupialier, Creodonten und Carnivoren des Europäischen Tertiärs und deren Beziehungen zu ihren lebenden und fossilen aussereuropäischen Verwandten. Wien, 1888–1890, S. 1–492 (S. 451–452) ()
- Leigh van Valen: New Paleocene Insectivores and Insectivore classification. Bulletin of the American Museum of Natural History 135, 1967, S. 217–284
- Malcolm C. McKenna und Susan K. Bell: Classification of mammals above the species level. Columbia University Press, New York, 1997, S. 1–631 (S. 212–213)