Steppennashorn
Das ausgestorbene Steppennashorn (Stephanorhinus hemitoechus) war eine pleistozäne Nashornart Eurasiens und Nordafrikas. Erstmals wurde es im Jahr 1868 durch den schottischen Paläontologen Hugh Falconer (1808–1865) wissenschaftlich unter dem Namen Rhinoceros hemitoechus beschrieben. Das Steppennashorn lebte im Gegensatz zum nah verwandten Waldnashorn (Stephanorhinus kirchbergensis) in eher offenen Landschaften und ernährte sich zum größeren Teil von harter Grasnahrung. Es starb wie das Waldnashorn erst im späten Pleistozän aus, überlebte aber regional neueren Untersuchungen zufolge bis zum Beginn des Holozäns.
| Steppennashorn | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Schädel | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Mittel- bis Jungpleistozän | ||||||||||||
| 500.000 bis 10.000 Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Stephanorhinus hemitoechus | ||||||||||||
| (Falconer, 1859, 1868) | ||||||||||||
Verbreitung und Lebensraum
Das Steppennashorn lebte im Mittel- und Jungpleistozän in weiten Teilen Europas und Asiens, von der Iberischen Halbinsel bis weit nach Mittelasien. Weiterhin ist es von Fundstellen in der Levante (Libanon, Syrien, Israel) sowie aus Nordafrika (Marokko, Libyen) bekannt.[1] In Nordafrika trat das Steppennashorn jedoch erst im Jungpleistozän auf, nachdem es wahrscheinlich aus dem Norden über den Nahen Osten eingewandert war.[2][3]
Wie sein naher Verwandter, das Waldnashorn, war auch das Steppennashorn eine an sub-mediterranes bis warm-gemäßigtes Klima angepasste Form. Seine angestammten Lebensräume waren das südliche Europa und die west- und mittelasiatischen Steppen, Busch- oder auch Waldgebiete. Unklar ist dabei, ob es, wie sein Name suggeriert, wirklich ein typisches Steppentier war oder eher ein Bewohner offener Waldhabitate und Parklandschaften.[4] Gegen eine starke Anpassung an Steppen spricht vor allem die Tatsache, dass Steppennashörner in Europa im Fossilbericht relativ häufig sind, aber in Asien, wo Steppengebiete auch im Pleistozän viel weiter verbreitet waren, eher selten gefunden wurden.[5] Allerdings scheint diese Tierart gegenüber kühlerem und trockenem Klima wesentlich resistenter gewesen zu sein als das Waldnashorn, da sie auch in spätinterglazialen bzw. kühleren Phasen der Warmzeiten nördlich der Alpen nachgewiesen ist und dann teilweise mit dem Wollnashorn (Coelodonta antiquitatis) zusammen auftritt. Während der Warmzeiten des Mittel- und Jungpleistozäns, so in der Holstein-Warmzeit (vor 340.000 bis 325.000 Jahren) und der Eem-Warmzeit (vor 126.000 bis 115.000 Jahren), verbreitete es sich auch bis weit in das nordalpine Europa (Mittel- und Osteuropa), wobei Fundplätze in den Niederlanden (Zwarte Water), in England (Illford, Barrington) und in Wales (Minchin-Höhle) sein nördlichstes Vorkommen markieren.[1][6] In den Kaltzeiten zog es sich jedoch aus den Gebieten nördlich der Alpen in den Süden zurück und wurde in den so frei werdenden Arealen durch das Wollnashorn vollständig ersetzt.[7]
Aufgrund der weiten Verbreitung im warmzeitlichen Europa koexistierte das Steppennashorn mit dem Waldnashorn. In Gebieten, wo beide Tierarten miteinander vorkamen, trat das Steppennashorn aber in den Hintergrund, da sich sein Verwandter offensichtlich besser durchsetzen konnte. Ausnahmen bilden Regionen, die das Waldnashorn nicht erreichte, etwa England in der Eem-Warmzeit oder die Iberische Halbinsel im Allgemeinen.[8] In diesen Gebieten bewohnte das Steppennashorn wahrscheinlich auch weniger offene Landschaften und dürfte mangels Konkurrenz auch in Waldgebiete vorgedrungen sein.[4]
Fundplätze, an denen das Steppennashorn auftritt, sind in Europa recht weit verbreitet, mit jeweils rund einem Dutzend Fundorten wurde es in Deutschland und Spanien am häufigsten registriert. Bekannt sind die eemzeitlichen Funde von Neumark-Nord 1 aus dem ehemaligen Tagebau Mücheln im Geiseltal (Sachsen-Anhalt),[9] wo neben zwei Schädeln auch 1988 bei Tagebauarbeiten ein weitgehend vollständiges Skelett aufgefunden wurde, welches mit einigen Feuersteinartefakten verbunden war, was für eine menschliche Manipulation des Kadavers spricht. Auch im Travertin von Ehringsdorf (Thüringen), der rund 200.000 Jahre alten Fundstelle des Ehringsdorfer Urmenschen, sind zahlreiche Reste des Steppennashorns überliefert, darunter auch ein fast vollständig erhaltener Schädel.[10] In der Fauna des seit 1969 wissenschaftlich erforschten archäologischen Fundplatzes von Bilzingsleben in Nordthüringen,[11] in der auch Homo erectus bzw. Homo heidelbergensis nachgewiesen ist und die ein Alter von etwa 350.000 Jahren aufweist, tritt das Steppennashorn (zusammen mit dem Waldnashorn) relativ häufig auf. Die Knochen und überwiegenden Gebissreste sind aber stark zerstückelt und selten vollständig,[12] was die damaligen Ausgräber unter Hinzuziehung anderer Hinweise dazu bewog, diese als Reste der Jagdbeute des frühen Menschen zu interpretieren. Andere bedeutende Fundstellen in Deutschland sind Steinheim an der Murr (Baden-Württemberg),[13] bekannt durch den Steinheimer Urmenschen, und Mosbach (Stadtgebiet Wiesbaden, Hessen).[14] An letzterer, eines der frühesten Nachweise des Steppennashorns in Europa, ist es aber nur durch wenige Reste repräsentiert, da hier ansonsten das Hundsheimer Nashorn dominiert, welches bald darauf ausstirbt.
Körperbau und Ernährungsweise
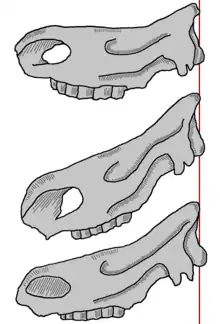
Vom Steppennashorn sind wie vom gleichzeitig auftretenden Waldnashorn nur Knochen- und Zahnreste bekannt, in einigen Fällen liegen mehr oder weniger vollständig überlieferte Skelette vor. Anhand dieser lässt sich sein Aussehen aber mit Hilfe anatomischer Untersuchungen und Vergleichen genauer rekonstruieren. Der Körper war kräftig gebaut, seine Gliedmaßen waren aber wesentlich kürzer und breiter als beim Waldnashorn und besaßen schwach ausgeprägte Gelenke, was eine Anpassung an das Leben in offenen Landschaften darstellt.[15] Im Bezug auf die Körpergröße war es etwas kleiner als sein ebenfalls fossiler Verwandter, das Waldnashorn, welches als der größte Vertreter der Gattung Stephanorhinus gilt[16] und vergleichbar mit dem heutigen Indischen Panzernashorn ist. Allgemein wird das Steppennashorn für ein Tier seiner Größe als recht schlank oder grazil beschrieben. Als rekonstruiertes Lebendgewicht werden etwa 1,4 Tonnen[17] bis 3,4 Tonnen[18] angegeben.
Der Schädel des Steppennashorns, der bis zu 76 cm lang wurde, war am Hinterhaupt stark verlängert, so dass es den Kopf schräg nach unten trug. Dadurch unterscheidet es sich sehr markant von den anderen Arten der Stephanorhinus-Linie, die den Kopf deutlich aufgerichteter hielten, und ähnelt mehr dem Wollnashorn oder dem heutigen Breitmaulnashorn (Ceratotherium simum).[1][14] Die Art besaß zwei Hörner, welche am Nasenbein und am mittleren Schädel lagen. In den gefundenen Fossilien sind die aus Keratin bestehenden, jedoch nicht konserviert überlieferten Hörner durch blumenkohlartig gewucherte Ansatzflächen markiert. Die Ausprägung dieser gerauten Oberflächen zeigt, dass vor allem das vordere Horn sehr groß gewesen sein muss, worauf auch die im Gegensatz zum Waldnashorn stärker verknöcherte Nasenscheidewand hinweist.[8][1]

Der Unterkiefer maß bis zu 56 cm in der Länge und war massiv gebaut, wobei der Unterkieferkörper bis zu 11 cm hoch wurde. Das Gebiss zeichnete sich wie bei allen Vertretern der Stephanorhinus-Linie durch das Fehlen der Schneidezähne aus, weiterhin durch die reduzierte Anzahl der Vorbackenzähne (drei je Kieferbogen) und der eigentlichen Backenzähne (ebenfalls drei). Die daraus resultierende Zahnformel lautet: Dabei waren sowohl der zweite Prämolar als auch der dritte Molar besonders grazil bei dieser Art.[12][19] Eine anormale Besonderheit, die vereinzelt beim Steppennashorn beobachtet wurde, ist die Ausbildung eines vierten Backenzahns, wie es eigentlich typisch ist für Beutelsäuger.[8] Die Zähne waren allgemein recht hochkronig und besaßen eine dicke Zementschicht – ein Merkmal, das bei dieser Tierart innerhalb der Stephanorhinus-Linie am deutlichsten ausgeprägt war – und sprechen dadurch, wie auch die Haltung des Schädels, für eine hauptsächlich grasfressende Ernährungsweise. Da aber die Zähne nicht immer horizontal gleichmäßig abgeschliffen sind, was bei allen grasfressenden Säugetieren durch die im Gras enthaltene harte Kieselsäure zu beobachten ist, sondern gelegentlich auch trogartige Eintiefungen vorkommen, wird davon ausgegangen, dass das Steppennashorn auch weichere Nahrung wie Blätter, Blüten oder Früchte zu sich genommen hat.[20][21]

Das postcraniale Skelett ist weitaus weniger bekannt. Die Wirbelsäule umfasste wenigstens 7 Hals-, 18 Brust- und 4 Lendenwirbel, die genaue Anzahl der Kreuzbein- und Schwanzwirbel ist nicht überliefert. Der Oberarmknochen konnte bis zu 46 cm lang werden, die Ulna 51 cm. Der Oberschenkelknochen erreichte 50 cm Länge und war relativ robust. Vermutlich waren die Hände und Füße, wie bei den heutigen modernen Nashörnern aber auch bei fossilen Arten von Stephanorhinus nachgewiesenen, dreistrahlig aufgebaut, wobei der Mittelstrahl (Metapodium III) besonders stark ausgeprägt war. Ein weitgehend vollständig überlieferter Fuß aus dem Geiseltal hatte einen 18 cm langen Mittelfußknochen.[8]
Über das weitere Aussehen dieser Art ist kaum etwas bekannt, da es keine überlieferten Weichteile gibt. So können keine Angaben über die Größe und Form der Ohren oder dem Aussehen der Haut gemacht werden. Spekulativ ist die Annahme, diese Art habe ein Fell besessen, das Sumatra-Nashorn als sein nächster heute lebender Verwandter ist allerdings behaart. Ob dies nun auch für das pleistozäne Steppennashorn zutrifft, kann nicht ohne weiteres gesagt werden. Sein Vorkommen auch in kühleren klimatischen Abschnitten der Warmzeiten,[1][6] die vor allem im nordalpinen Europa auch mit kalten Wintern verbunden sind, könnte dies wahrscheinlich machen.
Systematik und Stammesgeschichte
Die Gattung Stephanorhinus umfasst neben dem Steppennashorn auch das zeitgleich mit ihm auftretende Waldnashorn (Stephanorhinus kirchbergensis) sowie die stammesgeschichtlich älteren Arten Etruskisches Nashorn (Stephanorhinus etruskus), das Hundsheimer Nashorn (Stephanorhinus hundsheimensis) und weitere. Zusammen mit der Gattung Dicerorhinus und Coelodonta gehört Stephanorhinus zu den Dicerorhinina, einer Gruppe von zweihörnigen Nashörnern, deren einziger heute noch lebender Vertreter das in seinem Bestand stark gefährdete Sumatra-Nashorn (Dicerorhinus sumatrensis) ist.
Über den Ursprung der Art ist wenig bekannt. Es wird davon ausgegangen, dass sie sich in Asien aus einer Spätform des Etruskischen Nashorns entwickelte und später nach Europa einwanderte.[22][12] In Europa erschien sie erstmals vor 450.000 bis 500.000 Jahren im mittleren Pleistozän. Die ältesten Funde stammen aus Wiesbaden (Hessen, Fundkomplex Mosbach 3) und der Höhle von Arago (Frankreich). Bei älteren Fundstellen, die mit dem Steppennashorn in Verbindung gebracht wurden, wie beispielsweise Süßenborn (Thüringen),[1] handelt es sich höchstwahrscheinlich um Verwechslungen mit dem Hundsheimer Nashorn oder dem Etruskischen Nashorn, welche beide phylogenetisch ältere Formen sind.[8][23]
Während seines frühen Auftretens war das Steppennashorn noch relativ grazil und glich in der Variationsbreite den frühen Vertretern der Gattung Stephanorhinus. Fundstellen mit solchen Formen sind unter anderem Bilzingsleben (Thüringen), Steinheim an der Murr (Baden-Württemberg) und Orgnac (Frankreich). Vor allem im späten Mittelpleistozän und im Jungpleistozän nahm es dann an Robustheit zu. Solche großen Steppennashörner sind in Ehringsdorf (Thüringen), Geiseltal (Sachsen-Anhalt) und Crayford (England) nachgewiesen.[21][14] Aus diesem Grund wurden bereits in den 1960er Jahren zwei Subspezies eingeführt, wobei Stephanorhinus hemitoechus falconeri die ältere und Stephanorhinus hemitoechus aretinus die jüngere Form umfasst.[24]
Nach der Eem-Warmzeit zog sich das Steppennashorn wie viele wärmeadaptierten Tiere nach Süden ins Mittelmeergebiet zurück. Funde aus dieser Spätzeit dieser Tierart, die dann nur noch in isolierten Populationen vorkam, stammen aus den Karsthöhlen von Melpigano in der süditalienischen Gemeinde Cursi bzw. von der Pontinischen Ebene bei Rom und sind zwischen 100.000 und 70.000 Jahre alt.[25][26] Wann das Steppennashorn letztendlich ausgestorben ist, kann nicht genau gesagt werden. Ursprünglich ging man davon aus, dass es schon kurz nach Beginn oder in der ersten Hälfte der Weichsel-Kaltzeit (vor 115.000 bis 11.700 Jahren) verschwand.[1][27] Neuere Funde unter anderem aus La Ventana bei Madrid (Spanien) legen jedoch nahe, dass das Steppennashorn bis zum Ende des Spätpleistozäns (Epipaläolithikum) oder sogar bis zum Beginn des Holozäns überlebt haben könnte.[28] (siehe auch Hauptartikel: Quartäre Aussterbewelle)
Forschungsgeschichte

Die Erstbeschreibung als Rhinoceros hemitoechus erfolgte von Hugh Falconer 1868.[29] Seiner Bearbeitung lagen Funde aus der Minchin-Höhle auf der Gower-Halbinsel in Glamorgan (Wales) zugrunde. Aber bereits 1859 hatte Falconer Nashornfossilien aus „Höhlen in Glamorganshire“ (Wales) mit dem gleichen Namen vorgestellt, was auch in mehreren Publikationen jener Zeit erwähnt wird. Die Funde aus den „Höhlen in Glamorganshire“ stellen deshalb heute den Syntypus und jene aus der Minchin-Höhle den Lectotypus dieser Tierart dar.[8]
Schon vor Falconer, im Jahr 1846, hatte der englische Naturforscher Richard Owen (1804–1892) für das Steppennashorn den Namen Rhinoceros leptorhinus eingeführt, der aber heute nicht mehr gültig ist. Für seine Beschreibung hatte er Funde aus Clacton-on-Sea (England) herangezogen, die aber ebenfalls nicht mehr zu den Typusexemplaren gehören. Der Name ist außerdem ein Homonym der gleichen vom französischen Naturforscher Georges Cuvier (1769–1832) bereits 1822 benutzten Bezeichnung.
Der ursprünglich gebrauchte Gattungsname Rhinoceros wurde später durch Dicerorhinus ersetzt. Aufgrund zahlreicher morphologischer Unterschiede zum heute noch lebenden Sumatra-Nashorn als einzigen rezenten Vertreter, wurde 1942 vom ungarischen Paläontologen Miklós Kretzoi (1907–2005) der heute gültige Gattungsname Stephanorhinus eingeführt.[30] Diese Gattung umfasst die meisten der fossilen plio- und pleistozänen Nashörner des nördlichen Eurasiens und ist monophyletisch verwandt mit der Gattung Coelodonta, zu der auch das ebenfalls im Mittel- und Jungpleistozän auftretende Wollnashorn gehört.
Forschungsgeschichtlich gesehen gab es bezüglich des Steppen- und des mit ihm verwandten Waldnashorns häufig große Verwirrung. Im Lauf der Zeit wurde das Steppennashorn unter verschiedenen wissenschaftlichen Namen geführt, die sich teilweise mit denen des Waldnashorns überschneiden:[25]
- Rhinoceros leptorhinus Cuvier, 1822
- Rhinoceros megarhinus De Christol, 1834
- Dicerorhinus mercki Kaup, 1841
- Rhinoceros Mercki Kaup, 1841
- Rhinoceros leptorhinus Owen, 1846
- Rhinoceros lunellensis Gervais, 1848–1852
- Rhinoceros hemitoechus Falconer, 1868
- Rhinoceros subinermis Pomel, 1895
- Rhinoceros hemitoechus aretinus Azzaroli, 1962
- Rhinoceros hemitoechus falconeri Azzaroli, 1962.
Literatur
- Wighart von Koenigswald: Lebendige Eiszeit. Klima und Tierwelt im Wandel. Theiss-Verlag, Stuttgart 2002, ISBN 3-8062-1734-3.
Weblinks
Einzelnachweise
- H. Loose: Pleistocene Rhinocerotidae of W. Europe with reference to the recent two-horned species of Africa and S.E. Asia. Scripta Geologica 33, 1975, S. 1–59
- Denise Geraads: Rhinocerotidae. In: L. Werdelin und D. J. Sanders (Hrsg.): Cenozoic Mammals of Africa. Berkeley, 2010, S. 669–683
- Donald R. Prothero, Claude Guérin und Earl Manning: The history of Rhinocerotoidea. In Donald R. Prothero und R. M. Schoch (Hrsg.): The evolution of the Perissodactyls. New-York, 1989, S. 321–340
- Wighart von Koenigswald: Lebendige Eiszeit. Klima und Tierwelt im Wandel. Stuttgart, 2002, S. 54–61
- Diana Pushkina: The Pleistocene easternmost distribution in Eurasia of the species associated with the Eemian Palaeoloxodon antiquus assemblage. Mammal Review 37, 2007, S. 224–245
- Ralf-Dietrich Kahlke und Frédéric Lacombat: The earliest immigration of woolly rhinoceros (Coelodonta tologoijensis, Rhinocerotidae, Mammalia) into Europe and its adaptive evolution in Palaearctic cold stage mammal faunas. Quaternary Science Reviews 27, 2008, S. 1951–1961
- Ralf-Dietrich Kahlke: Die Entstehungs-, Entwicklungs- und Verbreitungsgeschichte des oberpleistozönen Mammuthus-Coelodonta-Faunenkomplexes in Eurasien (Großsäuger). Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft 546 Frankfurt am Main, 1994
- Jan van der Made: The rhinos from the Middle Pleistocene of Neumark-Nord (Saxony-Anhalt). In: Dietrich Mania u. a. (Hrsg.): Neumark-Nord: Ein interglaziales Ökosystem des mittelpaläolithischen Menschen. Veröffentlichungen des Landesmuseums für Vorgeschichte 62. Halle/Saale 2010, S. 433–527
- Dietrich Mania u. a.: Quartärforschung im Tagebau Neumark-Nord, Geiseltal (Sachsen-Anhalt) und ihre bisherigen Ergebnisse. In: Dietrich Mania u. a. (Hrsg.): Neumark-Nord – Ein interglaziales Ökosystem des mittelpaläolithischen Menschen. Veröffentlichungen des Landesmuseums für Vorgeschichte in Halle 62 Halle/Saale 2010, S. 11–70
- Walter Steiner: Der Travertin von Ehringsdorf und seine Fossilien. Lutherstadt Wittenberg 1981
- Dietrich Mania: Zur Geologie des altpaläolithischen Fundhorizontes von Bilzingsleben (Thüringen) unter der Berücksichtigung des geologischen Wirkfaktors „Mensch“. Hercynia 37, 2004, S. 143–184
- Jan van der Made: A preliminary note on the rhinos from Bilzingsleben. Praehistoria Thuringica 4, 2000, S. 41–64
- Karl-Dietrich Adam: Der Urmensch von Steinheim an de Murr und seine Umwelt. Ein Lebensbild aus der Zeit vor einer viertel Million Jahre. Jahrbuch des Römisch-Germanischen Zentralmuseums 35, 1988, S. 1–23
- Mikael Fortelius, Paul Mazza und Benedetto Sala: Stephanorhinus (Mammalia: Rhinocerotidae) of the Western European Pleistocene, with a revision of S. etruscus (Falconer, 1868). Palaeontographia Italica, 80, 1993, S. 63–155
- Frederic Lacombat: Phylogeny of the genus Stephanorhinus in the Plio-Pleistocene of Europe. Hallesches Jahrbuch für Geowissenschaften 23, 2007, S. 63–65
- Emmanuel M. E. Billia: Revision of the fossil material attributed to Stephanorhinus kirchbergensis (Jäger 1839) (Mammalia, Rhinocerotidae) preserved in the museum collections of the Russian Federation. Quaternary International 179, 2008, S. 25–37
- Mikael Fortelius (coordinator). Neogene of the Old World Database of Fossil Mammals (NOW). University of Helsinki, 2003 http://www.helsinki.fi/science/now/
- Andreas Wagner: Rekonstruktion von Körpermassen pleistozäner Rhinocerotidae in der Sammlung von Königswald. Frankfurt am Main, 2007
- Frederic Lacombat: Pleistocene Rhinoceroses in Mediterranean Europe and Massif Central (France). Courier des Forschungs-Instituts Senckenberg 256, 2006, S. 57–69
- René Grube: Pflanzliche Nahrungsreste der fossilen Elefanten und Nashörner aus dem Interglazial von Neumark-Nord. In: Jan Michal Burdukiewicz, Lutz Fiedler, Wolf-Dieter Heinrich, Antje Justus und Enrico Brühl (Hrsg.): Erkenntnisjäger. Festschrift für Dietrich Mania. Veröffentlichungen des Landesmuseums für Vorgeschichte in Halle 57. Halle/Saale 2003, S. 221–236
- Jan van der Made und René Grube: The rhinoceroses from Neumark-Nord and their nutrition. In: Harald Meller (Hrsg.): Elefantenreich – Eine Fossilwelt in Europa. Halle/Saale 2010, S. 382–394
- E. Cerdeño: Diversity and evolutionary trends of the family Rhinocerotidae Perissodactyla. Palaeo 141, 1997, S. 13–34
- Ralf-Dietrich Kahlke, Nuria García, Dimitris S. Kostopoulos, Frédéric Lacombat, Adrian M. Lister, Paul P.A. Mazza, Nikolai Spassov und, Vadim V. Titov: Western Palaearctic palaeoenvironmental conditions during the Early and early Middle Pleistocene inferred from large mammal communities, and implications for hominin dispersal in Europe. Quaternary Science Reviews, 2010, S. 1–28
- A. Azzaroli: Validità della specie Rhinoceros hemitoechus Falconer. Palaeontographia Italica 57, 1962, S. 21–34
- Carmelo Petronio und Luca Pandolfi: Stephanorhinus hemitoechus (FALCONER 1868) del Pleistocene Superiore dell'Area die Melpigano-Cursi E S. (Lecce, Italia). Geologica Romana 41, 2008, S. 1–12
- Simone Farina: Late Pleistocene-Holocene mammals from “Canale delle Acque Alte (Canale Mussolini)” (Agro Pontino, Latium). Bollettino della Società Paleontologica Italiana, 50 (1), 2011, S. 11–22
- Claude Guérin: Les Rhinocerotidae (Mammalia, Perissodactyla) du Miocène terminal au Pléistocène supérieur d’Europe occidentale comparés aux espèces actuelles: tendances évolutives et relations phylogénétiques. Géobios 15, 1982, S. 599–605
- A. Sánchez, S. Fraile, Jan van der Made, J. Morales, V. Quiralte, M. J. Salesa, I. M. Sánchez, B. Sanchiz, D. Soria, J. Jiménez, L. J. Barbadillo, C. Laplana, Z. Szyndlar: Primeros datos faunísticos del Neolítico madrileño: la cueva de la Ventana (Torrelaguna, Madrid). In: P. Arias Cabal, R. Ontañón Peredo, C. García-Moncó Piñeiro (Hrsg.) III Congreso del Neolítico en la Península Ibérica. Monografias del Instituto Internacional de Investigaciones Prehistóricas de Cantabria, 1, 2005, S. 155–165
- Hugh Falconer: On the European Pliocene and Pleistocene species of the genus Rhinoceros. In: C. Murchison (Hrsg.): Palaeontological memories and notes of the late Hugh Falconer. Volume 2. London, 1868, S. 309–403
- Miklós Kretzoi: Bemerkungen zum System der nachmiozänen Nashorn-Gattungen. Földtany Közlöny 72, 1942, S. 309–323